1
Biología Celular Eucariota
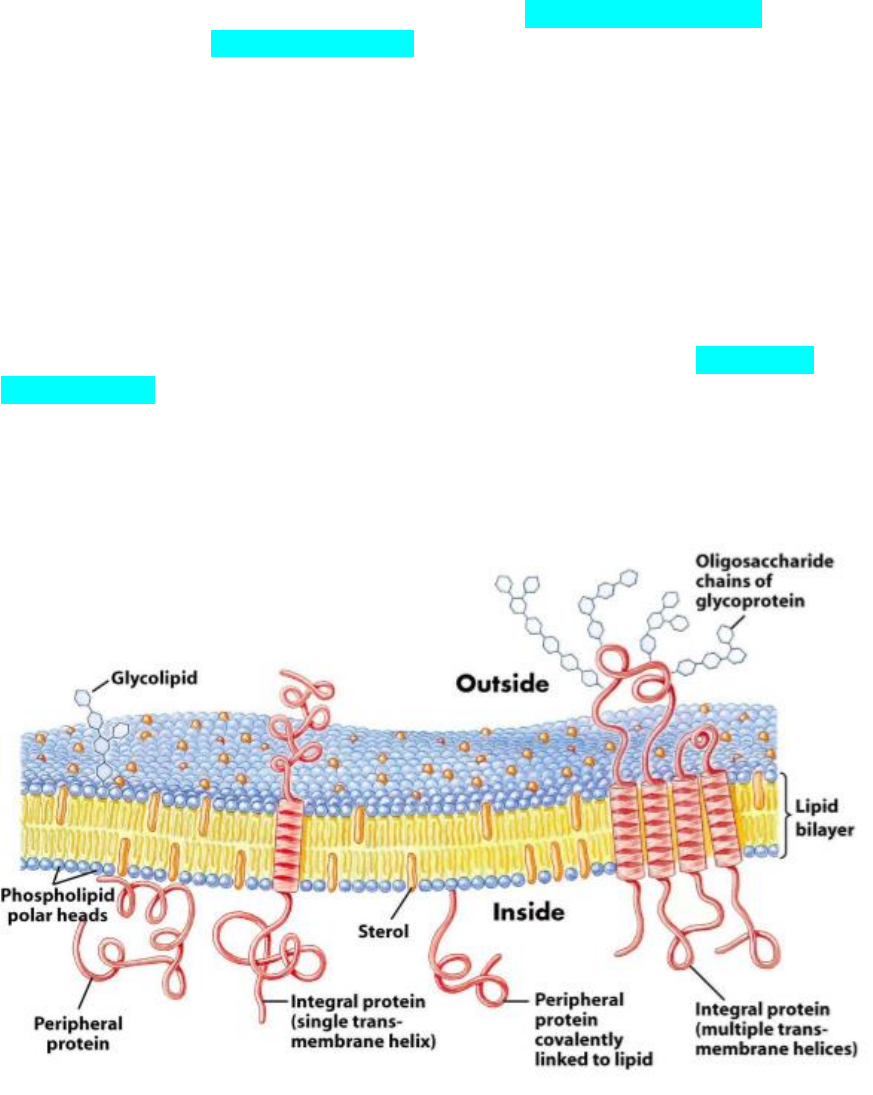
Todas las células están rodeadas por una membrana plasmática que está
constituida por una bicapa fosfolipidica con diversas proteínas intercaladas.
Los extremos de los fosfolípidos al ser polares interactúan con las partes
polares de las moléculas de agua (hidrofilicas) que están por fuera y por dentro de
las células. Por eso se orientan hacia el exterior de la bicapa.
Las largas colas hidrocarbonadas de los ácidos grasos son no polares, por
lo tanto, no pueden interactuar con los polos de las moléculas de agua
(hidrofóbicas) por eso se orientan hacia dentro de la bicapa. Los dobles enlaces
en los ácidos grasos, producen una curvatura que le otorgan mayor fluidez a la
membrana.
La estructura de la membrana celular recibe el nombre de modelo del
mosaico fluido ya que las moléculas no están en una estructura rígida, sino que
tienen movilidad.
UN MAYOR DETALLE BIOQUIMICO DE LAS MEMBRANAS BIOLOGICAS
2
Es esta imagen se puede observar con mayor detalle que la membrana
posee glucolipidicos que en conjunto constituyen el glucocalix que tiene funciones
inmunológicas y de reconocimiento entre células, entre otras. También se observa
que hay proteínas integrales que atraviesan la membrana y otras que son
periféricas y también se observa colesterol como un abundante constituyente de la
membrana. Las presencias del colesterol entre los fosfolípidos le otorgan mayor
estabilidad y rigidez a la membrana y el colesterol es abundante en la membrana
de los eucariotas, pero está ausente en las de los procariotas.
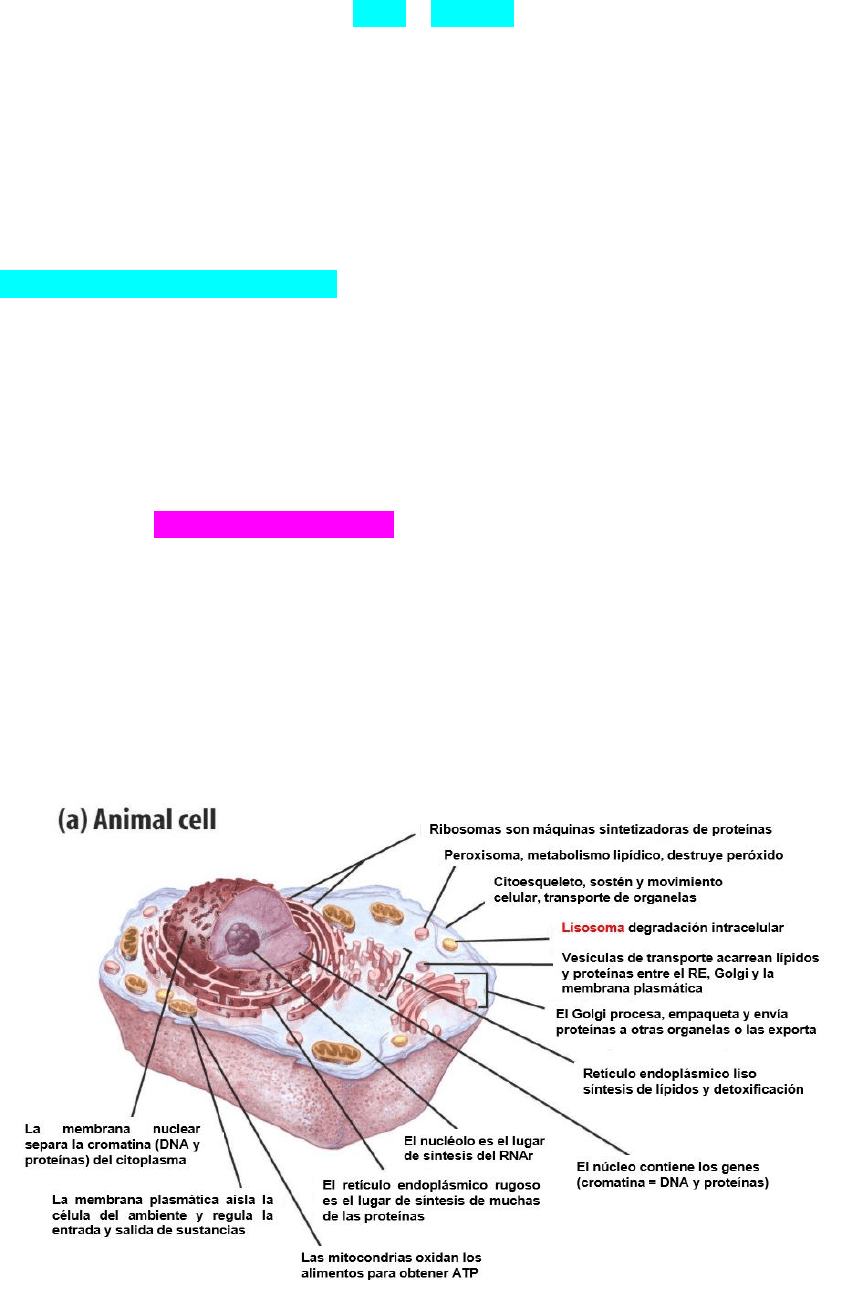
LA CELULA EUCARIOTA Y SUS ORGANULOS
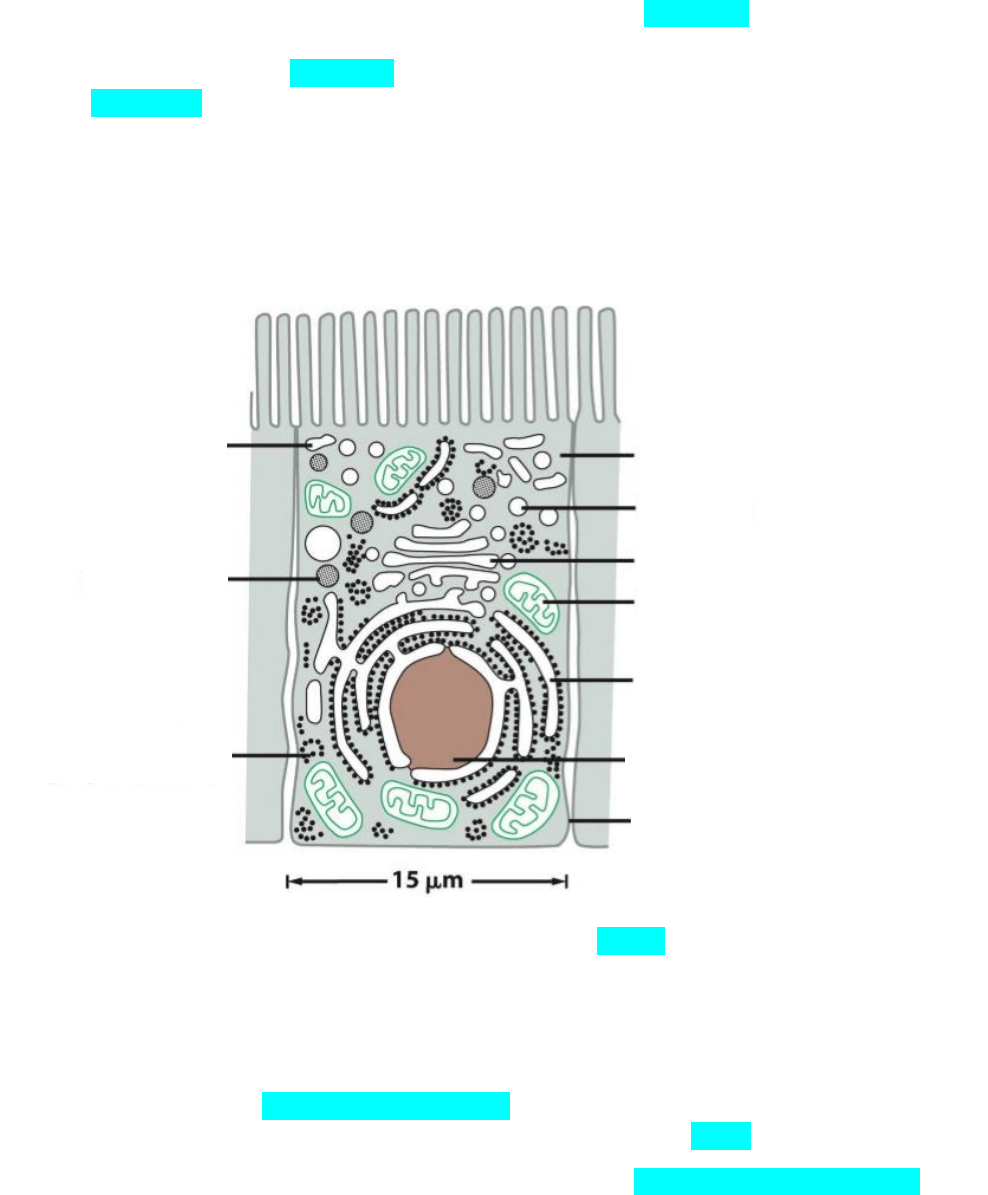
El interior de la célula este relleno por el citosol cuyo principal componente
es el agua en el que se encuentran los componentes moleculares de la célula y
donde se realiza gran parte del metabolismo. El citosol mas todos los orgánulos
que están por fuera del núcleo constituyen el citoplasma.
Todos los componentes del sistema vacuolar citoplasmático están
conectados por vesículas de transporte. Estas vesículas transportan proteínas de
membrana y diversas moléculas en su interior llamadas carga.
La formación de vesículas es producida por proteínas de recubrimiento que
se ubican en la zona de la membrana que va a formar la vesícula. Al ensamblarse
en complejos estas proteínas invaginan esta zona de la membrana que se termina
citosol
lisosoma
Aparato de Golgi
mitocondria
Retículo endoplásmico
rugoso
Nucleo
Membrane plasmatica
endosoma
peroxisoma
Polirribosomas libres
3
desprendiendo para formar la vesícula que queda cubierta por la proteína. Una de
las proteínas de recubrimiento de las vesículas es la clatrina. Otra proteína de
recubrimiento es la COPII. Nuevamente en estas vesículas proteínas de la
membrana, algunas de las cuales funcionan como receptores de la carga que se
unen a proteínas adaptadoras.
ENDOCITOSIS MEDIADA POR RECEPTOR – CATRINA DEPENDIENTE
En este tipo de endocitosis, la molécula especifica que necesita ser
endocitada es atraída y concentrada en la zona de la membrana con catrina
(donde se está formando la vesícula) por la acción de receptores y de las
proteínas adaptadoras (adaptin). Esto incrementa la cantidad de moléculas
endocitadas. Las vesículas endociticas pueden formarse gracias a la presencia de
clatrina por debajo de la zona de la membrana plasmática que va a formar dicha
vesícula.
Algunos autores identifican otro tipo de endocitosis que se produce por
pequeñas vesículas que ingieren moléculas disueltas al que denominan
pinocitosis. Es un proceso constitutivo que se produce a distintas velocidades en
casi todas las células.
Las vesículas pinociticas pueden formarse en parches de la membrana
plasmática ricas en una proteína integral de membrana llamada caveolina. Estas
vesículas se denominan caveolas.
Cuando por endocitosis se ingieren grandes partículas, como pueden ser
microorganismos patógenos, células senescentes o muertas por apoptosis, el
proceso se denomina fagocitosis. No es un proceso constitutivo, sino que es
producido cuando se lo necesita. La fagocitosis es utilizada por células del sistema
inmunitario tales como los neutrófilos para destruir bacterias. Hay receptores de
membrana que reconocen a los anticuerpos que se unieron al microorganismo
desencadenando la formación del fagosoma. Los macrófagos utilizan la fagocitosis
para ingerir material no biológico, células que han muerto por apoptosis o células
senescentes. Los fagosomas se unen a los lisosomas para degradar las partículas
fagocitadas. A los macrófagos y neutrófilos se los denomina fagocitos
profesionales.
El compartimiento al cual una vesícula se fusiona es especifico y debe de
estar regulado. Primero las proteínas Rab dirigen la vesícula al compartimiento
adecuado y luego las proteínas SNARE inducen la fusión de ambas membranas.
Las Rab son GTPasas que cuando están unidas a GDP están en el citosol y
cuando se une a GTP despliegan una hélice hidrofilica con la que se unen a sitio
específicos de las membranas. Estas proteínas son reconocidas por proteínas de
amarre que están en la membrana target. Una vez que las dos membranas están
4
cerca las proteínas vSNARE que están sobre la vesícula se unen a las tSNARE
que están sobre la membrana del compartimiento target, desplazando el agua que
queda entre las dos membranas e induciendo la fusión de las mismas con lo que
la vesícula se conecta con el compartimiento target y puede volcar su carga en él.
DESDE LA MEMBRANA PLASMATICA HASTA LOS LISOSOMAS
Las vesículas endociticas se conectan con los endosomas tempranos.
Estos clasifican las proteínas entregadas por las vesículas endociticas y las
reciclan nuevamente a la membrana para que sean reutilizadas (receptor del LDL)
o las incorporan en vesículas en su interior formando los cuerpos multivesiculares
en cuyo caso pasaran a los endosomas tardíos que se fusionaran o maduraran a
lisosomas donde serán degradados. Las proteínas contenidas en estas vesículas
internas terminarán dentro de los lisosomas y por lo tanto serán degradadas.
Los endosomas tempranos reciben la vesícula endociticas (con su carga) y
las vesículas que se forman a partir de ellos pueden tener tres destinos diferentes:
1- Reciclado con lo que algunas proteínas endocitadas vuelven a la
membrana plasmática (ej. Receptor de LDL).
2- Trancitosis, las vesículas son exocitadas en regiones de la membrana
plasmática distinta a la región donde fueron endocitadas (ej. Secreción
de inmunoglobulinas A en el aparato digestivo)
3- Degradación, las vesículas se unen o maduran a endosomas tardíos
para terminar en el lisosoma donde su carga es degradada (el factor de
crecimiento epidérmico EGF y su receptor).
Los lisosomas son el principal sitio de digestión intracelular de elementos
que se han vuelto obsoletos para la célula o el organismo. Los lisosomas
contienen un conjunto de enzimas llamadas hidrolasas acidas que degradan
prácticamente todos los componentes macromoleculares de la celula a sus
monómeros constituyentes. Estas enzimas se producen en el RER y son
trasladadas al Golgi que las empaqueta en vesículas y las envía a los lisosomas.
Estas enzimas trabajan solamente a pH acido. Los lisosomas son típicos de las
células animal. En las células vegetales sus funciones son llevadas a cabo por la
vacuola central, aunque estas últimas cumple otras funciones que no son
realizadas por los lisosomas. Los precursores inactivos de las hidrolasas
lisosomas pasan del RER al Golgi donde son marcados con un olisacarido que
tienen una manosa en su extremo, este azúcar es fosforilado para dar M6P. El
grupo fosfato es agregado con otra molécula que es inmediatamente eliminada.
En los últimos receptores se desprenden de la hidrolasa y es reciclado enviándolo
nuevamente al Golgi. Las hidrolasas quedan dentro del endosoma y a medida que
este madure a lisosoma, el pH descenderá, con lo que estas enzimas se activaran.
5
Los lisosomas reciben los materiales que degradan de vesículas que provienen de
tres fuentes distintas: la endocitosis, la fagocitosis y la autofagia.
Los proteosomas constituyen una vía alternativa de degradación de
proteínas. Existe una forma alternativa a la autofagia por medio de la cual las
proteínas pueden ser degradadas. Esta es llevada a cabo por los proteosomas.
Las proteínas que deban ser degradadas por este medio son marcadas como una
cadena de una proteína pequeña llamada ubiquitina. Estas ubiquitinas son
agregadas a la proteína target por enzimas denominadas ubiquitina ligasas. Las
proteínas ubiquitinadas son enviadas al proteosoma que es una estructura
cilíndrica hueca cuya pared está formada por proteasas. En el interior del
proteosoma la proteína ubiquitinada es degradada a aminoácidos o a pequeños
péptidos que son expulsados por el otro extremo del proteosoma.
El retículo endoplásmico es un conjunto de tubos y sacos aplanados
formados por membrana cuyos espacios interiores se comunican entre si y que
por fuera se continúan con la membrana plasmática y por dentro con la membrana
externa de la envoltura nuclear. El RE se divide estructural y funcional en RE
rugoso y RE liso.
El RER se caracteriza por poseer ribosomas adheridos sobre su cara
citosolica. Es el lugar donde se sintetizan todas las proteínas de las membranas
celulares y las proteínas que van a ser secretadas (exportadas al exterior celular).
El REL no posee ribosomas y es el lugar de síntesis de los lípidos
(incluyendo los que forman parte de las membranas celulares) y los esteroides.
También está involucrado en la detoxificacion gracias a las enzimas de la familia
del citocromo P450 que hacen que los compuestos toxico hidrofóbicos se vuelvan
hidrosolubles y puedan ser eliminados del cuerpo.
Las proteínas sintetizadas en el RER, ya sea las solubles o incorporadas a
la membrana, son trasladadas al Golgi por medio de vesículas cubiertas por
COPII. La membrana que va a formar estas vesículas tiene receptores que van a
unir las proteínas solubles que deben ser transportadas. Estas vesículas pierden
la cubierta luego de formarse, se fusionan entre sí (vesicular tubular clúster) y son
transportadas por proteínas motoras a lo largo de micro túbulos que las llevan
hasta la cara cis del Golgi donde se fusionan para entregar la proteína que
acarrean a este último compartimento. Las proteínas receptoras pueden retornar
desde el Golgi a RER para ser reutilizadas por medio de vesículas cubierta por
COPI.
El aparato de Golgi consiste de una serie ordenada de compartimientos.
Está formado por una serie de cisternas aplanadas dispuestas una sobre otra. La
6
cara cis es la más cercana al RE. Las vesículas cargadas provenientes del RE
ingresan al Golgi por su cara cis, luego van pasando por las cisternas medias y
finalmente salen en vesículas por la cara trans. Durante este pasaje los lípidos y
las proteínas son glicosilados. Finalmente, el Golgi decidirá el destino de las
vesículas. Si son enviada a la membrana plasmática para que secreten su
contenido al exterior, si llevan su contenido a los lisosomas, a los endosomas o a
la superficie celular.
El aparato de Golgi dirige el trafico vesicular del sistema vacuolar.
Exocitosis. Diagrama del camino seguido por las proteínas sintetizadas en
el RER hasta ser secretadas al exterior de la célula o integradas a la membrana
plasmática. Este camino se denomina VIA BIOSINTETICA SECRETO. La
exocitosis puede ser constitutiva o regulada. En este último caso un estímulo
(ligand) produce la apertura de canales de calcio. El ingreso de este último a la
célula estimula la fusión de la vesícula exocíticas a la membrana plasmática. La
proteína de cubierta COPII media el transporte desde el RER al Golgi. La COPI
media el transporte inverso
Los peroxisomas están presentes tanto en las células animales como en las
vegetales. Estas vesículas contienen enzimas que oxidan varios compuestos
produciendo peróxido de hidrogeno. Estas enzimas se encuentran en tan alta
concentración que forma los típicos cristales observados en estos orgánulos. El
H
2
O
2
es convertido en H
2
O y O
2
por encimas llamadas catalasas.
Las células vegetales también poseen peroxisomas donde pueden convertir
ácidos grasos en azucares gracias a un ciclo bioquímico llamado ciclo del
glioxilato. Este proceso es importante en las semillas donde la energía se
almacena en forma de triglicéridos. Por eso los peroxisomas vegetales suelen
recibir el nombre de glioxisomas.
Los ribosomas están compuestos por proteínas y ARN ribosómico (ARNr).
Es el orgánulo celular donde se realiza la síntesis de las proteínas (traducción).
Tanto las células procariotas como las eucariotas poseen ribosomas
formados por una subunidad mayor y otra menor. Sin embargo, las subunidades y
también la composición de los ribosomas procariotas son distintas de las de los
eucariotas. Las diferencias entre la estructura de los ribosomas procarióticos
(bacterianos) y los ribosomas eucarióticos fueron aprovechadas por los
investigadores, quienes descubrieron compuestos químicos (antibióticos) que se
fijan a los ribosomas bacterianos (pero no a los ribosomas eucariotas) y, de ese
modo, destruyen una infección bacteriana sin herir a las células de la persona
infectada.
7
Los ribosomas están compuestos por proteínas y ARNr. Es el orgánulo
celular donde se realiza la síntesis de proteínas (traducción) el ribosoma es una
ribozima que cataliza el enlace peptídico.
Cada ARNt se une y acarrea uno solo de los 20 aminoácidos. Durante la
síntesis de proteínas 3 ARNt se ubican dentro del ribosoma. El ARNt tiene tres
nucleótidos denominados anti codón que se aparean con cada 3 nucleótidos del
ARNm llamado codón de acuerdo al típico apareamiento de bases (A-U y C-G).
De manera que la secuencia de nucleótidos del ARN mensajero, que a su vez es
copiada de la secuencia del ADN, determina el orden de aminoácidos en la
proteína (estructura primaria).
Los ribosomas se pueden encontrar en el citosol donde forman
poliribosomas y sintetizan las proteínas que permanecerán en el citosol. Como ya
vimos, también se encuentran adosados a las cisternas del RE donde forman el
RER y sintetizan las proteínas que se incorporarán a las membranas o serán
secretadas al exterior de la célula.
El citoplasma de las células eucariotas contiene una red de filamentos
denominada CITOESQUELETO que le da sostén a las células, se encarga de la
motilidad celular y del tráfico intracelular de organelas. Estas variadas funciones
dependen del comportamiento de tres familias de proteínas que se ensamblan
para formar tres tipos de filamentos.
1. Los MICROFILAMENTOS o FILAMENTOS DE ACTINA están formados
por la polimerización de un monómero de la proteína ACTINA.
2. Los MICROTÚBULOS están formados por la polimerización de
heterodímeros de α y β tubulina. Son filamentos huecos más rígidos que
los microfilamentos de actina.
Los microtubulos actúan como vías para para el trafico intracelular de
vesículas y organelas con la ayuda de proteínas motoras (kinesina y dineina) que
utilizan ATP.
Los microtubulos se originan en estructuras conocidas como centros
organizadores de microtubulos (MTOC). La mayoría de las células tienen un
MTOC bien definido cerca del núcleo llamado Centrosoma.
El centrosoma está compuesto de una matriz del centrosoma rica en γ
tubulina que forma los anillos a partir de los cuales se polimeriza la α y β tubulina
para formar los microtúbulos. Dentro se encuentran los centriolos formados por
dos cilindros dispuesto en ángulo recto, cada uno de los cuales está formado por 9
tripletes de microtúbulos. Estas estructuras no parecen ser necesarias para la
formación de los microtúbulos ya que están ausentes en MTOCs de algunas
células animales, de muchas células vegetales (angiospermas y gimnospermas) y
de las células de los hongos.
8
La célula eucariota posee cilios y flagelos con una estructura
completamente diferente a la de los flagelos de la célula procariota.
Los cilios y flagelos son proyecciones de la célula eucariota que le sirven
para moverse o para crear corriente y mover otras estructuras. Ambos tienen la
misma estructura interna pero los cilios son más cortos y aparecen en grandes
cantidades en cambio los flagelos se encuentran solos y son más largos.
la estructura de cilios y flagelos conformada por nueve pares de
microtúbulos periféricos y un par central. Cilios y flagelos se forman a partir de
cuerpos basales o cinetosomas que están por debajo de la membrana plasmática.
Estos últimos tienen la misma estructura de los centriolos de 9 tripletes de
microtúbulos periféricos (ver abajo) y son intercambiables con los centriolos que
se encuentran en el centrosoma.
Los centrosomas y los microtubulos forman parte fundamentalmente del
huso mitótico que se encarga de producir la migración de los cromosomas hacia
los polos de la célula en división durante la mitosis y la meiosis.
3. Los filamentos intermedios están formados por una familia de proteínas
evolutivamente relacionadas. Estas proteinas, que están conformadas
por una alfa-helice central y dos extremos globulares, son los
monómeros que al polimerizar forman el filamento. Gracias a sus
estructuras los filamentos intermedios son muy resistentes al
estiramiento y otorgan resistencia mecánica a las celulas.
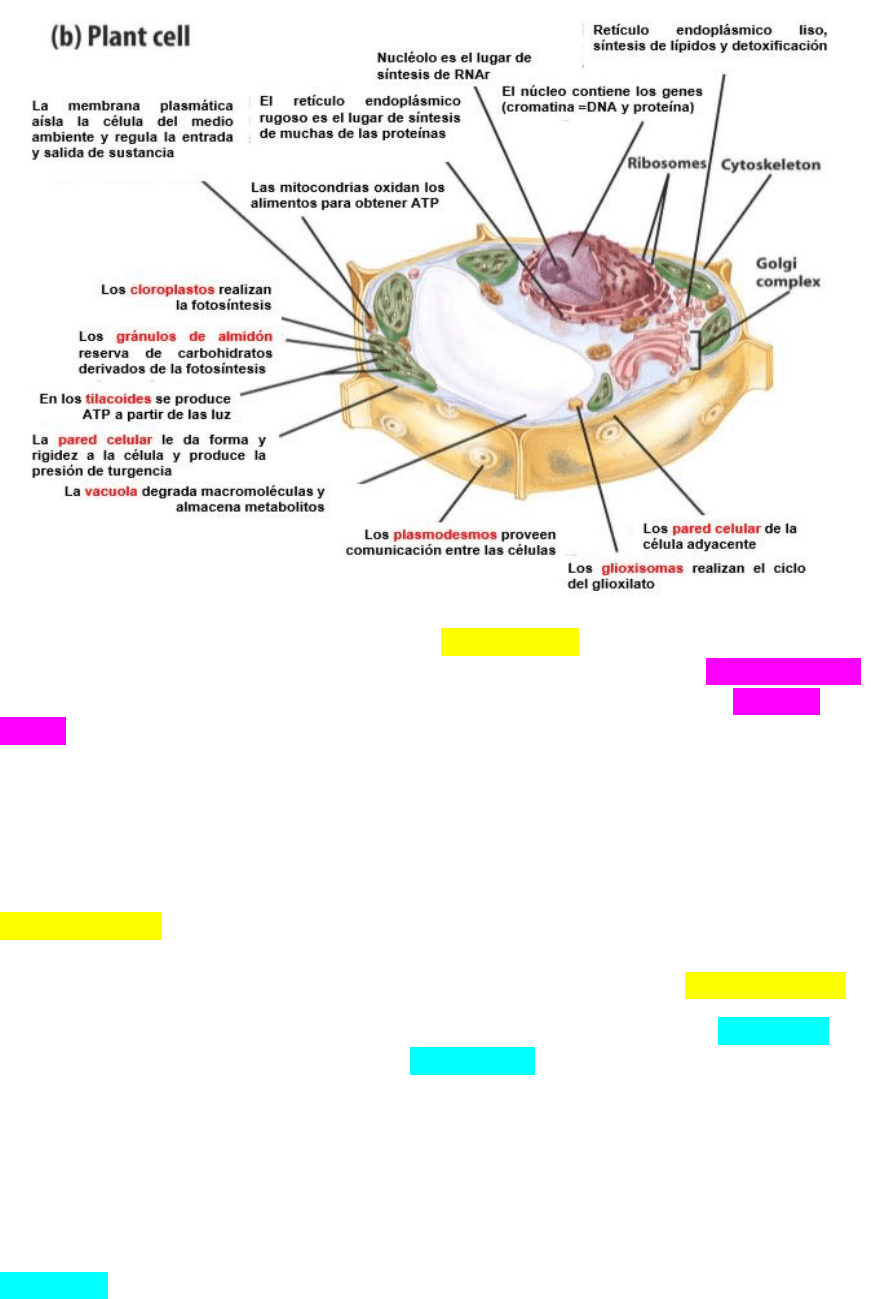
DIFERENCIAS ENTRE CELULAS ANIMALES Y VEGETALES
9
Las células vegetales poseen una pared celular por fuera de la membrana
plasmática. Por fuera de la membrana plasmática se encuentra la pared primaria
que se encuentra separada de la pared de las células vecinas por la laminilla
media. Muchas células vegetales al madurar depositan una pared secundaria por
dentro (pegada a la membrana plasmática) de la primaria. Esta última contiene
lignina que le otorga dureza y la hace impermeable al agua. La pared celular está
compuesta de microfibrillas de celulosa unidas entre sí por otro polisacárido
llamado glicano embebidos en otro azúcar llamado pectina.
Las paredes celulares están atravesadas por orificios llamados
plasmodesmos a través de los cuales se continua la membrana plasmática y el
citoplasma de las dos células vecinas, frecuentemente el plasmodesmo contiene
un túbulo que conecta el RE de las dos células vecinas llamado desmostubulos.
La pared celular se forma durante la división citoplasmática (citocinesis) que
ocurre luego de la división del núcleo (cariocinesis) en la mitosis. Varias vesículas
provenientes del Golgi cargadas con celulosa (y con los demás componentes de la
pared celular) son llevadas hacia el plano ecuatorial de la célula en división por los
microtúbulos. Una vez allí se fusionan entre sí de manera que sus membranas
pasan a formar parte de la nueva membrana plasmática y el contenido que
transportan pasa a constituir la nueva pared celular.
La célula vegetal posee una familia de orgánulos específicos llamados
plásmidos. Todos los plásticos derivan de un proplástido que se va diferenciando a
10
medida que la célula vegetal se especializa en una función determinada. Todos los
plástidos tiene doble membrana lo cual se considera una evidencia de su origen
endosimbionte. Cuando su ancestro procariota ingresó a la célula antecesora
eucariota conservó su propia membrana más la derivada de la invaginación de la
membrana plasmática de la célula eucariota progenitora. Los plastidos que
almacenan almidón reciben el nombre de leucoplastos o amiloplastos.
Los cloroplastos llevan a cabo la fotosíntesis. El cloroplasto posee tres
juegos de membrana. Las dos membranas externas más las membranas internas
que son un conjunto de cisternas denominadas tilacoides que en ciertos lugares
se disponen en forma apilada. El conjunto de tilacoides apilados recibe el nombre
de granas.
Se denomina metabolismo celular al conjunto de reacciones químicas que
ocurren en la célula. Una célula eucariota típica puede sintetizar 30.000 enzimas
(proteínas) que catalizan miles de reacciones químicas diferentes que involucran a
cientos de metabolitos, muchos de ellos compartidos por varias de las vías
metabólicas. Todas las reacciones químicas de las células están catalizadas por
enzimas (o ribozimas). Las reacciones y vías metabólicas pueden dividirse en
catabólicas y anabólicas. En las catabólicas las moléculas complejas (polímeros)
son degradadas a moléculas simples (monómeros) más pequeñas con liberación
de energía y en las anabólicas se sintetizan grandes moléculas complejas a partir
de moléculas pequeñas con utilización de energía.
El principal proceso catabólico de la célula eucariota es la respiración
celular. La respiración celular consta de 3 partes:
1- Glucolisis.
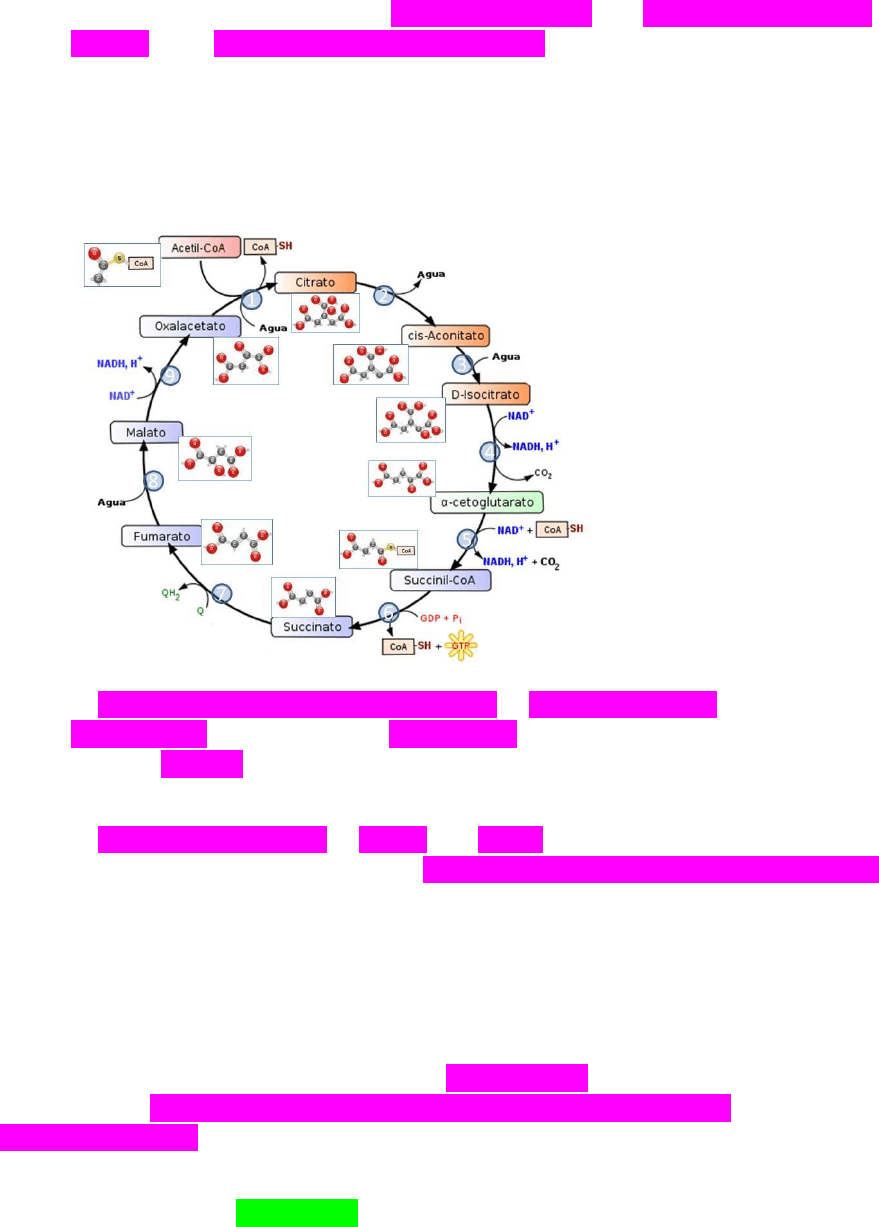
2- Ciclo de Krebs o del ácido cítrico.
3- Fosforilacion oxidativa.
La glucolisis: es la ruta metabólica encargada de oxidar la glucosa con la
finalidad de obtener energía para la célula. Consiste en 10 reacciones enzimáticas
consecutivas que convierten a la glucosa en dos moléculas de piruvato, el cual es
capaz de seguir otras vías metabólicas y así continuar entregando energía al
organismo. Esta ruta se realiza tanto en ausencia como presencia de oxígeno,
definido como proceso anaeróbico en este caso. El ácido pirúvico es las
condiciones anaeróbicas de ejercicio extremo puede ser convertido en ácido
láctico. Las levaduras también en anaerobiosis, lo convertirán en etanol. Estos
procesos reciben el nombre de fermentación. En la mayoría de las células en
presencia de O
2
será transferido al interior de la mitocondria donde será
descarboxilado (perdida de CO
2
) para dar acetil coenzima A que ingresara al ciclo
de Krebs.
11
El ciclo de Krebs ocurre en la matriz mitocondrial y la fosforilacion oxidativa
en las crestas de la membrana mitocondrial interna. La mitocondria consta de una
membrana externa y una interna que delimitan un espacio intermembranosa. La
membrana interna se pliega hacia el interior dando crestas. Por dentro de la
membrana interna se encuentra la matriz mitocondrial.
En el ciclo de Krebs o ciclo del ácido cítrico el acetil coenzima A (2C) se une
con el oxaloacetato (4C) para dar el ácido cítrico (6C) que será oxidado al perder
dos moléculas de CO2 para regenerar el oxaloacetato y formar 3 NADH, un FADH
y un GTP (este último cede su grupo P a un ADP para dar un ATP).
En la fosforilación oxidativa el NADH y el FADH generados en la glucolisis y en
el ciclo de Krebs entregan sus e- a una cadena de proteínas transportadoras de e-
que están en la membrana mitocondrial interna. Como algunas de estas proteínas
puede transportar los e- libres y otras lo tienen que transportar asociado a un H+
se va generando una alta concentración de protones en el espacio
intermembranoso que produce una presión osmótica y una diferencia de cargas
eléctricas (muchas más cargas + afuera que adentro). Esta diferencia de
concentración y de cargas genera una gran cantidad de energía potencial que es
utilizada por una bomba proteica llamada ATP sintetiza (6) para unir P al ADP y
generar ATP. Este mecanismo se conoce con el nombre de TEORIA
QUIMIOSMOTICA.
Las plantas utilizan la energía de la luz solar para sintetizar azucares. Este
proceso se denomina fotosíntesis y ocurre en los cloroplastos. En la etapa
lumínica (fosforilación fotosintética) que ocurre en las membranas de los tilacoides
la clorofila al ser excitada por los fotones de la luz cede e- a una cadena de
12
transportadores que generan ATP y NADPH. El H20 es utilizada para reponer los
e- de la clorofila generando O2 libre en el proceso. La energía almacenada en el
ATP y NADPH es utilizada en el ciclo de Calvin que se realiza en el estroma del
cloroplasto para reducir el CO2 proveniente de la atmosfera y sintetizando
azucares. En esta última instancia toda la energía necesaria para la vida proviene
de la luz solar.
REACCIONES DE LA CELULA CON EL MEDIO Y TRANSPORTE DE
SUSTANCIA A TRAVES DE LAS MEMBRANAS:
Difusion es el fenómeno por el cual las moléculas tienden a ir de una zona o
región de altas concentraciones hacia otra de menor concentración. Se dice que
las sustancias se mueven a favor de un gradiente de concentración. Es una
manifestación de la capacidad que tiene las moléculas de moverse. Se dice que
una membrana es semipermeable si permite el paso del H
2
O pero no de los
solutos. La difusión del H
2
O a través de una membrana semipermeable (como la
membrana plasmática) siguiendo su gradiente de concentración se denomina
OSMOSIS. La concentración del H2O en una solución dependerá de la
concentración de soluto. Cuanta más concentración de soluto menos
concentración de H2O y viceversa. En la figura de la izquierda arriba el recipiente
contiene H2O pura y el interior del tubo H2O con soluto disuelto por lo tanto la
concentración de H2O es mayor en el recipiente que en el interior del tubo y
difundirá a través de la membrana siguiendo este gradiente e ingresando al tubo
como se observa en. La presión necesaria para evitar que el agua ingrese al
compartimento se denomina presión osmótica.
Las membranas celulares, incluyendo la membrana plasmática, debido a su
parte central hidrofóbica, solo permiten el paso de moléculas no polares y
pequeñas. Cuanto más grande y más polar es una molécula más difícil es que
pueda atravesar las membranas por difusión. Es decir que las membranas
biológicas funcionan como membranas semipermeables.
En el TRANSPORTE PASIVO las sustancias atraviesan las membranas a favor
de un gradiente de concentración por lo tanto este proceso no requiere energía.
El H
2
O atraviesa las membranas biológicas principalmente por osmosis que
también es una forma de transporte pasivo.
Si una célula se encuentra en una solución isotónica (hay igual concentración
de soluto dentro de la célula que en la solución que la rodea) el H2O difundirá
hacia fuera y hacia adentro a igual velocidad y el volumen de la célula no
cambiará.
Si una célula se encuentra en una solución hipertónica (mayor concentración
de soluto en el medio que en el interior de la célula) el H2O estará más
13
concentrada dentro de la célula y difundirá hacia el medio que la rodea y el
volumen celular disminuirá.
Si una célula se encuentra en una solución hipotónica (con menor
concentración de soluto) el H2O estará más concentrada en el medio que rodea a
la célula y difundirá hacia el interior de la misma aumentando el volumen celular.
Si la presión osmótica es grande la célula puede llegar a reventar.
Muchas moléculas atraviesan las membranas a través de otro tipo de
transporte pasivo denominado DIFUSION FACILITADA en la cual dos tipos de
proteínas de membrana denominados transportadores y canales facilitan el pasaje
de estas moléculas a través de las membranas.
Un transportador se une específicamente a la molécula que debe transportar y
cambia su conformación para exponer el sitio de unión alternativamente a ambos
lados de la membrana. Un ejemplo es el transportador de glucosa.
Un canal no se une a la molécula que debe transportar y forma un poro
continuo a través de la membrana que ofrece un paso hidrofílico a través del cual
las moléculas polares pueden atravesar las membranas. En algunos canales este
poro puede cerrarse o abrirse en forma controlada.
Cuando se necesita transportar una molécula a través de una membrana en
contra de su gradiente de concentración o eléctrico el proceso se denomina
TRANSPORTE ACTIVO y requiere energía. Las proteínas de membrana
encargadas de este proceso se denominan bombas.
Muchas macromoléculas, moléculas y grandes partículas pueden entrar o salir
de la célula por medio del TRANSPORTE MEDIADO POR VESÍCULAS o
TRANSPORTE VESICULAR que también se utiliza para transportar moléculas
entre los diferentes compartimentos celulares y renovar la membrana plasmática.
ENDOCITOSIS es el mecanismo por el cual la célula capta partículas del
medio externo mediante la invaginación de la membrana plasmática que
posteriormente se estrangula para formar la vesícula que transporta su contenido
hacia el interior de la célula.
EXOCITOSIS es el mecanismo por el cual las macromoléculas contenidas en
vesículas secretoras generadas en el aparato de Golgi son transportadas hasta la
membrana plasmática, donde ambas membranas se fusionan para liberar su
contenido al exterior de la célula. De esta manera se secretan hormonas, enzimas
digestivas, neurotransmisores, etc.).
EL NUCLEO CELULAR
14
El núcleo celular está delimitado por la membrana nuclear que está constituida
por una doble membrana (cuatro capas fosfolipídicas) que se continua
directamente con el RE y puede tener ribosomas adheridos a su superficie
citoplasmática. La membrana nuclear posee poros nucleares que comunican el
nucleoplasma con el citoplasma. En el interior se observa la presencia de
cromatina que se presenta en dos formas: heterocromatina o eucromatina. Es
conspicua la presencia de un nucléolo. La cara interior de la membrana nuclear
esta tapizada por la lámina nuclear.
La LAMINA NUCLEAR tapiza internamente la membrana nuclear. Está
formada por una red bidimensional de subunidades proteicas llamadas láminas
similares a las proteínas de los filamentos intermedios. Esta red está adherida a
las proteínas de la membrana nuclear interna, otorgándole estabilidad, y a la
cromatina. Durante la mitosis las láminas nucleares son fosforiladas, lo que
conduce al desarmado de la red, lo que a su vez libera la membrana nuclear que
entonces se incorpora a las cisternas de RE. Este es el mecanismo por el cual el
núcleo desaparece durante la mitosis.
En muchos lugares la membrana nuclear externa se continua con la interna
delimitando un orificio que comunican el nucleoplasma con el citoplasma y que se
denomina PORO NUCLEAR. El orificio contiene un conjunto de proteínas que
conforman el COMPLEJO DEL PORO NUCLEAR cuya función es regular el
pasaje de moléculas y macromoléculas entre el núcleo y el citoplasma.
El complejo del poro consta de 8 unidades idénticas que se repitan alrededor
del mismo. Cada unidad está formada por una subunidad columnar que se
conecta con dos subunidades de los anillos que forma dos anillos del lado
citosólico y nuclear del poro. La subunidad luminal se proyecta al espacio
intermembranoso de la membrana nuclear y ancla a la subunidad columnar a la
membrana. La subunidad columnar también conecta con una subunidad anular
hacia el interior del poro. De cada subunidad de los anillos surge una proteína
fibrilar. El conjunto de fibrillas del anillo nuclear se une para dar una estructura con
forma de cesto de básquet.
EL TRANSPORTE A TRAVES DEL PORO NUCLEAR
Las proteínas que tiene que entrar o salir del núcleo tienen en unos de sus
extremos varios aminoácidos que constituyen señales de localización nuclear y
señales de exportación nuclear. Estas señales son reconocidas específicamente
por receptores de importación nuclear y receptores de exportación nuclear que se
unen a las proteínas del complejo del poro permitiendo el pasaje de la proteína
que transportan (cargo). La molécula RAN-GTP al cambiar la conformación de los
receptores para que puedan descargar o cargar las proteínas que tienen que
transportar, aporta la energía necesaria para el proceso de transporte.
15
El nucléolo es una fábrica de ribosomas y de otros complejos
ribonucleoproteicos.
Los genes que codifican al rARN se denominan organizadores nucleolares. De
la transcripción de estos genes surge un rARN precursor de 45S que por un
proceso de corte y empalme conocido como splicing dará origen los rARN
maduros que formaran parte del ribosoma (excepto el rARN de 5S que no es
producido en el nucléolo). Las proteínas que componen el ribosoma y las
proteínas enzimáticas que catalizan las reacciones necesarias para la formación
de los ribosomas se producen en el citoplasma y son importadas al núcleo a
través de los poros nucleares para que puedan llegar al nucléolo. Una vez que las
subunidades menor y mayor están formadas son exportada al citoplasma donde
pueden unirse para formar el ribosoma y realizar la síntesis de proteínas. Otros
complejos ribonucleo-proteicos como la enzima telomerasa que alarga los
extremos de los cromosomas (telomeros) también se sintetizan en el nucléolo.
En el nucléolo se pueden distinguir centros fibrilares rodeados de un
componente fibrilar denso e inmersos en el componente granular. La transcripción
de los genes para el rARN se produce en el centro fibrilar. Los transcriptos de
rARN se acumulan en el componente fibrilar denso y el ensamblaje de las
subunidades ribosómicas ocurre en el componente granular.
En las células eucariotas el ADN se encuentra enrollado alrededor de un centro
de proteínas llamadas histonas conformando una estructura denominada
nucleosomas.
Los nucleosomas están conectados entre sí por los sectores de la molécula de
ADN que no están enrollados alrededor del complejo de histonas.
En el núcleo interfasico (entre la mitosis) el ADN y las histonas forman la
cromatina. Se distinguen 2 tipos de cromatina: eucromatina y heterocromatina.
La transcripción solo ocurre en el estado de eucromatina. El enrollamiento de
eucromatina a heterocromatina y viceversa está estrictamente regulado por
mecanismos moleculares especiales y es uno de las formas de control de la
expresión de los genes.
Durante la división celular (mitosis y meiosis) la cromatina se condensa para
formar los cromosomas.
la replicación (duplicación del ADN) ocurre en la interfase dando origen a las
CROMATIDES que quedarán unidas hasta que sean separadas en la mitosis
subsiguiente. Cada cromátide posee una de las dos moléculas de ADN hijas que
resultaron de la replicación.
La interface es el periodo de la vida de la célula que va entre dos mitosis. El
cromosoma durante la interface está en estado de cromatina (eucromatina o
Este documento contiene más páginas...
Descargar Completo
Unidad 3. Biologia Celular Eucariota.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.