pág. 1 Agustina Colombo
Dra Said. Transporte celular
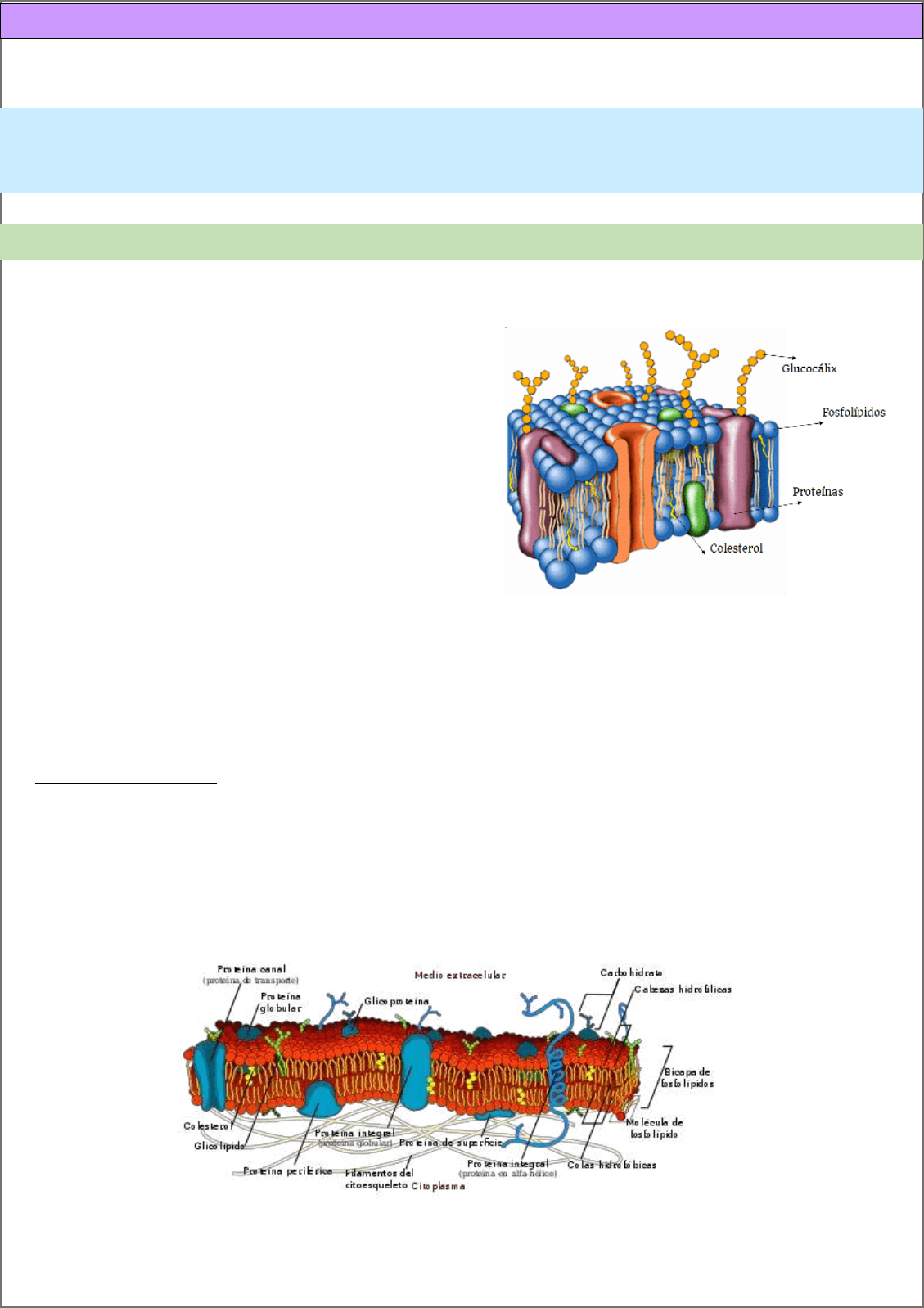
La membrana plasmática es una bicapa lipídica (fosfolípidos principalmente) en los cuales hay proteínas embebidas.
Otros componentes en menor cantidad son carbohidratos.
El modelo de mosaico fluido, es el modelo de membrana
existente, que está constituido por moléculas antipáticas
intercaladas entre proteínas. En donde los lípidos pueden
moverse de lugar y rotar sobre su propio eje.
Dependiendo el tipo de ácido graso presentes en los
fosfolípidos y de la cantidad de colesterol presentan mayor o
menor fluidez de membrana.
¿para qué sirve la membrana?
Aislamiento físico: separa el líquido intracelular del
extracelular
Permeabilidad selectiva: debido a su interior
hidrofóbico, la bicapa lipídica es una barrera altamente impermeable para la mayoría de las moléculas
polares
Transporte: el intercambio de materia entre el interior de la célula y su ambiente externo.
Soporte estructural: mantenimiento de la forma celular
Reconocimiento y comunicación: gracias a moléculas situadas en la parte externa de las membranas, que
actúan como receptoras.
Proteínas de membrana: se pueden clasificar en:
Integrales: embebidas en la membrana, tienen conexión con el espacio intra/extra celular. No se pueden
extraer con tratamientos con sal, para lograrlo debemos disolver la membrana con detergentes. Encontramos:
o Proteínas transmembranas: atraviesan la bicapa lipídica una o más veces.
o Proteínas que integran la membrana, pero no la atraviesan por completo.
o Proteínas ancladas a lípidos por enlaces covalentes. Ej: Glucosilfosfatidilinositol (GPI).
Periféricas: se adosan a otras proteínas integrales o a lípidos, pero por uniones NO covalentes. Con
tratamientos suaves se pueden extraer.
Funciones de proteína integrales de membranas:
Transporte celular a través de membrana.
Conocimientos básicos membrana
Músculo esquelético y unión neuromuscular. Electrofisiología general. (TP1)
pág. 2 Agustina Colombo
o Receptores: receptores de unión a ligando (transductor de señal). Receptores acoplados a proteínas G.
fármacos pueden interactuar inhibiendo desde este receptor.
o Moléculas de adhesión: forma contacto físico con la matriz extracelular circundante o con sus vecinos
celulares. Ej: integrinas – cadherinas.
o Movimiento transmembrana de sustancias hidrosolubles: canales y poros, conductos que permiten el paso de
agua, iones, proteínas. Transportadores y bombas.
o Enzimas: las bombas iónicas son enzimas que catalizan la hidrólisis del ATP.
o Señalización intracelular: algunas se asocian con ácidos grasos o grupos fenilo.
Función de proteína periférica: participan en la señalización intracelular y pueden formar un citoesqueleto
submembranoso.
La membrana diferencia el interior del exterior, genera compartimiento cerrado. La concentración de iones es muy
diferente en ambos compartimientos:
Involucra el paso o movimientos de iones y moléculas a través de la membrana celular, independientemente de su
tamaño y del mecanismo mediante el cual la atraviesan.
Las moléculas polares y cargadas: difícil que atraviesen la membrana (el núcleo hidrofóbico)
Las moléculas de agua pueden cruzar la membrana a baja velocidad gracias a su tamaño pequeño.
Las moléculas apolares pequeñas: difunden.
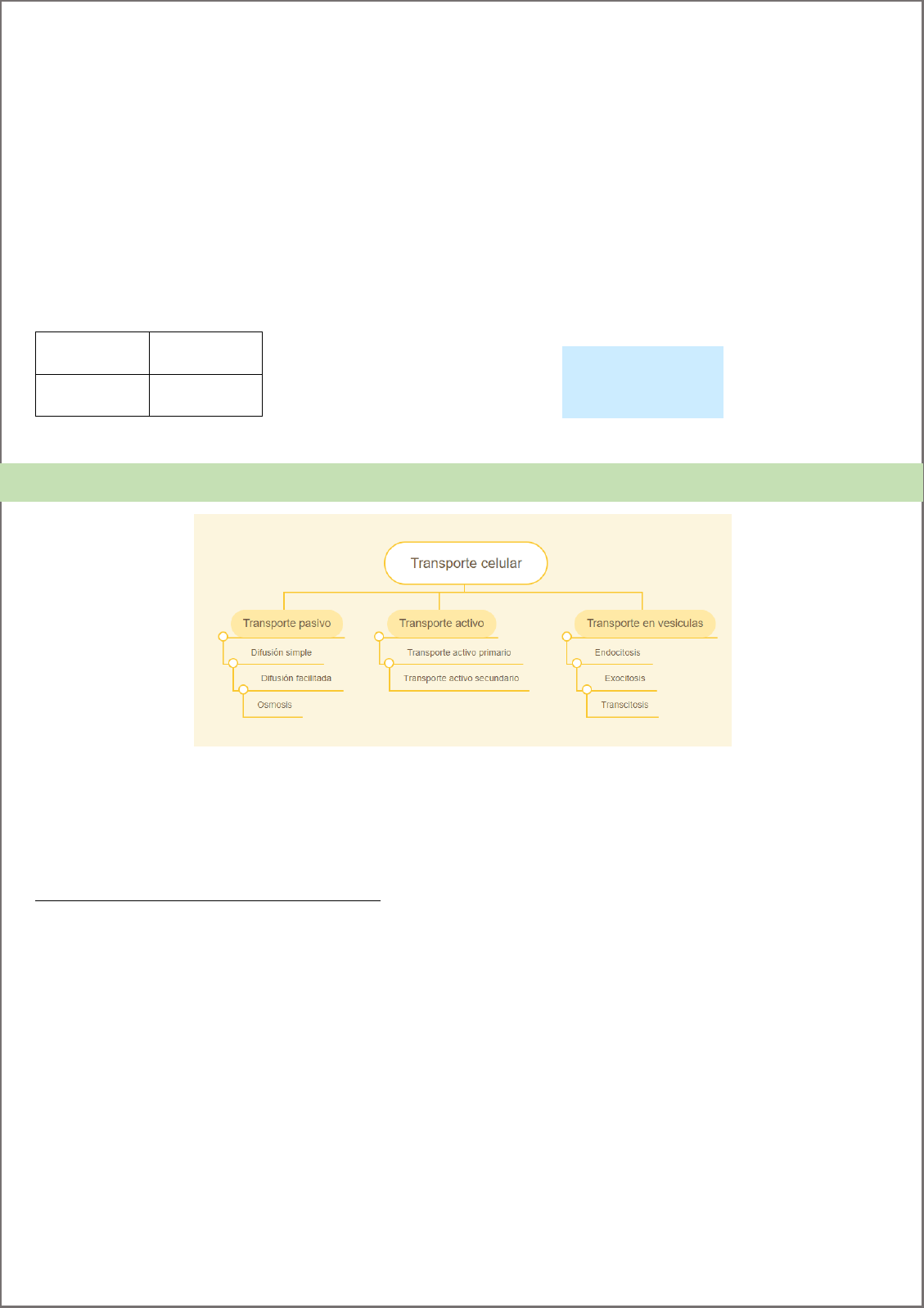
Clasificación según su requerimiento de energía
No requiere energía. Transporte pasivo, se da a por una fuerza impulsora favorable (a favor de su gradiente
electroquímico):
o Difusión simple: atraviesa la membrana sin ayuda
o Difusión facilitada: necesita de una proteína para atravesar membrana
Dentro de la
célula
K+, glucosa
Exterior
celular
Na+, Cl-,
Ca2+
“Los K adentro y
la CaNa afuera”
Transporte biológico
pág. 3 Agustina Colombo
Requiere energía
o Transporte activo primario: (ATP).
o Transporte activo secundario: fuerza
impulsora procedente del acoplamiento
contra gradiente de dicho soluto con el
movimiento a favor del gradiente de uno o
varios solutos.
o Para moléculas más grandes (vesícula):
Endocitosis: se invagina y se forma
una vesícula dentro del citoplasma.
Se denomina:
Pinocitosis: pequeñas
moléculas o solutos
Fagocitosis: moléculas
grandes.
Exocitosis: se fusiona con la
membrana plasmática y el
contenido se libera hacia el espacio
extracelular.
Difusión: Movimiento browniano: toda materia del universo
está en constante movimiento.
En la difusión simple: movimiento desde una región de alta
concentración a una región de menor concentración. Es proporcional a un gradiente. La diferencia de concentración
a los lados de la membrana genera un potencial químico, que va a permitir la difusión a través de la membrana de
sustancias sin carga e hidrófobas como compuestos endógenos (hormonas esteroideas), los gases y fármacos.
La difusión neta es el moviente que genera la sustancia a favor del gradiente de concentración.
La rapidez con la que se mueve la sustancia se describe por su flujo (Jx): número de moles de sustancia x que atraviesa
una unidad de superficie de membrana por unidad de tiempo.
Fórmula para el flujo:
Cuando se llega al equilibrio, el flujo debe ser neto debe ser 0.
Transporte mediado:
-pasivo: difusión facilitada.
o Poros: abierto la mayor parte del tiempo. Ej: porinas, perforinas, complejos de poros nucleares, aquaporinas
(canales agua).
o Canal: alterna entre canal abierto/cerrado, también pueden inactivarse.
Dependientes de voltaje
dependientes de ligando: se abren o cierran por efecto de hormonas, segundos mensajeros o NT. Por
ejemplo, el receptor nicotínico de acetilcolina (ACh) en la placa motora terminal es un canal iónico
que se abre cuando la ACh se une a él. Cuando está abierto, es permeable a Na+ y K+, lo que
provoca la despolarización de la placa motora.
activados por estímulos mecánicos.
Los canales iónicos pueden estar abiertos o cerrados.
La conductancia de un canal depende de la probabilidad de que el canal esté abierto. Cuanto mayor
es la probabilidad de que un canal esté abierto, más alta será la conductancia o permeabilidad. La
apertura y el cierre de los canales son controlados por compuertas
o Transportadores o acarreadores: compuertas nunca abiertas al mismo tiempo. Lo encierra y cambia de
conformación, se abre del lado opuesto.
Algunos transportadores facilitan la difusión pasiva de pequeños solutos. Los transportadores que no hidrolizan ATP o no
se acoplan a una cadena de transporte de electrones pertenecen a una superfamilia de transportadores de solutos
(SLC).
Factores que influyen la difusión:
- Gradiente de concentración
- Naturaleza química de la sustancia.
- Superficie de membrana
- Tamaño molecular
- Solubilidad de lípidos
Jx=Px ([X]e – [x]i)

pág. 4 Agustina Colombo
o Cotransportadores: transportan dos o mas solutos en el mismo sentido
o Intercambiadores (antiporter). Transportan solutos en sentidos opuestos.
-activo:
Transporte activo primario: ATPasas:
o P: bomba Na-K, Ca, H-K.
o V: transportan H+. rodean orgánulos intracelulares como lisosomas, endosomas, vesículas secretoras,
etc.
o F: en la membrana interna de la mitocondria, transportan H+
o Transportadores ABC: objetivos de muchos fármacos, ya que están presentes en muchos tejidos.
Ejemplo: Na-K ATPasa: se encarga de expulsar 3 iones Na+ por dos iones K+ que entran, con el hidrolisis intracelular de
ATP.
pág. 5 Agustina Colombo
Electrofisiologia introducción. Profesora Dra Villa-Abrille.
Galvani hizo un experimento al darle corriente al musculo y al nervio, viendo que generaba actividad.
Los iones son quienes van a transmitir la corriente dentro de las células, a través de transporte de membrana.
Potencial electroquímico: cuando las fuerzas de la tendencia a moverse se
equilibrio. OJO, las concentraciones no son las que se igualan. En este momento no
habrá FLUJO NETO DEL ION.
Potencial eléctrico: es la tendencia a moverse como consecuencia de la
diferencia de potencial eléctrico.
Potencial químico: tendencia a moverse de A a B debido al gradiente de
concentración. Alcanza el equilibrio químicos cuando es igual a ambos
lados de la membrana
El potencial electroquímico siempre se da cuando la sustancia a difundir por la membrana tiene carga.
Electrodifusión: proceso de difusión iónica a través de la membrana.
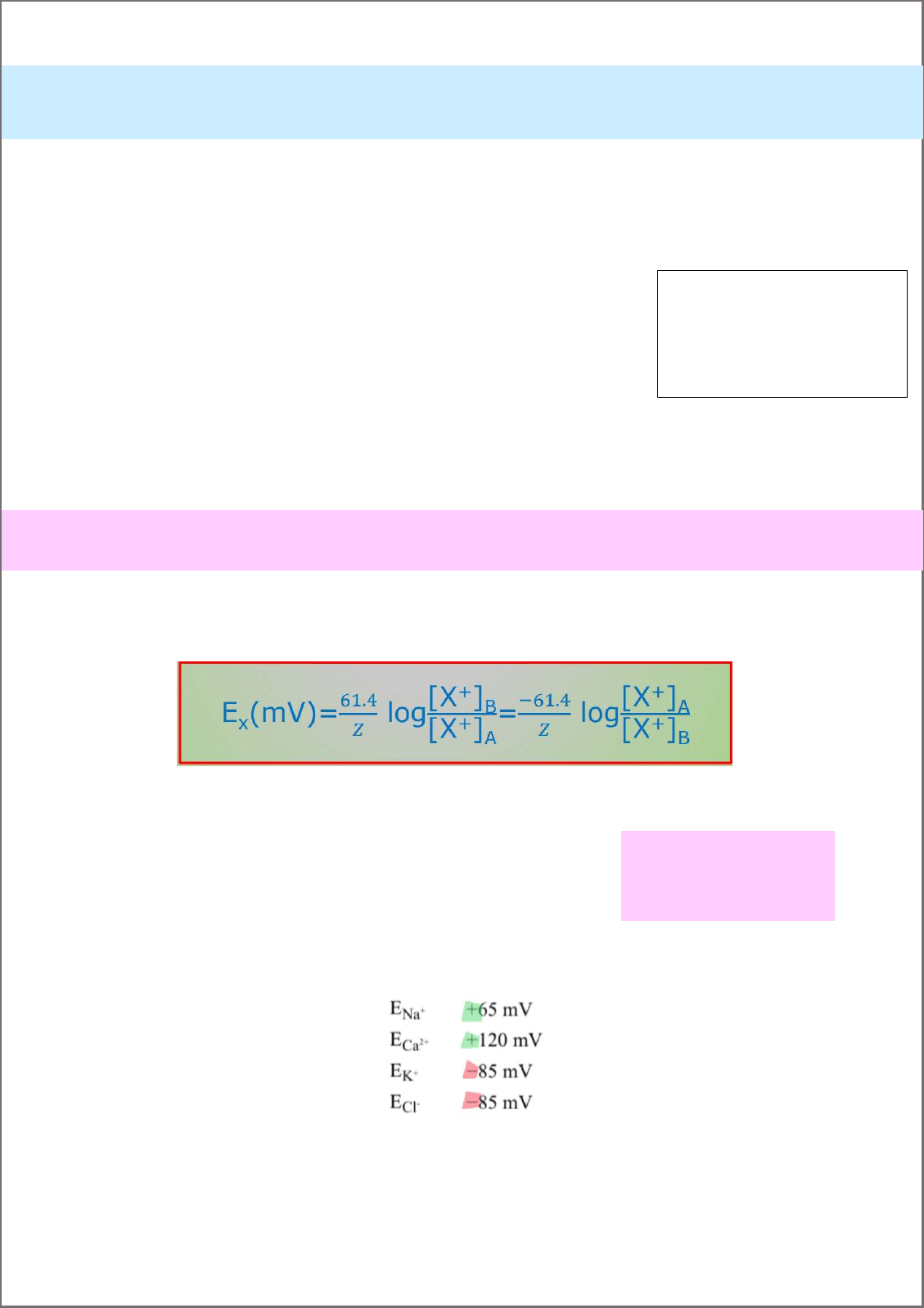
Ecuación de Nernst: me indica el potencial de equilibrio electroquímico(Ex) o potencial de Nernst para cualquier ion. Es
decir, me indica cuando un ion esta en equilibrio a través de la membrana.
61,4 a 37° (310K) siempre. El potencial de equilibrio cambia con la diferencia de temperatura.
T: temperatura absoluta en K
R: constante de gases: 8,31Joules/Kmol
F: constante de Faraday:96500C/mol
Voltios V_ Joules/C.
Valores aproximados de potenciales de equilibrio en el nervio y el musculo.
Potencial de membrana (Vm): diferencia de voltaje a través de la membrana celular (entre el citoplasma y el espacio
extracelular). Por convención, se expresa como el potencial intracelular con respecto al potencial extracelular (ej:
PMR: -70mV significa 70Mv más negativos en la célula.
Las modificaciones del potencial de membrana resultan de cambios en la conformación de proteínas estructurales de
la membrana plasmática llamados canales iónicos. Estos cambios conformacionales dan lugar a un aumento de la
Manitol: se agrega muchas
veces para igualar las
osmolaridades a ambos lados
de la membrana, evitando el
flujo de agua.
Electrofisiologia general.
Equilibrio electroquímico: se igual y oponen ambas fuerzas.
Tendencia a moverse debía al gradiente de concentración= tendencia a moverse por la diferencia de potencial eléctrico
Z: valencia del ion, es
decir, la cantidad de
carga

pág. 6 Agustina Colombo
permeabilidad selectiva de la membrana a iones que se encuentran distribuidos asimétricamente a ambos lados de la
membrana. Los cambios de permeabilidad y la distribución asimétrica de los iones dan lugar a corrientes iónicas que
median las modificaciones del potencial de membrana. Este tipo de actividad eléctrica de membrana es la base de
lo que se conoce con el nombre de respuestas activas
Los potenciales de membrana en reposo (PMR): valor constante de la membrana, donde no presenta respuesta
activa. Potencial que se tiene cuando no existe un estímulo o una distorsión externa de ese potencial. El PMR de las
células musculares, cardiacas y neuronas oscilan aproximadamente entre -60 y -90 m V; en el músculo liso se sitúa
cerca de los – 55m V. y los eritrocitos -9Mv.
Cada ion tiene a llevar la membrana hacia su potencial de equilibrio. Los iones con las permeabilidades o
conductancias más altas serán los que más contribuirán al potencial de membrana en reposo, y los iones con las
permeabilidades más bajas contribuirán poco o nada.
El PMR se da gracias a la alta conductancia en reposo al K+, que impulsa el potencial de membrana hacia el
potencial de equilibrio de K+. En reposo, las compuertas de activación de canales Na+ están cerradas, por lo que la
conductancia de Na+ es baja.
El ritmo de movimiento iónico a través de la membrana depende de la concertación interna y externa del ion X, del
voltaje transmembrana y de un coeficiente de permeabilidad para el ion X. Teniendo en cuenta estas afirmaciones:
1. la membrana es un medio homogéneo con un grosor a.
2. la diferencia de voltaje varia linealmente con la distancia a través de la membrana. Es decir, el campo
eléctrico (cambio de voltaje con la distancia) es constante a lo largo del grosor de la membrana
3. principio de independencia: el movimiento del ion a través de la membrana es independiente del movimiento
del resto de iones.
4. El coeficiente de permeabilidad (P
X
) es una constante. No varia con las fuerzas impulsoras químicas o
eléctricas. Px describe la capaciendad de un ion para disolverse en la membrana y de difundir de un lado a
otro a lo largo de la distancia a.
La ecuación de Goldman- Hodgkin –Katz: obtenemos el potencial de membrana, relacionando la corriente de los
iones a través de la membrana con las concentraciones internas y externas, el voltaje transmembrana y la
permeabilidad de la membrana.
T: temperatura absoluta a 37°C (310K)
R: constante de gases: 8.31 Joules/K mol
F: constante de Faraday: 96500 C/mol
P
x
: permeabilidad

pág. 7 Agustina Colombo
Todas las células tienen un potencial de membrana porque los iones se distribuyen alrededor de ella. Pero, no toda es
células excitables.
Célula excitable: Clásicamente se define como aquellas que frente a un estímulo
eléctrico cambian su potencial de membrana y producen potencial de acción. Pero
esto solo es para células nerviosas y musculares,
Pero, también existen otras células que ante el estímulo generan una respuesta. Como:
las B del páncreas, que ante el estímulo se despolariza liberando su secreción
(insulina). Entonces:
Definimos célula excitable cualquier célula que corresponde ante un cambio en el
potencial de membrana.
Fuerza impulsora:
Diferencia entre el potencial de membrana y potencial de equilibrio.
Cuando la fuerza impulsora neta es negativa los cationes entrarán a la célula y los aniones saldrán (Cuando el Vm es
más negativo que Ex un catión tendrá que entrar y un anión que salir).
Cuando la fuerza impulsora neta es positiva los cationes salen y los aniones entran.
Mayor sea la fuerza impulsora mayor es la corriente de iones observada.
o Corriente de entrada: corriente de iones que fluye hacia dentro de la célula. Cuando los valores de Vm son
más negativos, se genera una corriente de entrada + hacia la célula.
o Corriente de salida: corriente que fluye hacia afuera. Cuando el Vm es más positivo, la fuerza impulsora es
positiva, generando una corriente de salida +
PMR: -80,14
Si PM (Em)= Px (Ex) NO HABRÁ FLUJO DE X.
Fuerza impulsora: Vm (potencial de membrana) – Ex (potencial de eq del ion)
Células excitables:
Musculares lisas y
esqueléticas
Células
secretoras
neuronas
pág. 8 Agustina Colombo
Si PM > Px calculado con la ecuación de Nernst y con mismo signo la fuerza eléctrica es mayor que la química, X se
mueve en sentido de la fuerza eléctrica.
Si OM < Px calculado con la ecuación de Nerst con mismo signo la fuerza eléctrica es menor que la química, X se
mueve en sentido de la fuerza química.
Si PM tiene signo opuesto al Px, la dirección y el flujo de X es determinado por ambas fuerzas.
Cuando exista una diferencia de potencial eléctrico transmembrana, un ion puede estar distribuido en equilibrio aun
cuando su concentración no sea la misma en ambos compartimientos.
Flujo de corriente: solo ocurre cuando:
Una FUERZA impulsa el ion
La membrana es PERMEABLE al ion
Equilibrio Gibbs-Donnan: pasivo. Se tienen cuenta especies impermeables a la membrana.
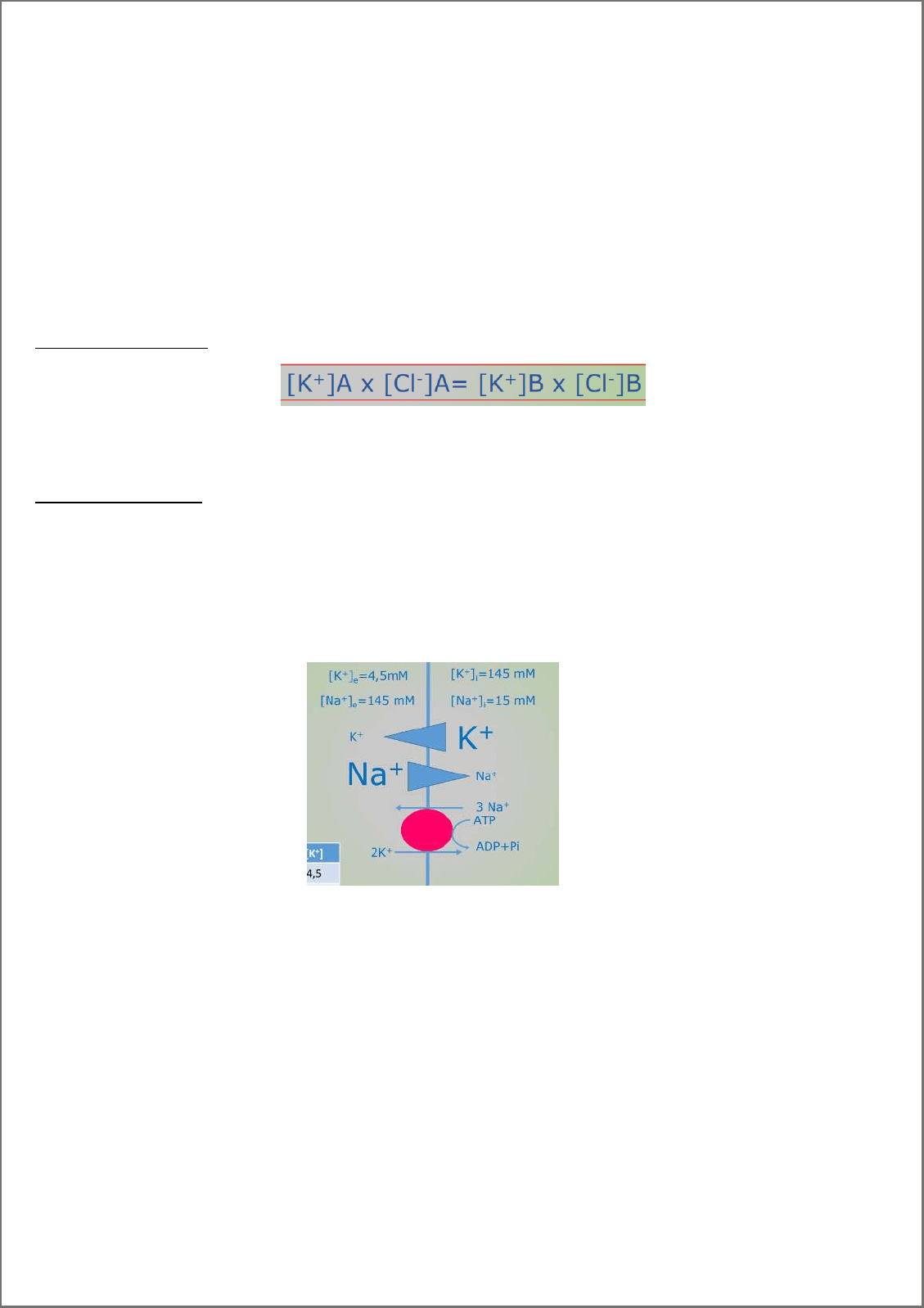
Bomba Na*/K+ ATPasa.
Transporte activo que bombea 3 NA+ afuera y 2K+ adentro de la célula, en contra de su gradiente. Sacando una
carga positiva, es una bomba electrogenica, o sea, genera un cambio de cargas.
No mantiene el potencial de membrana de reposo, pero contribuye a mantener los gradientes iónicos (contribución
indirecta con el PMR). Regula volumen celular y al bloquearla produce una despolarización.
pág. 9 Agustina Colombo
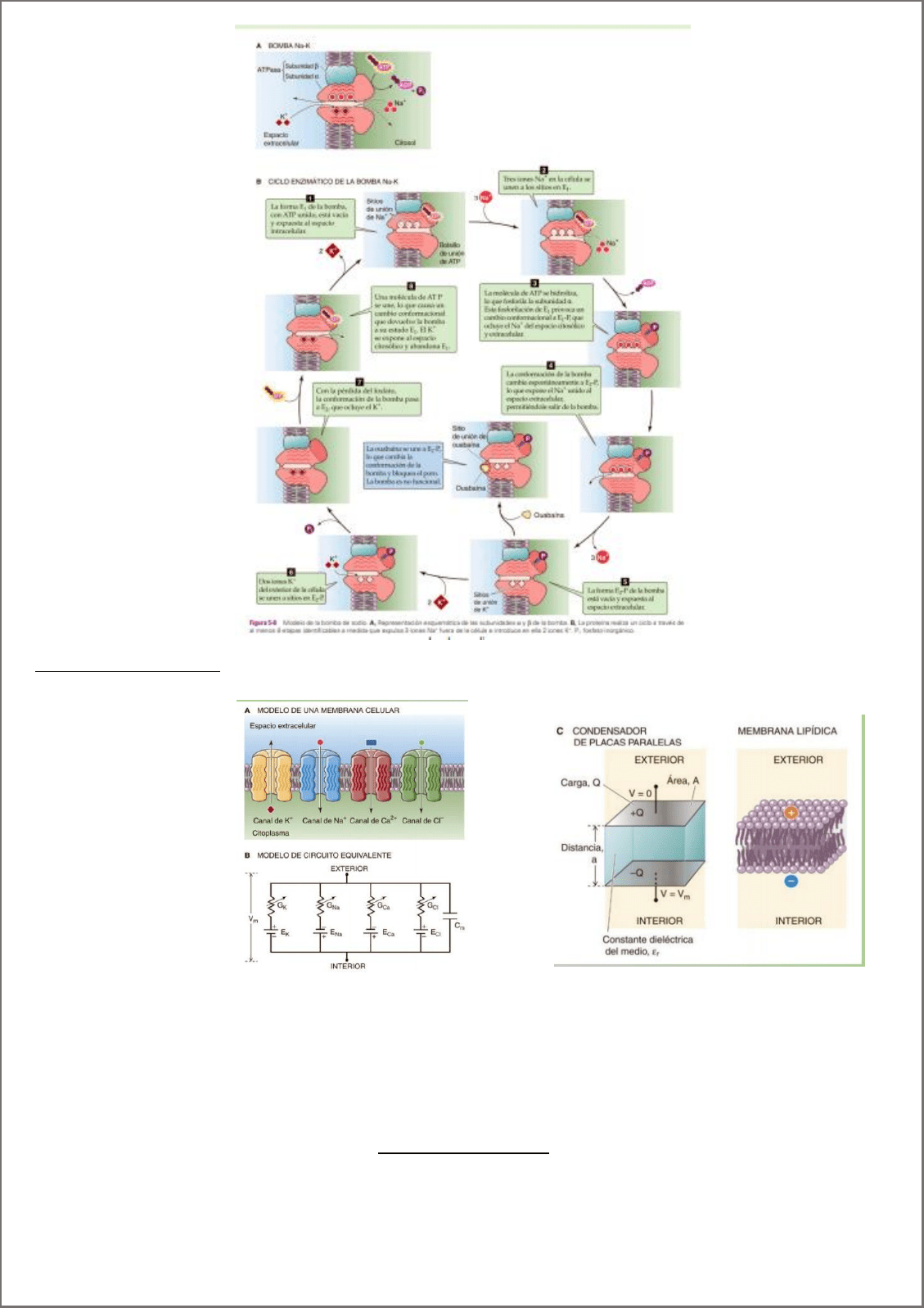
La membrana plasmática actúa como capacitor, tiene capacidad de separar cargas. Produciendo un potencial de
membrana.
-adentro
+afuera
Definiciones importantes:
Hiperpolarización: cambio negativo del Vm.
Despolarización: cambio postivo del Vm.
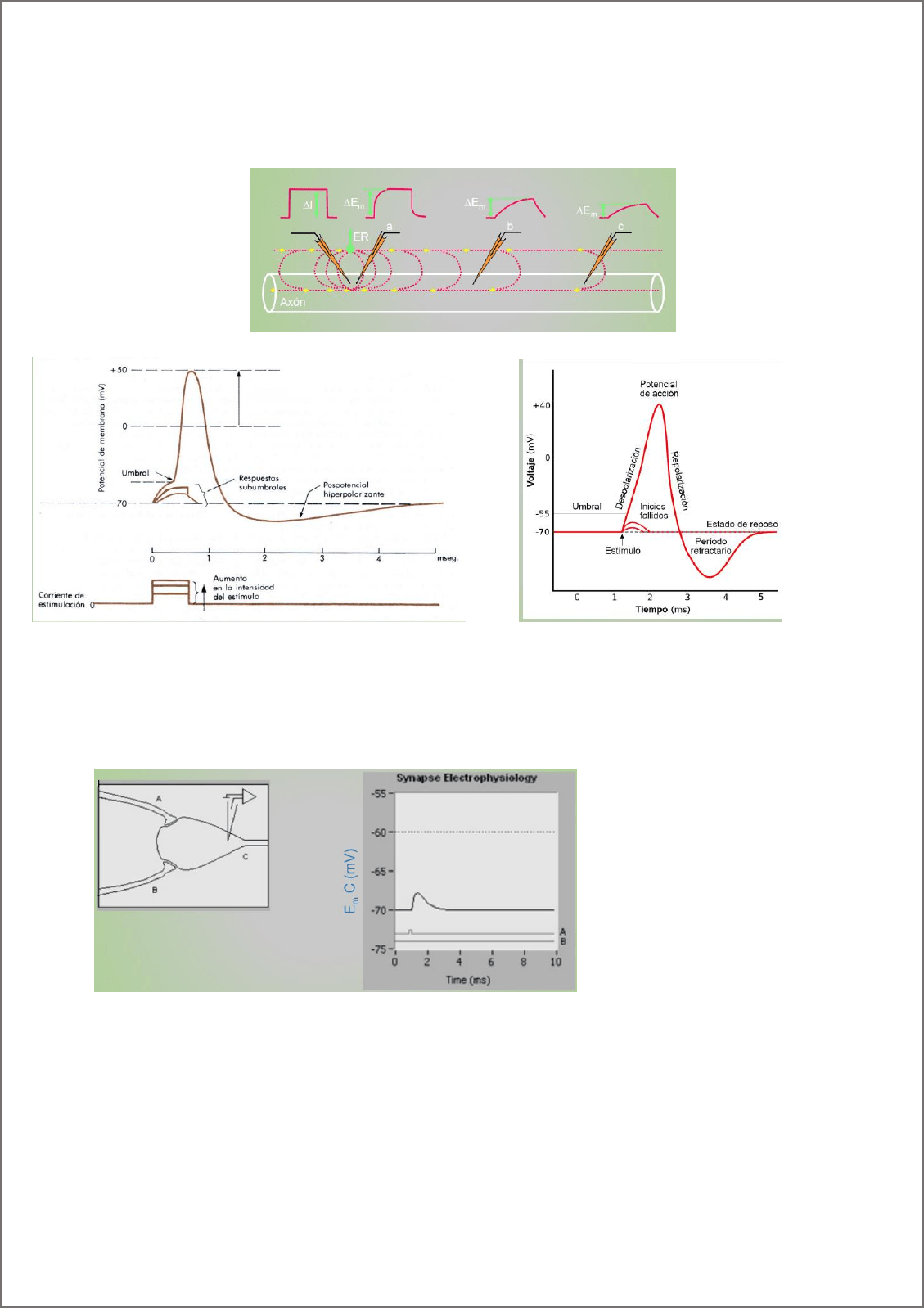
Potencial electrotónico:
Distorsión del potencial de membrana en reposo dada por una corriente catódica o anodica. A medida que discurre
el tiempo pierde su amplitud a medida que me alejo del sitio de inyección del estímulo (decrece en tiempo y espacio).
Características:
pág. 10 Agustina Colombo
Son locales, no se propagan
Subumbrales, no superan el valor umbral para dosparar al potencial de acción
Son pasivos: no cambia la polaridad de la membrana
Son SUMABLES en TIEMPO y ESPACIO.
Puede ser hiperpolarizante o despolarizante.
A nivel del sistema nervioso tenemos como potencial:
PEPS: potenciales post-sinápticos excitatorio. Despolarizantes. Si la sumatoria espacial y temporal superan el
potencial umbral se da el potencial de acción.
Se producen por la apertura de canales permeables a Na+ y K+, despolarizándose la membrana hasta un
valor medio entre los potenciales de equilibrio de Na+ y K+.
Neurotransmisores exitadores: Ach, noradrenalina, adrenalina, dopamina, glutamato, serotonina.
PIPS: potenciales post-sinapticos inhibitororio. Nunca llegaran al PA. Causados por la apertura de canales CL-.
El Vm se hiperpolariza hacia el potencial de equilibrio de CL-.
Neurotransmisores inhibidores: ácido-y-aminobutírico (GABA) y glicina
pág. 11 Agustina Colombo
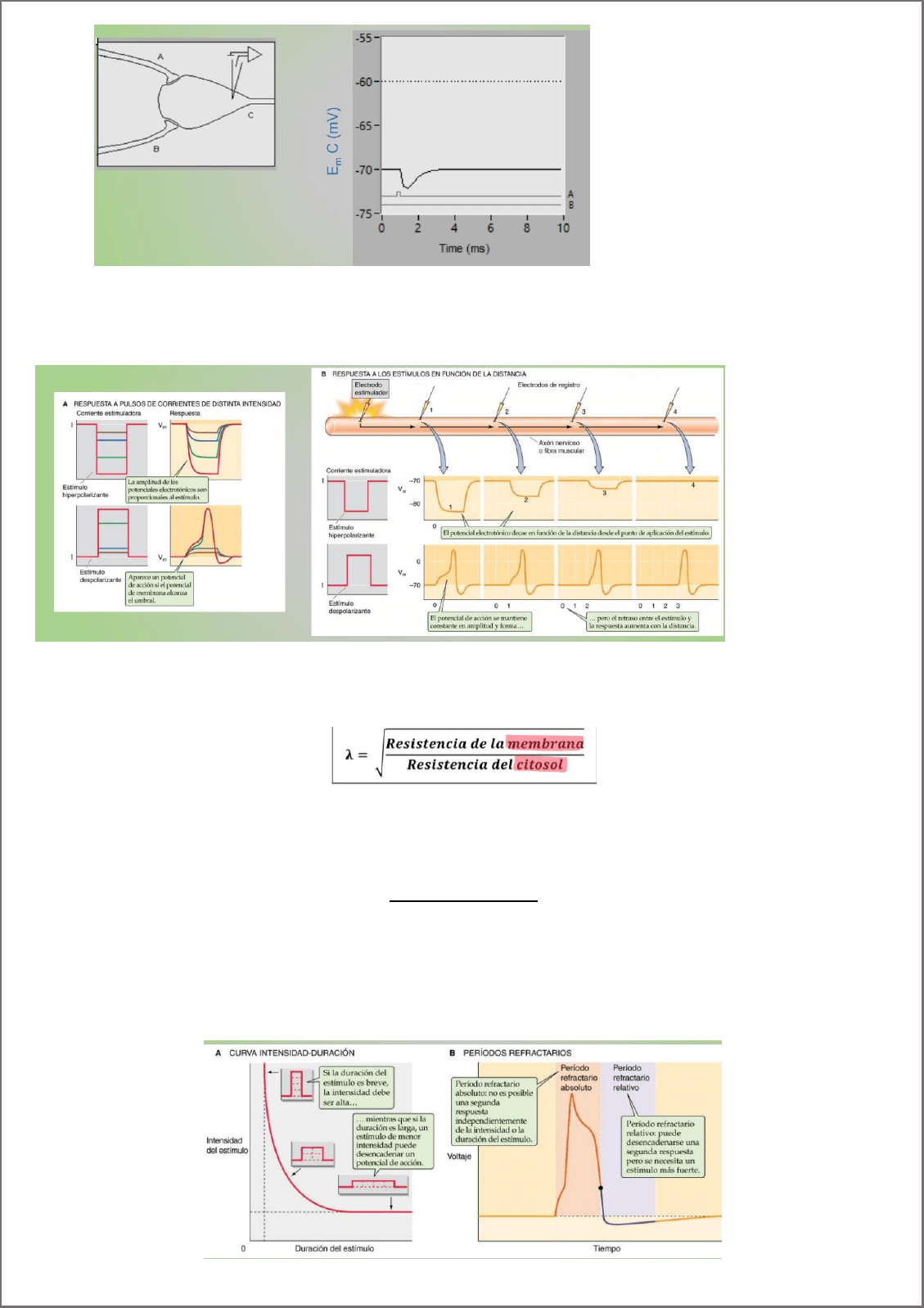
Si aplicamos un estímulo de cierta intensidad y por un cierto tiempo, podemos llegar a despolarizar la membrana para
que se produzca un potencial de acción
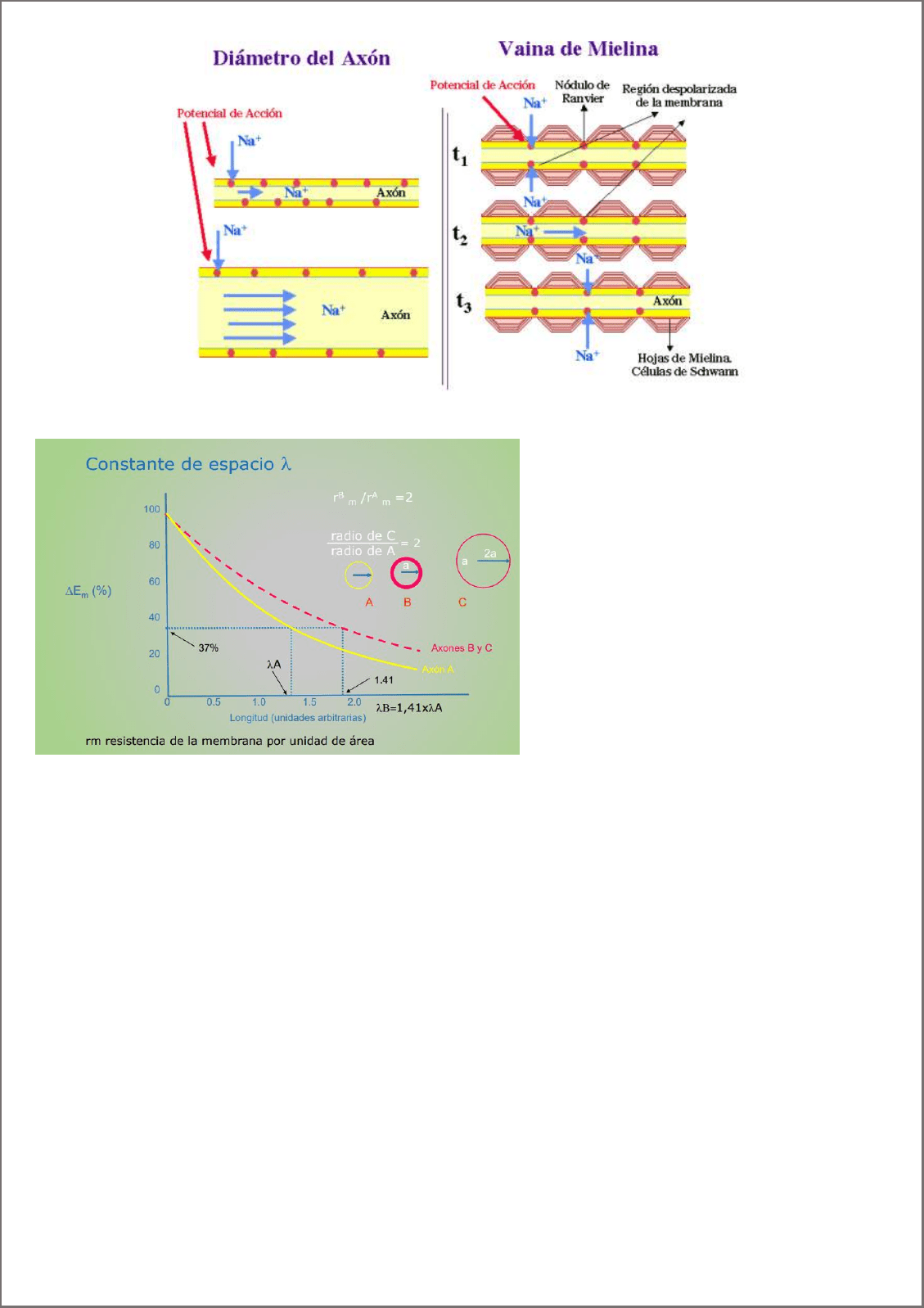
Constante espacio de membrana (λ): distancia del origen de un potencial electrotónico a la cual la amplitud que
llega del potencial es sólo el 37% de su valor máximo. Dado que la conductancia depende de la resistencia del citosol
y de la membrana.
Constante de tiempo (t): La constante temporal (τ), por otra parte, es una medida de la demora de la membrana en
volver a su potencial en reposo, y es el tiempo que tarda la variación del potencial en llegar al 63% de su valor final
Potencial de acción:
Propiedad de células excitables que es una modificación del potencial de membrana que se propagan manteniendo
siempre la misma duración y amplitud. Consiste en una rápida despolarización o fase de ascenso, seguida por
repolarización del potencial de membrana.
Producen una reacción. Se caracteriza por ser del tipo todo o nada, es decir, se da o no se da, si yo pongo una
mayor intensidad no se amplifica el PA o, en caso de ser más chica la intensidad, no puedo generarlo en absoluto).

pág. 12 Agustina Colombo
Estímulos:
Subumbral: Si no se produce el potencial de acción. Por debajo del
umbral.
Exitatorios: permiten llegar al potencial umbral.
Los estímulos pueden variar en:
Duración
Intensidad
Reobase: La mínima intensidad de estímulo que produce un potencial de acción
Cronaxia: cronos (tiempo) es el tiempo que le debo aplicar a un estímulo cuya intensidad es dos veces la rebase
Periodo refractario: No responde a un estímulo.
Absoluto: no se produce otro potencial de acción por más grande que sea el estímulo. Las compuertas de
desactivación de Na+ están cerradas cuando el potencial de acción se despolariza. Permanecen cerradas
hasta la repolarización
Relativo: si aplico una intensidad MUY grande, produzco un potencial de acción. Se encuentra entre el
preiodo refractario absoluto y el PMR. La conductancia para el K+ es mas alta que en reposo y el Vm está mas
cerca de su potencial de equilibrio, por lo tanto, más lejos del umbral. Entonces, necesito una corriente más
grande para llegar al umbral.
Fenómeno de acomodación: Ocurre cuando la membrana celular se mantiene a un nivel de despolarización tal que
se rebasa el potencial umbral sin que se dispare un potencial de acción. Se produce porque la despolarización cierra
las compuertas de desactivación en los canales de Na+. Se pone de manifiesto en situaciones de hiperpotasemia, en
que la alta concentración sérica de K+ despolariza la membrana de la célula musculoesquelética. Aunque el
potencial de membrana está más cerca del umbral, no se producen potenciales de acción porque la despolarización
cierra las compuertas de desactivación en los canales de Na+, lo que provoca debilidad muscular
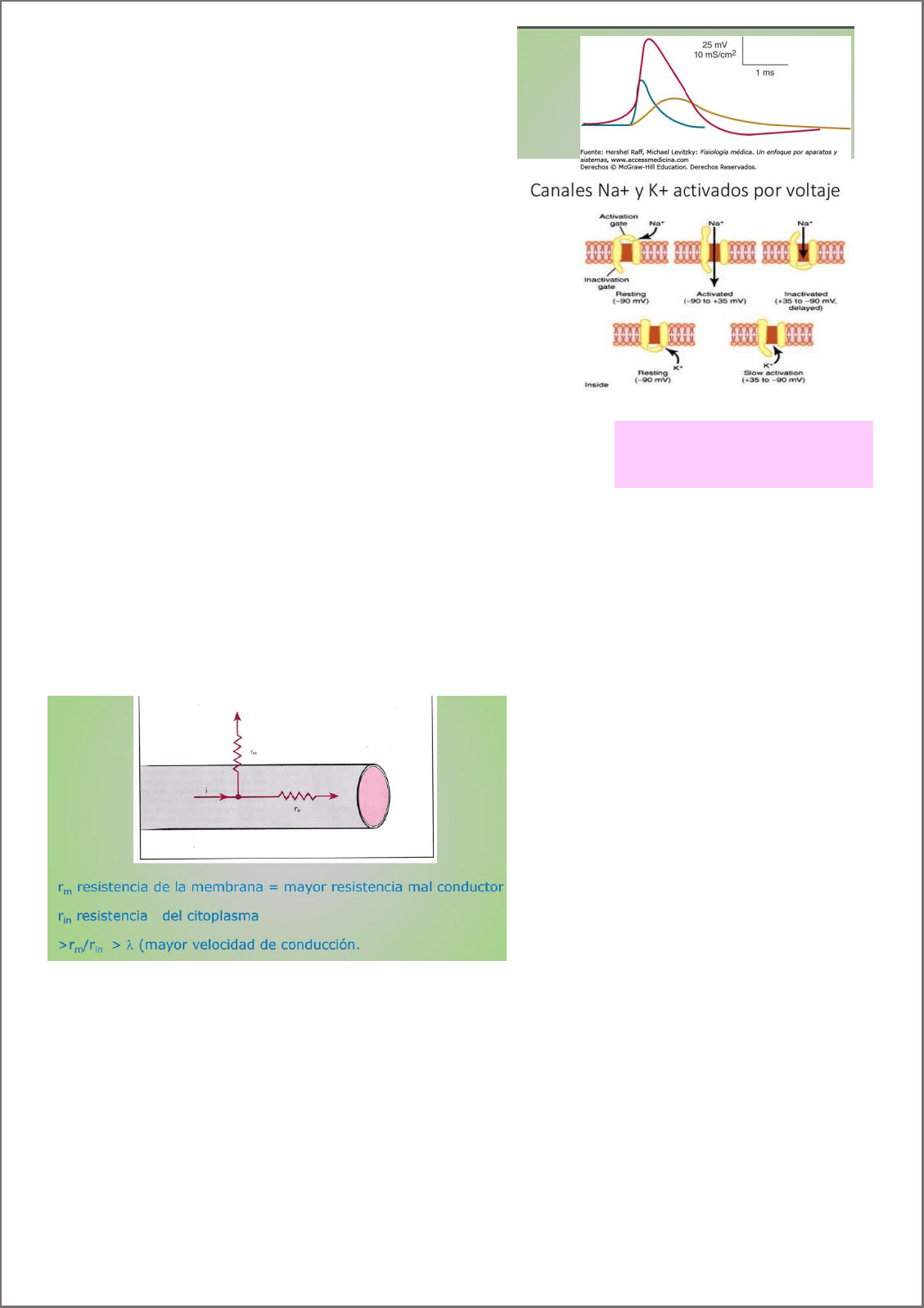
Potencial de acción:
Al llevar el potencial de membrana hacia una intensidad
umbral. Se activación de canales de NA+
¿como?
Dos compuertas:
Activación:
Inactivación.
Configuración cerrada: potencial de membrana en reposo.
La compuerta cerrada es la de activación y la de
inactivación abierta.
Configuración activa/abierta: se abre la puerta de
activación y el sodio entra por su gradiente electroquímico.
Despolarización de la membrana
Configuración inactivada: el mismo estimulo de
despolarización genera que la compuerta de inactivación se
cierre. La cinética con la que la compuerta de inactividad se
cierra es más lenta que la apertura de la de activación.
Permitiendo que en un lapso de tiempo estén abiertas las
dos. Canal inactivo.
¿Por qué no se llega a su potencial de equilibro el Na+?
Porque se inactivan los canales de Na+
UMBRAL: potencial de membrana al
que hay que llegar para obtener la
descarga del PA. La corriente de
entrada se hace más grande que la
de salida.
La despolarización se hace
autosostenidad.
pág. 13 Agustina Colombo
Porque se activan canales de K+
Los canales de K+, involucrados en la repolarización. Canales
operados por voltaje, que se activan a potenciales mucho mas
positivos.
Tiene una compuerta que se abre y el potasio sale a favor de
gradiente electroquímico.
Los canales de Na+, solo pueden ser activados cuando esta la
configuración cerrada, NO EN INACTIVO. Vuelven a pasar de
inactivo a cerrado, cuando vuelve el potencial en reposo.
ENTONCES: fases del PA:
1. Corriente de entrada despolariza la membrana
2. Apertura de Canales de Na+ (aumenta su
permeabilidad)
3. Se desplaza el potencial de membrana acercándose a
potencial de equilibrio del Na+.
4. Cierre de las compuertas de inactivación (conductancia mínima)
5. La despolarización abre lentamente canales K+ (aumento de su
conductancia)
6. Sale K+, repolarizando la célula.
7. La conductancia de K+, permite acercar el potencial de membrana a su potencial de equilibrio, lo que puede
generar una hiperpolariazción.
A medida que el potencial de acción se propaga depende de:
Resistencia de la membrana (r
m
)
Resistencia citoplasmática (R
in
)
>relación Rm/Rin >velocidad de conducción. Ya que se recorre más distancia disipándose menos.
Conductancia= permeabilidad
del ion
pág. 14 Agustina Colombo
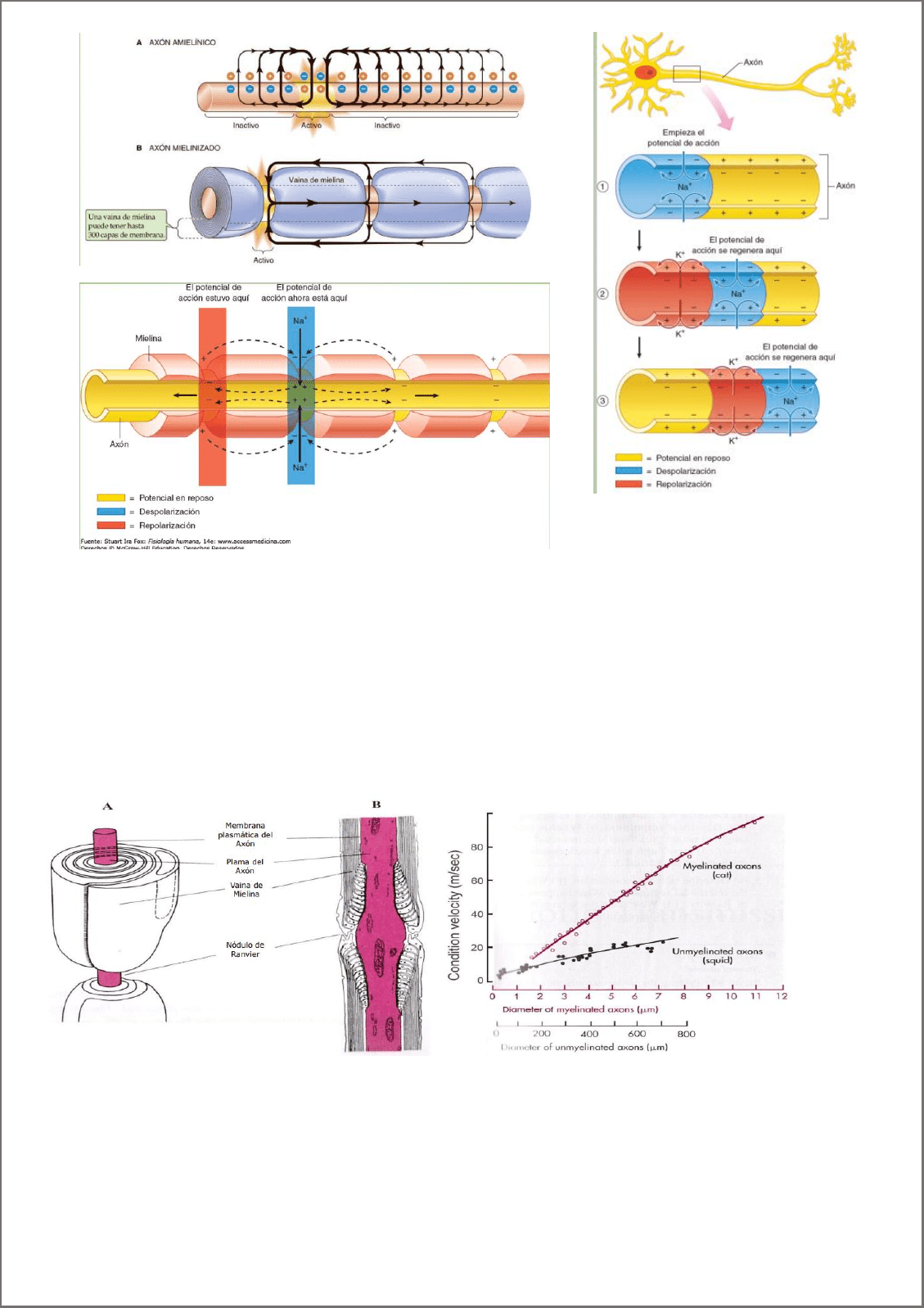
La propagación de los PA Ocurre por propagación de corrientes locales a zonas adyacentes de la membrana, que
entonces se despolarizan hasta el umbral y generan potenciales de acción
Entonces:
Mayor espesor de la membrana (diámetro de la fibra nerviosa), genera que se recorra más distancia, porque
se disipa menos. Mayor velocidad El radio diámetro mayor, (axón más ancho) mayor velocidad.
Mielinización: Las vainas de mielina actúan como aislantes, y se encuentran entre los nodos de ranvier. Entre
ellas, no se produce el potencial de acción, la conducción se da por potenciales electrónicos, pero es muy
rápida y se disipa menos, así que no se pierde tanta intensidad hasta el próximo nodo de ranvier en donde se
produce el potencial de acción. Esta conducción se denomina conducción saltatoria.
velocidad de conducción 1
pág. 15 Agustina Colombo
Este parámetro se mide con la constante de espacio.
Este documento contiene más páginas...
Descargar Completo
unidad 1 terminada.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.