
PATOLOGIA HUMANA
Page
1
of
10
UNIDAD 1: INTRODUCCIÓN A LA PATOLOGÍA
LA CÉLULA COMO UNIDAD DE SALUD Y ENFERMEDAD
Definición de patología: estudio del sufrimiento, o el estudio de la enfermedad. Virchow la denomino patología celular
para destacar que todas las enfermedades tienen origen en la célula, siendo este el principio que rige a la patología
moderna, entonces es el estudio de las anomalías celulares.
1. GENOMA:
Contiene aprox. 3.200 millones de pares de bases de ADN. Del total del genoma, solo el 1.5% corresponde a ADN
codificante. Este 1,5% también se puede entender como 20.000 genes que van a codificar 20.000 proteínas (este numero
es tentativo ya que no siempre es un gen = una proteína) que funcionaran como enzimas, componentes estructurales o
moléculas transmisoras de señales, que se usan para ensamblar y mantener todas las células del organismo. Hay
organismos que con menos pares de bases que los de los seres humanos, poseen, de igual manera, 20.000 genes que
formaran 20.000 proteínas, entendiéndose que la diferencia entre distintos organismo vivos, esta dada por el ADN NO
CODIFICANTE.
1.1 Adn no codificante:
Corresponde a 98.5% del genoma que no codifica proteínas, y que se encuentra entre los genes que si las codifican.
Las principales clases de secuencias codificantes no proteínicas comprenden las siguientes:
Regiones promotoras e intensificadoras que aportan lugares de unión para factores de transcripción.
Lugares de unión para factores que mantienen estructuras cromatínicas.
ARN reguladores no codificantes: es ARN que no se traduce en proteínas, pero regulan la expresión génica,
como por ej micro-ARN, ARN no codificantes largos.
Elementos genéticos móviles (transposones).
Regiones estructurales especiales del ADN: telómeros y centrómeros.
Es importante destacar que la gran mayoría de variaciones genéticas (polimorfismos) asociadas a enfermedades están
situadas en las regiones del genoma que no codifican proteínas, por lo que la regulación génica podría ser el principal
precursor de enfermedades.
Otro dato importante es que los seres humanos compartimos el 99.5% de información genética, lo que indica que la
variación interindividual entre un humano y otro, incluida la distinta susceptibilidad a las enfermedades y la respuesta a
compuestos y sustancias ambientales, esta codificada en menos del 0.5% de nuestro ADN, lo que corresponde a 15
millones de pares de bases.
Las dos formas mas frecuentes de variación del ADN son:
Polimorfismos de nucleótido único (PNU): variantes
de posiciones de un único nucleótido que casi
siempre suelen ser bialelicas, es decir, que solo
existen dos posibles nucleotidos para ese alelo, (Si
no es G es C). Pueden estar presentes en todo el
genoma (exones, intrones y regiones intergénicas),
pero los que se encuentran en regiones no
codificantes pueden estar en elementos reguladores
del genoma, influenciando directamente sobre la
susceptibilidad a enfermedades. Aunque existen PNU neutrales, los cuales no generarían efectos sobre la
función génica o el fenotipo en el portador, podrían ser marcadores de interés si se heredaran
conjuntamente con un gen asociado enfermedades especificas debido al desequilibro de ligamiento,
Variaciones en el número de copias (VNC): consiste en secuencias largas de ADN que se encuentran
repetidas en números variables entre una persona y otra. Suelen ser bialelicas y ademas suelen darse por
duplicación, eliminación o reordenamiento. Se entienden como la gran razón para la diferencia fenotipica
entre individuos debido a que se encuentra mayormente en en secuencia codificantes de genes la mayoría

PATOLOGIA HUMANA
Page
2
of
10
afectan a secuencias codificantes variaciones fenotípicas.
Epigenética: son los cambios heritable en la expresión génica que no están causados por variaciones en la
secuencia de ADN. Siendo esta la explicación de la variación fenotipica entre gemelos monocigoticos,
explicándose así la diversidad genética.
1.2 Organización de las histonas
Histona: Tipo de proteína que se encuentra en los
cromosomas. Las histonas se unen al ADN, ayudan a dar su
forma a los cromosomas y ayudan a controlar la actividad
de los genes.
Histonas y factores modificadores de histonas:
son segmentos de ADN de 147 pb envueltos
alrededor de una estructuras central de histonas.
El complejo ADN-Histona resultante quedan
unidas por pequeños fragmentos de ADN no unido
a histonas (cromatina). En la mayoría de los casos
no esta enrollado uniformemente, debido a esto
podemos encontrar que la cromatina adopta dos
formas básicas: Heterocromatina y eucromatina.
Existen complejos remodeladores de la cromatina que regulan las histonas, pudiendo recolocar los
nucleosomas en el ADN, exponiendo o regulando elementos reguladores génicos. Ademas existen escritores
de cromatina que generan marcas en las histonas, las cuales son alteraciones covalentes (metilación,
acetilación y fosforilación de residuos de aminoácidos específicos), con el fin de indicar el ADN accesible para
replicación a las ARN-pol (también marca el que no es accesible). Estas marcas son reversibles mediante la
actividad de borradores de cromatina, ademas existen también lectores de cromatina que se unen a las
histonas con determinadas marcas regulando así la expresión génica.
Metilación de histonas: las lisinas y argininas en las histonas pueden ser metiladas para indicar la activación o
represión de la transcripción
Acetilación de histonas: incrementan la transcripción mediante la acción de la histona acetiltransferasa(HAT) la
cual genera la apertura de la cromatina. Es revertido por la histona desacetilasa (HDAC)
Fosforilación de histonas: los residuos de serina pueden fosforilarse con el fin abrirse para transcripción o
condensarme para quedar inactivo dependiendo del residuo al que se una.
Metilación del ADN: Un nivel elevado de metilación de ADN en los elementos reguladores de genes produce
silenciamiento de la transcripción. Posee los mismos reguladores que la metilación de histonas.
Factores organizadores de la cromatina: son proteínas unidas a regiones no codificantes que controlan la
formación de bucles de ADN, para regular las relaciones espaciales entre potenciadores y promotores
génicos.
1.3 Micro-arn y arn largo no codificante
Es importante para la regulación génica, este es ADN que no se codifica pero que si se transcribe.
Los miARN son ARN cortos (21-30 nucleótidos); no codifican proteínas, sino que están implicados en el
silenciamiento postranscripcional de la expresión génica. Los miARN pueden regular multiples genes
codificantes de proteínas. La transcripción de los miARN requieren varios pasos: producción de un miARN
primario, recorte mediado por la enzima DICER, generación de miARN maduro que que se asocia a un
complejo multiproteínico denominado: silenciador inducido por ARN (RISC), el cual induce el la escisión del
ARNm o reprimir su traducción. Los ARN de interferencia pequeños (pARNi) es ARN sintético, son breves
secuencias de ARN que pueden introducirse en las células, actuando de modo análogo a los miARN
endógenos.
El ARN largo no codificante (ARNInc) está implicado en la modulación de la expresión génica al unirse a
regiones de cromatina, restringiendo el acceso de la ARN pol a genes codificantes de region. al facilitar la
unión al factor de transcripción (favoreciendo activación génica) y facilita la modificación de la cromatina o
proporciona un andamiaje para estabilizar su estructura.
PATOLOGIA HUMANA
Page
3
of
10
2. MANTENIMIENTO CELULAR
Las funciones de mantenimiento de todas la células diferenciadas son:
Protección frente al entorno
Obtención de nutrientes
Comunicación
Movimiento
Renovación de moléculas envejecidas
Catabolismos celular y generación de energía
Muchas de estas funciones están están compartimentalizadas dentro de orgánulos intracelulares rodeados de membrana
1.1 Membrana plasmática: Protección y obtención de nutrientes
La MP es una bicapa fluida de fosfolípidos anfipáticos con cabezas hidrófilas enfrentadas al entorno acuoso y colas
lipídicas hidrófobas que interactuan entre si para formar una barrera a la difusión pasiva de moléculas grandes o con
carga
Existe una composición heterogénea asimétrica de los fosfolípidos dada por:
Fosfatidilinositol: en la cara interna de la MP, importante en la formación del andamio para proteínas celulares cuando
este es fosforilado y también para la transducción de señales debido a que generan inositoltrifosfato y diacilglicerol
mediando por fosfolipasa C
Fosatidilserina: restringido a la cara interna, confiriéndole una carga negativa a esta cara. Cuando se gira a la cara
extracelular es señal para los fagocitos en la apoptosis
Glucolipidos y esfingomielina: en cara extracelular, son importantes para las interacciones entre células y entre celula-
matriz, entre ellos el reclutamiento de células inflamatorias o las interacciones espermatozoide-ovulo.
En la membrana plasmática se insertan libremente muchas proteínas y glucoproteínas distintas implicada en: transporte
de iones y metabolitos, captación de macromoléculas en fase liquida y mediada por receptores, y interacciones entre
células y ligandos, la matriz u otras células.
Las proteínas de membrana se relacionan con las membranas lipídicas mediante interacciones:
Las proteínas transmembrana tienen uno o más segmentos alfa-helicoidales hidrófobos que atraviesan la MP.
Interacciones por unión postraduccional a grupos prenilo o ácidos grasos, insertados en la MP.
Las proteínas de membrana periféricas pueden asociarse de forma no covalente a las proteínas transmembrana.
Varias proteínas de la MP actúan de forma conjunta con complejos ensamblados por ejemplo en el RER o
seforman por difusión lateral en la MP.
La cara extracelular de la MP contiene hidratos de carbono en las glucoproteínas y los glucolípidos, así como ·[ de
cadenas de polisacáridos unidas a los proteoglucanos integrantes de la membrana. Este glucocáliz actúa como
barreraquímica y mecánica, y media las interacciones intercelulares y célula-matriz.
Difusión pasiva a través de la membrana: Las moléculas pequeñas no polares (O2 y CO2) y las hidrófobas (esteroides y
vitamina D) difunden a través de la MP a favor de su gradiente de concentración. Las moléculas pequeñas polares
(agua, etanol y urea) atraviesan la membrana, y cuando se trata de grandes volúmenes de agua se requieren de
acuaporinas.
Transportadores y canales: Las moléculas polares (azúcares y nucleótidos) e iones precisan transportadores proteínicos
especializados:
Proteínas canal: crean poros hidrófilos que se abren, permitiendo el paso de solutos (limitado por tamaño
ycarga). Los gradientes de concentración y/o eléctrico impulsan el movimiento del soluto.
Proteínas transportadoras: se unen a un soluto específico y experimentan un cambio conformacional
paratransportar al ligando a través de la MP. Utilizan ATP o energía liberada del gradiente iónico acoplado.
Dado que las membranas son plenamente permeables a las moléculas polares pequeñas, el agua las atraviesa
siguiendo las correspondientes concentraciones de solutos. Por tanto, la presencia de una mayor cantidad de sal
extracelular en relación con el citosol (hipertonía) provoca una salida neta de agua de las células, mientras que la
hipotonía induce unaentrada neta del agua hacia su interior.
PATOLOGIA HUMANA
Page
4
of
10
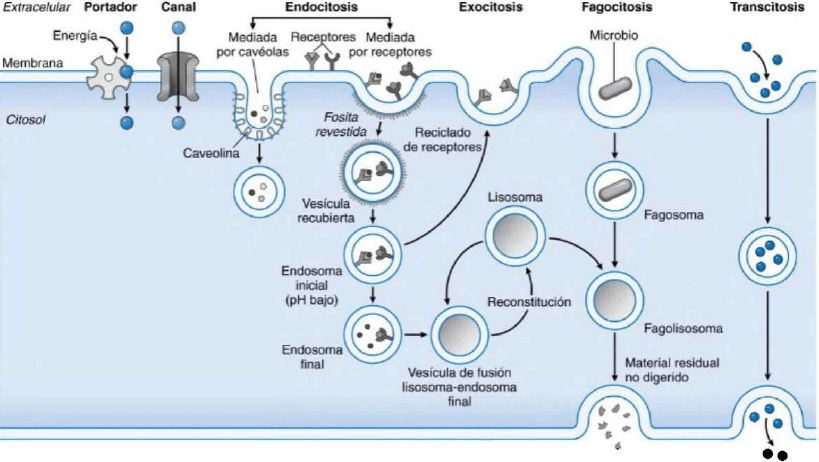
Captación en fase líquida y mediada por receptores: captación de fluidos o macromoléculas por parte de la célula
denominada endocitosis
La endocitosis permite la importación de macromoléculas:
Endocitosis mediada por caveólas: las caveólas son invaginaciones de la MP sin revestimiento asociadas a
moléculas unidas a GPI, proteínas de unión a AMPc, kinasas de la familia SRC y receptor de folato. Estas
atrapanmoléculas y liquido extracelular.
Pinocitosis: es un proceso en fase líquida en la que la MP se invagina hasta separarse de la membrana hacia
el interior celular, formando vesículas citoplasmáticas. La pinocitosis y la endocitosis mediada por receptores
comienza en la “fosita revestida por clatrina” de la MP. Estas vesículas pierden su revestimiento y se
fusionan con los endosomas iniciales, los cuales procesan parcialmente el contenido antes de pasar a los
lisosomas.
Endocitosis mediada por receptores: tras unirse a los receptores localizados en las fositas revestidas por
clatrina,la LDL y transferrina son endocitadas en vesículas, hasta que en un medio acido se liberan y luego
pueden ser reciclados a la MP.
La exportación celular de macromoléculas se denomina exocitosis, un proceso que lo utilizan, por ejemplo, aquellas
proteínas sintetizadas y empaquetadas en el RER y aparato de Golgi (las vesículas se fusionan con la MP y expulsan el
contenido). La transcitosis, es el movimiento de las vesículas endocitadas desde la cara apical hacia la basolateral de
lascélulas, permitiendo el transporte de grandes cantidades de proteínas intactas a través de las barreras epiteliales
(por ejanticuerpos ingeridos de la leche materna).
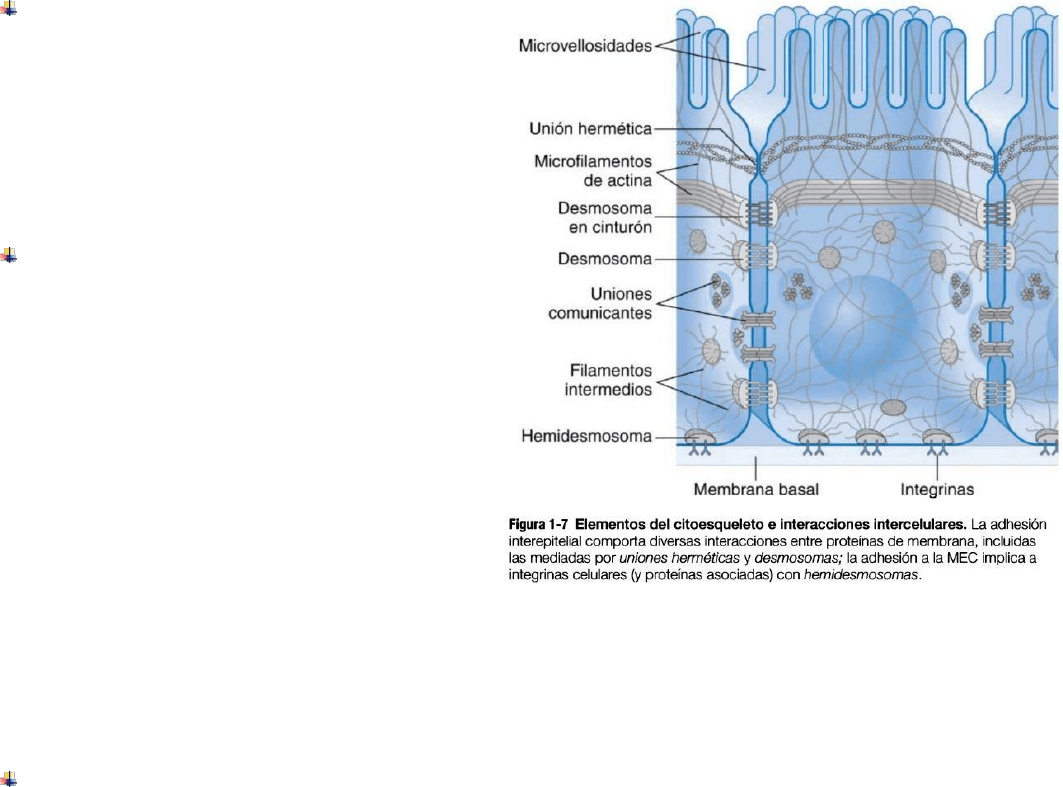
1.2 Interacciones intercelulares y citoesqueleto La forma y polaridad de la célula, el tráfico intercelular y la
motilidad dependen de las proteínas del citoesqueleto:
Microfilamentos de actina: fibrillas formadas a partir de actina globular (G-actina). Los monómeros de G-
actina se polimerizan de forma no covalente para generar filamentos largos (F-actina), que forman hélices
polares; se añaden o sacan subunidades globulares. En las cél no musculares, las proteínas unidas a actina la
organizan en haces y redes que controlan la forma y el movimiento celular. En las musculares, se produce
contracción por acción de la miosina que se desliza sobre los filamentos de actina.
Filamentos intermedios: fibrillas que presentan una forma polimerizada y soportan la tensión mecánica
(importante en piel y pelo)
Microtúbulos: fibrillas compuestos por dímeros polimerizados no covalentemente de tubulina alfa y beta.
Tienen polaridad definida, donde el extremo – encaja con un centro organizados de microtúbulos
(centrosoma) próximo al núcleo asociado a centriolos emparejados, mientras que el extremo + se alarga o
retrae, añadiendo o eliminando dímeros de tubulina. Generan “andamios” para proteínas motoras que
desplazan vesículas y orgánulos por las células: las cinesinas son proteínas motoras destinadas al transporte
anterógrado, mientras que las dineínas se encargan del transporte retrógrado; también participan en la
separación de cromátidas hermanas y forman cilios móviles o flagelos.
PATOLOGIA HUMANA
Page
5
of
10
Las células interactúan y se comunican por medio de complejos de unión:
Uniones oclusivas (uniones herméticas): sellan
células adyacentes entre sí, creando una barrera
continua que limita el movimiento paracelular de
iones y otras moléculas. Son uniones formadas
por proteínas transmembranas: ocludinas,
claudinas. Se disocian o reforman a fin de
facilitar la proliferación epitelial o migración de
células inflamatorias.
Uniones de anclaje (desmosomas): unen
mecánicamente las células a otras o a la MEC.
Mácula adherente: formados por asociación
de cadherinas (glucoproteínas
transmembranas) las cuales se unen a
filamentos intermedios intracelulares para
distribuir las fuerzas extracelulares entre
múltiples células.
Desmosomas en cinturón: cadherinas que se
asocian a microfilamentos de actina
intracelulares, que pueden determinar la
forma y la motilidad.
Hemidesmosomas: integrinas (proteínas de conexión transmembrana) se unen a filamentos
intermedios,vinculando el citoesqueleto a la MEC.
Complejos de adhesión focal: complejos macromoleculares localizados en los hemidesmosomas e
incluyen proteínas que generan señales intracelulares cuando las células están expuestas a fuerzas
mecánicas.
Uniones comunicantes(uniones en hendidura): median el paso de señales químicas o eléctricas entrecélulas,
formadas por conexinas. La permeabilidad de las uniones comunicantes permite la comunicación intercelular.
1.3 Maquinaria biosintetica: reticulo endoplasmatico y aparato de golgi
Las proteínas estructurales y las enzimas de la célula se renuevan constantemente mediante una síntesis
continua cuidadosamente equilibrada con su degradación intracelular
El RE es el lugar de síntesis de todas la proteínas transmembranas y los lípidos de la mp y de los organilos
celulares, incluido el propio RE, también produce todas las moléculas destinadas a salir de la célula.
Es un laberinto interconectado a modo de malla que se compone por dos dominios contiguos pero distintos: el
RER y REL
El proceso de traducción del ARNm dado por los ribosomas en la cara citosolica del RER produce proteínas
destinadas a la luz del RER o para su membrana, proceso dirigido por las secuencia señal en el extremo N-
terminal de las proteínas nacientes. Las que no poseen la señal se traducen en los ribosomas libres
(generalmente polirribosomas) del citosol los cuales suelen quedar en el citoplasma.
En el RE las proteínas deben plegarse y pueden formar complejos polipeptidicos (oligomerizacion), ademas se
forman enlaces disulfuro y se añaden oligosacaridos unidos a N, las moléculas chaperonas son las encargadas
de retener las proteínas en el RE hasta que todas sus modificaciones hayan sido completadas. El ejemplo de
mutación patógena mas frecuente es el del transportador de membrana en la fibrosis catequista, que ademas
el acumulamiento de proteína mas plegadas en el RE es un inductor a la respuesta al estrés del RE que
conlleva a la apoptosis.
El aparato de golgi se encarga del transporte de los productos del RE. Esta constituido por cisternas apiladas
que modifican progresivamente las proteínas de una forma ordenada desde la cara cis (proxima al RE) a la
trans (mas lejana), y que se contactan entre si por medio de vesículas. Las modificaciones que realiza durante
el transporte son es que los oligosacaridos unidos a N son podados y modificados, ademas de añaden
oligosacaridos unidos a O, lo cual es importante para el transporte de las moléculas a los lisosomas o para la
interacción intercelulares o célula*matriz
PATOLOGIA HUMANA
Page
6
of
10
La red golgi cis ademas puede reciclar proteínas y la red golgi trans clasifica proteínas y lípidos y los envía a
otros orgánulos. El complejo de golgi es principalmente importante en células especializadas en la secreción,
como las células caliciformes del intestino.
El REL es relativamente escaso en las cellas y existe básicamente como una zona de transición entre el REL y el
aparato de golgi, excepto en células donde se sintetizan hormonas asteroide (gónadas o glándulas
suprarrenales) o catabolizan moléculas liposolubles (hígado) donde pueden ser especialmente visibles.
1.4 Eliminación de desechos: Lisosomas y proteosomas
Lisosomas: orgánulos rodeados pormembrana que contienen hidrolasas ácidas (enzimas que funcionan mejor con ph
ácidos, las cuales incluye: proteasas, nucleasas, lipasas, glucosidasas, fosfatasas y sulfatasas. Estas se sintetizan en la
luz del REL y se marcan con un residuo de manosa 6fosfato y se transportan mediante vesículas a los lisosomas
muchas son proteínas modificadas por M6P.
Las vesículas generadas porpinocitosis en fase líquida o endocitosis mediada porreceptores pasa por el endosoma
inicial y luego por el endosoma final, los cuales maduran y constituyen lisosomas. El nivel de acidez va a aumento
en el recorrido hasta llegar al punto de acidez máximo en el lisosoma.
Los orgánulos senescentes y los grandes complejos de proteínas desnaturalizadas penetran los lisosomas por
autofagia.
La fagocitosis de microorganismos/fragmentos de matriz/residuos tiene lugar en fagocitos (macrófagos o
neutrófilos), los cuales forman un fagosoma que se fusiona con los lisosoma.
Proteosomas: son complejos de proteasas con múltiples subunidades que degradan las proteínas citosólicas,
incluidaslas desnaturalizadas o mal plegadas, así como otras macromoléculas. Estas generan fragmentos de 6 a 12
aminoácidos que serán reciclados.
3. METABOLISMO CELULAR Y FUNCIÓN MITOCONDRIAL
Las mitocondrias evolucionaron de procariotas ancestrales que fueron engullidas por eucariotas primitivas, lo cual explica
que estas tengan su propio ADN, el cual es circular y constituye el 1% del total del ADN de la célula que codifica un 1% de
proteínas celulares y el 20% de las proteínas implicadas en la fosforilación oxidativa
1.1 Generación de energía
La mayor parte de la energía celular procede del metabolismo mitocondrial. Sus sustratos se oxidan a CO2, transfiriendolos
electrones de alta energía de la molécula original al oxígeno molecular y generando electrones de agua, de baja energía.
La oxidación de varios metabolitos activa las bombas de protones, que transfieren H+ de la matriz al espacio
intermembrana. A medida que los iones H+ fluyen en contra de su gradiente electroquímico, la energía liberada se utiliza
para sintetizar ATP. El transporte de electrones no siempre genera ATP. La termogenina, proteína de la membrana
interna presente a concentraciones elevadas en ciertos tejidos (p. ej., la grasa parda), desacopla el proceso, produciendo
calor en vez de ATP.
1.2 Metabolismo intermediario
La glucosa ademas de dirigirse a formación de ATP en la mitocondria, produce intermediarios se desvían para la
producción de lípidos ácidos nucleicos y proteínas. Por lo tanto según el estado de crecimiento de la célula, el
metabolismo mitocondrial se modula para dar soporte al mantenimiento o al crecimiento celular, lo cual es regulado por
medio de factores de crecimiento, reserva de nutrientes y oxigeno, y de la via de transmisión de señales y sensores que
responden a factores exógenos.
1.3 Muerte celular
Las mitocondrias también regulan el equilibrio entre superviviencia y muerte de la célula mediante dos vías principales:
NECROSIS
APOPTOSIS
PATOLOGIA HUMANA
Page
7
of
10
4. ACTIVACIÓN CELULAR
Las señales extracelulares determinan si la célula vive o muere, o si permanece en reposo o es estimulada para una
función especifica. La perdida de Comunicación celular puede provocar un crecimiento no controlado (cancer) o una
respuesta ineficaz frente a estresores extrinsecos (shock)
1.1 Transmisión de señales celulares
Las células responden a las siguientes señales:
Patógenos y daño de células próximas
Contacto con células próximas
Contacto con la MEC mediado x integrinas
Moléculas secretadas (factores de crecimiento, citocinas hormonas)
Las vías de señalización se clasifican como:
Paracrinas: se ven afectadas las células en proximidad inmediata.
Autocrinas: las moléculas secretadas por una célula afectan a esta misma célula.
Sinápticas: las neuronas secretan neurotransmisores hacia las células diana en las uniones celulares
especializadas.
Endocrinas: el mediador es liberado al torrente circulatorio y actúa sobre las células diana a distancia.
Las moléculas de señalización (ligandos) se unen a sus respectivos receptores e inician una cascada de episodios
intracelulares independientemente del tipo de estimulo extracelular. Los receptores pueden ser:
Receptores intracelulares: son factores de transcripciones activados por ligandos liposolubles que atraviesan
con facilidad las MP.
Receptores de superficie celular: suelen ser proteínas transmembranas con dominios de unión a ligandos
extracelulares. La unión a ligandos puede: abrir canales iónicos, activar una proteína G, activar una enzima
endógena o asociada a tirosina o cinasa o desencadenar un episodio proteolítico o un cambio en la unión a
proteínas y en la estabilidad de las mismas.
5. VÍAS DE TRANSDUCCIÓN DE LA SEÑAL
La unión de ligandos a receptores de superficie media la señalización por inducción de agregación de receptores
(enlaces cruzados en el receptor) o por otros tipos de perturbaciones físicas, lo que provoca cambios bioquímicos
intracelulares, activando en última instancia factores de transducción que penetran en el núcleo para alterar la expresión
génica. Los receptores celulares se clasifican según los mecanismos señaladores que utilizan y las vías bioquímicas
intracelulares que activan:
Receptores asociados a actividad cinasa: Las alteraciones en los receptores inducen actividad proteína cinasa
intrínseca o favorecen la actividad enzimática de cinasas intracelulares reclutadas, originando adición de
residuos fosfato cargados a las moléculas diana. Las tirosina cinasa adhieren fosforilan residuos de tirosina
específicos, mientras que las serina/treonina cinasas lo hacen a residuos serina/treonina determinados y las
lípidos cinasas fosforilan sustratos lipídicos, para cada fosforilación existe una fosfatasa.
a) Receptores tirosina cinasa: son proteína integrales de membrana (ejemplo: receptores de insulina), la
union del ligando activan tirosina cinasa localizados en las colas citoplasmáticas.
b) Varios receptores carecen de actividad catalítica intrínseca (ejemplo: receptores inmunitarios). En estos
casos proteínas intracelulares distintas (tirosina cinasa no receptora) fosforilan residuos específicos del
receptor o de otras proteínas
Receptores acoplados a la proteína G: son polipéptidos que tipicamente atraviesan la mp siete veces, tras unirse
al ligando, el receptor se asocia a una proteína de unión a GTP(proteína G), que contiene GDP. Esto induce su
activación, por intercambio de GDP a GTP, con generación anterógrada de AMPc.
Receptores nucleares: los ligandos liposolubles pueden difundir por el interior de las células, donde
interaccionan con proteína intracelulares para formar un complejo receptor-ligando que se une
directamente al ADN nuclear generando la activación o represión de la transcripción.
PATOLOGIA HUMANA
Page
8
of
10
Otra clase de receptores: importantes para el desarrollo embrionario y la determinación del destino celular
a) Las proteínas receptoras de la familia notch
b) Los ligandos de la proteína wnt
1.2 proteínas, conectores y nodos modulares de transmisión de señales
El fin de las vías de transducción clasicamente se resumen en cambios en la expresión génica y respuesta
biológica deseada, pero resulta en multiples efectos
activación de enzimas o inactivación
Localizacion nuclear o citoplasmática de factores de transcripción
activación de factores de transcripción o inactivación
Polimerización de actina o despolimerización
degradación de proteínas o estabilización
activación de circuitos de retroalimentación inhibidores o estimuladores.
Las proteínas adaptadoras funcionan como conectores moleculares que enlazan físicamente distintas enzimas y
promueven el ensamblaje de complejos, que pueden ser proteínas integrales de membrana o citosólicas. El adaptador
puedo tener unos cuantos dominios específicos que median interacciones entre proteínas. Por analogía con las redes
informáticas, los complejos proteínicos se pueden considerar nodos y los procesos bioquímicos dirigidos o emanados
de estos serian los conectores.
1.3 Factores de transcripción
La mayoría de las vías de transcripción de la señal afectan a las función de celular modulando la transcripción
génica a través de la activación y localización nuclear de factores de transcripción, su cambio de conformación
permite su traslado al núcleo o bien exponen dominios específicos de union al ADN o proteínas
6. FACTORES DE CRECIMIENTO Y RECEPTORES
Su función principal es estimular la actividad de genes necesarios para e crecimiento y la division de células. La
actividad de los factores de crecimiento es mediada por la unión a receptores específicos, que induce la expresión de
genes que:
Favorecen la entrada en el ciclo celular.
Eliminan los bloqueos para la progresión del ciclo celular.
Previenen la apoptosis.
Fomentan la biosíntesis de constituyentes celulares.
Regulan múltiples actividades diferentes del crecimiento, como migración, diferenciación y capacidad
desíntesis.
Los factores de crecimiento están implicados en la proliferación de células en estado de equilibrio, así como después
deuna lesión, a fin de reponer las células dañadas. Numerosos genes de las vías de los factores de crecimiento son
protooncogenes. Cuando la producción de un factor de crecimiento se descontrola y sus vías de señalización se
activande forma constitutiva, se produce una proliferación celular no regulada.
7. INTERACCION CON LA MATRIZ EXTRACELULAR
Las interacciones celulares con la MEC son esenciales en el desarrollo y la cicatrización, así como en el mantenimiento
de la arquitectura tisular normal:
Soporte mecánico: permite la fijación y la migración celulares, y el mantenimiento de la polaridad celular.
Control de la proliferación celular: la MEC enlaza factores de crecimiento que pueden ser
liberados/activadospor proteólisis y también emite señales mediante las integrinas celulares.
Base estructural para la renovación tisular: la integridad de la membrana basal y el estroma de las
célulasparenquimatosas es fundamental para la regeneración organizada de los tejidos.
PATOLOGIA HUMANA
Page
9
of
10
Establecimiento de microambientes tisulares: la membrana basal no es solo un soporte pasivo entre epitelio
ytejido conjuntivo; también es funcional.
La MEC presenta dos formas básicas: matriz intersticial y membrana basal:
Matriz intersticial: sintetizada por cél mesenquimatosas, se sitúan en los espacios intercelulares del tejido
conectivo y entre el epitelio parenquimatoso y las estructuras de soporte subyacentes. Sus principales
componentes son colágenos fibrilares y no fibrilares, junto con fibronectina, elastina, proteoglucanos y
hialuronato.
Membrana basal: se sintetiza a partir del epitelio que la cubre y las células mesenquimatosas a ella
subyacentes.Principales constituyentes: colágeno tipo IV y no fibrilar amorfo y laminina.
1.1 Componentes de la MEC
Los componente pertenecen a tres grupos de proteínas
proteínas estructurales fibrosas: como colágeno y elastina, para la fuerza de tension y elasticidad
Geles hidratados en agua: hilvanaron y proteoglucanos, para resistencia a la comprensión y lubricación
Glucoproteínas adhesivas: conectan los elementos de la MEC entre si y con las células
8. MANTENIMIENTO DE LAS POBLACIONES CELULARES
1.1 Proliferación y ciclo celular
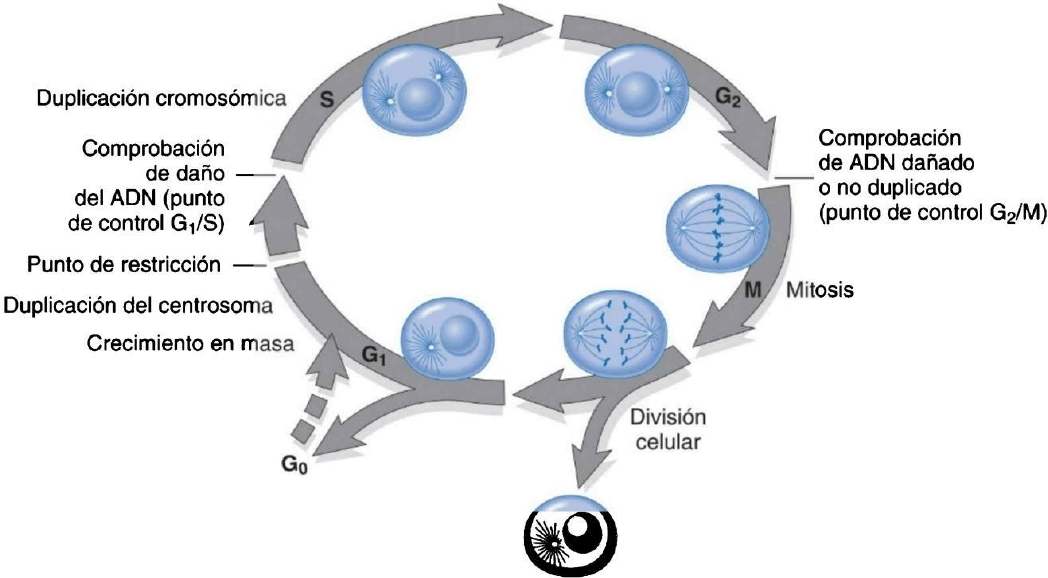
Ciclo celular: secuencia de procesos que culmina en la division celular
Es fundamental para el desarrollo, el mantenimiento de la homostasia tisular en estado de equilibrio y el
remplazo de células muertas o dañadas.
Los elementos clave de la proliferación del ciclo celular son:
1. Replicación precisa del ADN
2. Síntesis coordinada de los demás constituyentes celulares (ej orgánulos).
3. Distribución equitativa del ADN y otros elementos celulares a las cel hijas.
Fases del ciclo celular: G1 (crecimiento presintético); S (Síntesis del ADN); G2 (crecimiento premitosis) y M (mitosis).
Esteciclo esta regulado por activadores e inhibidores y su progresión es dirija por: proteínas ciclinas (cíclicas) y kinasas
dependientes de ciclinas (CDK) las cuales son enzimas que fosforilan.
El aumento transitorio de la síntesis de una ciclina concreta provoca una mayor actividad cinasa de pareja CDK, a medida
PATOLOGIA HUMANA
Page
10
of
10
que la CDK completa su ronda de fosforilación, la ciclina asociada se degrada y la actividad de la CDK disminuye.
Durante el ciclo celular, existen puntos de control que aseguran que las células con defectos genéticos no completen su
replicación. El punto de control G1/S evalúa la integridad del ADN para su replicación y el punto de restricción G”/M
garantiza que la replicación genética haya sido adecuada antes de que se divida la célula. Cuando se detectan
imperfecciones, se activan mecanismos de reparación del ADN. Esta reparación puede no resultar, por lo que la célula
sufre apoptosis, ó pueden entrar en un estado no replicativo (senescencia) sobre todo a través de mecanismos
dependientes de p53.
El refuerzo de los puntos de control del ciclo celular es la misión de los inhibidores de CDK, lo consiguen modulando la
actividad del complejo CDK-Ciclina.
1.2 Células madre
Las células madre se caracterizan por dos importantes propiedades:
La autorrenovación, que permite que las células madre mantengan su número.
La división asimétrica, en la que una célula hija entra en una vía de diferenciación y da lugar a células
maduras,mientras que la otra permanece indiferenciada y conserva su capacidad de autorrenovación.
Variedades de células madre: las cél madre embrionarias, derivadas de la masa celular interna del blastocito, son
totipotenciales (generan cualquier célula del cuerpo) y cél madre tisulares (limitadas en cuanto a diferenciación).

UNIDAD 1 - PATOLOGIA.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.