1
TEJIDO CARTILAGINOSO
El cartílago es un tejido avascular que consiste en condrocitos y una matriz extracelular extensa especializada. Más del
95% del volumen del cartílago corresponde a la MEC, elemento funcional de este tejido. Los condrocitos son escasos pero
indispensables para la producción y mantenimiento de la MEC.
La MEC del cartílago es sólida y firme pero también algo maleable. Como no existe una red vascular en el cartílago, la
supervivencia de los condrocitos depende de la composición de la MEC. La gran proporción de glucosaminoglucanos (GAG)
con respecto a las fibras de colágeno tipo II en la matriz del cartílago permite la difusión de sustancias desde los vasos
sanguíneos del tejido conjuntivo circundante a los condrocitos dispersos en la matriz, manteniendo la viabilidad del tejido.
Existen interacciones entre dos clases de moléculas estructurales: la red de fibrillas de colágeno resistentes a la tensión y
la gran cantidad de proteoglucanos muy hidratados (débiles contra fuerzas de cizallamiento, pero buenos para soportar
peso).
Según las características de su matriz, se distinguen 3 tipos de cartílago que difieren en cuanto a su aspecto y sus
propiedades mecánicas:
1. CARTÍLAGO HIALINO: matriz que contiene fibras de colágeno tipo II, GAG, proteoglucanos y glucoproteínas
multiadhesivas.
2. CARTÍLAGO ELÁSTICO: fibras elásticas y laminillas elásticas además de material de matriz de cartílago hialino.
3. CARTÍLAGO FIBROSO (FIBROCARTÍLAGO): abundantes fibras de colágeno tipo I, además de material de matriz
del cartílago hialino.
CARTÍLAGO HIALINO
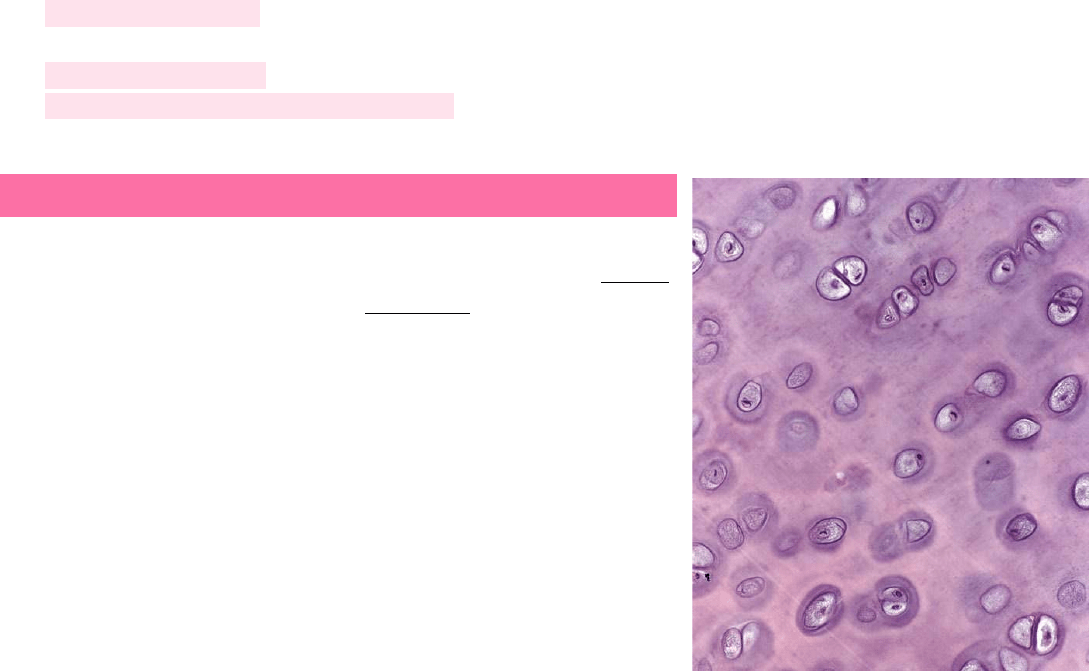
La matriz de cartílago hialino tiene aspecto vidrioso en el estado vivo. En
toda la extensión de la matriz cartilaginosa hay espacios llamados lagunas,
dentro de las cuales se encuentran los condrocitos. Este tipo de cartílago
provee una superficie de baja fricción, participa en la lubricación de las
articulaciones sinoviales y distribuye las fuerzas aplicadas al hueso
subyacente. Si bien su capacidad de reparación es limitada, en
circunstancias normales, no exhibe indicios de desgaste abrasivo durante
toda la vida. Una excepción es el cartílago articular, el cual, en muchas
personas, se degrada con la edad.
Las macromoléculas de la matriz del cartílago hialino consisten en colágeno
(con predominio de fibrillas de colágeno tipo II y otras moléculas de
colágeno específicas), aglomeraciones de proteoglucanos que contienen
GAG y glucoproteínas multiadhesivas (proteínas no colágenas).
2
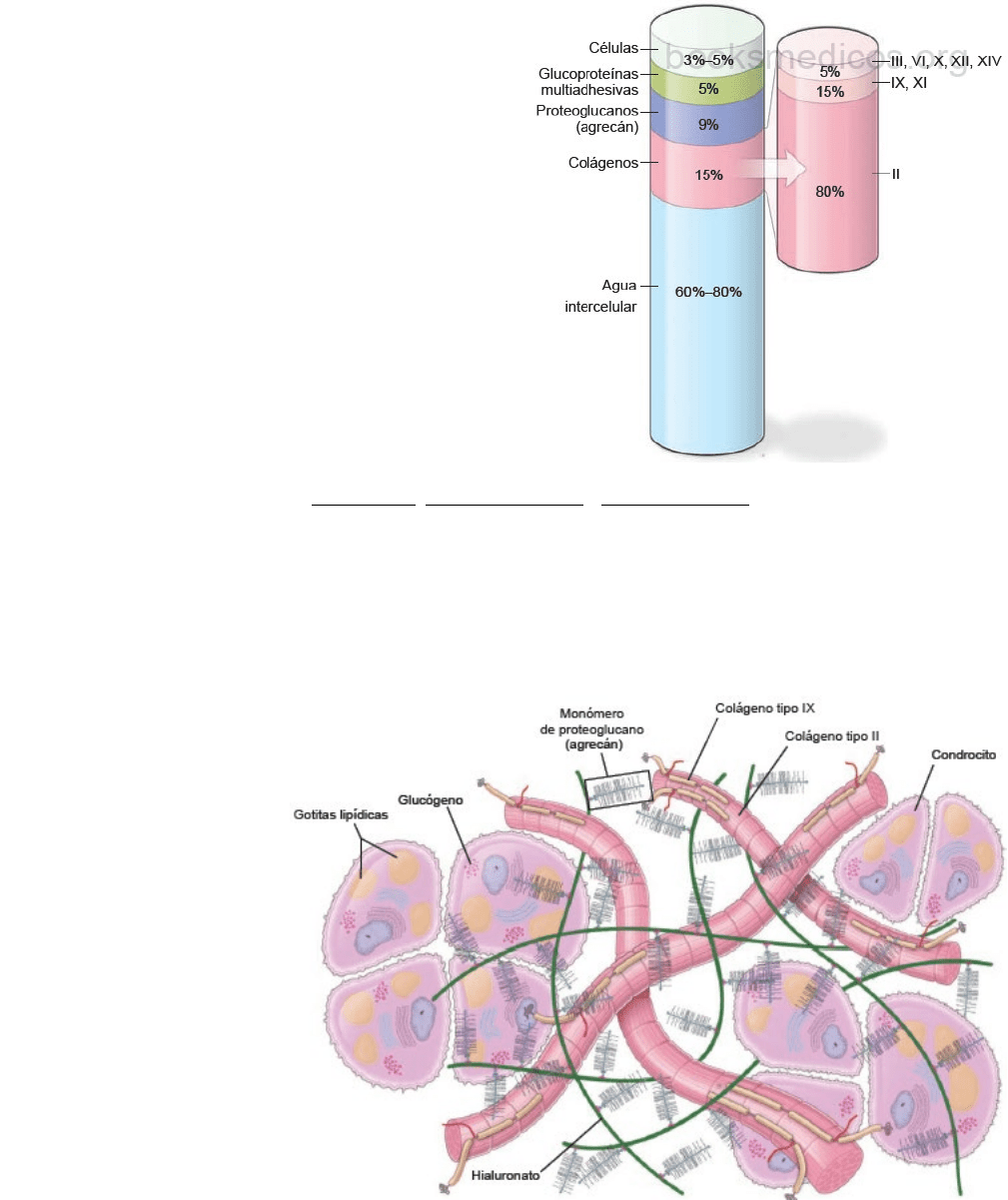
En la matriz del cartílago hialino se distinguen tres clases de
moléculas:
1. Moléculas de colágeno: proteína principal de la matriz. 4
tipos de colágeno participan en la formación de una red
de fibrillas cortas y delgadas. La mayor parte de éstas está
constituida por colágeno tipo II. El colágeno tipo XI regula
el tamaño de las fibrillas y el colágeno tipo X organiza las
fibrillas en una red hexagonal, decisiva para su función
mecánica eficaz. El colágeno tipo VI predomina en la
periferia de los condrocitos, en donde contribuye a la
adhesión de éstas al armazón matricial. A los tipos II, VI,
IX, X y XI se les llama moléculas de colágenos específicos
del cartílago, ya que se encuentran en cantidades
importantes sólo en la matriz cartilaginosa.
2. Proteoglucanos: la sustancia fundamental de cartílago
hialino contiene tres tipos de GAG: hialuronato, condroitín sulfato y queratán sulfato. Como en la matriz del tejido
conjuntivo laxo, el condroitín sulfato y el queratán sulfato de la matriz se unen a una proteína central para formar
un monómero de proteoglucanos. El más importante de estos monómeros del cartílago hialino es el agrecán, que
tiene gran afinidad por las moléculas de agua. Cada molécula de hialuronato se asocia con una gran cantidad de
moléculas de agrecán para formar aglomeraciones de proteoglucanos (agrecano-hialuronato), unidas a las fibrillas
de colágeno de la matriz por interacciones electrostáticas y glucoproteínas multiadhesivas. Esta es la causa de las
propiedades biomecánicas del cartílago hialino. La matriz cartilaginosa también contiene otros proteoglucanos
(decorina, biglicán y
fibromodulina), que no
forman agregados, pero se
unen a moléculas y
contribuyen a estabilizar la
matriz.
3. Glucoproteínas
multiadhesivas
(glucoproteínas no ligadas a
proteoglucanos): influyen
sobre las interacciones entre
los condrocitos y las moléculas
de la matriz. Las
glucoproteínas son
marcadores de recambio y de
la degeneración del cartílago.
La matriz del cartílago hialino está muy hidratada para permitir la elasticidad y difusión de metabolitos pequeños. Del 60
al 80% del peso del cartílago hialino corresponde a agua intercelular. Gran parte de esta agua está fuertemente unida a
las aglomeraciones de agrecano-hialuronato, produciendo alta presión osmótica y son responsables de la elasticidad del
cartílago. La red de fibrillas de colágeno tipo II es responsable de la forma del cartílago, de su resistencia a la tensión y
también le provee un armazón para resistir la presión osmótica de las moléculas de agrecán.
3
Las compresiones aplicadas al cartílago, como ocurre en las articulaciones sinoviales, crean señales mecánicas, eléctricas
y químicas que contribuyen a dirigir la actividad sintética de los condrocitos. No obstante, a medida que el organismo
envejece, la composición de la matriz cambia y los condrocitos pierden la capacidad de responder a estos estímulos.
CONDROCITOS
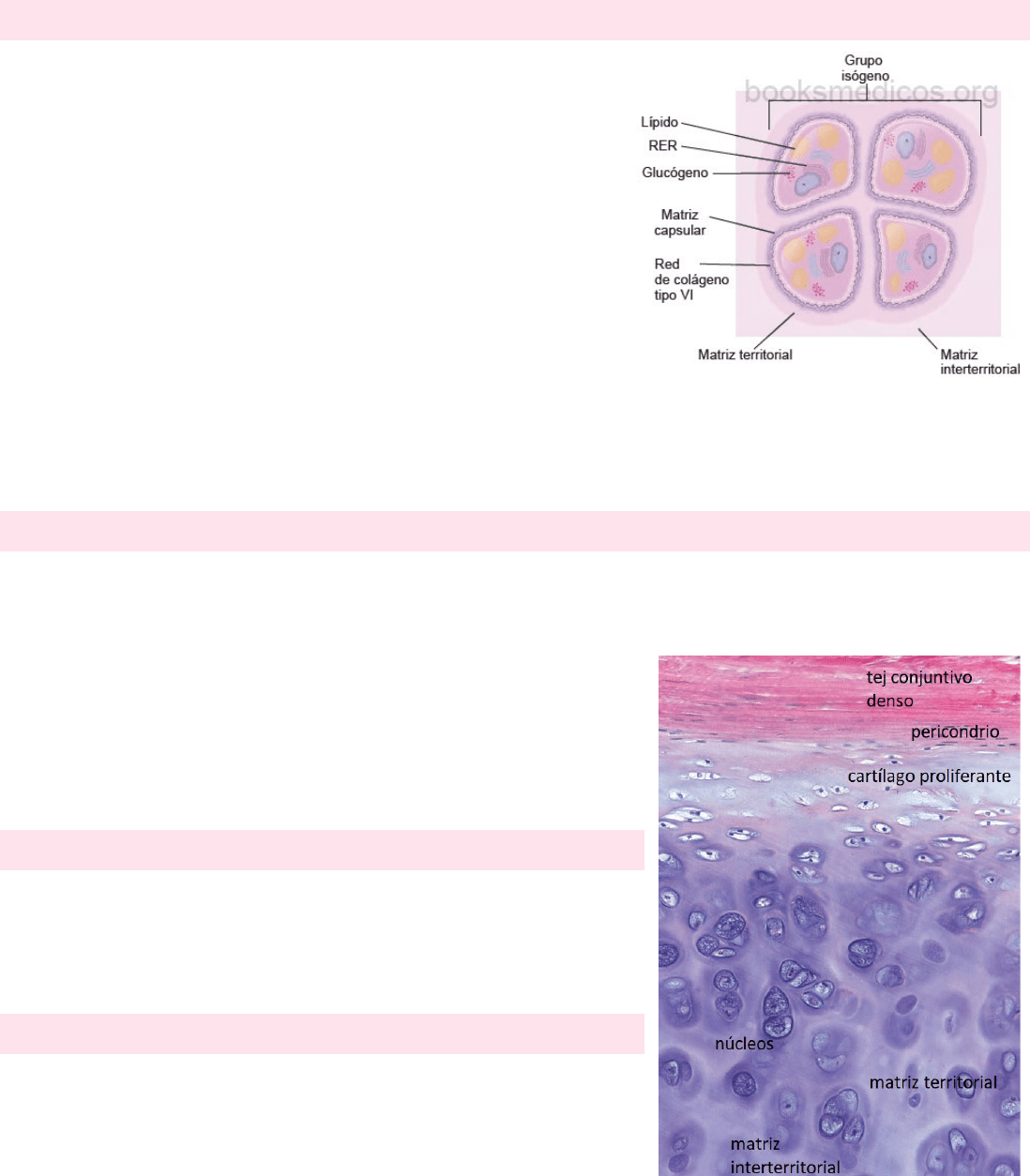
Son células especializadas que producen y mantienen la MEC. En el
cartílago hialino se distribuyen solos o en cúmulos llamados grupos
isógenos. Cuando los condrocitos están presentes en estos grupos, son
células que acaban de dividirse. A medida que van sintetizando la matriz
que los rodea, los condrocitos recién divididos se dispersan. También
secretan metaloproteinasas, enzimas que degradan la matriz
cartilaginosa para permitir que las células se expandan y se reubiquen
dentro del grupo isógeno en crecimiento.
Los condrocitos que están activos en la producción de la matriz tienen
regiones de basofilia citoplasmática (se tiñen fácilmente), que indican
síntesis proteica, y regiones claras, que corresponden al aparato de Golgi
grande. Los condrocitos no sólo secretan el colágeno de la matriz, sino también todos los GAG y proteoglucanos.
En células menos activas, el aparato de Golgi es más pequeño, las regiones citoplasmáticas son claras, cuando se ven,
indican sitios donde se han extraído lípidos o depósitos de glucógeno.
EL CARTÍLAGO HIALINO PROVEE UN MOLDE PARA EL ESQUELETO EN DESARROLLO DEL FETO
En las etapas iniciales del desarrollo fetal, el cartílago hialino es el precursor del tejido óseo que se origina por el proceso
de osificación endocondral. Al principio, la mayoría de lo que serán los huesos largos no son más que moldes de cartílago
que se asemejan a la forma del hueso maduro.
Durante el proceso de desarrollo, cuando gran parte del cartílago es
reemplazada por hueso, un resto de tejido cartilaginoso en la parte proximal
y distal del hueso sirve como sitio de crecimiento: es la placa epifisiaria de
crecimiento. Este cartílago permanece funcional siempre y cuando el hueso
crezca en longitud.
PERICONDRIO
El pericondrio es un tejido conjuntivo denso irregular compuesto por células
que no pueden distinguirse de los fibroblastos. Durante el crecimiento
activo, el pericondrio aparece dividido en una capa interna celular, que da
origen a células cartilaginosas nuevas y una capa externa fibrosa.
CARTÍLAGO ARTICULAR
El cartílago hialino que cubre las superficies articulares de las diartrosis se
denomina cartílago articular. La diferencia entre ambos tipos de cartílago,
es que el articular carece de pericondrio.
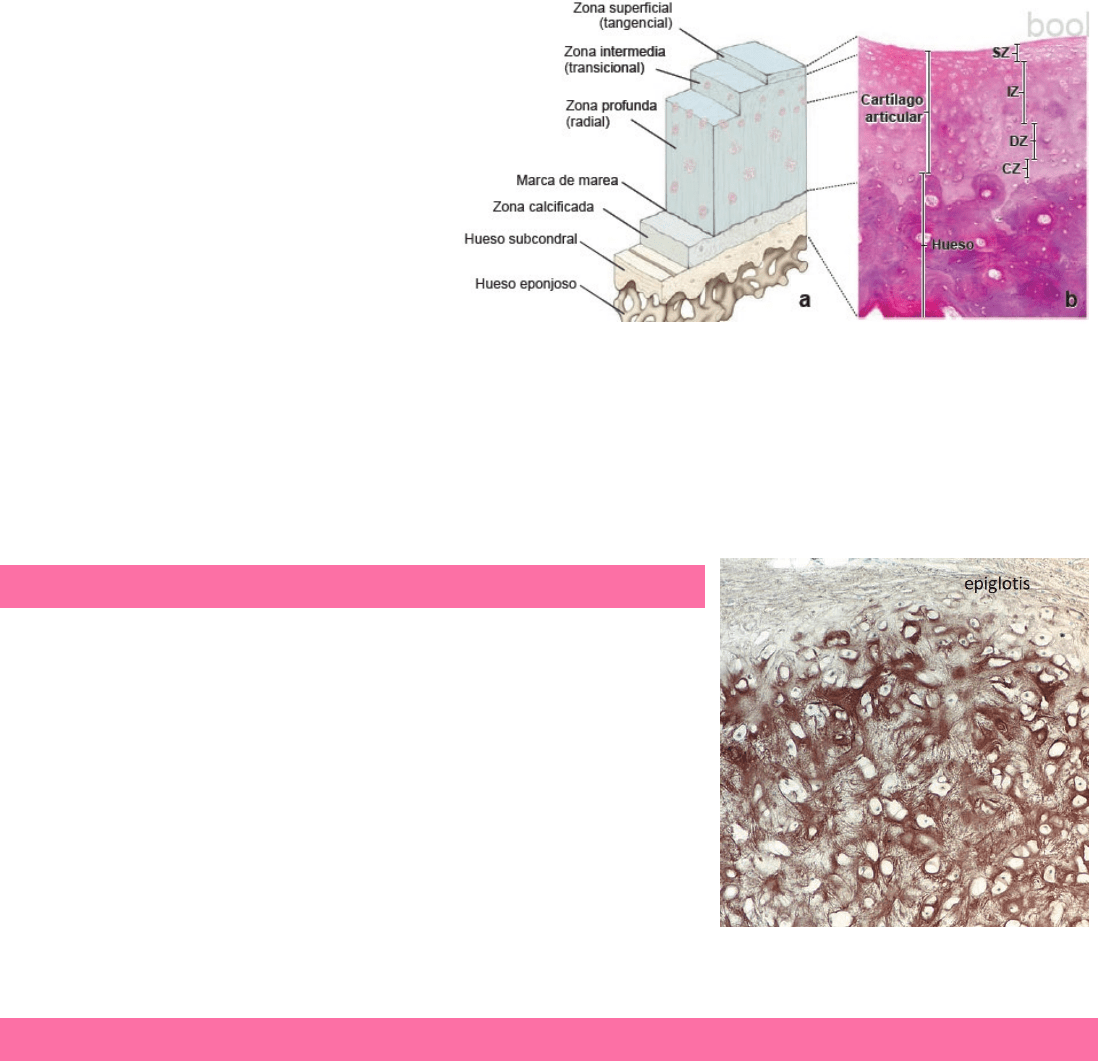
4
En los adultos, el cartílago articular se divide en 4
zonas:
1. Zona superficial (tangencial): región
resistente a la compresión, cercana a la
superficie articular. Contiene abundantes
condrocitos alargados y aplanados rodeados
por una condensación de fibrillas de
colágeno tipo II.
2. Zona intermedia (transicional): debajo de la
zona superficial y contiene condrocitos
redondos distribuidos al azar dentro de la matriz y fibrillas de colágeno.
3. Zona profunda (radial): tiene condrocitos redondeados pequeños dispuestos en columnas cortas perpendiculares a la
superficie libre del cartílago, y fibrillas de colágeno entre las columnas.
4. Zona calcificada: caracterizada por una matriz calcificada con la presencia de condrocitos pequeños. Separada de la
zona profunda por la línea de marea. Por arriba de esta línea, la proliferación de los condrocitos dentro de las lagunas
del cartílago provee las células nuevas para el crecimiento intersticial. En la renovación del cartílago articular, los
condrocitos migran desde esta región hacia la superficie de la articulación.
CARTÍLAGO ELÁSTICO
Además de contener los componentes normales de la matriz del cartílago
hialino, la matriz del cartílago elástico también contiene una densa red de
fibras elásticas ramificadas y anastomosadas y láminas interconectadas de
material elástico. Este material imparte propiedades elásticas el cartílago,
además de distensibilidad y maleabilidad, características del cartílago hialino.
Este tipo de cartílago se encuentra en el pabellón auricular, en las paredes del
CAE, en la tuba de Eustaquio y en la epiglotis. El cartílago de todos estos sitios
está rodeado por un pericondrio similar al que se encuentra alrededor de la
mayoría de los cartílagos hialinos. La matriz de este cartílago no se calcifica
durante el proceso de envejecimiento.
CARTÍLAGO FIBROSO O FIBROCARTÍLAGO
Es una combinación de tejido conjuntivo denso modelado y cartílago hialino. Los condrocitos están dispersos entre las
fibrillas de colágenos solos, en hileras y formando grupos isógenos. Este tipo de cartílago tiene mucho menos material de
matriz asociado con los condrocitos.
En los cortes de fibrocartílago, es normal ver una población de células con núcleos redondos y una pequeña cantidad de
matriz circundante. Estos núcleos pertenecen a los condrocitos. Dentro de las regiones fibrosas se ven núcleos aplanados
o alargados, estos son los núcleos de los fibroblastos.

5
El cartílago fibroso es típico de los discos intervertebrales, la sínfisis
pubiana, los discos articulares de las articulaciones esternoclavicular y
temporomandibular, los meniscos de la rodilla, el complejo
fibrocartilaginoso triangular de la muñeca y ciertos sitios donde los
tendones se insertan en los huesos. La presencia del cartílago en estos sitios
indica que el tejido debe soportar fuerzas de compresión y distensión,
actuando como amortiguador.
Las células del cartílago fibroso sintetizan una gran variedad de moléculas
de EC no sólo durante su etapa de desarrollo sino también durante su etapa
madura bien diferenciada.
Esto permite que el fibrocartílago responda a los cambios en el medio
externo (fuerzas mecánicas, modificaciones nutricionales y
concentraciones variables de hormonas y factores de crecimiento). La MEC
del cartílago fibroso contiene cantidades importantes de colágeno tipo I
(característico de la matriz del tejido conjuntivo) y de colágeno tipo II
(característico del cartílago hialino). La proporción entre los colágenos de los tipos I y II en el fibrocartílago varía con la
edad.
CONDROGÉNESIS Y CRECIMIENTO DEL CARTÍLAGO
La condrogénesis, proceso de desarrollo del cartílago, comienza con la aglomeración de células mesenquimatosas
condroprogenitoras para formar una masa densa de células redondeadas.
El sitio de formación del cartílago hialino se reconoce inicialmente por una aglomeración de células mesenquimatosas
conocida como nódulo condrogénico. La expresión del factor de transcripción SOX-9 desencadena la diferenciación de
estas células en condroblastos, que secretan matriz cartilaginosa. Los condroblastos se van separando progresivamente
conforme depositan matriz a su alrededor. Una vez que el material de matriz los ha rodeado por completo reciben el
nombre de condrocitos. El tejido mesenquimatoso que hay justo alrededor del nódulo condrogénico da origen al
pericondrio.
La condrogénesis está regulada por muchas moléculas, entre las cuales hay ligandos extracelulares, receptores nucleares,
factores de transcripción, moléculas adhesivas y proteínas de la matriz. También, el crecimiento y desarrollo del esqueleto
de cartílago son afectados por las fuerzas biomecánicas, que regulan la forma, la regeneración y el envejecimiento del
cartílago, y modifican las interacciones célula-matriz extracelular dentro de este tejido.
Con el inicio de la secreción de la matriz, el crecimiento del cartílago continúa por una combinación de dos procesos:
• Crecimiento por aposición: proceso en el cual se forma cartílago nuevo sobre un cartílago preexistente.
• Crecimiento intersticial: proceso de formación de cartílago nuevo en el interior de un cartílago preexistente.
Las células cartilaginosas nuevas producidas durante el crecimiento por aposición derivan de la capa interna del
pericondrio circundante. Las células se asemejan a los fibroblastos en cuanto a forma y función y producen el componente
de colágeno del pericondrio (tipo I). Sin embargo, cuando el crecimiento se inicia, las células sufren un proceso de
diferenciación guiado por la expresión del factor de transcripción SOX-9. Las evaginaciones citoplasmáticas desaparecen,

6
el núcleo se redondea y el citoplasma aumenta de tamaño. Estos cambios determinan la conversión de la célula en un
condroblasto. Los condroblastos sintetizan la matriz cartilaginosa, incluida la secreción de colágeno tipo II. La nueva
matriz aumenta la masa de cartílago, mientras se van produciendo nuevos fibroblastos para mantener la población celular
del pericondrio.
Las células cartilaginosas nuevas producidas durante el crecimiento intersticial surgen de la división de los condrocitos
dentro de sus lagunas. Al principio, las células hijas de los condrocitos en división ocupan la misma laguna. A medida que
se secreta una matriz nueva, se forma una división entre ambas células hijas; en este momento, cada célula ocupa su
propia laguna. Conforme se secreta una cantidad mayor de matriz, las células se van separando cada vez más. En
consecuencia, el crecimiento global del cartílago es el producto de la secreción intersticial del nuevo material de matriz
por los condrocitos y de la aposición de matriz secretada por condroblastos recién diferenciados.
REPARACIÓN DEL CARTÍLAGO HIALINO
El cartílago puede tolerar bastante la acción de las fuerzas intensas y repetidas. Sin embargo, cuando se lesiona, el cartílago
manifiesta una llamativa incapacidad para sanar, aún en las lesiones más leves. Esta falta de respuesta a la lesión se
atribuye a la avascularidad del cartílago, la inmovilidad de los condrocitos y la capacidad limitada de los condrocitos
maduros para proliferar. Es posible cierto grado de reparación, pero sólo si el defecto comprende el pericondrio. En estas
lesiones, la reparación es el resultado de la actividad de las células progenitoras pluripotenciales ubicadas en el
pericondrio. La reparación comprende sobre todo la producción de tejido conjuntivo denso.
A nivel molecular, la reparación de cartílago es un equilibrio tentativo entre el depósito de colágeno tipo I, en forma de
tejido cicatrizal, y la restauración por la expresión de los colágenos específicos del cartílago. Sin embargo, en los adultos,
es común que se formen vasos sanguíneos nuevos en el sitio de la herida en proceso de curación, lo que estimula el
desarrollo de tejido óseo en vez de una reparación del cartílago.
El cartílago hialino es propenso a calcificarse, procedo en el que cristales de fosfato de calcio se depositan en la matriz
cartilaginosa. La matriz del cartílago hialino sufre normalmente calcificación en 3 situaciones bien definidas:
1. La porción del cartílago articular que está en contacto con el tejido óseo en los huesos en crecimiento y en el
adulto, pero no la porción superficial, está calcificada.
2. La calcificación siempre ocurre en el cartílago que está por ser reemplazado por tejido óseo (osificación
endocondral) durante el período de crecimiento de una persona.
3. El cartílago hialino en el adulto se calcifica con el tiempo como parte del proceso de envejecimiento.
Los condrocitos normalmente obtienen todas sus sustancias nutritivas y eliminan los desechos por difusión de materiales
a través de la matriz. Cuando la matriz se calcifica mucho, se impide la difusión y los condrocitos sufren tumefacción y
mueren.
7
TEJIDO ÓSEO
Es una forma especializada de tejido conjuntivo que, al igual que otros tejidos conjuntivos, se compone de células y MEC.
Lo que distingue el tejido óseo de otros tejidos conjuntivos es la mineralización de su matriz (fosfato de calcio en forma
de cristales de hidroxiapatita), que produce un tejido muy duro capaz de proporcionar sostén y protección.
Gracias a su contenido mineral, este tejido también sirve como sitio de almacenamiento de calcio y fosfato. Tanto el calcio
como el fosfato se pueden movilizar de la matriz ósea y ser captados por la sangre según sea necesario para mantener la
homeostasis.
El principal componente estructural de la matriz ósea es el colágeno tipo I y, en menor medida, el colágeno tipo V.
También se han encontrado de tipo III, XI y XIII). Todas las moléculas de colágeno constituyen alrededor del 90% del peso
total de las proteínas de la matriz ósea.
La matriz también contiene otras proteínas no colágenas que componen la sustancia fundamental del tejido óseo. Ésta
constituye sólo un 10% del peso total de proteínas de la matriz ósea, sin embargo, es esencial para el desarrollo,
crecimiento, remodelado y reparación ósea. Los 4 grupos principales de proteínas no colágenas que se encuentran en la
matriz ósea son:
o Macromoléculas de proteoglucanos: contienen una proteína central con cantidades diversas de cadenas laterales
de GAG (hialuronato, condroitín sulfato y querantán sulfato) unidos en forma covalente. Contribuyen a que el
hueso ofrezca resistencia a la compresión. También son responsables de la fijación de los factores de crecimiento
y puede inhibir la mineralización.
o Glucoproteínas multiadhesivas: intervienen en la adhesión de las células óseas y las fibras colágenas a la sustancia
fundamental mineralizada. Las más importantes son la osteonectina (adhesivo entre el colágeno y los cristales de
hidroxiapatita), la podoplanina (producida por los osteocitos en respuesta al estrés mecánico), la proteína de la
matriz de dentina (mineralización de la matriz ósea) y las sialoproteínas, como la osteopontina (media la adhesión
de las células a la matriz ósea e inicia la formación de fosfato de calcio durante la mineralización).
o Proteínas dependientes de vitamina K osteoespecíficas: incluida la osteocalcina (captura el calcio de la circulación
y atrae y estimula los osteoclastos en el remodelado óseo), la proteína S (que contribuye a la eliminación de las
células que sufren apoptosis) y la proteína Gla de la matriz (participa en el desarrollo de las calcificaciones
vasculares.
o Factores de crecimiento y citosinas: pequeñas proteínas reguladoras, como el factor de crecimiento similar a la
insulina (IGF), el factor de necrosis tumoral α (TNF α), el factor de crecimiento transformante β (TGF-β), los
factores de crecimiento derivados de plaquetas (PDFG), las proteínas morfogénicas óseas (BMP), la esclerostina
(antagonista de BMP) y las interleucinas (IL-1, IL-6). Los más importantes son las BMP ya que inducen la
diferenciación de células mesenquimatosas en osteoblastos, formadoras del hueso. La BMP-7 (conocida como
proteína osteogénica-1) se utiliza clínicamente para inducir el crecimiento óseo tras la cirugía de defectos óseos
mayores, fusiones de columna o la implantación de materiales de injerto.
En la matriz ósea hay espacios llamados lagunas, cada una de las cuales contiene un osteocito. Éste extiende una gran
cantidad de evaginaciones en pequeños túneles llamados canalículos. Los canalículos atraviesan la matriz mineralizada,
conectando lagunas contiguas y permitiendo el contacto entre las evaginaciones de los osteocitos contiguos, formándose

8
una red continua de canalículos y lagunas con células y sus evaginaciones en todo el tejido mineralizado. Las evaginaciones
de los osteocitos están comunicadas mediante uniones de hendidura.
Además de los osteocitos, en el tejido hay otros 4 tipos celulares:
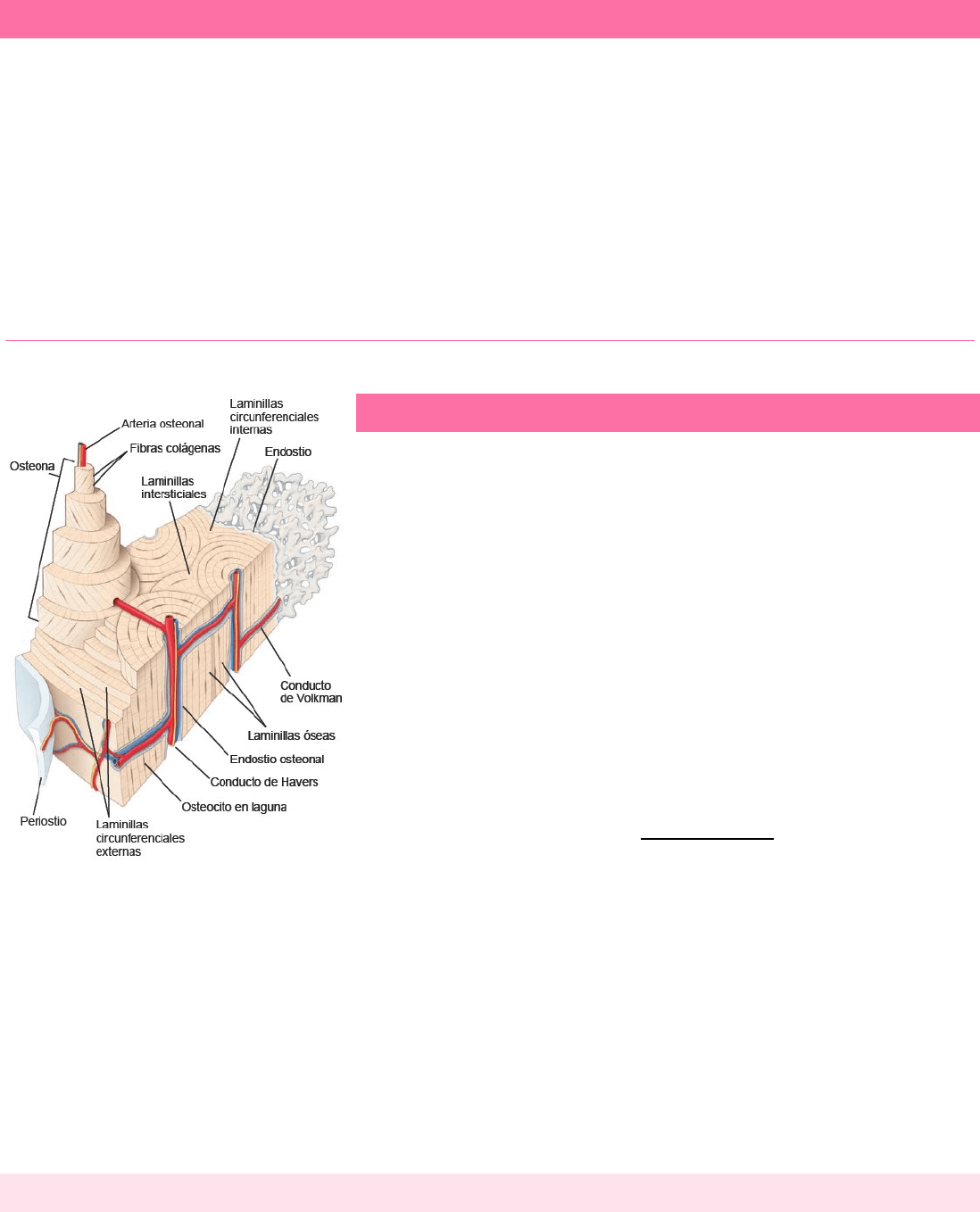
1. Células osteoprogenitoras: derivadas de las células madre mesenquimales. Dan origen a los osteoblastos.
2. Osteoblastos: secretan la MEC del hueso. Una vez que la célula queda rodeada por la matriz secretada, pasa a
llamarse osteocito.
3. Células de revestimiento óseo: permanecen en la superficie ósea cuando no hay crecimiento activo. Derivan de
aquellos osteoblastos que quedan después del cese del depósito óseo.
4. Osteoclastos: células de resorción ósea presentes en las superficies óseas donde el hueso se está eliminando o
remodelando, o donde el hueso ha sido dañado.
Tanto las células osteoprogenitoras como los osteoblastos son precursores del desarrollo de los osteocitos.
ESTRUCTURA GENERAL DE LOS HUESOS
En general, el hueso está compuesto por tejido óseo y otros tejidos conjuntivos, incluidos el tejido hematopoyético,
adiposo, vasos sanguíneos y nervios. Si el hueso forma parte de una articulación sinovial, entonces hay cartílago hialino.
Si se corta un hueso, pueden reconocerse dos organizaciones estructurales distintas de tejido óseo. Una capa densa,
compacta, forma la superficie ósea externa (hueso compacto); una malla de aspecto esponjoso compuesta por cordones,
forma el interior del hueso (hueso esponjoso). Los espacios dentro de la malla están comunicados y, en un hueso vivo,
contienen la médula y los vasos sanguíneos.
Los huesos largos tienen un cuerpo (diáfisis) y dos extremos expandidos llamados epífisis. La superficie articular de la
epífisis está cubierta de cartílago hialino. Una gran cavidad ocupada por médula ósea, llamada cavidad medular, forma la
parte interna del hueso. En las diáfisis, casi todo el espesor del tejido óseo es compacto. En las epífisis el hueso esponjoso
es abundante y el compacto apenas forma una delgada cubierta externa.
Los huesos cortos poseen una corteza de tejido óseo compacto y en su interior hay tejido óseo esponjoso y espacios
medulares. Sus superficies articulares están cubiertas por cartílago hialino. El resto de la superficie externa del hueso está
cubierto por una cápsula de tejido conjuntivo fibroso, el periostio.
SUPERFICIE EXTERNA DE LOS HUESOS
Los huesos están revestidos por periostios excepto en las regiones donde se articulan con otro hueso, donde la superficie
articular está cubierta por cartílago. El periostio se compone de una capa fibrosa externa y de una capa interna que
contiene células osteoprogenitoras. Si no se está formando tejido óseo en la superficie del hueso, la capa fibrosa es el
componente principal y la capa interna no está bien definida.
En general, las fibras colágenas del periostio son paralelas a la superficie del hueso y forman una cápsula. La naturaleza
del periostio es diferente en los sitios donde los ligamentos y los tendones se unen al hueso. Las fibras colágenas de estas
estructuras se extienden de manera oblicua o en ángulos rectos al eje más largo del hueso, y se continúan con las fibras
colágenas de la matriz extracelular. Estas fibras se denominan fibras perforadoras o de Sharpey.
Se extienden hacia las
laminillas intersticiales y circunferenciales externas, pero por lo general no entran en las osteonas.
9
CAVIDADES ÓSEAS
El tejido de revestimiento, tanto del hueso compacto que delimita la cavidad medular como el de los cordones del hueso
esponjoso, se conoce como endostio. Éste suele ser sólo una capa de células de espesor y consiste en células
osteoprogenitoras que pueden diferenciarse en células secretoras de matriz ósea, los osteoblastos, y células de
revestimiento óseo.
La médula ósea roja se compone de células sanguíneas en diferentes etapas de diferenciación y una red de células
reticulares y fibras que funcionan como un armazón de sostén para el desarrollo de células y vasos. Conforme el individuo
crece, la cantidad de médula roja no aumenta en proporción con el crecimiento óseo. En etapas posteriores del
crecimiento y en los adultos, cuando la producción de células sanguíneas disminuye, la cavidad medular es ocupada en su
mayor parte por tejido adiposo, pasándose a llamar médula ósea amarilla.
TIPOS DE TEJIDO ÓSEO
HUESO MADURO O LAMINILLAR
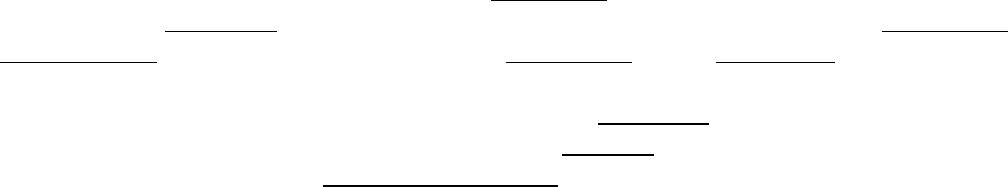
Compuesto, en gran parte, por unidades cilíndricas llamadas osteonas o
sistemas de Havers. Las osteonas consisten en laminillas concéntricas de
matriz ósea alrededor de un conducto central, el conducto osteonal (de
Havers), que contiene el suministro vascular y nervioso de la osteona. Los
canalículos que contienen las evaginaciones de los osteocitos se disponen
generalmente siguiendo un patrón radial con respecto al conducto. El sistema
de canalículos que se abre al conducto osteonal también sirve para el
intercambio de sustancias entre los osteocitos y los vasos sanguíneos. Entre
las osteonas hay restos de laminillas concéntricas antiguas llamadas
laminillas intersticiales.
Las fibras colágenas de cada una de las laminillas concéntricas de una osteona
son paralelas entre sí en cualquier laminilla dada, pero orientadas en una
dirección diferente a la que adoptan las fibras de las laminillas contiguas. Esta
disposición le confiere un aspecto madera terciada a la superficie del corte
del hueso laminillar y le da gran resistencia a la osteona.
El hueso laminillar también se encuentra en otros sitios fuera de la osteona. Las laminillas circunferenciales
siguen la
totalidad de las circunferencias interna y externa de la diáfisis de un hueso largo, y se ven parecidas a los anillos de
crecimiento de un árbol. Los conductos perforantes (de Volkmann) son túneles en el hueso laminillar a través de los cuales
pasan vasos sanguíneos y nervios desde las superficies del periostio y endostio para alcanzar el conducto osteonal (de
Havers); también conectan los conductos de Havers entre sí. Suelen extenderse más o menos perpendiculares al eje
longitudinal de las osteonas y del hueso. Los conductos de Volkmann no están rodeados por laminillas concéntricas, una
característica clave en su identificación histológica.
El hueso esponjoso maduro es similar en estructura al hueso compacto maduro, excepto que el tejido se distribuye
formando cordones. La matriz del hueso es laminillar.
IRRIGACIÓN SANGUÍNEA DE LA DIÁFISIS
10
El foramen nutricio es un orificio en el hueso a través del cual pasan los vasos sanguíneos para alcanzar la médula ósea.
La mayoría de ellos se encuentran en la diáfisis y las epífisis. Las arterias metafisarias complementan la irrigación sanguínea
del hueso. El drenaje venoso se produce por medio de venas que abandonan el hueso a través del foramen nutricio o a
través del tejido óseo de la diáfisis y luego corren por el periostio.
Los conductos de Volkmann proveen la vía de entrada principal para los vasos que atraviesan el tejido óseo compacto. Los
vasos más pequeños se introducen en los conductos de Havers. Una menor irrigación proviene de los vasos del periostio,
que suelen irrigar sólo la porción más externa del tejido óseo compacto. El tejido óseo carece de vasos linfáticos; sólo el
periostio posee drenaje linfático.
HUESO INMADURO, ENTRETEJIDO O FASCICULADO
Es el tejido óseo que se forma primero en el esqueleto de un feto en desarrollo. Difiere del hueso maduro en:
o El hueso inmaduro no exhibe aspecto laminillar organizado. Por la disposición de sus fibras colágenas
(entrelazada), esta variedad ósea se denomina no laminillar.
o El hueso inmaduro contiene una cantidad algo mayor de células por unidad de volumen que el hueso maduro.
o Las células del hueso inmaduro tienen la tendencia a distribuirse de forma aleatoria, mientras que en el hueso
maduro las células se orientan con su eje mayor paralelo a las laminillas.
o La matriz del hueso inmaduro posee más sustancia fundamental que la del hueso inmaduro.
CÉLULAS DEL TEJIDO ÓSEO
CÉLULAS OSTEOPROGENITORAS
La osteogénesis es el proceso de
formación del hueso nuevo, lo cual es
esencial para una función ósea normal. Se
requiere una población de células
osteoprogenitoras renovables (células
precursoras de osteoblastos) que son
sensibles a los estímulos moleculares que
las transforman en célula formadoras de tejido óseo.
Las células osteoprogenitoras derivan de células madre mesenquimatosas de la médula ósea que tienen el potencial de
diferenciarse en diferentes tipos celulares, como fibroblastos, osteoblastos, adipocitos, condrocitos y células musculares.
El factor clave que desencadena la diferenciación de estas células es un factor de transcripción llamado factor fijador
central alfa 1 (CBFA1) o factor de transcripción 2 relacionado con runt (RUNX2). Esta proteína impulsa la expresión de
genes que son característicos del fenotipo del osteoblasto. Los IGF-1 e IGF-2 estimulan la proliferación de las células
osteoprogenitoras y la diferenciación de los osteoblastos.
Las células osteoprogenitoras se encuentran en las superficies externa e interna de los huesos y también pueden residir
en el sistema microvascular que irriga el tejido óseo. Desde el punto de vista morfológico, comprenden las células del
periostio que forman la capa más interna del periostio y las células del endostio que revisten las cavidades medulares, los
conductos osteonales (de Havers) y los conductos perforantes.

11
OSTEOBLASTOS
Es una célula secretora versátil que conserva la capacidad de dividirse. Secreta tanto el colágeno tipo I como las proteínas
de la matriz ósea, que constituyen la matriz no mineralizada inicial, llamada osteoide. Las proteínas de la matriz ósea
producidas por el osteoblasto incluyen proteínas fijadoras de calcio como la osteocalcina y la osteonectina, glucoproteínas
multiadhesivas como las sialoproteínas óseas (BSP 1 y BSP 2), la trombospondina, proteoglucanos y sus agregados y la
fosfatasa alcalina (ALP). Las concentraciones de ALP y osteocalcina en circulación sanguínea se utilizan en clínica como
marcadores de actividad de los osteoblastos.
El osteoblasto también es responsable de la calcificación de la matriz ósea. El proceso de calcificación es iniciado por el
osteoblasto mediante la secreción hacia la matriz de pequeñas vesículas matriciales. Las vesículas contienen gran cantidad
de ALP y se secretan activamente sólo durante en el período en el que la célula produce matriz ósea. Los osteoblastos se
reconocen por su forma cuboide o poliédrica y su distribución monoestratifcada en la superficie donde se está formando
tejido óseo. En contraste con los osteoblastos secretores que se ven donde hay depósito activo de la matriz, los
osteoblastos inactivos son células aplanadas que revisten la superficie ósea. Los osteoblastos responden a estímulos
mecánicos para mediar los cambios en el crecimiento óseo y el remodelado de los huesos. Sólo entre el 10 y 20% de los
osteoblastos se convierten en osteocitos, otros se transforman en células inactivas y se convierten en células de
revestimiento óseo.
El citoplasma del osteoblasto se caracteriza por una gran cantidad de RER y ribosomas libres, lo que concuerda con su
función en la síntesis de colágeno y proteoglucanos para la MEC.
OSTEOCITOS
Una vez que el osteoblasto está totalmente rodeado
por el osteoide o matriz ósea, pasa a llamarse
osteocito. El osteoblasto produce una gran cantidad
de MEC, disminuye su tamaño y la cantidad de
orgánulos y desarrolla largos procesos celulares que se
irradian de su cuerpo celular. Después de la
mineralización de la matriz ósea, cada osteocito ocupa
un espacio o laguna. Los osteocitos extienden sus
evaginaciones citoplasmáticas a través de los
canalículos de la matriz.
Se comunican mediante estas evaginaciones con los
osteocitos vecinos y las células de revestimiento óseo a través de las uniones de hendidura formada por la familia de
conexinas que se expresan en el tejido óseo (uniones célula-célula). Los osteocitos contienen hemiconductos que
proporcionan comunicación entre las células y la MEC (uniones célula-matriz). También pueden comunicarse de forma
indirecta con osteoblastos, células endoteliales del sistema vascular de la médula ósea, los pericitos de los vasos
sanguíneos y con otras células distantes a través de la expresión de diversas moléculas de señal (óxido nítrico y
transportadores de glutamato).
Los osteocitos son células metabólicamente activas y multifuncionales. Intervienen en el proceso de
mecanotransducción, en el cual los osteocitos responden a las fuerzas mecánicas aplicadas al hueso. Debido a la poca
flexibilidad ósea, las fuerzas mecánicas aplicadas causan el flujo del líquido intersticial de los canalículos y lagunas hacia
12
el lado comprimido del hueso. Este movimiento a través del sistema canalicular genera un potencial eléctrico transitorio
(potencial de flujo) en el momento en que se aplica la fuerza. El potencial de flujo abre los conductos de calcio
dependientes de voltaje en las membranas de los osteocitos, sobre las que fluye líquido tisular. Los aumentos resultantes
en el calcio intracelular, el ATP, la concentración de óxido nítrico y la síntesis de prostaglandina E2 alteran la expresión de
los genes c-fos y cox-2 responsables de la formación de hueso. La expresión del gen IGF-1 genera un aumento de la
producción de sí mismo, que promueve la conversión de células osteoprogenitoras en osteoblastos. Por lo tanto, las
regiones óseas sobre las que se aplica más fuerza, son las que tendrán mayor deposición de hueso nuevo.
Un osteocito responde a una fuerza mecánica reducida al secretar metaloproteinasas de la matriz (MMP). El espacio vacío
que rodea a los osteocitos es el resultado de la degradación enzimática de la matriz ósea por las MMP. Los osteocitos son
responsables del remodelado reversible de su matriz ósea pericanalicular y perilagunar, proceso denominado remodelado
osteocítico.
ESTADOS FUNCIONALES DE LOS OSTEOCITOS EN EL REMODELADO OSTEOCÍTICO
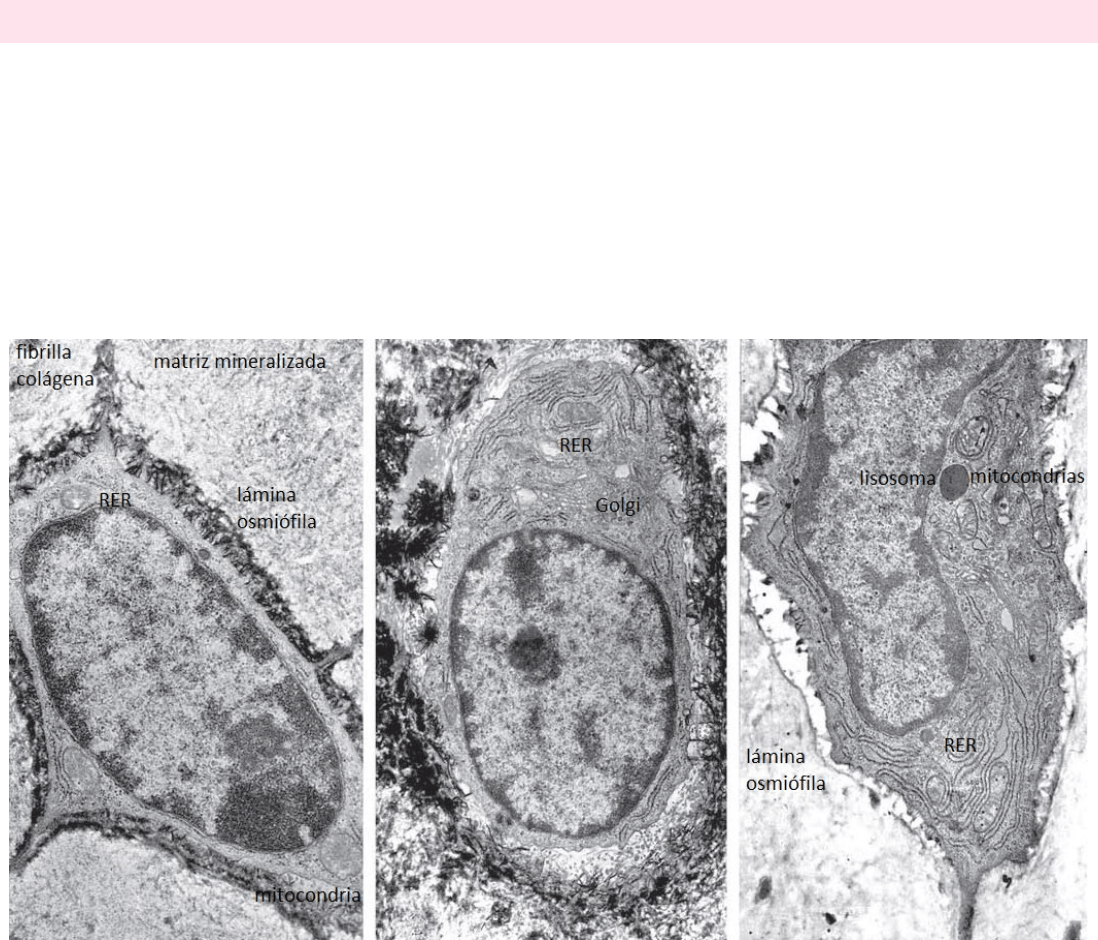
Existen 3 estados funcionales, cada uno con una morfología característica:
o Osteocitos latentes: escaso RER y pequeño aparato de Golgi. Una lámina osmiofílica que representa la matriz madura
calcificada es vista cercanamente opuesta a la membrana celular. (primera foto)
o Osteocitos formativos: exhiben indicios de formación de matriz y presentan características similares a los
osteoblastos. RER y aparato de Golgi más abundantes y se observa osteoide en el espacio pericelular dentro de la
laguna. (segunda foto)
o Osteocitos resortivos: gran cantidad de cisternas del retículo endoplásmico y un aparato de Golgi bien desarrollado.
Lisosomas visibles. (tercera foto)

13
CÉLULAS DE REVESTIMIENTO ÓSEO
Se ubican en los sitios en los que no se está produciendo remodelado del tejido óseo, se presentan como una capa de
células aplanadas con poco citoplasma y escasos orgánulos en las superficies óseas. Las células de revestimiento óseo
ubicadas en las superficies externas de hueso reciben el nombre de células del periostio y las que tapizan las superficies
internas se denominan células del endostio.
Constituyen una población celular que deriva de los osteoblastos. Intervienen en el mantenimiento y el soporte
nutricional de los osteocitos incluidos en la matriz ósea subyacente y regulan el movimiento del calcio y el fosfato desde
y hacia el hueso. Los procesos celulares de las células de revestimiento óseo se extienden dentro de los canalículos del
hueso contiguo y se comunican por medio de uniones de hendidura con las evaginaciones de los osteocitos.
OSTEOCLASTOS
Son células multinucleadas grandes que aparecen en los sitios donde ocurre la resorción ósea. Están apoyados
directamente sobre la superficie ósea en proceso de resorción. Como resultado de la actividad de los osteoclastos, se
forma una excavación llamada laguna de resorción que se puede observar en el hueso directamente bajo al osteoclasto.
Derivan de la fusión de células progenitoras hematopoyéticas monocelulares, las células progenitoras de
granulocitos/macrófagos (GMP, CFU-GM) que dan origen a los linajes de granulocitos y monocitos. La formación de
osteoclastos se produce en asociación con las células del estroma de la médula ósea, que secretan citosinas esenciales
para la diferenciación, tanto de los osteoclastos como de los macrófagos, a partir de células progenitoras GMP, que
incluyen el factor estimulante de colonias de monocitos (M-CSF), el TNF y varias IL.
El osteoblasto recién formado tiene que activarse para convertirse en una célula de resorción ósea. Cuando resorben
hueso en forma activa, los osteoclastos exhiben 3 regiones especializadas:
1. Borde festoneado: porción de la célula en contacto directo con el hueso. Contiene pliegues profundos de la MP
que forman estructuras del tipo de microvellosidades, responsables del aumento de la extensión de superficie
para la exocitosis de enzimas hidrolíticas, la secreción de protones por las bombas dependientes de ATP y la
endocitosis de productos de degradación y detritos óseos.
2. Zona clara: citoplasma en forma de anillo contiguo al borde festoneado que delimita la superficie ósea en
resorción. Es donde se produce la resorción y degradación de la matriz. Contiene abundantes microfilamentos de
actina dispuestos en forma de anillo rodeados por proteínas fijadoras de actina. LA MP de la zona clara contiene
moléculas de adhesión célula-matriz extracelular, responsables de proporcionar un sello hermético entre la MP y
la matriz ósea mineralizada.
3. Región basolateral: interviene en la exocitosis del material digerido.
La mayoría de las vesículas del osteoclasto son los lisosomas. Sus contenidos se liberan al espacio extracelular de las
hendiduras que hay entre pliegues del borde festoneado, como ejemplo, las enzimas lisosómicas. Una vez liberadas, estas
enzimas hidrolíticas degradan el colágeno y otras proteínas de la matriz ósea.
Sin embargo, antes de que pueda producirse la digestión, la matriz ósea tiene que descalcificarse a través de la
acidificación de la superficie ósea, que inicia la disolución de la matriz mineral. El citoplasma del osteoclasto contiene
anhidrasa carbónica II, que produce ácido carbónico (H2CO3) a partir de CO2 y H20. El ácido carbónico se disocia en
bicarbonato (HCO3-) y H+. Con la ayuda de bombas de protones dependientes de ATP, los protones se transportan a través
del borde festoneado, generando un pH bajo (4 o 5) en el ambiente de la laguna en resorción. Este ambiente ácido, creado

14
en el espacio extracelular entre el hueso y el osteoclasto, está protegido por la zona clara. El medio ácido inicia la
degradación del componente mineral del hueso (principalmente hidroxiapatita) para convertirlo en iones de calcio,
fosfatos inorgánicos solubles y agua. Cuando la resorción del tejido óseo se ha completado, los osteoclastos sufren
apoptosis.
Los materiales digeridos del hueso resorbido se transportan en vesículas endocíticas por todo el osteoclasto. El contenido
de ellas que se originan en el borde festoneado se libera en la membrana basal, que generalmente está en contacto con
los vasos sanguíneos.
FORMACIÓN DEL HUESO
El desarrollo del hueso tradicionalmente se clasifica en endocondral o intramembranosa. La distinción entre ambas radica
en si un modelo de cartílago sirve como el precursor óseo (osificación endocondral) o si el hueso está formado por un
método más sencillo, sin la intervención de un cartílago precursor (osificación intramembranosa). Huesos de las
extremidades y partes del esqueleto axial que soportan peso se desarrollan por osificación endocondral. Huesos planos
del cráneo y de la cara, la mandíbula y la clavícula se desarrollan por osificación intramembranosa.
OSIFICACIÓN INTRAMEMBRANOSA
La primera evidencia de la osificación intramembranosa en los seres humanos se ve alrededor de la 8va semana de
gestación dentro del mesénquima. Las células mesenquimatosas migran y se acumulan en áreas específicas, donde
forman los centros de osificación, iniciando el proceso de osificación intramembranosa.
Las células mesenquimatosas en estos centros se alargan y diferencian en células osteoprogenitoras, que expresan el
factor de transcripción CBFA1, esencial para la diferenciación de osteoblastos y la expresión de genes necesarios para la
osificación. El citoplasma de las células cambia de eosinófilo a basófilo, y el aparato de Golgi se observa como una región
clara evidente. Estos cambios resultan del osteoblasto diferenciado que, entonces, secreta los colágenos, sialoproteínas
óseas, la osteocalcina y otros componentes de la matriz ósea (osteoide= porción sin mineralizar). Los osteoblastos se
acumulan en la periferia del centro de osificación y continúan secretando osteoide. A medida que continúa el proceso, el
osteoide se mineraliza y los osteoblastos se convierten en osteocitos. Dentro de la matriz ósea, los osteocitos se separan
cada vez más unos de otros mientras se produce más matriz, pero permanecen en contacto a través de sus evaginaciones.
Con el tiempo, la matriz se calcifica y los procesos citoplasmáticos intercomunicados de los osteocitos quedan contenidos
dentro de los canalículos.
OSIFICACIÓN ENDOCONDRAL
También comienza con la proliferación y acumulación de células mesenquimatosas en el sitio donde se desarrollará el
futuro hueso. Bajo la influencia de los factores de crecimiento de fibroblastos (FGF) y diferentes proteínas morfogénicas
óseas (BMP) las células mesenquimatosas expresan inicialmente colágeno tipo II y se diferencian en condroblastos que, a
su vez, producen matriz cartilaginosa.
Una vez establecido, el modelo cartilaginoso experimenta crecimiento intersticial y por aposición. El aumento en la
longitud del modelo cartilaginoso se atribuye al crecimiento intersticial. El aumento de espesor se debe a la adición de
matriz cartilaginosa producida por los nuevos condrocitos diferenciados a partir de la capa condrogénica del pericondrio
que rodea la masa de cartílago.
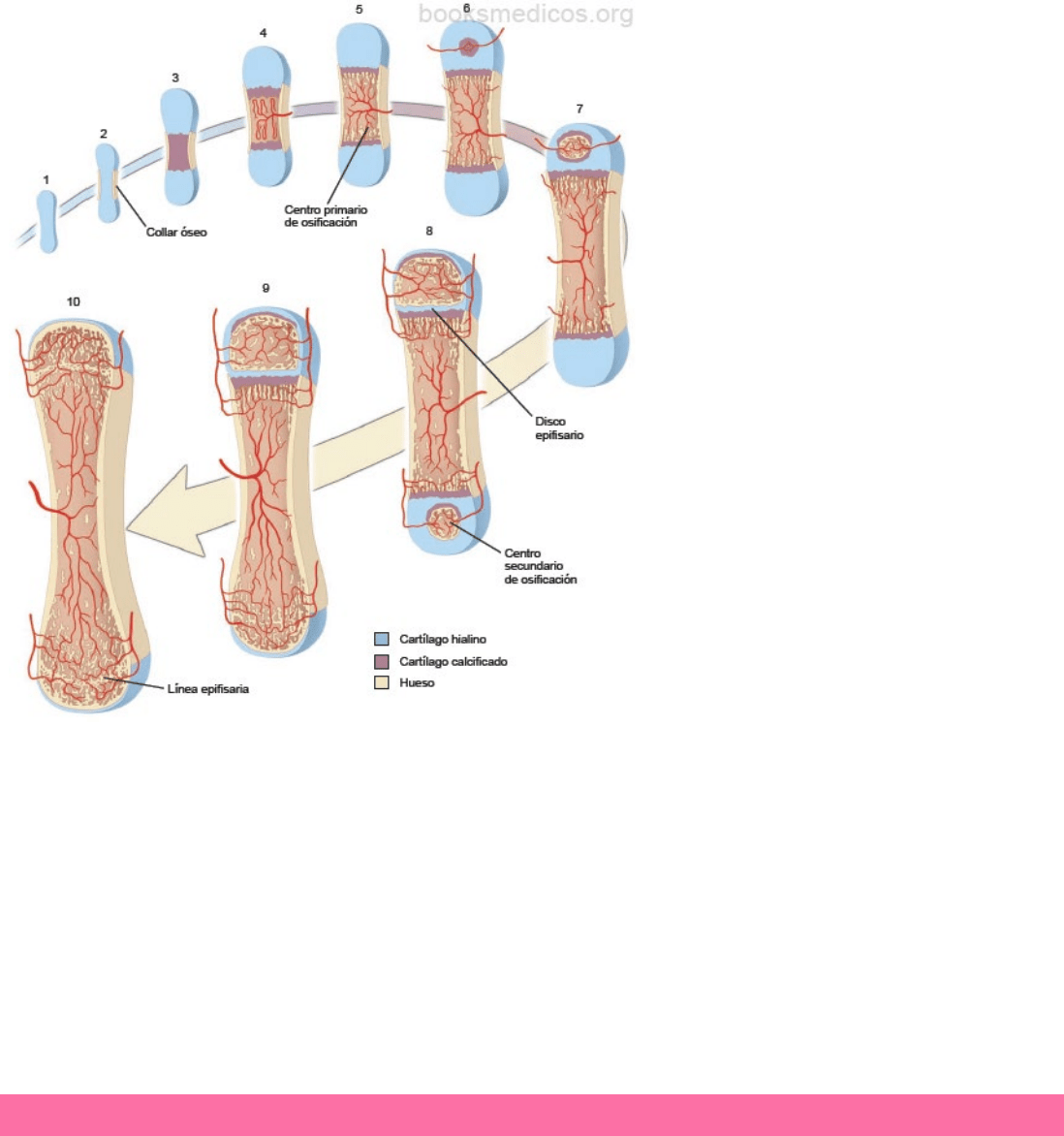
15
En esta etapa, las células del pericondrio en la
región media del modelo cartilaginoso dejan de
producir condrocitos. En su lugar, se originan
osteoblastos. Por lo tanto, el tejido conectivo
que rodea esta porción funcionalmente ya no
es pericondrio, sino que es periostio. Como
consecuencia, se forma una delgada capa de
tejido óseo alrededor del modelo cartilaginoso,
que puede denominarse hueso subperióstico.
En caso de un hueso largo, alrededor del
modelo cartilaginoso en la porción de la diáfisis
del hueso en desarrollo, se forma una cubierta
de tejido óseo subperióstico, llamado collar
óseo.
A medida que los condrocitos aumentan de
tamaño, la matriz cartilaginosa circundante se
resorbe, formando delgadas placas de cartílago
entre las células hipertróficas. Éstas comienzan
a sintetizar fosfatasa alcalina, al mismo tiempo,
la matriz cartilaginosa circundante se calcifica.
La matriz cartilaginosa calcificada impide la
difusión de las sustancias nutritivas y causa la
muerte de los condrocitos en el modelo del
cartílago. Con esto, gran parte de la matriz se degrada y las lagunas vecinas confluyen para forman una cavidad cada vez
más grande. Mientras se produce esto, uno o varios vasos sanguíneos proliferan a través del collar óseo en la diáfisis para
vascularizar la cavidad.
Las células madre mesenquimatosas que residen en el periostio en desarrollo, migran junto con los vasos sanguíneos
invasores y se diferencian en células osteoprogenitoras en la cavidad medular. Las células madre hematopoyéticas (HSC)
también llegan a la cavidad a través del nuevo sistema vascular y abandonan la circulación para dar origen a la médula
ósea que incluye todos los linajes de las células sanguíneas. A medida que se degrada el cartílago calcificado y se elimina
parcialmente, quedan restos con el aspecto de espículas irregulares. Cuando las células osteoprogenitoras se adhieren a
estas espículas, se convierten en osteoblastos y comienzan a sintetizar tejido óseo (osteoide). El hueso formado de esta
manera se le denomina hueso endocondral. Este sitio donde comienza a formarse tejido óseo en la diáfisis de un hueso
largo se llama centro primario de osificación.
CRECIMIENTO DEL HUESO ENDOCONDRAL
La etapa inicial de osificación endocondral comienza alrededor de la 12va semana de gestación. El crecimiento del hueso
endocondral se inicial en el segundo trimestre de la vida fetal y continúa después del nacimiento hasta el principio de la
vida adulta.
A medida que la cavidad medular de la diáfisis se agranda, pueden reconocerse distintas zonas de cada extremo de la
cavidad. El remanente tejido cartilaginoso, denominado cartílago epifisiario, muestra zonas bien definidas. Durante la
Este documento contiene más páginas...
Descargar Completo
tejido cartilaginoso y oseo.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.