
María Paula Lorenzo
El tejido adiposo es un tejido conjuntivo especializado que cumple una función importante en la
homeostasis energética.
En el tejido conjuntivo laxo se encuentran células adiposas o
adipocitos, de manera individual o formando
grupos. El tejido en el que los adipocitos son el tipo celular primario se denomina tejido adiposo. Los
adipocitos desempeñan un papel fundamental en la homeostasis energética.
Para poder sobrevivir, el cuerpo necesita asegurar la entrega continua de energía a pesar del suministro
muy variable de sustancias nutritivas desde el medio externo. Para satisfacer la demanda de energía del
cuerpo cuando hay escasez de alimentos, el tejido adiposo almacena con mucha eficiencia el exceso de
energía. El organismo posee una capacidad limitada para almacenar hidratos de carbono y proteínas; por
lo tanto, las reservas de energía se almacenan dentro de las
gotitas de lípidos de los adipocitos en la
forma de triglicéridos. Éstos representan una forma dinámica de almacenamiento de energía, que se
incrementa cuando la ingesta de alimentos es mayor que el consumo energético, y se utiliza cuando el
consumo energético es mayor que la ingesta de alimentos. La energía depositada en los adipocitos puede
liberarse con rapidez para ser usada en otros sitios del organismo.
Los
triglicéridos son la forma más concentrada de almacenamiento de energía metabólica disponible para
el ser humano. Dado que carecen de agua, poseen alrededor del doble de la densidad energética de los
hidratos de carbono y las proteínas. La densidad energética de los
triglicéridos es de aproximadamente
37,7 kJ/g (9 cal/g), en tanto que la de los hidratos de carbono y las proteínas es de 16,8 kJ/g (4 cal/g). En
el caso de la privación de alimentos (inanición), los triglicéridos son una fuente esencial de agua y energía.
Algunos animales pueden depender sólo del agua metabólica obtenida a partir de la oxidación de ácidos
grasos para mantener su equilibrio hídrico. Por ejemplo, las jorobas del camello están formadas
principalmente por tejido adiposo, y a partir de ellas obtiene el agua y la energía necesarias para sobrevivir
en el desierto.
Los
adipocitos cumplen otras funciones además de su papel como reservorio para el almacenamiento de
grasa. También regulan el metabolismo energético mediante la secreción de sustancias paracrinas y
endocrinas. El reciente descubrimiento de las funciones secretoras de los adipocitos ha cambiado las
perspectivas acerca del tejido adiposo, y en la actualidad se le considera un órgano endocrino importante.
Existe evidencia que relaciona el incremento en la actividad endocrina de los adipocitos con las
complicaciones metabólicas y cardiovasculares asociadas con la obesidad.
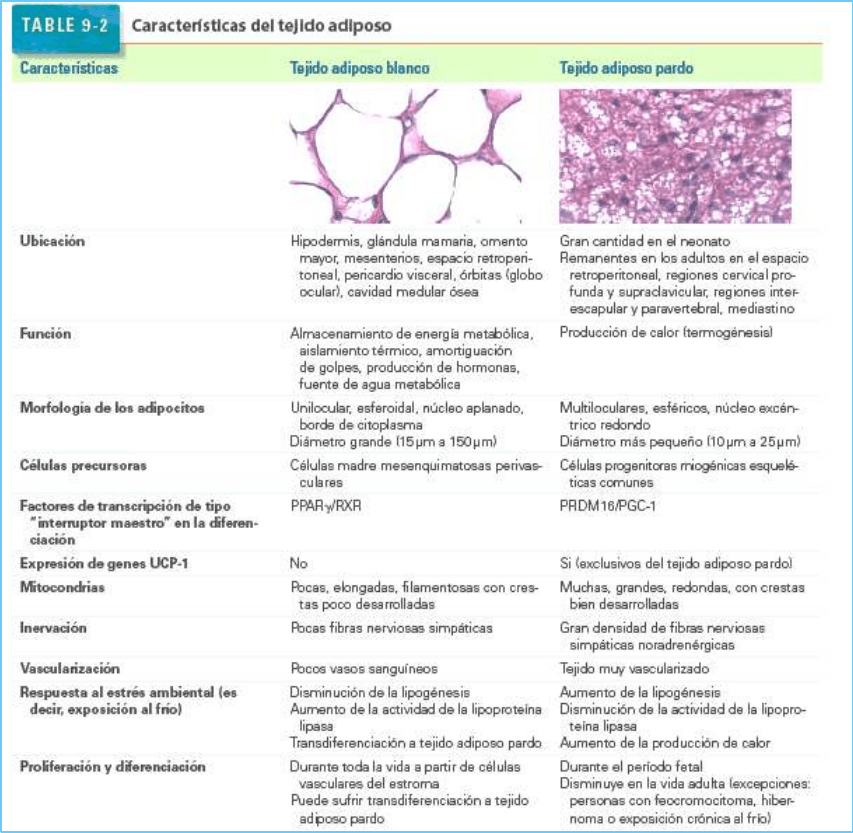
Existen dos tipos de tejido adiposo: blanco (unilocular) y pardo (multilocular).
Los dos tipos de tejido adiposo se denominan tejido adiposo blanco y tejido adiposo pardo debido a su
color en estado vivo:
• El tejido adiposo blanco es el tipo predominante en el ser humano adulto.
• El tejido adiposo pardo está presente en el ser humano en grandes cantidades durante la vida
fetal. Disminuye a lo largo de la primera década de vida pero continúa presente en cantidades
variadas, en especial, alrededor de los órganos internos.

María Paula Lorenzo
Función del tejido adiposo blanco
Las funciones del tejido adiposo blanco incluyen almacenamiento de energía, aislamiento térmico,
amortiguamiento de los órganos vitales y secreción de hormonas.
El tejido adiposo blanco (unilocular) representa al menos el 10 % del peso corporal total de un individuo
saludable normal. Forma la capa adiposa de la
fascia subcutánea (superficial) llamada panículo adiposo (lat.
panniculus, tela fina) en el tejido conjuntivo subcutáneo. Dado que la conductividad térmica del tejido
adiposo es sólo alrededor de la mitad de la del músculo esquelético, la fascia subcutánea (hipodermis)
provee un aislamiento importante contra el frío porque reduce la pérdida de calor. Este tejido adiposo se
concentra bajo la piel del abdomen, la región glútea, las axilas y los muslos. Las diferencias en el espesor
de esta capa adiposa según el sexo están dadas, en parte, por las diferencias entre las siluetas masculina
y femenina. En ambos sexos, la región mamaria es un sitio preferencial para la acumulación del tejido
adiposo; este tejido es el componente principal de la
mama no lactante. En la mujer lactante, la almohadilla
de grasa mamaria desempeña un papel importante en el sustento de la función de las mamas. Provee
lípidos y energía para la producción de leche y también es un sitio de síntesis de diferentes factores de
crecimiento que modulan las respuestas a los distintos esteroides, proteínas y hormonas que actúan sobre
la función de la glándula mamaria.
En los órganos internos, el tejido adiposo se localiza de forma preferencial en el omento mayor, el
mesenterio y el espacio retroperitoneal, en donde suele ser abundante alrededor de los riñones. También
se encuentra en la
médula ósea y entre otros tejidos, donde rellena espacios. En la palma de las manos y
la planta de los pies, por debajo del pericardio visceral (que tapiza la superficie externa del corazón) y en la
órbita del globo ocular, el tejido adiposo funciona como una almohadilla protectora. Mantiene esta función
estructural inclusive durante la ingesta calórica reducida; cuando el tejido adiposo de otros sitios agota sus
lípidos, el tejido adiposo estructural no disminuye.
El tejido adiposo blanco secreta una variedad de adipocinas, que incluyen hormonas, factores de
crecimiento y citocinas.
Los
adipocitos sintetizan y secretan adipocinas, un grupo de sustancias biológicamente activas que incluyen
hormonas, factores de crecimiento y citocinas. Por esta razón, el tejido adiposo es considerado muy
importante en la homeostasis energética, la adipogénesis, el metabolismo de esteroides, la angiogénesis y
la respuesta inmunitaria. El miembro más importante de las adipocinas es la leptina (gr. leptos, delgado),
una hormona peptídica de 17 kDa descubierta en 1994. La leptina interviene en la regulación de la
homeostasis energética y es un producto exclusivo de los adipocitos. La leptina inhibe la ingesta de
alimentos y estimula la tasa metabólica y la pérdida de peso corporal. De esto modo, la leptina cumple los
criterios de un
factor de saciedad circulante que controla la ingesta de alimentos cuando el depósito de
energía del organismo es suficiente. La leptina también participa en un mecanismo de señalización
endocrino que informa sobre el estado energético del tejido adiposo a los centros cerebrales
que regulan la ingesta de alimentos. Actúa sobre el sistema nervioso central, fijándose a receptores
específicos ubicados principalmente en el
hipotálamo. Además, la leptina informa sobre el estado de
reserva de combustible en los adipocitos de los sitios de almacenamiento de lípidos a otros tejidos
metabólicamente activos (es decir, del tejido adiposo al muscular de un sitio diferente).
Además de la leptina, el tejido adiposo secreta una variedad de
adipocinas como
adiponectina, resistina, proteína de unión a
retinol 4 (RBP4), visfatina, apelina, inhibidor del activador de
plasminógeno-1 (PAI-1), factor de necrosis tumoral (TNF),
interleucina-6 (IL-6), proteína quimiotáctica de monocitos-1
(MCP-1) y
angiotensinógeno (AGE). La leptina también produce
hormonas esteroides (testosterona, estrógeno y
glucocorticoides). Algunas adipocinas también se sintetizan en
María Paula Lorenzo
otros tejidos. Por ejemplo, el AGE se sintetiza en el hígado; el
aumento de la producción de este péptido hormonal contribuye a
la hipertensión (tensión arterial elevada), que es una complicación
frecuente de la obesidad. Las hormonas sexuales y los
glucocorticoides no se sintetizan de novo; en cambio, surgen de la
conversión de formas inactivas
por la acción de enzimas específicas expresadas en los adipocitos.
Por consiguiente, estas enzimas pueden influir sobre el perfil de
esteroides sexuales de las personas obesas. En la obesidad, el
aumento de la secreción de
factores de crecimiento (factor de
necrosis tumoral a [TNF-a], factor de crecimiento transformante b
[TGF-b], y factor de crecimiento similar a la insulina I (IGF-I)) y
citocinas (IL-6 y prostaglandinas) podría estar relacionado con
alteraciones metabólicas y la aparición de diabetes.
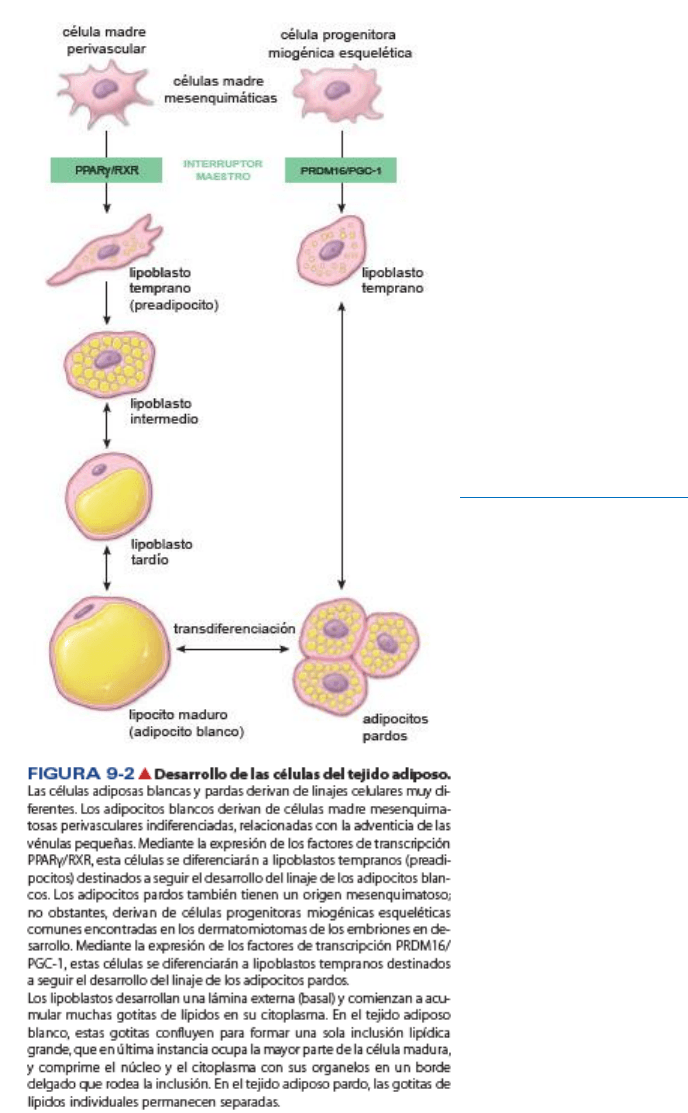
Diferenciación de adipocitos
Los adipocitos blancos se diferencian a partir de las células madre
mesenquimatosas bajo el control de los factores de transcripción
PPARg/RXR.
Durante el desarrollo embrionario, los
adipocitos blancos se
forman a partir de las células madre
mesenquimatosas
perivasculares indiferenciadas que se encuentran en la adventicia
de las vénulas pequeñas. La información actual
sugiere que un
factor de transcripción llamado
receptor gamma activado por
proliferador peroxisómico (PPARg), en un complejo con el receptor
X de retinoides (RXR), desempeña un papel decisivo en la
diferenciación de los adipocitos
y la iniciación del metabolismo
de los lípidos. Este
complejo induce la maduración de los
lipoblastos (adipoblastos) tempranos o preadipocitos hacia
células adiposas del tejido adiposo blanco. La mayoría de los
genes diana del PPARg en el tejido adiposo ejerce un efecto
sobre los mecanismos
lipógenos e inicia el almacenamiento de
triglicéridos. En consecuencia, el complejo PPARg/RXR se
considera un regulador de tipo
“interruptor maestro” en la
diferenciación
de los adipocitos blancos.
El tejido adiposo blanco comienza a formarse en la vida fetal.
Los
lipoblastos, se desarrollan inicialmente en el feto a partir de
células del estroma vascular situadas a lo largo de los vasos
sanguíneos pequeños, y no poseen lípidos. A pesar de ello, estas
células están destinadas a convertirse en adipocitos ya en esta
etapa temprana mediante la expresión de los factores de
transcripción PPARg/RXR. En ocasiones, la agrupación de estas células se denomina como
órganos adiposos
primitivos. Se caracterizan por la presencia de lipoblastos tempranos y capilares que proliferan en forma
activa. La acumulación de lípidos en los lipoblastos produce la morfología típica de los adipocitos
.
Los lipoblastos tempranos se parecen a los fibroblastos pero desarrollan inclusiones lipídicas pequeñas y
una lámina externa delgada.
Los estudios realizados con el microscopio electrónico de transmisión (MET) revelaron que
los lipoblastos
tempranos tienen configuración alargada, evaginaciones citoplasmáticas múltiples y gran abundancia de
retículo endoplásmico y de aparato de Golgi. A medida que se inicia la diferenciación de los lipoblastos,
aumenta la cantidad de vesículas y disminuye el retículo endoplásmico rugoso (RER). En un polo del
citoplasma aparecen
inclusiones lipídicas pequeñas. También aparecen vesículas pinocíticas y una lámina
externa. La presencia de una lámina externa es una característica que distingue aún más a los adipocitos
de las células propias del tejido conjuntivo.
María Paula Lorenzo
Los lipoblastos intermedios se tornan ovoides conforme la acumulación de lípidos cambia las dimensiones
celulares.
Con el desarrollo continuo, los lipoblastos tempranos asumen una configuración oval. La característica más
distintiva en esta etapa, es una gran concentración de vesículas y de
pequeñas gotitas lipídicas alrededor
del núcleo que se extienden hacia ambos polos de la célula. En la periferia de las inclusiones lipídicas
se presentan partículas de glucógeno, y se tornan más obvias las vesículas pinocíticas y la lámina basal.
Estas células se denominan lipoblastos intermedios.
El adipocito maduro se caracteriza por una sola inclusión lipídica muy grande rodeada por un reborde
delgado de citoplasma.
En la etapa final de la diferenciación, las células aumentan de tamaño y se tornan más esféricas. Las
pequeñas gotitas de lípido confluyen para formar una sola gota lipídica grande que ocupa la porción central
del citoplasma. El retículo endoplásmico liso (REL) es abundante, en tanto que el RER es menos prominente.
Estas células se denominan lipoblastos tardíos. Después de un tiempo, la masa lipídica comprime el núcleo
y lo desplaza hacia una posición excéntrica, lo cual produce el aspecto de un anillo de sello en las
preparaciones teñidas con hematoxilina y eosina (H&E). Debido a que estas células poseen una sola
inclusión lipídica, reciben el nombre de
adipocitos uniloculares (lat. unus, uno; loculus, sitio o lugar
pequeño) o lipocitos maduros.
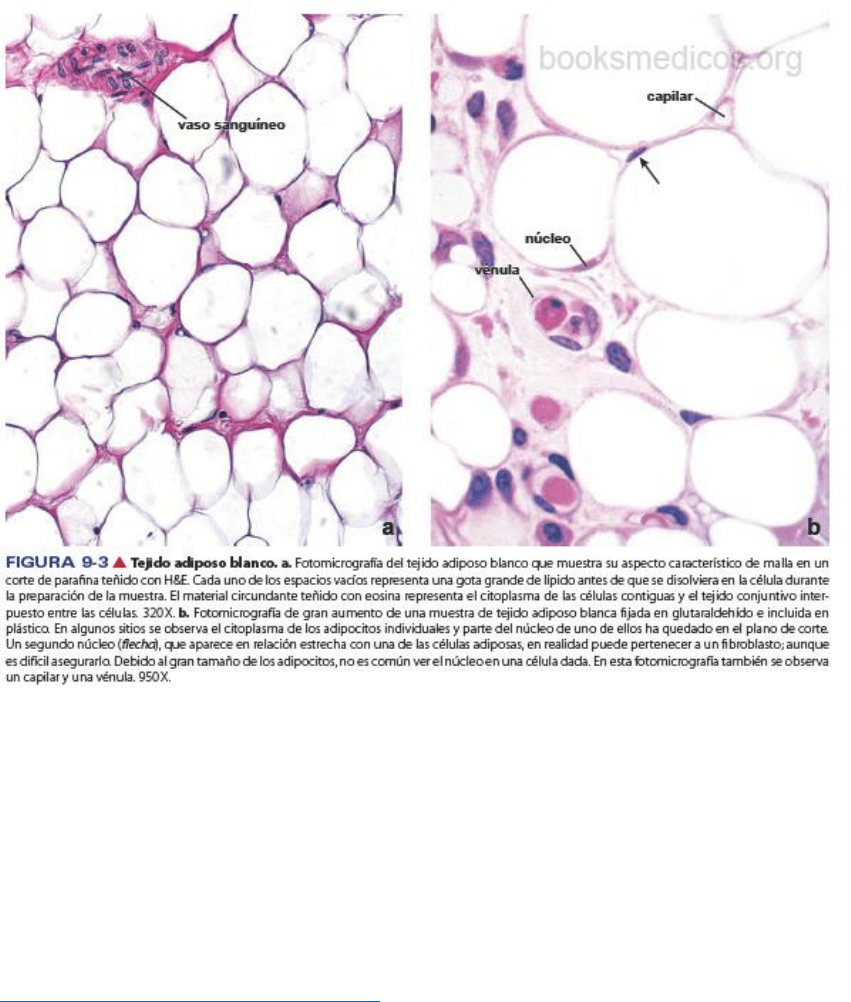
Estructura de los adipocitos y del tejidoadiposo
Los adipocitos uniloculares son células grandes, en ocasiones con un diámetro de 100 μm o más.
Cuando se encuentran aislados, los adipocitos uniloculares son esféricos, pero adoptan una forma ovalada
o poliédrica al agruparse en el tejido adiposo. El gran tamaño de estas células se debe al lípido acumulado
en las mismas. El núcleo se aplana y se desplaza hacia un lado de la masa lipídica; el citoplasma forma un
borde delgado alrededor del lípido. En las preparaciones histológicas de rutina, las grasas se disuelven por
acción de los solventes orgánicos, como el xileno; por consiguiente, el aspecto del tejido adiposo es el de
una delicada malla con diseños poligonales. La fina hebra de la malla que separa los adipocito contiguos
María Paula Lorenzo
corresponde al citoplasma de ambas células y a una pequeña cantidad de matriz extracelular. No obstante,
esta hebra suele ser tan delgada que sus componentes no se pueden determinar con el microscopio óptico.
El tejido adiposo recibe irrigación abundante a través de los vasos sanguíneos, y los capilares se pueden
observar en los ángulos de la malla donde convergen adipocitos contiguos. Las impregnaciones argénticas
muestran que los adipocitos están rodeados por fibras reticulares (colágeno tipo III), que son secretadas
por ellos mismos. Otros métodos especiales también revelan la presencia de fibras nerviosas amielínicas y
gran cantidad de mastocitos.
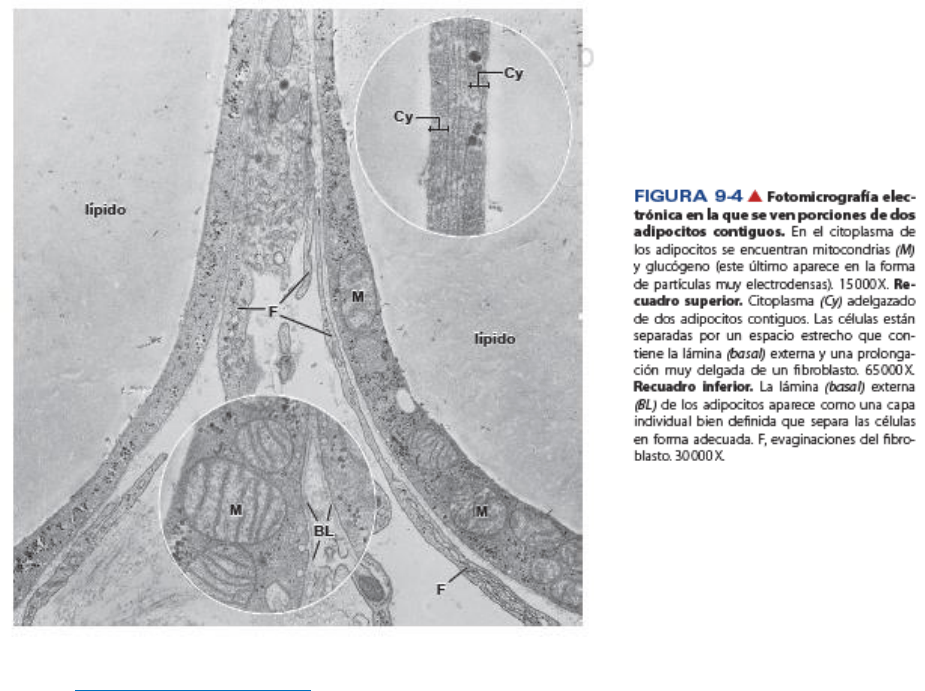
La masa lipídica del adipocito no está rodeada por membrana.
El MET demuestra que la interfaz entre la grasa contenida y el citoplasma circundante del adipocito está
compuesta por una capa de lípidos condensados de 5 nm de espesor, reforzada por
filamentos de
vimentina paralelos con un diámetro de 5 nm a 10 nm. Esta capa separa el contenido hidrófobo de la gotita
lipídica de la matriz citoplasmática hidrófila.
El citoplasma perinuclear del adipocito contiene un aparato de Golgi pequeño, ribosomas libres, perfiles
de RER cortos, microfilamentos y filamentos intermedios. En el reborde encuentran formas filamentosas
de mitocondrias y muchos perfiles de REL.
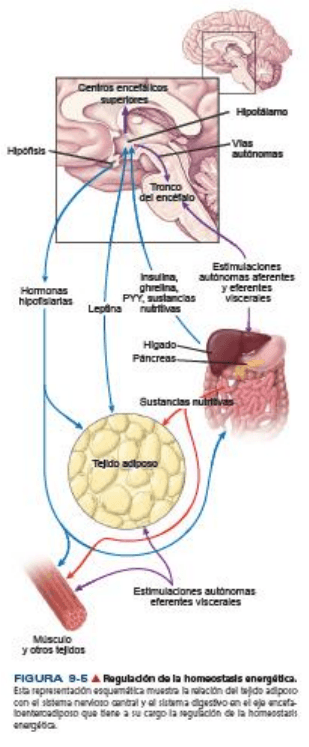
Regulación del tejido adiposo
Es casi imposible separar la regulación del tejido adiposo de los procesos digestivos y de las funciones del
sistema nervioso central. Estas señales hormonales y nerviosas interconectadas que surgen del tejido
adiposo, del tubo digestivo y del sistema nervioso central forman el eje encefaloenteroadiposo que regula
el apetito, el hambre, la saciedad y la homeostasis energética.
La cantidad de tejido adiposo en una persona está determinada por dos sistemas fisiológicos: uno asociado
con la regulación del peso a corto plazo y el otro asociado con la regulación del peso a largo plazo.
La cantidad de tejido adiposo en una persona es regulada por dos sistemas fisiológicos. El primer sistema,
que está asociado con la
regulación del peso a corto plazo, controla el apetito y el metabolismo en forma
María Paula Lorenzo
cotidiana. Recientemente, se han vinculado con este sistema dos hormonas peptídicas sintetizadas en el
tubo digestivo, conocidas como ghrelina, un estimulante del apetito y el
péptido YY (PYY), un supresor
del apetito. El segundo sistema, que está asociado con la
regulación del peso a largo plazo, controla el
apetito y el metabolismo en forma continua (durante meses o años). Dos hormonas principales, la
leptina
y la insulina, ejercen su efecto sobre este sistema junto con otras hormonas, como las hormonas tiroideas,
los glucocorticoides y las hormonas hipofisiarias.
La ghrelina y el péptido YY controlan el apetito como parte del sistema de
regulación del peso corporal a corto plazo.
El potente estimulante del apetito llamado
ghrelina, descubierto hace poco
tiempo, es un polipéptido pequeño de 28 aminoácidos producido por las células
epiteliales gástricas. Además de su función estimulante del apetito, actúa sobre el
lóbulo anterior de la glándula hipófisis para que libere la hormona del crecimiento.
En el ser humano, la ghrelina actúa a través de receptores ubicados en el
hipotálamo para aumentar la sensación de hambre. Por lo tanto, se considera que
es un factor “iniciador de la alimentación”.
Una mutación genética en el
cromosoma 15 causa el síndrome de Prader-Willi, en el cual una producción
excesiva de ghrelina conduce a una obesidad mórbida. En individuos con este
síndrome, suele observarse una alimentación compulsiva y una obsesión por los
alimentos desde muy jóvenes. El deseo de comer de estas personas es fisiológico
y abrumador, y resulta muy difícil de controlar. Si no se tratan, estos pacientes
mueren con frecuencia antes de los 30 años por complicaciones atribuibles a la
obesidad.
La pequeña hormona gastrointestinal de 36 aminoácidos de longitud llamada
péptido YY es producida por el intestino delgado y cumple una función importante
en la promoción y el mantenimiento de la pérdida de peso, ya que induce una
mayor sensación de saciedad poco después de una comida. También actúa a
través de receptores en el hipotálamo que suprimen el apetito. Disminuye la
ingesta alimentaria de las personas porque induce la saciedad y el deseo de dejar
de comer. En estudios clínicos experimentales, se ha demostrado que la infusión
de PYY en seres humanos reduce la ingesta de alimentos en un 33 % en un período
de 24 horas.
Dos hormonas, la leptina y la insulina, tienen a su cargo la regulación del peso
corporal a largo plazo.
El descubrimiento del
gen de la leptina (ob) que codifica un ARN mensajero
(ARNm) adiposo-específico para leptina, ha mejorado los conocimientos sobre el
mecanismo de la homeostasis energética. En modelos con animales de experimentación, la adición de
leptina recombinante a ratones obesos ob/ob con deficiencia de leptina, hace que reduzcan la ingesta de
alimentos y pierdan alrededor del 30 % de su peso corporal total, después de dos semanas de tratamiento.
A diferencia de lo que ocurre con los ratones mutantes, en la mayoría de las personas obesas, tanto la
concentración del ARNm de leptina en el tejido adiposo, como la concentración de leptina en suero, se
encuentran elevadas. Esto se comprobó en todos los tipos de obesidad, sin importar si las causas eran
factores genéticos, lesiones hipotalámicas o un aumento en la eficiencia de la utilización de los alimentos.
Por causas que aún no se conocen, los adipocitos en estas personas obesas son resistentes a la acción de
la leptina, y la administración de esta hormona no reduce la cantidad de tejido adiposo. En cambio, estudios
en personas cuyo peso había disminuido y en pacientes con
anorexia nerviosa, mostraron una disminución
significativa en la concentración del ARNm de leptina en su tejido adiposo y en la concentración de leptina
en suero. Recientes hallazgos clínicos indican que es muy probable que la leptina proteja el organismo
contra la pérdida de peso en los períodos de privación de alimento.
La insulina, la hormona pancreática que regula la concentración de glucosa en sangre (glucemia), también
participa en la regulación del metabolismo del tejido adiposo. Estimula la conservación de glucosa en los
triglicéridos de la gotita lipídica por el adipocito. Al igual que la leptina, la insulina
regula el peso porque
actúa sobre centros nerviosos superiores en el hipotálamo.

María Paula Lorenzo
A diferencia de la leptina, la insulina es necesaria para la acumulación de tejido adiposo. En la actualidad,
el diseño de fármacos contra la obesidad está centrado en sustancias que puedan inhibir los mecanismos
de señalización de la insulina y la leptina en el hipotálamo.
Factores neurales y hormonales influyen en el depósito y la movilización de los lípidos.
Una de las principales funciones metabólicas del tejido adiposo comprende la
captación de ácidos grasos
de la sangre y su
conversión en triglicéridos dentro del adipocito. Después, los triglicéridos se almacenan
en la gotita lipídica de la célula. Cuando el tejido adiposo es estimulado por mecanismos neurales u
hormonales, los triglicéridos se desdoblan en glicerol y ácidos grasos, un proceso denominado movilización.
Los ácidos grasos atraviesan la membrana del adipocito para introducirse en un capilar. Aquí se unen a la
proteína transportadora
albúmina y son transportados a otras células que utilizan los ácidos grasos como
combustible metabólico.
La movilización neural es de particular importancia durante los períodos de ayuno y de exposición a frío
intenso. Durante las etapas iniciales de la inanición experimental en roedores, las células de una
almohadilla adiposa desnervada continúan acumulando grasa. Los adipocitos de la almohadilla
contralateral intacta movilizan los lípidos. En la actualidad, se sabe que la
noradrenalina (liberada por los
axones de las neuronas del sistema nervioso simpático) inicia una serie de pasos metabólicos que conducen
a la activación de la
lipasa. Esta enzima desdobla los triglicéridos, que constituyen más del 90 % de los
lípidos almacenados en el adipocito. Esta actividad enzimática es uno de los primeros pasos en la
movilización de los lípidos.
La movilización hormonal comprende un sistema complejo de hormonas y enzimas que controla la
liberación de ácidos grasos desde los adipocitos. Este sistema incluye la
insulina, las hormonas tiroideas y
los
esteroides suprarrenales. La insulina es una hormona importante que promueve la síntesis de lípidos
mediante la estimulación de la síntesis de enzimas de la lipogénesis (ácido graso sintetasa, acetil-CoA
carboxilasa), y suprime la degradación de lípidos debido a que inhibe la acción de la lipasa sensible a
hormonas y bloquea así la liberación de ácidos grasos. El glucagón, otra hormona pancreática, y la
hormona
de crecimiento de la glándula hipófisis, aumentan la utilización de los lípidos (lipólisis). Además, las
concentraciones elevadas del factor de
necrosis tumoral a (TNF-a) se han señalado como un factor causal
en el desarrollo de la resistencia a la insulina relacionada con la obesidad y la diabetes.
El tejido adiposo pardo, abundante en los neonatos, se encuentra muy reducido en los adultos.
El tejido adiposo pardo es un tejido termógeno esencial que se encuentra presente en grandes cantidades
en el neonato, lo que ayuda a protegerlo de la gran pérdida de calor que es resultado de la desfavorable
relación entre su superficie y su masa, y evita la hipotermia mortal (un importante riesgo de muerte en
los lactantes prematuros). En los neonatos, el tejido adiposo pardo representa casi el 5 % de la masa
corporal total y se localiza en el dorso, a lo largo de la mitad superior de la columna vertebral, y extendido
hacia los hombros. La cantidad de tejido adiposo pardo disminuye en forma gradual a medida que el
cuerpo crece, pero su distribución es amplia durante la primera década de vida en las regiones cervical,
axilar, paravertebral, del mediastino, esternal, y abdominal. Luego desaparece de casi todas partes,
excepto alrededor de los riñones, las glándulas suprarrenales y los grandes vasos (p. ej., aorta), y en
regiones de cuello (cervical profunda y supraclavicular), del dorso (interescapular y para vertebral) y del
tórax (mediastino).
La tomografía de emisión de positrones (PET), que se utiliza para detectar células
neoplásicas en base a su absorción de grandes cantidades de glucosa marcada radiactivamente (18F-FDG),
es capaz de detectar patrones característicos del tejido adiposo pardo dentro de las regiones del
organismo adulto que se mencionan arriba. Estos hallazgos se confirmaron con la biopsia del tejido.
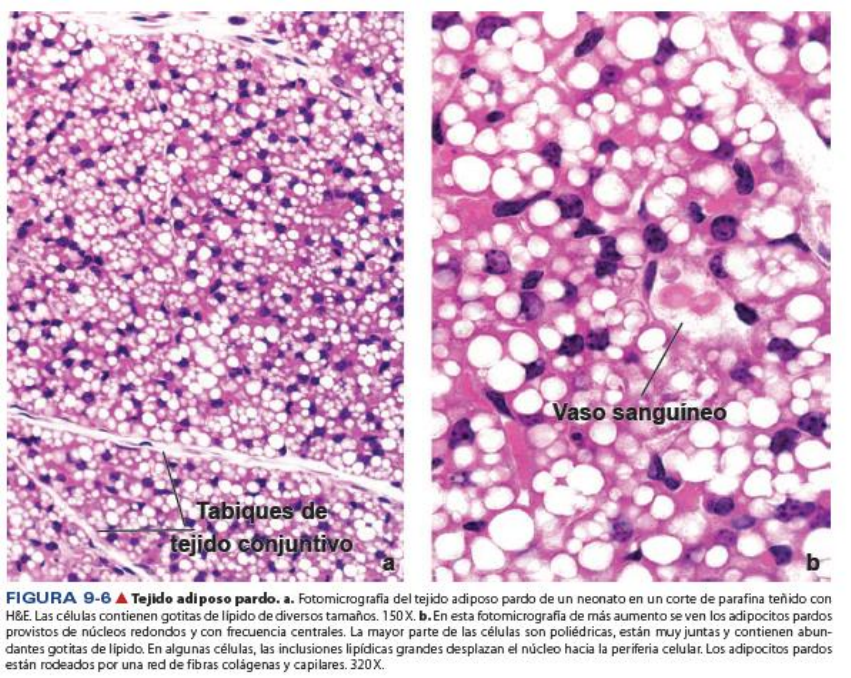
María Paula Lorenzo
Los adipocitos del tejido pardo (multilocular) contienen muchas gotitas de lípidos.
Las células del
tejido adiposo pardo (multilocular) son más pequeñas que las del tejido adiposo blanco
(unilocular). El citoplasma de cada célula contiene muchas gotitas de lípido pequeñas, de allí el nombre
de multilocular, en oposición al adipocito blanco unilocular que contiene una sola gota lipídica muy
grande. En general, el núcleo del adipocito pardo maduro es excéntrico pero no está aplanado como el
núcleo del adipocito blanco. En los cortes de rutina teñidos con H&E, el citoplasma de los adipocitos
pardos está compuesto sobre todo de espacios redondeados vacíos porque los lípidos que habitualmente
ocupan esos espacios, se pierden durante la preparación. El adipocito pardo contiene muchas
mitocondrias esféricas grandes con una gran cantidad de crestas, un aparato del Golgi pequeño, y sólo
pequeñas cantidades de RER y REL. Las mitocondrias contienen abundante cantidad de citocromo oxidasa,
la cual le confiere el color pardo a las células.
El tejido adiposo pardo está subdividido en lobulillos a través de tabiques de tejido conjuntivo, pero el
estroma conjuntivo entre las células de un mismo lobulillo es escasa. El tejido posee un extenso suministro
de capilares que realzan su color. Entre los adipocitos, se hallan presente abundantes fibras nerviosas
amielínicas adrenérgicas del sistema nervioso simpático.
Los adipocitos pardos se diferencian a partir de las células madre mesenquimatosas bajo el control de
los factores de transcripción PRDM16/PGC-1.
Los
adipocitos pardos también derivan de las células madre mesenquimatosas pero de un linaje distinto
del que derivan los adipocitos blancos. Los experimentos de rastreo de linaje demuestran que el tejido
adiposo pardo y el músculo esquelético derivan de
células progenitoras miogénicas esqueléticas
comunes presentes en los dermatomiotomas del embrión en desarrollo. A diferencia de lo que ocurre con
los adipocitos blancos, la diferenciación de los adipocitos pardos está bajo el control directo de un par
distinto de factores de transcripción. Cuando la proteína con dedos de zinc llamada PRDM16 (PR domain
containing 16 =
proteína 16 con dominio PR) se activa, células progenitoras miogénicas sintetizan varios
miembros de la familia PGC-1 (PPAR ү coactivator-1 = coactivador 1 de PPARg) de factores de
transcripción. En consecuencia,
PRDM16/PGC-1 se considera un regulador de tipo “interruptor maestro”
de la diferenciación de los adipocitos pardos. Estos factores, a su vez regulan la expresión de genes (p. ej.,
UCP-1) que codifican una proteína mitocondrial específica llamada
proteína desacoplante (UCP-1 =
uncoupling protein 1) o
termogenina (una proteína de 33 kDa de la membrana mitocondrial interna), que
es indispensable para el metabolismo de los adipocitos pardos (termogénesis). Las observaciones clínicas
confirman que en condiciones normales, el tejido adiposo pardo puede expandirse en respuesta al
María Paula Lorenzo
aumento de la concentración sanguínea de noradrenalina. Esto se torna obvio en los pacientes que tienen
un feocromocitoma, un tumor endocrino de la médula suprarrenal que secreta cantidades excesivas de
adrenalina y noradrenalina. En estos pacientes, el gen UCP-1 se activa por la estimulación de la
noradrenalina, que también protege los adipocitos pardos mediante la inhibición de la apoptosis.
Anteriormente, se creía que las proteínas desacoplantes sólo se expresaban en el tejido adiposo pardo.
Recientemente, se han descubierto varias proteínas desacoplantes similares en otros tejidos. La UCP-2
está vinculada con la hiperinsulinemia y la obesidad, y podría participar en la regulación del peso corporal.
La UCP-3 se expresa en el músculo esquelético y podría ser la causa de los efectos termógenos de la
hormona tiroidea. Las UCP-4 y -5 son moléculas mitocondriales específicas del encéfalo.
El metabolismo de los lípidos en el tejido adiposo pardo genera calor en el proceso conocido como
termogénesis.
Los animales que hibernan poseen una gran cantidad de
tejido adiposo pardo. Este tejido les sirve como
una fuente disponible de lípidos. Al oxidarse, produce calor para aumentar la temperatura de la sangre
que circula a través de la grasa parda al momento de despertar de la hibernación, y para mantener la
temperatura corporal durante la exposición al frío. Este tipo de generación de calor se conoce como
termogénesis sin temblor. El tejido adiposo pardo también está presente en los animales que no hibernan
y en los seres humanos, e igualmente sirve como fuente de calor. El sistema nervioso simpático estimula
los adipocitos pardos para movilizar los lípidos y generar calor, como ocurre en el tejido adiposo blanco.
María Paula Lorenzo
Por lo tanto, es probable que el tejido adiposo pardo normalmente presente pueda inducirse y funcionar
en el contexto de la termogénesis adaptativa humana. Las investigaciones futuras están siendo dirigidas
hacia la búsqueda de mecanismos para aumentar la diferenciación del tejido pardo, lo cual puede
convertirse potencialmente en un tratamiento atractivo tanto para la obesidad inducida por la dieta como
para la adquirida en forma genética.
La actividad termógena del tejido adiposo pardo es facilitada por la UCP-1 que se encuentra en la
membrana mitocondrial interna.
Las mitocondrias de las células eucarióticas producen y almacenan energía en la forma de un gradiente
electroquímico de protones a través de la membrana mitocondrial interna. Como ya se describió (v. pág.
57-61), esta energía se utiliza para sintetizar adenosina trifosfato (ATP) cuando los protones retornan a la
matriz mitocondrial a través de la enzima ATP sintetasa ubicada en la membrana mitocondrial interna.
Las peculiares
mitocondrias grandes, redondas, que se encuentran en el citoplasma de las células del
tejido adiposo pardo contienen
la proteína desacoplante (UCP-1), que desacopla la oxidación de los
ácidos grasos de la producción de ATP. De este modo, los protones pueden retornar desde el espacio
intermembranal hacia la matriz mitocondrial junto con el gradiente sin pasar a través de la ATP sintetasa
y, por ende, sin producir ATP. Esto puede ocurrir porque se encuentra disponible una vía alternativa para
el retorno de los protones a través de la UCP-1, que facilita el transporte protónico a través de la
membrana mitocondrial interna. La salida de los protones del espacio intermembranal disipa el gradiente
protónico mitocondrial, y así desacopla la respiración de la síntesis de ATP. La energía producida por la
mitocondria entonces se disipa como calor en un proceso denominado
termogénesis.
La actividad metabólica del tejido adiposo pardo es regulada por el sistema nervioso simpático y está
relacionada con la temperatura ambiental exterior.
La actividad metabólica del tejido adiposo pardo es regulada, en gran medida, por la noradrenalina
liberada por las terminaciones nerviosas simpáticas, la cual estimula la lipólisis y la hidrólisis de los
triglicéridos, y aumenta la expresión y la actividad de las moléculas de
UCP-1 en las mitocondrias. En
animales de experimentación, se ha comprobado que la actividad de la UCP-1 aumenta durante la
exposición al frío. Además, el frío estimula la utilización de la glucosa en los adipocitos pardos por la
expresión excesiva de transportadores de glucosa (GLUT-4).
Estudios clínicos recientes que utilizaron
tomografía de emisión de positrones (PET) en adultos, han demostrado una relación directa entre la
temperatura exterior y la cantidad de grasa parda acumulada en el organismo. Se ha reportado un
aumento de la cantidad de tejido adiposo pardo en el cuello en general y en la región supraclavicular
durante los meses invernales, en especial en las personas delgadas. Este fenómeno se sustenta también
en los hallazgos e
autopsias, de una cantidad mayor de grasa parda en quienes trabajan a la intemperie y se exponen al frío.
En la actualidad, las técnicas modernas de obtención de imágenes moleculares permiten que los clínicos
identifiquen con precisión los sitios de distribución de la grasa parda en el organismo, lo cual
es indispensable para el diagnóstico adecuado de las lesiones neoplásicas.

Sistema Linfático ROSS.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.