Gluconeogénesis
Importancia biológica
Determinados tejidos NECESITAN un aporte CONTINUO de glucosa:
Cerebro: depende de glucosa como combustible primario
Eritrocito: utiliza glucosa como único combustible
Consumo glucosa
Cerebro: 120 g/dia
Organismo: 160 g/dia
Reservas de glucosa
Liquidos corporales: 20 g
Glucógeno: 160 g
Las reservas directas de glucosa solo son suficientes para cubrir las
necesidades de un día!!!: períodos más largos de ayuno implican la
necesidad de sistemas alternativos de obtener glucosa
GLUCONEOGENESIS: síntesis de glucosa a partir de precursores que no
sean hidratos de carbono:
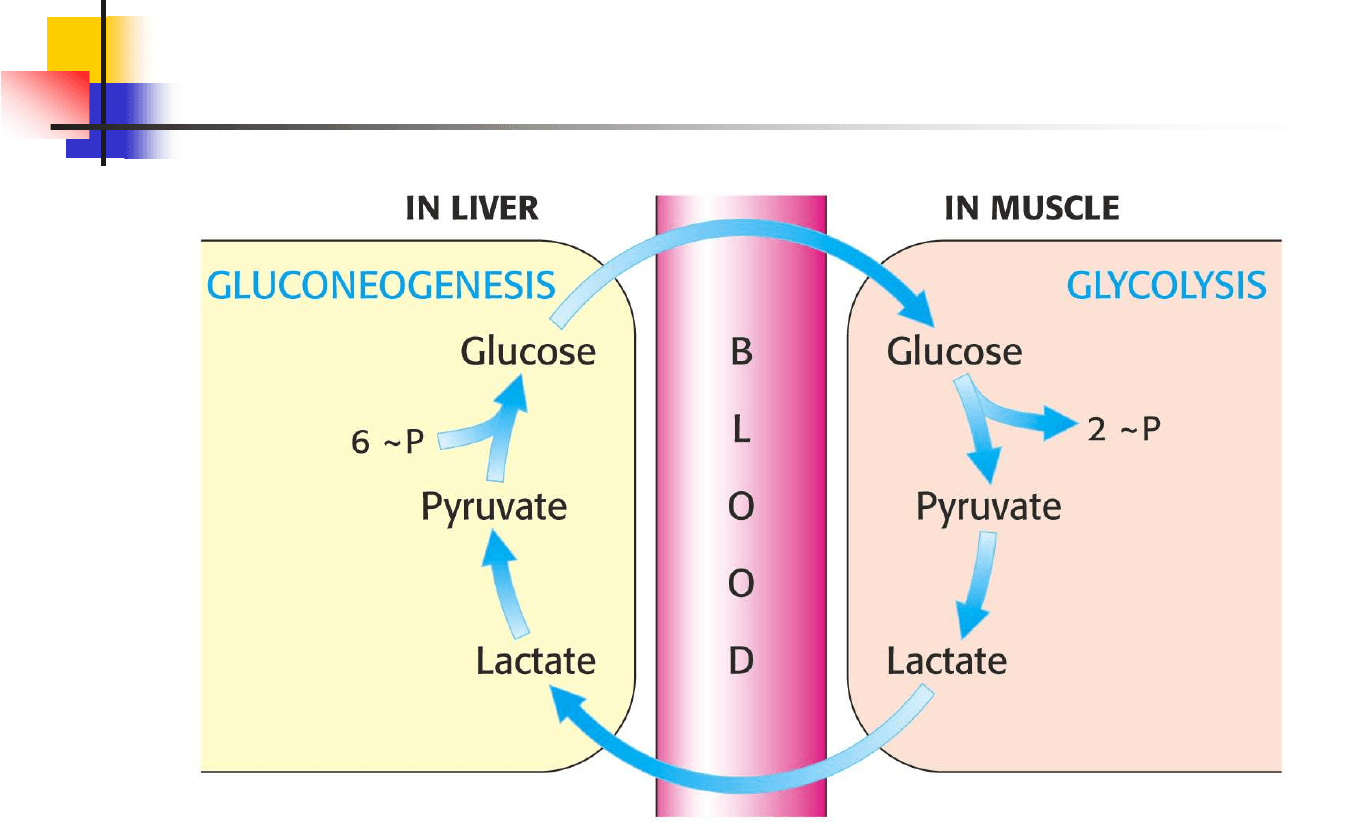
LACTATO: músculo esquelético activo cuando
Glicolisis > fosforilación oxidativa
AMINOACIDOS: degradación de proteínas de la dieta o
proteínas de músculo esquelético.
GLICEROL: hidrólisis triacilglicéridos en células adiposas.
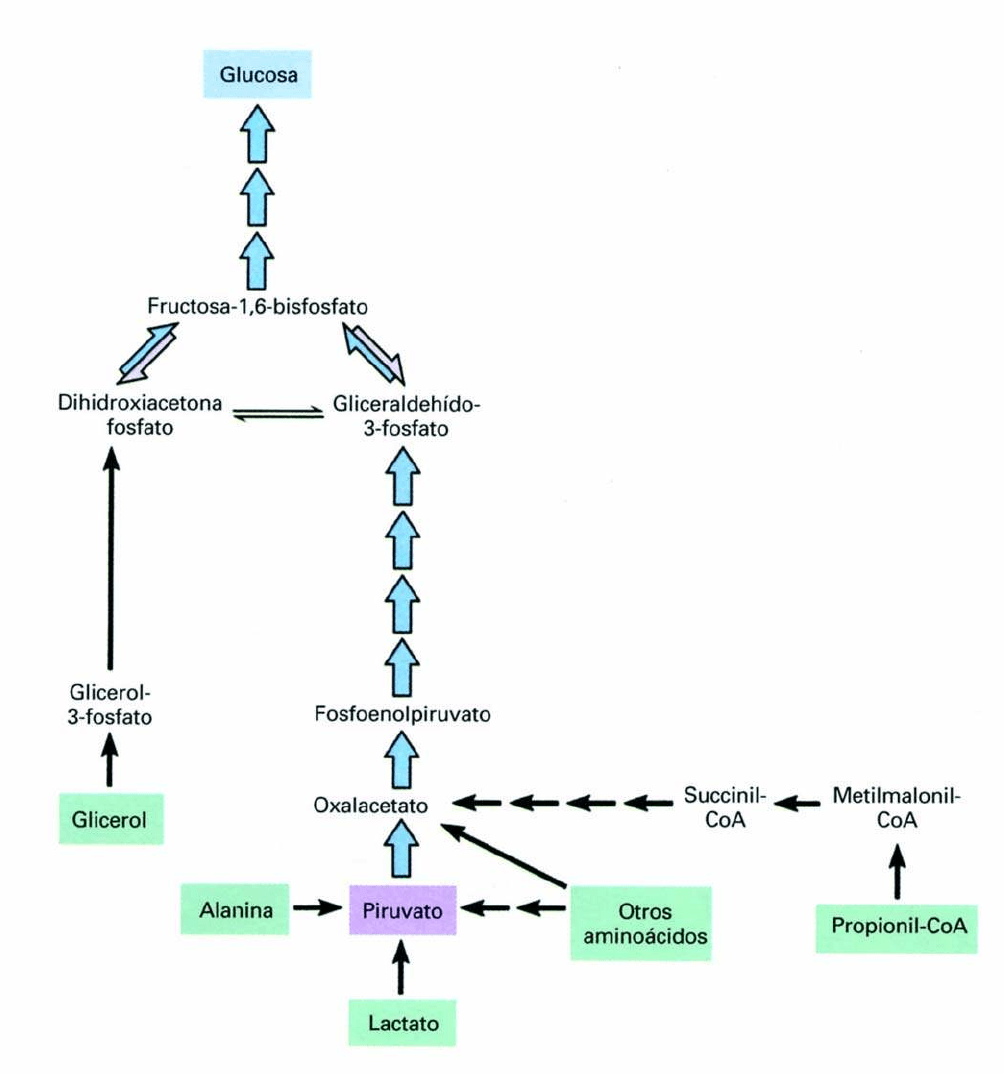
GLUCONEOGENESIS:
síntesis de glucosa a partir de piruvato.
• Los precursores gluconeogénicos se convierten a
piruvato, o bien entran en la ruta por conversión a
oxalacetato o dihidroxiacetona fosfato
• Cualquier metabolito que pueda ser convertido a
piruvato u oxalacetato puede ser un precursor de
glucosa
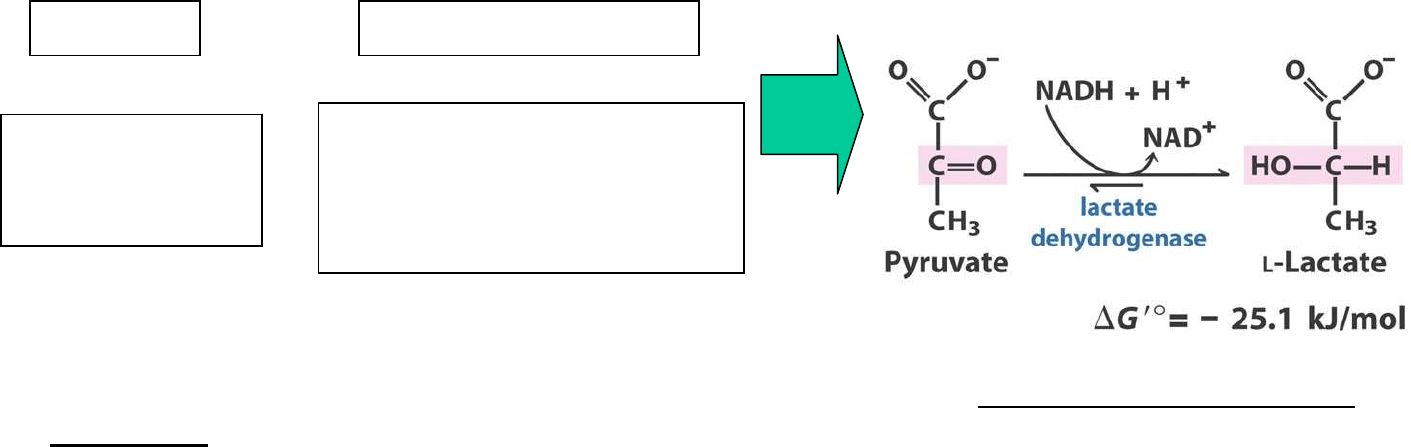
Lactato como precursor gluconeogénico
Durante ejercicio físico vigoroso, cuando se contrae el músculo esquelético:
NADH es regenerado a NAD
+
por
LACTATO DESHIDROGENASA
>
Glicolisis Ciclo del ácido cítrico
Regeneración a NAD+ por
el metabolismo aerobio
(Ciclo del ácido cítrico +
cadena transporte)
Formación de
NADH por la
Glicolisis
>
Lactato como tal queda como punto muerto en el metabolismo: debe convertirse de nuevo
en piruvato para poder ser metabolizado: es reconvertido a piruvato en el hígado
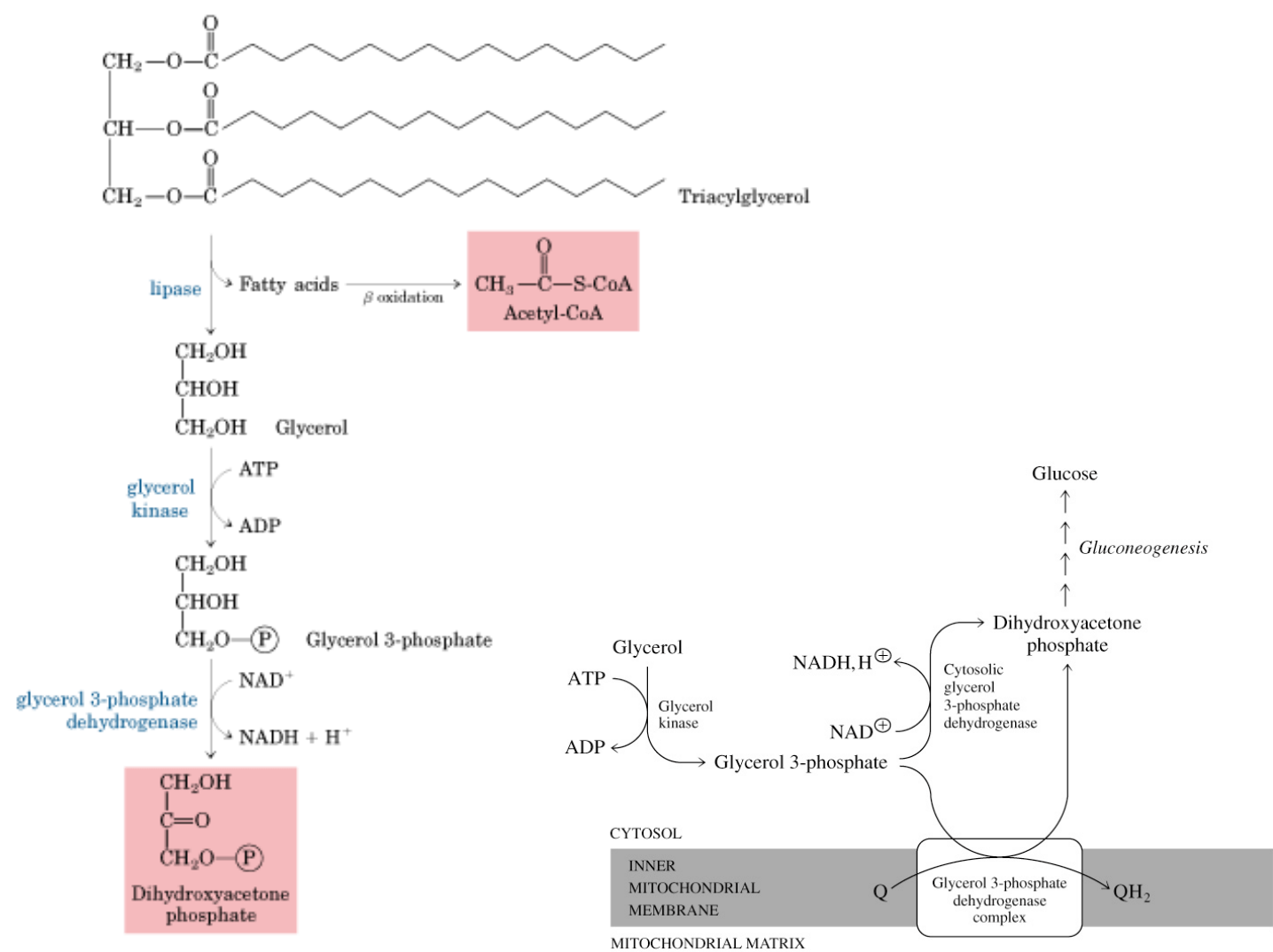
Glicerol como
precursor
gluconeogénico
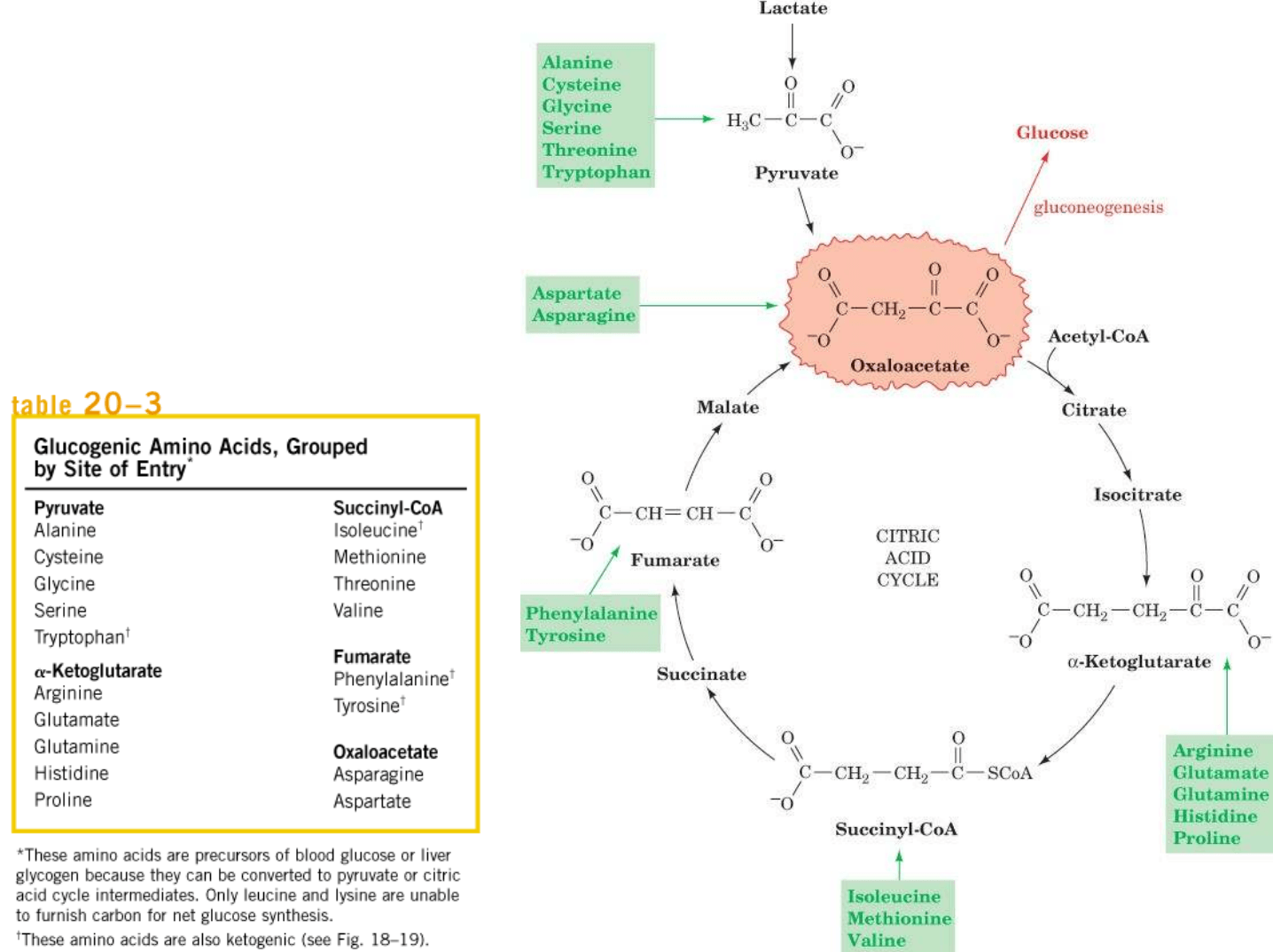
Aminoácidos
precursores
de glucosa
Localización tisular
Hígado (90%) y riñón (10%)
son los órganos donde tiene
lugar principalmente la
gluconeogénesis
En Cerebro, músculo esquelético y
músculo cardíaco tiene lugar muy
poca gluconeogénesis
CEREBRO
MUSCULO ESQUELETICO
MUSCULO CARDIACO
GLUCOSA EN
SANGRE
GLUCONEOGENESIS
HIGADO/RIÑON
La gluconeogénesis en hígado y riñón ayuda a mantener el nivel de
glucosa necesario en sangre para que cerebro y músculos puedan extraer
la suficiente glucosa para atender a sus demandas energéticas
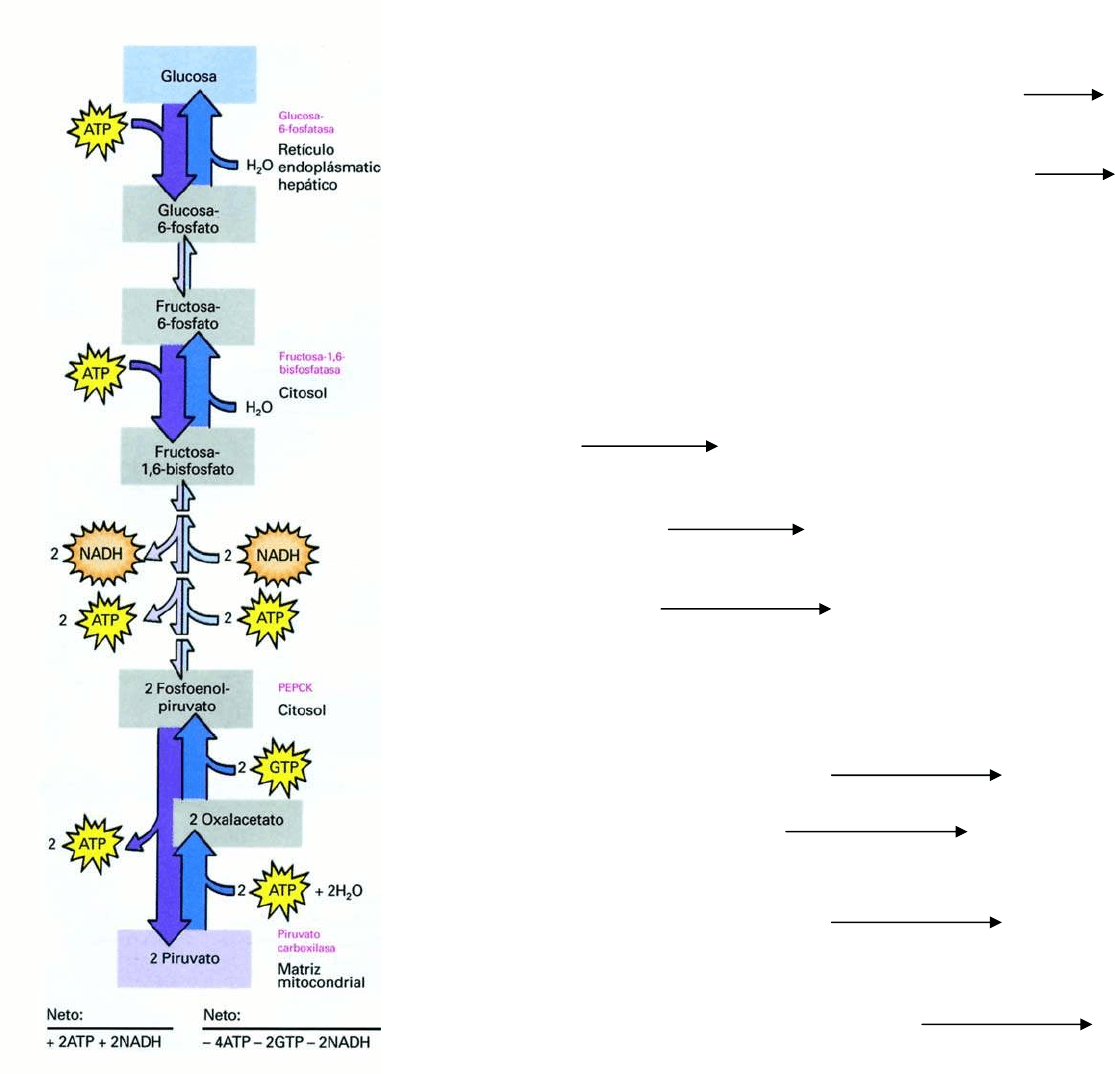
GLICOLISIS: Glucosa Piruvato
GLUCONEOGENESIS: Piruvato Glucosa
Sin embargo, la gluconeogénesis no es el proceso inverso de la
glicolisis
Razon termodinámica: 3 reacciones de la
glicolisis estan muy
desplazadas del equilibrio, practicamente irreversibles
Hexoquinasa
Glucosa + ATP Glucosa-6-fosfato + ADP + Pi + 2H
+
∆G= -8 Kcal/mol
fosfofructoquinasa
Fructosa-6-fosfato + ATP Fructosa-1,6-bifosfato + ADP ∆G= -5,3 Kcal/mol
Piruvato quinasa
Fosfoenolpiruvato + ADP Piruvato + ATP ∆G= -4 Kcal/mol
En la gluconeogénesis estas reacciones son sustituidas por
reacciones nuevas:
Formación de Fosfoenolpiruvato:
Piruvato carboxilasa
Piruvato + CO
2
+ ATP + H
2
O Oxalacetato + ADP + Pi + 2H
+
Fosdoenolpiruvato carboxiquinasa
Oxalacetato + GTP Fosfoenolpiruvato + GDP + CO
2
Formación de Fructosa-6-fosfato:
Fructosa 1,6 Bifosfatasa
Fructosa-1,6-Bifosfato + H
2
O Fructosa-6-fosfato + Pi
Formación de Glucosa:
Glucosa 6-fosfatasa
Glucosa-6-fosfato + H
2
O Glucosa + Pi
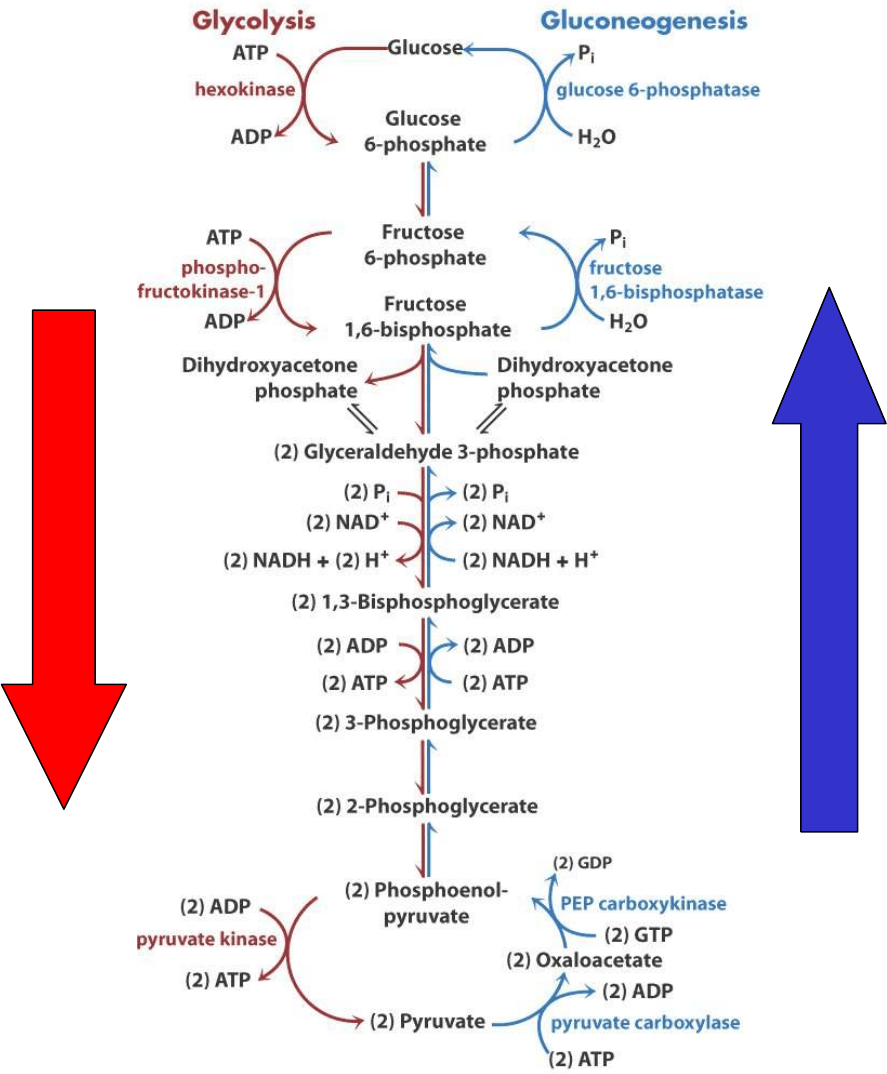
Glicolisis
Gluconeogénesis
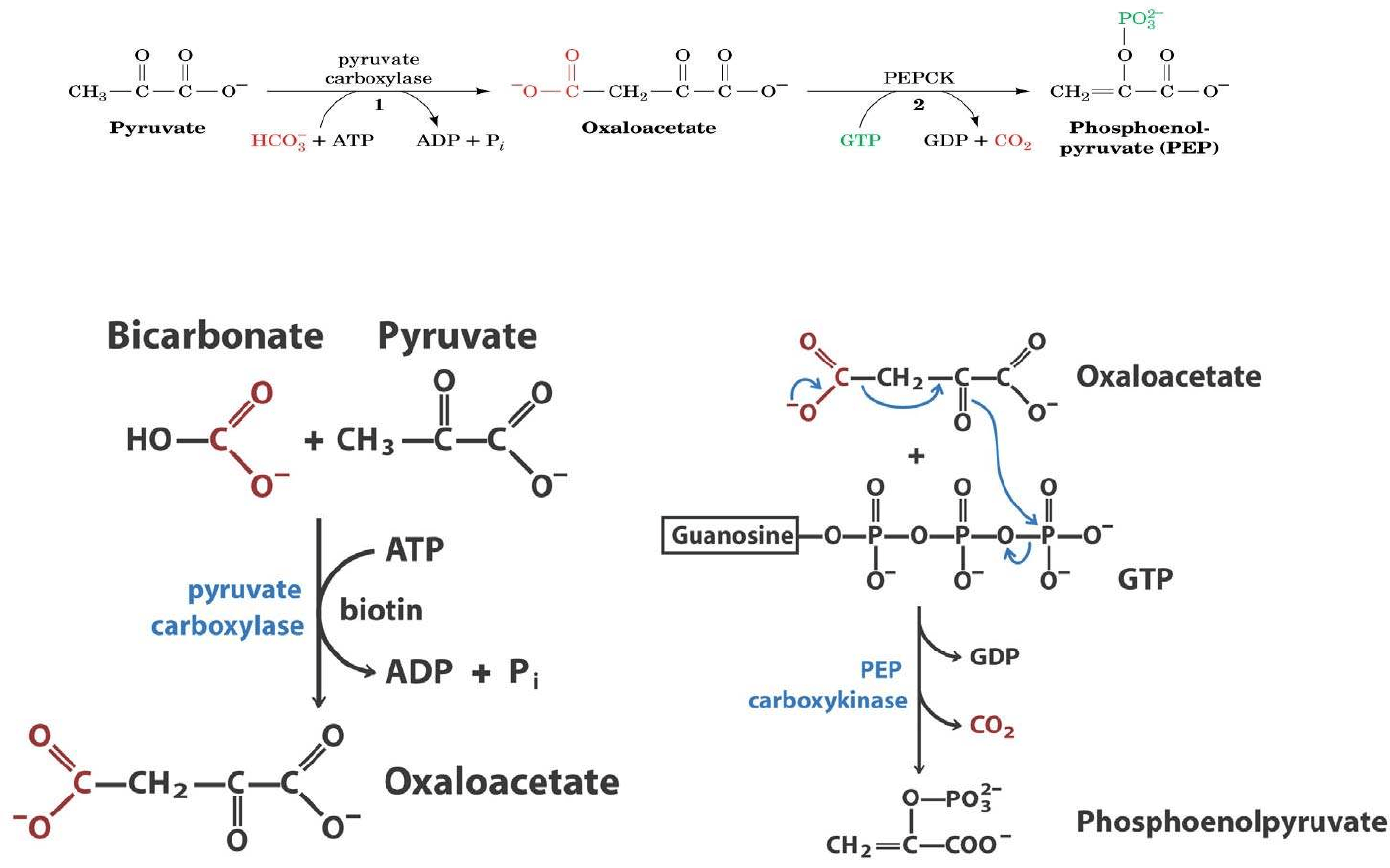
Conversión de Piruvato en fosfoenolpiruvato
Se realiza en dos pasos:
a) Carboxilación del piruvato,
consumiendo ATP
b) Descarboxilación y fosforilación del
oxalacetato, consumiendo GTP
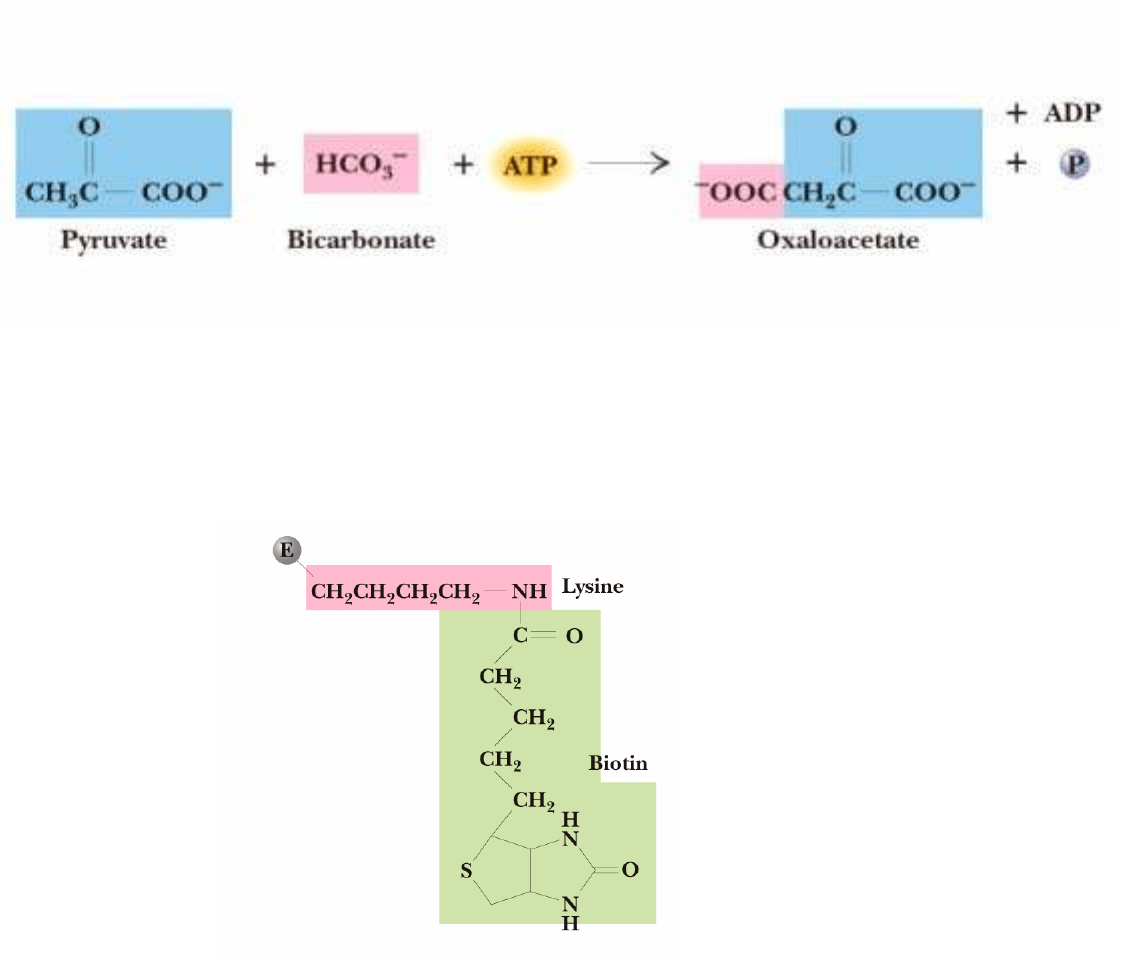
Se realiza en la matriz mitocondrial.
a) Carboxilación del piruvato
Catalizado por PIRUVATO CARBOXILASA:
Estructura:
Region N-terminal 300-350 aa : Dominio de captación de ATP
Región C-terminal: dominio de unión de Biotina
BIOTINA:
transportador de CO
2
activado.
Unida al enzima por una
cadena larga y flexible.
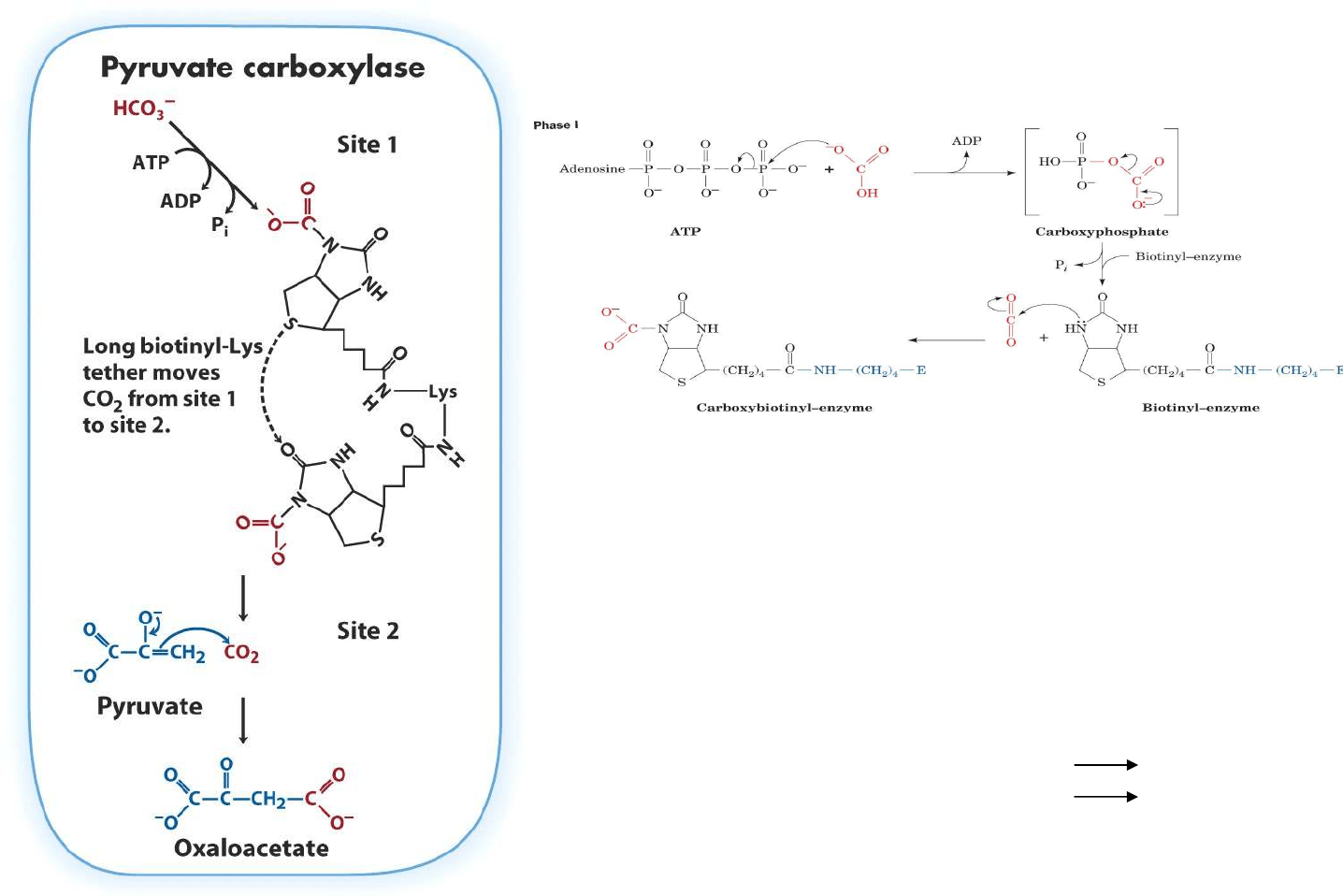
Mecanismo de la Piruvato carboxilasa
Mecanismo de tres etapas:
1. Activación del CO
2
2. Unión del CO
2
activado a la biotina
3. Paso del CO
2
desde biotina al piruvato. El brazo
unido a biotina permite el transporte del CO
2
entre los dos centros activos del enzima.
La presencia de Acetil CoA: control fisiológico
-carga energética alta: oxalacetato glucosa
-carga energética baja: oxalacetato ciclo del ácido
citrico
La etapa de carboxilación de biotina depende de la unión
previa de Acetil CoA: ACTIVACION ALOSTERICA
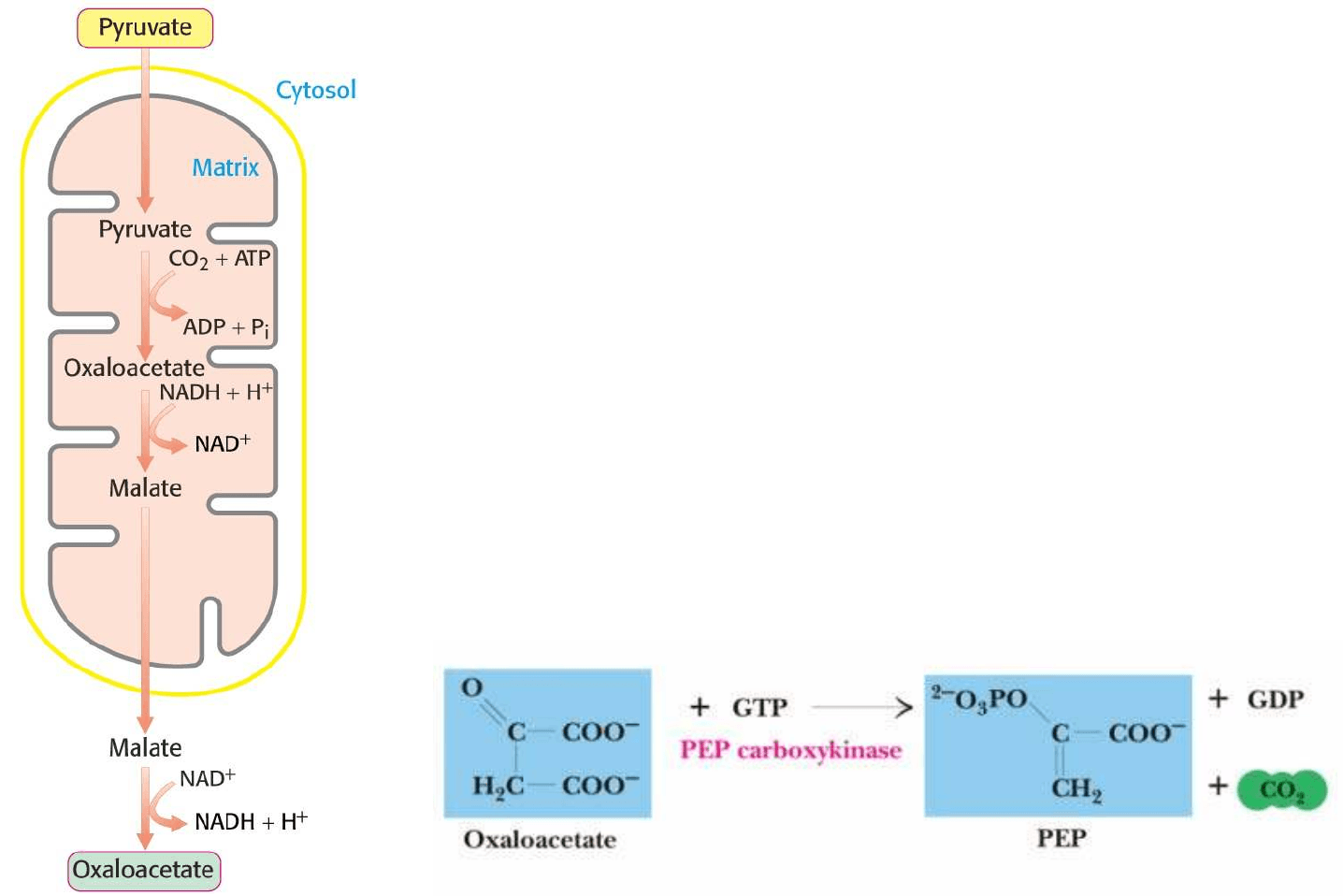
Piruvato carboxilasa es un enzima mitocondrial, mientras que
el resto de enzimas de la gluconogénesis son citosólicos: Se
debe transportar el oxalacetato producido fuera de la
mitocondria:
1. Oxalacetato es reducido a malato por una malato
deshidrogenasa mitocondrial ligada a NADH
2. Malato es transportado al citosol por el sistema
lanzadera malato-aspartato
3. Una vez en el citosol, el malato es reoxidado a
oxalacetato por una malato deshidrogenasa citosólica
ligada a NAD
+
Oxalacetato es descarboxilado y fosforilado simultaneamente por
FOSFOENOLPIRUVATO CARBOXIQUINASA (PEP
carboxiquinasa). La hidrólisis del GTP y liberación de CO
2
desplazan al reacción hacia la formación de PEP.
B) Transporte de Oxalacetato al citosol y conversión a
fosfoenolpiruvato
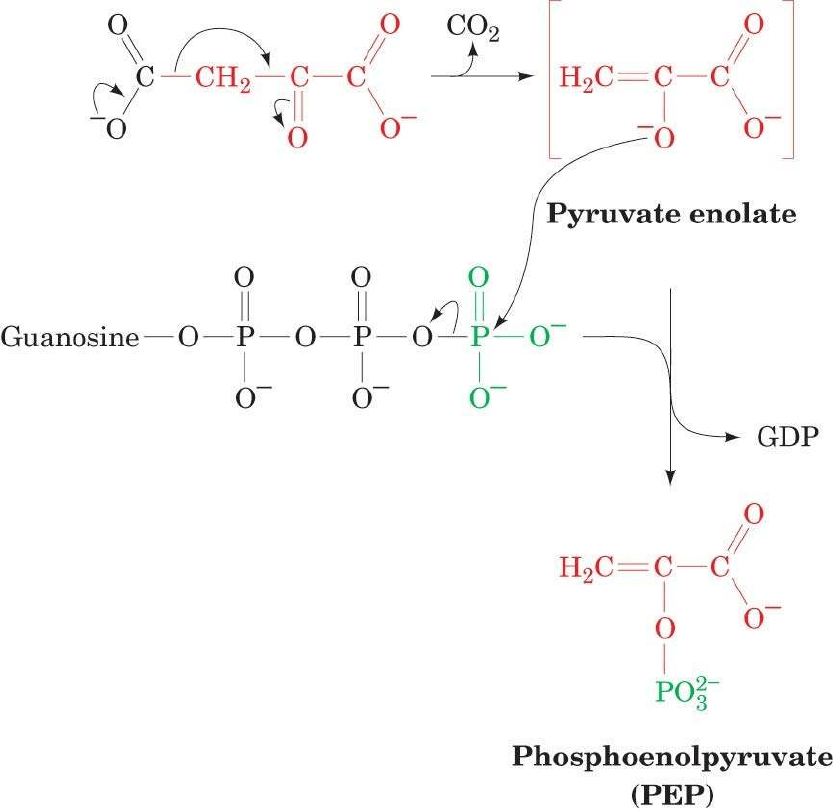
Mecanismo de la
FOSFOENOLPIRUVATO CARBOXIQUINASA
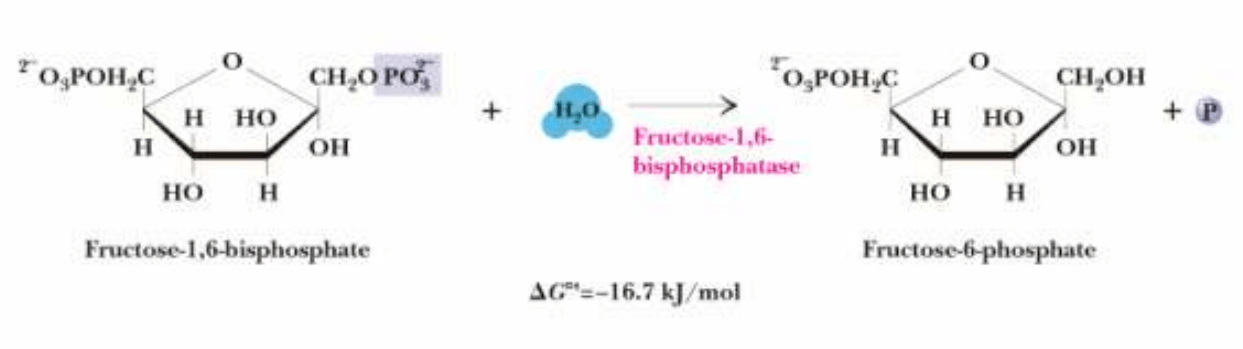
Conversión de Fructosa-1,6-bifosfato en Fructosa-6-fosfato
Una vez formado, el Fosfoenolpiruvato es metabolizado por los enzimas de la
glicolisis pero en sentido inverso (reacciones en equilibrio).
El siguiente paso irreversible es la hidrólisis de Fructosa-1,6-bifosfato en Fructosa-
6-fosfato y Pi:
Catalizado por FRUCTOSA-1,6-BIFOSFATASA, enzima alostérico.
Requiere Mg
2+
.
Inhibido por AMP, fructosa 2,6-bifosfato
activado por ATP, citrato
Este documento contiene más páginas...
Descargar Completo
Tema17 - Gluconeogenesis.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.