
APARATO URINARIO
FUNCIONES GENERALES
● Los riñones regulan el volumen y la composición de la sangre, ayudan a regular la presión arterial, el pH
y la glucemia, producen dos hormonas (calcitriol y eritropoyetina) y excretan los desechos en la orina.
● Los uréteres transportan la orina desde los riñones hasta la vejiga.
● La vejiga almacena la orina y la excreta a través de la uretra.
● La uretra expulsa la orina del cuerpo.
➤ RIÑÓN
Funciones:
★ Regulación de la composición iónica de la sangre: en especial sodio (Na+), potasio (K+), calcio (Ca2+),
cloruro (Cl–) y fosfato (HPO4 2–).
★ Regulación del pH sanguíneo: excretan una cantidad variable de iones hidrógeno (H+) hacia la orina y
conservan los iones bicarbonato (HCO3 –).
★ Regulación de la volemia: a través de la conservación o la eliminación de agua en la orina. El aumento
de la volemia incrementa la tensión arterial y un descenso de ésta disminuye la tensión arterial.
★ Regulación de la tensión arterial: mediante la secreción de la enzima renina, que activa el sistema
renina-angiotensina aldosterona.
★ Mantenimiento de la osmolaridad de la sangre: través de la regulación de la pérdida de agua y, por otro
sistema, de la pérdida de solutos en la orina.
★ Producción de hormonas: el calcitriol, la forma activa de la vitamina D, ayuda a regular la homeostasis
del calcio y la eritropoyetina estimula la producción de eritrocitos
★ Regulación de la glucemia: pueden utilizar el aminoácido glutamina para la gluconeogénesis, que es la
síntesis de nuevas moléculas de glucosa, y luego liberar glucosa hacia la sangre para mantener una
glucemia normal.
★ Excreción de desechos y sustancias extrañas: algunos de los desechos excretados con la orina son el
producto de reacciones metabólicas, como el amoníaco y la urea, que se forman luego de la
desaminación de los aminoácidos, la bilirrubina procedente del catabolismo de la hemoglobina, la
creatinina de la degradación de la creatina fosfato en las fibras musculares y el ácido úrico del
catabolismo de los ácidos nucleicos.
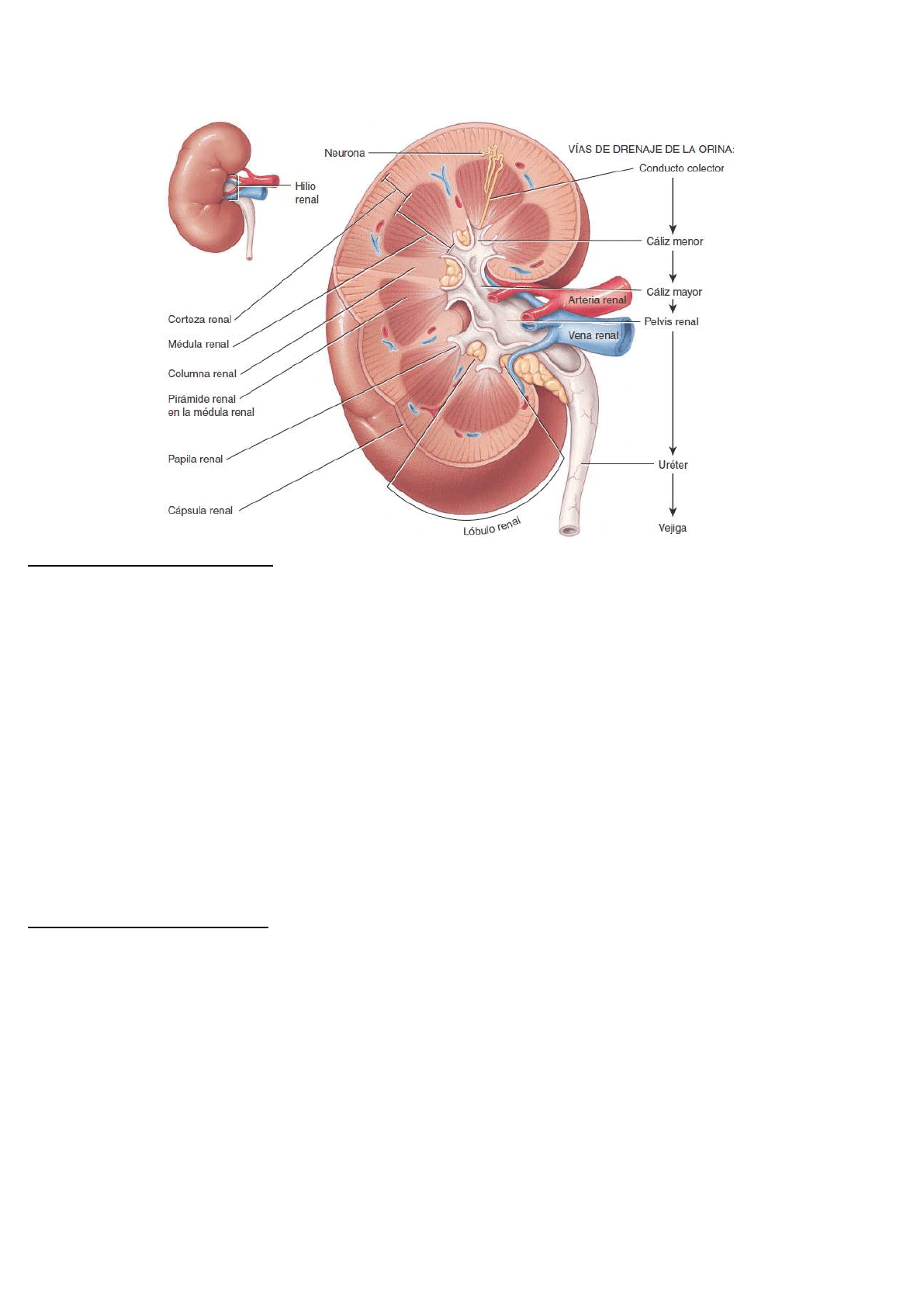
Anatomía externa de los riñones
El riñón típico de un adulto mide entre 10 y 12 cm de longitud. El borde medial cóncavo de cada riñón se
orienta hacia la columna vertebral. Cerca del centro de este borde cóncavo hay una escotadura llamada hilio
renal, a través del cual emerge el uréter junto con los vasos sanguíneos, los vasos linfáticos y los nervios.
Cada riñón está cubierto por tres capas de tejido:
❖ capa más profunda o cápsula renal, es una lámina lisa y transparente de tejido conectivo denso
irregular, que se continúa con la capa externa del uréter. Esta lámina sirve como barrera contra los
traumatismos y ayuda a mantener la forma del órgano.
❖ capa intermedia o cápsula adiposa, es una masa de tejido adiposo que rodea la cápsula renal. También
protege al riñón de los traumatismos y lo sostiene con firmeza en su sitio, dentro de la cavidad
abdominal.
❖ capa superficial o fascia renal es otra capa delgada de tejido conectivo denso irregular que fija el riñón a
las estructuras que lo rodean y a la pared abdominal. En la superficie anterior de los riñones, la fascia
renal es profunda con respecto al peritoneo.
Anatomía interna de los riñones
Un corte frontal del riñón muestra dos regiones distintas:
➢ un área superficial, de color rojo claro, llamada corteza renal (corteza = cubierta)
➢ una región profunda, de color pardo rojizo, denominada médula renal (médula = porción interna)
La médula renal está compuesta por entre 8 y 18 pirámides renales de forma cónica. La corteza renal es el
área de textura lisa que se extiende desde la cápsula hasta las bases de las pirámides renales y hacia los
espacios entre ellas. Se divide en una zona cortical externa y una zona yuxtamedular interna. Estas porciones
de la corteza renal que se extienden entre las pirámides renales se denominan columnas renales.
Un lóbulo renal consta de una pirámide renal, la región suprayacente de la corteza y la mitad de cada columna
renal adyacente. Juntas, la corteza y las pirámides renales de la médula constituyen el parénquima o porción
funcional del riñón. Dentro del parénquima se encuentran las unidades funcionales: las nefronas. El filtrado que
se forma en las nefronas drena en conductos papilares grandes, que se extienden a través de las papilas
renales de las pirámides. Los conductos papilares desembocan en estructuras en forma de copa llamadas
cálices menores y mayores. Cada riñón tiene entre 8 y 18 cálices menores y 2 o 3 cálices mayores.
❖ Un cáliz menor recibe orina de los conductos papilares de una papila renal y la envía a un cáliz mayor.
Una vez que ingresa el filtrado en los cálices se convierte en orina porque no experimenta más
reabsorción, ya que el epitelio simple de la nefrona y los conductos se convierte en el epitelio de
transición de los cálices.
❖ A partir de los cálices mayores, la orina drena en una cavidad más grande denominada pelvis renal y
luego, a través del uréter hacia la vejiga.
El hilio desemboca en una cavidad dentro del riñón que se denomina seno renal y que contiene parte de la
pelvis, los cálices y ramas de los vasos sanguíneos y los nervios renales.
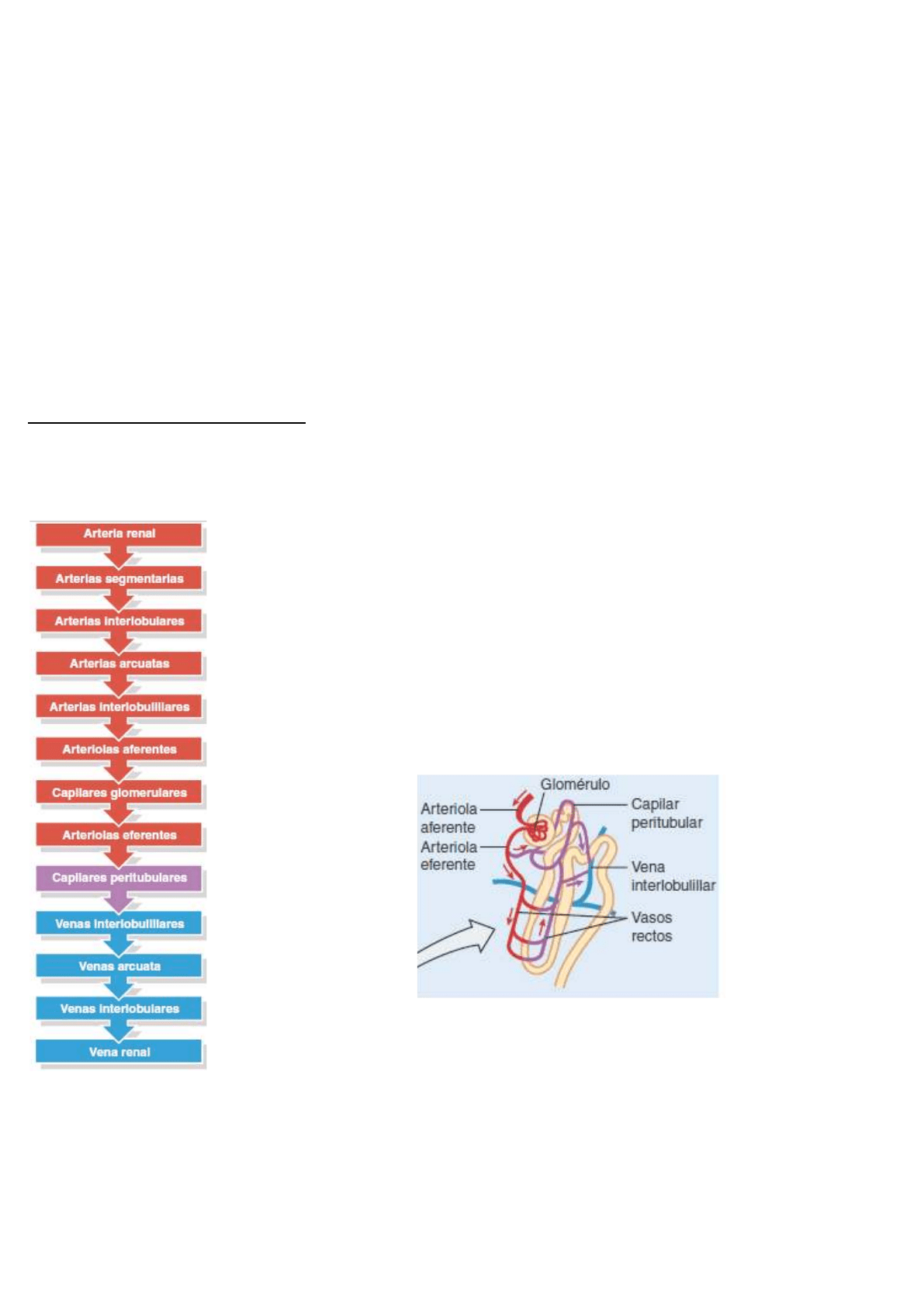
Irrigación e inervación de los riñones
En los adultos, el flujo sanguíneo renal, que es el flujo de sangre que atraviesa ambos riñones, es de alrededor
de 1200 mL por minuto.
¡ Los capilares glomerulares son únicos entre los capilares del cuerpo porque
están situados entre dos arteriolas, en lugar de interponerse entre una arteriola
y una vénula.
Muchos nervios renales se originan en el ganglio renal y pasan a través del plexo
renal hacia los riñones, junto con las arterias. Los nervios renales pertenecen a la
división simpática del sistema nervioso autónomo y en su mayor parte son nervios
vasomotores que regulan el flujo sanguíneo a través del riñón, lo que provoca
vasoconstricción o vasodilatación de las arteriolas renales.

➤ NEFRONA
Partes de la nefrona
Cada nefrona consta de dos partes: un corpúsculo (cuerpo diminuto) renal, donde se filtra el plasma sanguíneo,
y un túbulo renal, hacia el que pasa el líquido filtrado.
Los dos componentes del corpúsculo renal son el glomérulo (red capilar) y la cápsula glomerular (de Bowman),
que es una bolsa epitelial en forma de copa de pared doble, que rodea los capilares glomerulares.
El plasma sanguíneo se filtra en la cápsula glomerular y luego el líquido filtrado ingresa en el túbulo renal, que
tiene tres sectores principales.
1. el túbulo contorneado proximal
2. el asa de Henle (conecta tubulos)
3. el túbulo contorneado distal.
El corpúsculo renal y ambos túbulos contorneados se encuentran dentro de la corteza renal, mientras que el
asa de Henle se extiende hacia la médula renal, gira en forma de U y luego regresa a la corteza renal.
Los túbulos contorneados distales de diversas nefronas desembocan en un solo túbulo colector. Los túbulos
colectores luego se unen y convergen en varios cientos de conductos papilares grandes, que drenan
a su vez en los cálices menores.
Los conductos colectores y los papilares se extienden desde la corteza a través de la médula hacia la pelvis
renal, de manera que un riñón tiene alrededor de un millón de nefronas, pero
un número mucho menor de conductos colectores y aún menor de conductos
papilares.
La primera porción del asa de Henle penetra en la médula renal, donde recibe
el nombre de rama descendente. Luego gira en forma de U y regresa a la
corteza renal como la rama ascendente. Entre el 80 y el 85% de las nefronas
son nefronas corticales. Sus corpúsculos renales se encuentran en la región
externa de la corteza renal y tienen asas de Henle cortas, que se localizan
sobre todo en la corteza y atraviesan sólo la región externa de la médula. Las
asas de Henle cortas reciben su irrigación de los capilares peritubulares que
emergen de las arteriolas eferentes.
El otro 15-20% de las nefronas son yuxtamedulares. Sus corpúsculos
renales se encuentran en la profundidad de la corteza, cerca de la médula, y
tienen un asa de Henle larga que se extiende hasta la región más profunda de la médula. Las asas de Henle
largas reciben su irrigación de los capilares peritubulares y de los vasos rectos que emergen de las arteriolas
eferentes. Asimismo, la rama ascendente del asa de Henle de las nefronas yuxtamedulares consta de dos
porciones: una rama ascendente delgada, seguida por una rama ascendente gruesa.
La luz de la rama ascendente delgada es igual que en otras áreas del túbulo renal, sólo que el epitelio es más
fino. Las nefronas con asas de Henle largas les permiten a los riñones excretar orina muy diluida o muy
concentrada.
Histología de la nefrona y el túbulo colector
CÁPSULA GLOMERULAR: está constituida por
➔ la capa visceral está compuesta por células epiteliales pavimentosas simples modificadas, llamadas
podocitos. Las numerosas proyecciones en forma de pie de estas células (pedicelos) rodean la capa
simple de células endoteliales de los capilares glomerulares y forman la pared interna de la cápsula.
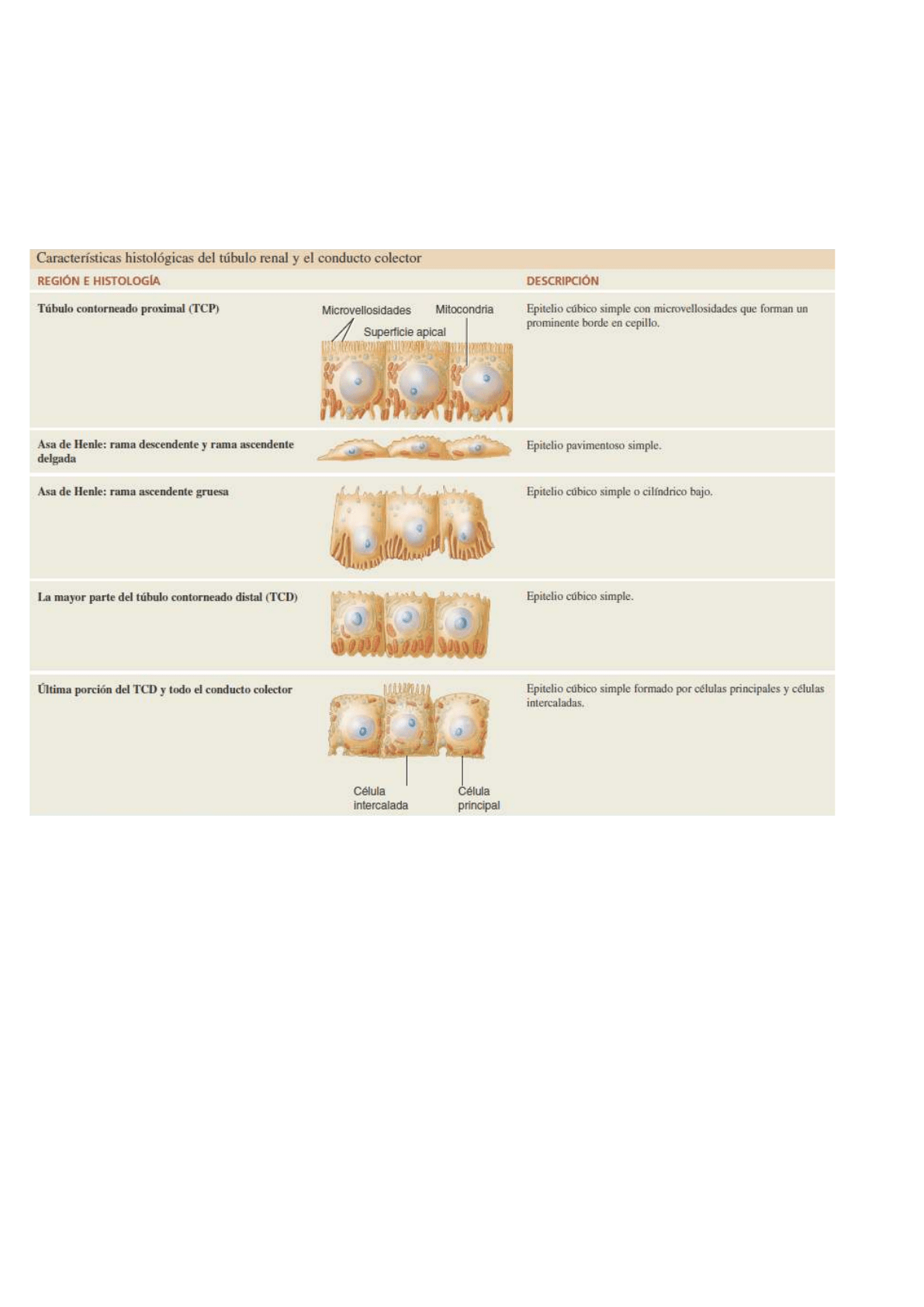
➔ La capa parietal de la cápsula glomerular está formada por epitelio pavimentoso simple y constituye la
pared externa de la cápsula. El líquido filtrado a través de los capilares glomerulares entra en el espacio
capsular, que se encuentra entre las dos capas de la cápsula glomerular y se considera la luz de la vía
urinaria.
TÚBULO RENAL Y TÚBULO COLECTOR
Para producir orina, las nefronas y los túbulos colectores desarrollan tres procesos básicos
1. Filtración glomerular: es el primer paso en la producción de orina. El agua y la mayor parte de los
solutos del plasma atraviesan la pared de los capilares glomerulares, donde se filtran e ingresan en la
cápsula de Bowman y luego, en el túbulo renal.
2. Reabsorción tubular. A medida que el líquido filtrado fluye a lo largo de los túbulos renales y los túbulos
colectores, las células tubulares reabsorben cerca del 99% del agua filtrada y diversos solutos útiles. El
agua y los solutos regresan a la sangre mientras ésta fluye a través de los capilares peritubulares y los
vasos rectos.
3. Secreción tubular. A medida que el líquido filtrado fluye a lo largo de los túbulos renales y los túbulos
colectores, las células tubulares secretan otras sustancias, como desechos, fármacos y compuestos
iónicos presentes en concentraciones excesivas, hacia el líquido filtrado.

La fracción del plasma que atraviesa las arteriolas aferentes de los riñones y se transforma en filtrado
glomerular es la fracción de filtración. A pesar de que la fracción de filtración típica normal oscila entre 0,16 y
0,2 (16-20%), el valor varía en forma considerable, tanto en condiciones de salud como de enfermedad.
En promedio, el volumen diario de filtrado glomerular en los adultos es de 150 L en las mujeres y de 180 L en
los hombres. Más del 99% del filtrado glomerular retorna a la corriente sanguínea por reabsorción tubular, de
modo que sólo 1-2 L se excretan como orina.
Membrana de filtración
Los capilares glomerulares y los podocitos, que rodean por completo los capilares, forman en conjunto una
barrera permeable denominada membrana de filtración. Esta configuración “en sándwich” permite la filtración
de agua y solutos pequeños, pero impide que se filtren la mayor parte de las proteínas del plasma, las células
sanguíneas y las plaquetas.
Las sustancias que se filtran de la sangre atraviesan tres barreras:
1) Las células endoteliales glomerulares son bastante permeables porque tienen grandes fenestraciones
(poros) que miden entre 0,07 y 0,1 μm de diámetro. Este tamaño les permite a todos los solutos del
plasma salir de los capilares glomerulares, pero impide la filtración de las células sanguíneas y las
plaquetas. Entre los capilares glomerulares y la hendidura entre las arteriolas aferente y eferente hay
células mesangiales que ayudan a regular la filtración glomerular.
2) La lámina basal es una capa de material acelular que se encuentra entre el endotelio y los podocitos y
está compuesta por fibras pequeñas de colágeno, proteoglucanos y una matriz de glucoproteínas; las
plasmáticas más grandes con carga negativa.
3) Miles de procesos en forma de pie llamados pedicelos se extienden desde cada podocito y envuelven
los capilares glomerulares. Los espacios entre los pedicelos son las hendiduras de filtración. Una
membrana delgada, la membrana de la hendidura, se extiende a lo largo de cada hendidura de filtración
y permite el pasaje de moléculas con diámetro menor de 0,006-0,007 μm, como agua, glucosa,
vitaminas, aminoácidos, proteínas plasmáticas muy pequeñas, amoníaco, urea e iones. Menos del 1%
de la albúmina, que es la proteína plasmática más abundante, atraviesa la membrana de la hendidura,
ya que tiene un diámetro de 0,007 μm y es demasiado grande para pasar.
El principio de filtración, que es el uso de presión para obligar a los líquidos y los solutos a que atraviesan una
membrana, es el mismo en los capilares glomerulares que en el resto de los capilares del cuerpo. Sin embargo,
el volumen de líquido filtrado por el corpúsculo renal es mucho mayor que en otros capilares, debido a tres
razones:
❖ Los capilares glomerulares tienen una gran superficie para la filtración porque son largos y extensos.
Las células mesangiales regulan la proporción de esta superficie disponible para la filtración. Cuando
las células mesangiales están relajadas, la superficie es máxima y la filtración glomerular es muy alta.
La contracción de dichas células reduce la superficie disponible y, por ende, la filtración glomerular.
❖ La membrana de filtración es delgada y porosa. A pesar de tener varias capas, su espesor es sólo de
0,1 μm. Los capilares glomerulares también son 50 veces más permeables que los capilares de la
mayor parte de los tejidos, principalmente, debido a sus grandes fenestraciones.
❖ La presión en el capilar glomerular es alta. Como el diámetro de la arteriola eferente es menor que el de
la arteriola aferente, la resistencia al flujo sanguíneo fuera del glomérulo es elevada. Como resultado, la
presión sanguínea en los capilares glomerulares es bastante más alta que en los capilares de cualquier
otro sitio del cuerpo.
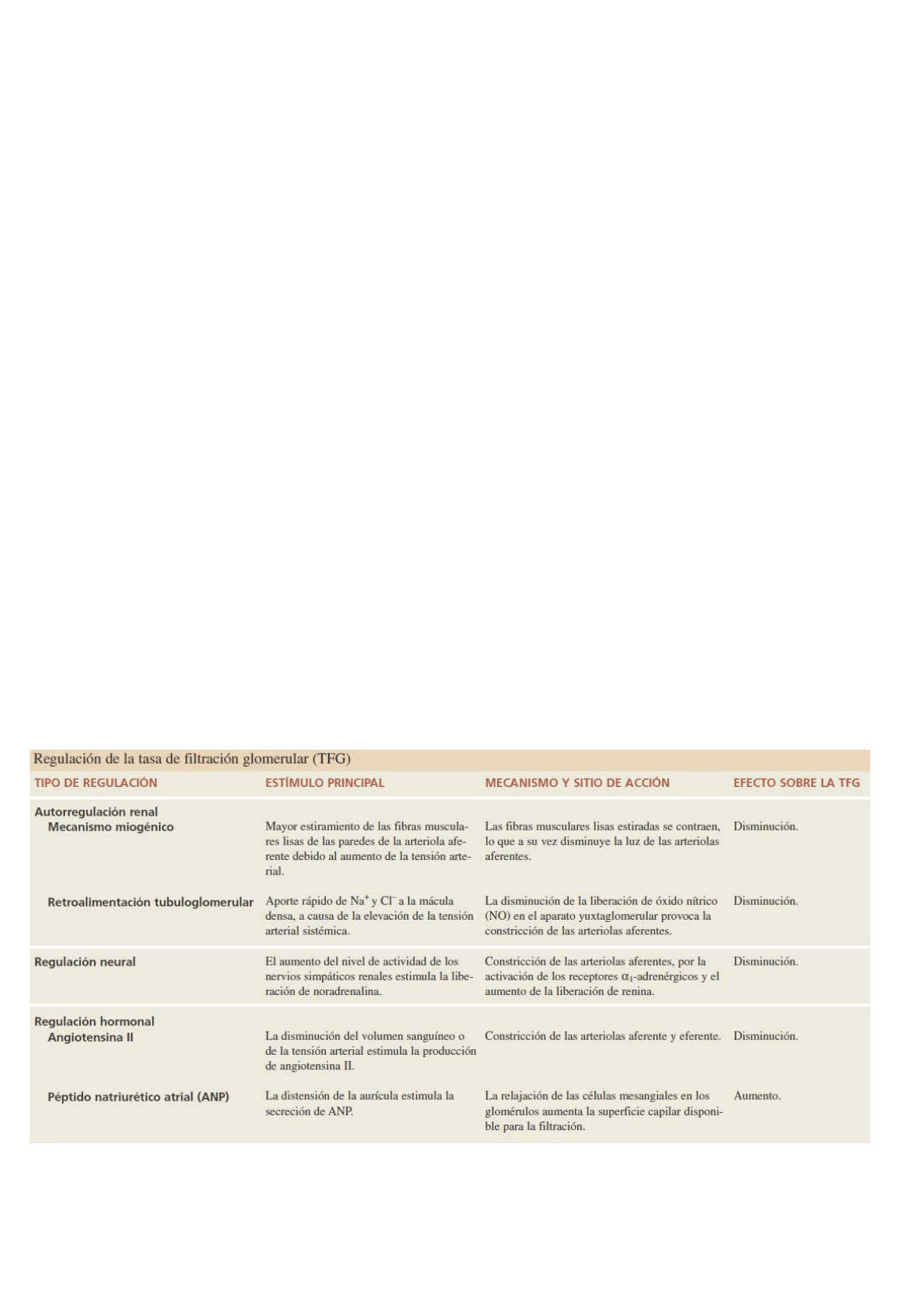
Presión de filtración neta
La filtración glomerular depende de tres presiones principales. Una de ellas promueve la filtración, y las dos
restantes se oponen a ella:
1) La presión hidrostática de la sangre glomerular (PHG) es la presión sanguínea en los capilares
glomerulares. Su valor suele aproximarse a 55 mm Hg. Promueve la filtración, al forzar la salida del
agua y los solutos del plasma, a través de la membrana de filtración.
2) La presión hidrostática capsular (PHC) es la presión hidrostática ejercida contra la membrana de
filtración por el líquido que ya está en el espacio capsular y el túbulo renal. Esta presión se opone a la
filtración y representa una “presión retrógrada” de alrededor de 15 mm Hg.
3) La presión osmótica coloidal de la sangre (POC), secundaria a la presencia de proteínas como la
albúmina, las globulinas y el fibrinógeno en el plasma, también se opone a la filtración. La presión
osmótica coloidal de la sangre promedio en los capilares glomerulares es de 30 mm Hg.
La presión de filtración neta (PFN), es decir, la presión total que promueve la filtración, se determina de la
siguiente manera:
(PFN) = PHG – PHC – POC
Tasa de filtración glomerular
REABSORCIÓN Y SECRECIÓN
La reabsorción, que es el retorno de la mayor parte del agua y de muchos de los solutos filtrados hacia la
corriente sanguínea, es la segunda función básica de la nefrona y el túbulo colector. En condiciones normales,
alrededor del 99% del agua filtrada se reabsorbe. Las células epiteliales a lo largo del túbulo renal y del túbulo
colector llevan a cabo la reabsorción, pero las células del túbulo contorneado proximal realizan la mayor
contribución.
La mayoría de las proteínas pequeñas y de los péptidos que pasan a través del filtro también se reabsorben, en
general, por pinocitosis.
La tercera función de las nefronas y los túbulos colectores es la secreción tubular, que es la transferencia de
sustancias desde la sangre y las células tubulares hacia el filtrado glomerular. Las sustancias secretadas son
iones hidrógeno (H+), K+ y amonio (NH4 +), creatinina y algunos fármacos como penicilina.
La secreción tubular tiene dos consecuencias importantes:
❖ la secreción de H+ ayuda a controlar el pH sanguíneo
❖ la secreción de otras sustancias contribuye a eliminarlas del cuerpo.
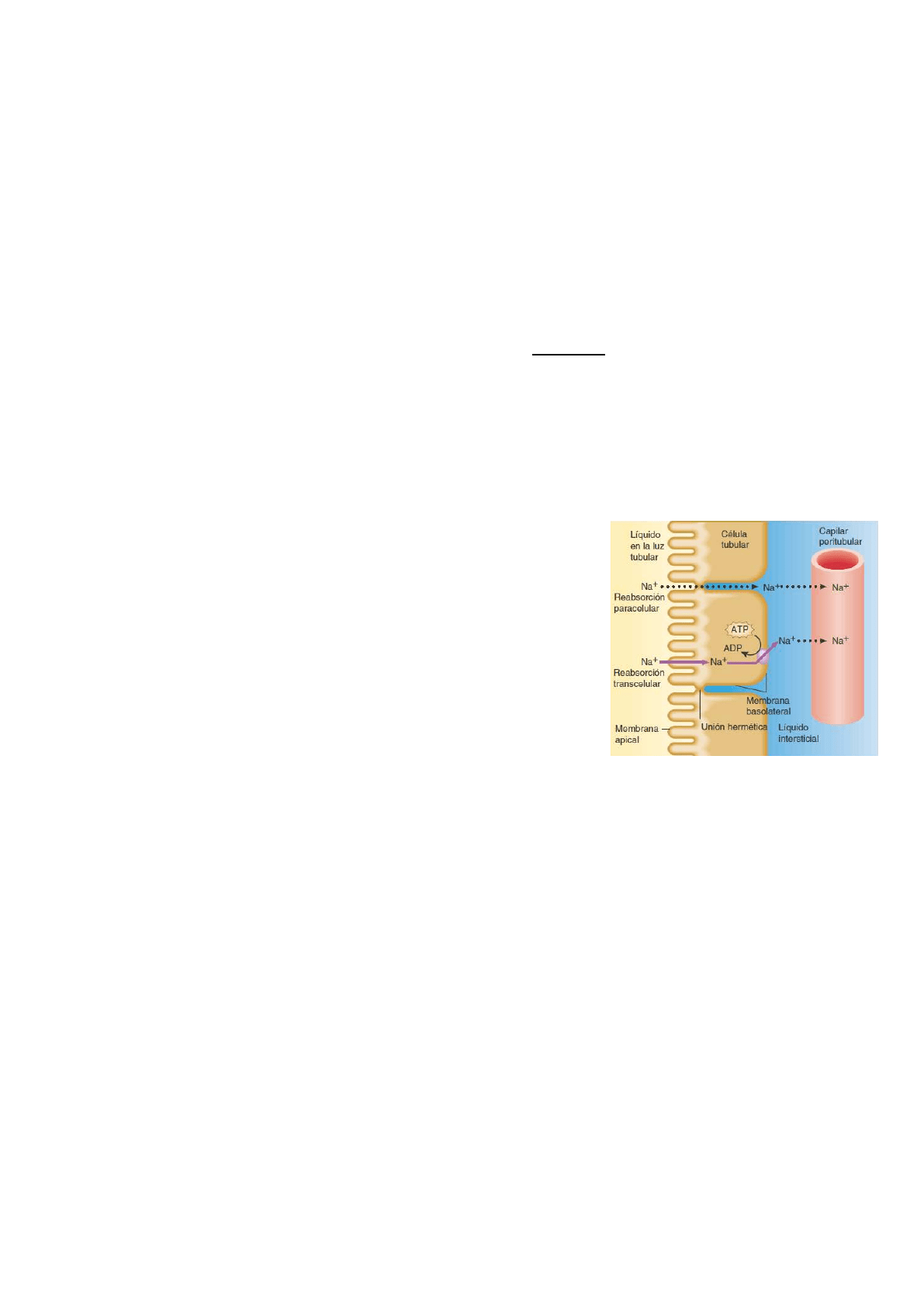
Vías de reabsorción
Una sustancia reabsorbida del líquido, en la luz del túbulo, puede seguir
uno de dos caminos antes de ingresar en el capilar peritubular:
puede desplazarse entre células tubulares adyacentes o a través de una
célula tubular.
A lo largo del túbulo renal, las uniones herméticas rodean y vinculan las
células contiguas entre sí. La membrana apical está en contacto con el
líquido tubular, y la membrana basolateral contacta con el líquido
intersticial en la base y los lados de la célula.
El líquido puede filtrarse entre las células, mediante un proceso pasivo
conocido como reabsorción paracelular.
Aunque las células epiteliales están conectadas por uniones herméticas,
estas uniones entre las células del túbulo contorneado proximal son
“permeables” y permiten la reabsorción de algunas sustancias en las células hacia los capilares peritubulares.
En la reabsorción transcelular, una sustancia pasa desde el líquido de la luz tubular a través de la membrana
apical de una célula tubular y del citosol hacia el líquido intersticial, desde donde atraviesa la membrana
basolateral.
Mecanismos de transporte
Cuando las células renales transportan solutos, dentro o fuera del líquido tubular, movilizan sustancias
específicas en una sola dirección.
La reabsorción de Na+ en los túbulos renales es muy importante, dada la gran cantidad de iones de sodio que
atraviesan los filtros glomerulares. Las células que revisten los túbulos renales, al igual que otras células del
cuerpo, tienen una baja concentración de Na+ en su citosol por la actividad de las bombas de sodio-potasio
(Na+/K+ATPasas). La ausencia de bombas de sodio-potasio en la membrana apical asegura que la
reabsorción de Na+ sea un proceso unidireccional. La mayor parte de los iones de sodio que atraviesan la
membrana apical se expulsan hacia el líquido intersticial, por la acción de la bomba en la base y los lados de
la célula. La cantidad de ATP que utilizan las bombas de sodio-potasio en los túbulos renales es el 6% del
consumo total del ATP del cuerpo en reposo.
La reabsorción de solutos rige la reabsorción de agua, ya que ésta se produce sólo por ósmosis.

➔ Cerca del 90% de la reabsorción del agua filtrada por los riñones tiene lugar junto con la reabsorción de
solutos como Na+, Cl– y glucosa. La reabsorción del agua junto con solutos en el líquido tubular se
denomina reabsorción de agua obligatoria porque el agua se ve “obligada” a seguir los solutos durante
su reabsorción. Este tipo de reabsorción se produce en el túbulo contorneado proximal y la rama
descendente del asa de Henle, puesto que estos segmentos de la nefrona siempre son permeables al
agua.
➔ La reabsorción del 10% restante del agua, un total de 10-20 L por día, se llama reabsorción de agua
facultativa. La reabsorción de agua facultativa es regulada por la hormona antidiurética y se lleva a
cabo, predominantemente, en los túbulos colectores.
⭄ Reabsorción y secreción en el túbulo contorneado proximal
La mayor parte de los solutos se reabsorben en el túbulo contorneado proximal, junto con el Na+. El transporte
del Na+ se produce por medio de cotransportadores y contratransportadores, en el túbulo contorneado
proximal. En condiciones normales, la glucosa, los aminoácidos, el ácido láctico, las vitaminas hidrosolubles y
otros nutrientes filtrados no se pierden con la orina, sino que se reabsorben por completo en la primera mitad
del túbulo contorneado proximal, a través de diversos tipos de cotransportadores de Na+ localizados en la
membrana apical.
Otros cotransportadores de Na+ presentes en el túbulo contornado proximal captan el HPO4 2– (fosfato) y SO4
2– (sulfato), todos los aminoácidos y el ácido láctico.
En otro proceso de transporte activo secundario, los contratransportadores
de Na+/H+ transportan el Na+ filtrado a favor de su gradiente
de concentración hacia las células del túbulo contorneado proximal,
junto con los H+, que se movilizan desde el citosol hacia la luz
(Figura 26.13a), lo que hace que el Na+ se reabsorba hacia la sangre y
los H+ se secreten hacia el líquido tubular.
La reabsorción de solutos en los túbulos contorneados proximales promueve la ósmosis de agua. Cada soluto
reabsorbido aumenta la osmolaridad, en primer lugar, dentro de la célula tubular, luego en el líquido intersticial
y por último en la sangre. Así, el agua se desplaza rápidamente desde el líquido tubular, tanto por la vía
paracelular como por la transcelular, hacia los capilares peritubulares y restablece el balance osmótico
Las células que tapizan el túbulo contorneado proximal y la rama descendente del asa de Henle son más
permeables al agua porque tiene muchas moléculas de acuaporina-1.
En la segunda mitad del túbulo contorneado proximal, los gradientes electroquímicos para Cl–, K+, Ca2+,
Mg2+ y urea promueven su difusión pasiva hacia los capilares peritubulares, tanto por la vía paracelular como
por la transcelular. Entre estos iones, el Cl– es el que se encuentra en mayor concentración. La difusión de las
moléculas de Cl– con carga negativa hacia el líquido intersticial, por la vía paracelular, hace que el líquido
intersticial sea más negativo que el líquido tubular. Esta negatividad estimula la reabsorción pasiva paracelular
de cationes, como K+, Ca2+ y Mg2+. Aunque en el sudor hay pequeñas cantidades de amoníaco y urea, la
excreción de estos productos de desecho nitrogenados se realiza por vía urinaria. La urea y el amoníaco
presentes en la sangre se filtran en el glomérulo y se secretan en las células del túbulo contorneado proximal,
hacia el líquido tubular.
⭄ Reabsorción en el asa de Henle
Como todos los túbulos contorneados proximales reabsorben cerca del 65% del agua filtrada. La composición
química del líquido tubular, en este sitio, es distinta de la del filtrado glomerular porque la glucosa, los
aminoácidos y otras sustancias ya no están presentes. Sin embargo, la osmolaridad del líquido tubular todavía
se parece a la de la sangre, ya que la reabsorción de agua por ósmosis se produce a la misma velocidad que la
reabsorción de solutos a todo lo largo del túbulo contorneado proximal.

El asa de Henle reabsorbe alrededor del 15% del agua filtrada, entre el 20 y el 30% del Na+ y el K+, el 35% del
Cl–, entre el 10 y el 20% del HCO3 – y una cantidad variable del Ca2+ y el Mg2+ filtrados. En este sitio, por
primera vez, la reabsorción de agua por ósmosis no se acopla en forma automática con la reabsorción de los
solutos filtrados porque parte del asa de Henle es relativamente impermeable al agua.
En el asa de Henle se inicia una etapa de regulación independiente, tanto del volumen como de la osmolaridad
de los líquidos corporales. Los gradientes electroquímicos promueven la reabsorción
pasiva de solutos, tanto por la vía paracelular como por la
transcelular.
Como hay muchos canales de K+ en la membrana apical, la mayor parte del K+ se moviliza por la acción de
cotransportadores que lo regresan al líquido tubular, a favor de su gradiente de concentración. En
consecuencia, el principal efecto de los cotransportadores de Na+- K+-2Cl– es la reabsorción de Na+ y Cl–.
Si bien alrededor del 15% del agua filtrada se reabsorbe en la rama descendente del asa de Henle, poco o
nada se reabsorbe en la rama ascendente. En este segmento del túbulo, las membranas apicales son casi
impermeables al agua y como se reabsorben iones pero no agua, la osmolaridad del líquido tubular se reduce
de manera progresiva, a medida que el líquido fluye hacia el final de la rama ascendente.
⭄ Reabsorción en la porción inicial del túbulo contorneado distal
El 80% del agua filtrada ya se reabsorbió. La porción inicial del túbulo contorneado distal reabsorbe alrededor
del 10-15% del agua filtrada, el 5% del Na+ filtrado y el 5% del Cl– filtrado. La reabsorción de Na+ y Cl– se
realiza a través de cotransportadores de Na+-Cl– en las membranas apicales.
La porción inicial del túbulo contorneado distal también es el principal sitio donde la hormona paratiroidea
(PTH) estimula la reabsorción del Ca2+. La cantidad total del Ca2+ reabsorbido en esta región varía de
acuerdo con las necesidades corporales.
⭄ Reabsorción y secreción en la porción final del túbulo contorneado distal y el túbulo
Cuando el líquido llega al final del túbulo contorneado distal, entre el 90 y el 95% del agua y los solutos filtrados
ya retornaron a la corriente sanguínea. Las células principales reabsorben Na+ y secretan K+; las células
intercaladas reabsorben K+ y HCO3 – y secretan H+.
En la porción distal del túbulo contorneado distal y en los túbulos colectores, la cantidad de agua y solutos
reabsorbidos y secretados varía, en función de las necesidades corporales.
A diferencia de lo que ocurre en segmentos anteriores de la nefrona, el Na+ atraviesa la membrana apical de
las células principales mediante canales de Na+, más que por medio de transportadores (cotransportadores o
contratransportadores).
En condiciones normales, la reabsorción transcelular y paracelular en el túbulo contorneado proximal y el asa
de Henle devuelven la mayor parte del K+ filtrado a la sangre. Parte del ion difunde a favor de su gradiente de
concentración hacia el líquido tubular, donde su concentración es muy baja. Este mecanismo de secreción es la
fuente principal de iones de K+ excretados con la orina.
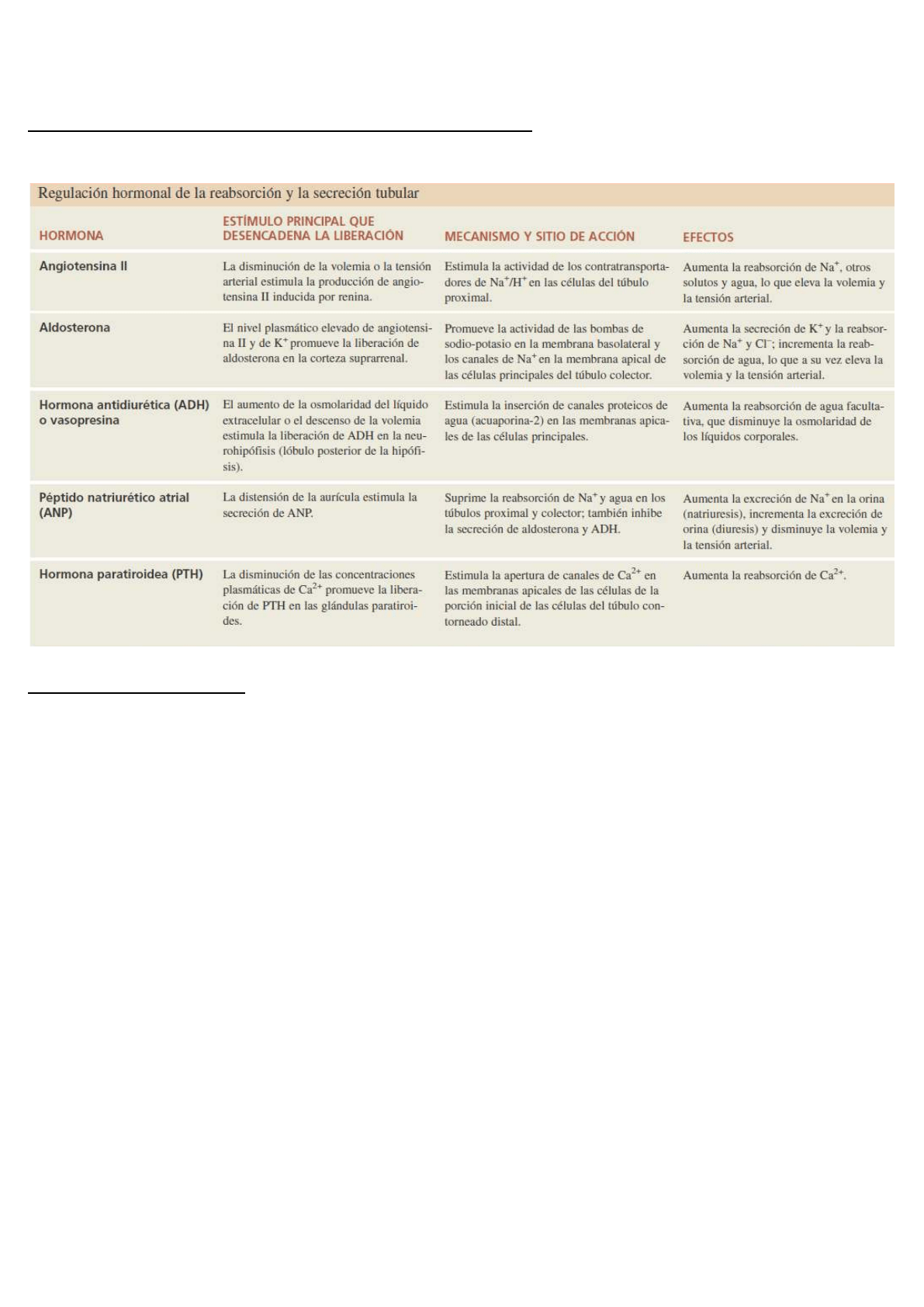
Regulación hormonal de la reabsorción y la secreción tubular
Cinco hormonas afectan la cantidad de Na+, Cl– , Ca2+ y agua reabsorbidos, y también la cantidad de K+
secretado en los túbulos renales:
Formación de orina diluida
Cuando se forma orina diluida, la osmolaridad del líquido en la luz tubular aumenta a medida que fluye a través
de la rama descendente del asa de Henle, vuelve a disminuir en su trayectoria por la rama ascendente y se
reduce aún más, cuando fluye a través del resto de la nefrona y el túbulo colector. Estos cambios en la
osmolaridad son el resultado de los siguientes mecanismos, a lo largo de la trayectoria del líquido tubular:
1. Como la osmolaridad del líquido intersticial de la médula renal aumenta en forma progresiva, se
reabsorbe cada vez más agua por ósmosis, a medida que el líquido tubular fluye a lo largo de la rama
descendente del asa de Henle hacia la punta del asa. Como consecuencia, el líquido que queda en la
luz se concentra cada vez más.
2. Las células que revisten la rama ascendente gruesa del asa de Henle poseen cotransportadores que
reabsorben en forma activa Na+, K+ y Cl– del líquido tubular (véase la Figura 26.15). Los iones pasan
del líquido tubular a las células de la rama ascendente gruesa, luego al líquido intersticial y por último
parte de ellos difunden a la sangre por los vasos rectos.
3. Aunque los solutos se reabsorben en la rama ascendente gruesa del asa de Henle, la permeabilidad al
agua de este sector de la nefrona siempre es baja, de manera que el agua no puede pasar por ósmosis.
A medida que los solutos, pero no las moléculas de agua, abandonan el líquido tubular, su osmolaridad
desciende hasta alrededor de 150 mOsm/L. En consecuencia, el líquido que entra en el túbulo
contorneado distal está más diluido que el plasma.
4. Mientras el líquido fluye a lo largo del túbulo contorneado distal, se reabsorben más solutos y sólo
pocas moléculas de agua. Las células de la porción inicial del túbulo contorneado distal no son muy
permeables al agua y no están reguladas por la ADH.

5. Por último, las células principales de la porción distal de los túbulos colectores son impermeables al
agua, cuando el nivel de ADH es muy bajo. Por ende, el líquido tubular se diluye cada vez más, a
medida que circula por los túbulos. Cuando el líquido tubular llega a la pelvis renal, su concentración
puede haber descendido hasta 65-70 mOsm/L, es decir, que puede ser hasta cuatro veces más diluido
que el plasma o el filtrado glomerular.
Formación de orina concentrada
Cuando la ingestión de agua disminuye o su pérdida es elevada (como durante la sudoración intensa), los
riñones deben conservar agua mientras eliminan desechos y el exceso de iones. Bajo la influencia de la ADH,
los riñones producen un pequeño volumen de orina muy concentrada. La orina puede ser cuatro veces más
concentrada (hasta 1200 mOsm/L) que el plasma o el filtrado glomerular (300 mOsm/L).La capacidad de la
ADH para excretar orina concentrada depende de la presión de un gradiente osmótico de solutos en el líquido
intersticial de la médula renal.
Multiplicación por contracorriente
La multiplicación por contracorriente es el proceso por medio del cual se crea un gradiente osmótico creciente
en forma progresiva en el líquido intersticial de la médula renal, como consecuencia del flujo de contracorriente.
Esta multiplicación requiere las asas de Henle largas de las nefronas yuxtaglomerulares.
La producción de orina concentrada se produce de la siguiente manera:
1. Los cotransportadores en las células de la rama ascendente gruesa del asa de Henle promueven la
acumulación de Na+ y Cl– en la médula renal. En la rama ascendente gruesa del asa de Henle, los
cotransportadores de Na+-K+-2Cl– reabsorben Na+ y Cl– del líquido tubular
2. El flujo de contracorriente, a través de las ramas descendente y ascendente del asa de Henle, establece
un gradiente osmótico en la médula renal. Como el líquido tubular se mueve en forma continua desde la
rama descendente a la rama ascendente gruesa del asa de Henle, la rama ascendente gruesa
reabsorbe Na+ y Cl– de manera constante. En consecuencia, el Na+ y el Cl– reabsorbidos se
concentran de manera progresiva en el líquido intersticial de la médula renal, y se crea un gradiente
osmótico que oscila entre 300 mOsm/L en la porción externa de la médula y 120 mOsm/L en la
profundidad de la porción interna de la médula. El líquido tubular se concentra cada vez más, a medida
que fluye a lo largo de la rama descendente y se diluye en forma progresiva, cuando atraviesa la rama
ascendente.
3. Las células, en los túbulos colectores, reabsorben más agua y urea. Cuando la ADH aumenta la
permeabilidad al agua de las células principales, el agua se desplaza con rapidez por ósmosis, fuera el
líquido intersticial del túbulo colector hacia el líquido intersticial de la porción interna de la médula y
luego, hacia los vasos rectos.
4. El reciclado de urea promueve su acumulación en la médula renal. A medida que se acumula urea en el
líquido intersticial, parte de ella difunde hacia el líquido tubular en las ramas descendente y ascendente
delgada de las asas de Henle largas, que también son permeables a la urea. A medida que el líquido
fluye a través de los túbulos colectores, continúa la reabsorción de agua por ósmosis, por la presencia
de ADH. Esta reabsorción de agua incrementa aún más la concentración de urea en el líquido tubular,
difunde más urea hacia el líquido intersticial de la porción interna de la médula renal, y el ciclo se repite.
La transferencia constante de urea entre los segmentos del túbulo renal y el líquido intersticial de la
médula se denomina reciclado de la urea. Como resultado, los solutos que quedan en la luz se
concentran en forma significativa y se excreta un pequeño volumen de orina concentrada.

Intercambio por contracorriente
El intercambio por contracorriente es el proceso por medio del cual los solutos y el agua se intercambian, en
forma pasiva, entre la sangre de los vasos rectos y el líquido intersticial de la médula renal, como consecuencia
del flujo de contracorriente.
Al igual que el líquido tubular, que fluye en direcciones opuestas en el asa de Henle, la sangre fluye en
direcciones contrarias en las porciones ascendentes y descendentes de los vasos rectos. Como el flujo de
contracorriente entre las zonas ascendente y descendente de los vasos rectos permite el intercambio de
solutos y agua entre la sangre y el líquido intersticial de la médula renal, se considera que los vasos rectos
funcionan como intercambiadores de contracorriente. A medida que fluye a lo largo de la porción descendente
hacia la médula renal, donde el líquido intersticial se concentra cada vez más, los iones de Na+ y Cl– y la urea
difunden desde el líquido intersticial hacia la sangre, y el agua difunde desde la sangre hacia el líquido
intersticial. No obstante, una vez que aumenta la osmolaridad, la sangre fluye hacia la porción ascendente de
los vasos rectos. En esta región, la sangre fluye a través de un área donde el líquido intersticial está cada vez
menos concentrado. Como consecuencia, los iones de Na+, Cl– y urea difunden desde la sangre hacia el
líquido intersticial, y el agua regresa a los vasos rectos desde el líquido intersticial.
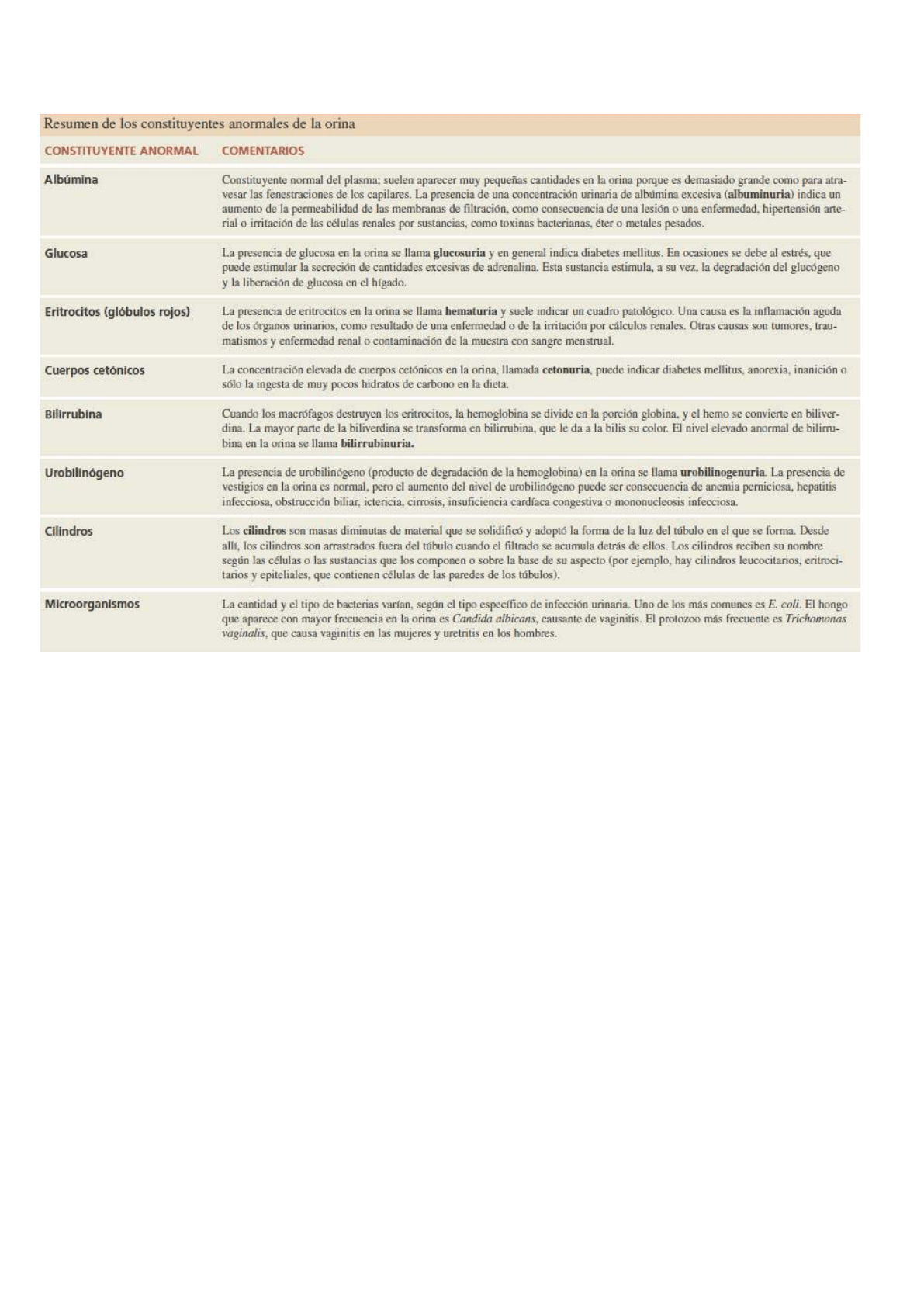
CARACTERISTICAS DE ORINA NORMAL:
TRANSPORTE, ALMACENAMIENTO Y ELIMINACIÓN DE LA ORINA
➤ URETERES
Cada uréter conduce orina desde la pelvis renal hasta la vejiga. Las contracciones peristálticas de las paredes
musculares de los uréteres impulsan la orina hacia la vejiga, y también la presión hidrostática y la gravedad.
Las ondas peristálticas que pasan desde la pelvis renal a la vejiga tienen una frecuencia que oscila entre uno y
cinco por minuto, según la rapidez con que se forma la orina. Los uréteres miden entre 25 y 30 cm de largo
(10-12 pulgadas), sus paredes son gruesas y su diámetro es pequeño; fluctúa entre 1 y 10 mm a lo largo de su
trayectoria entre la pelvis renal y la vejiga.
Al igual que los riñones, los uréteres son retroperitoneales. En la base de la vejiga, los uréteres giran en sentido
medial y adoptan una dirección oblicua, a través de la pared vesical posterior. Aunque no existe una válvula
anatómica en la desembocadura de cada uréter, en la vejiga hay una válvula fisiológica eficaz. A medida que la
vejiga se llena de orina, la presión en su interior comprime los orificios oblicuos de los uréteres e impide el
reflujo de orina.
La pared de los uréteres está formada por tres capas de tejido.
❖ La capa más profunda, la mucosa, es una membrana compuesta por epitelio de transición, y la lámina
propia subyacente e tejido conectivo areolar presenta una cantidad considerable de colágeno, fibras
elásticas y tejido linfático. El epitelio de transición puede distenderse, lo que representa una ventaja
notable para cualquier órgano que deba adaptarse a un volumen cambiante de líquido. El moco secretado
por las células caliciformes de la mucosa evita que las células tomen contacto con la orina, cuya
concentración de solutos y pH puede variar en forma significativa con respecto al citosol de las células que
forman las paredes de los uréteres.
❖ La capa intermedia, la muscular, está compuesta por capas longitudinal interna y circular externa de fibras
musculares lisas. Esta disposición es inversa a la del tubo digestivo, que contiene una capa circular interna
y una longitudinal externa. La muscular consta de una capa longitudinal interna, una circular media y una
longitudinal externa. La peristalsis es la función principal de la túnica muscular.
❖ La cubierta superficial de los uréteres es la adventicia, una capa de tejido conectivo que contiene vasos
sanguíneos, vasos linfáticos y nervios destinados a la muscular y la mucosa. La adventicia se mezcla con
el tejido conectivo circundante y fija los uréteres en su posición.
➤ VEJIGA
La vejiga es un órgano muscular hueco y distensible, situado en la cavidad pelviana, por detrás de la sínfisis
del pubis. En los hombres, se encuentra por delante del recto y en la mujer es anterior a la vagina e inferior al
útero.
Los repliegues peritoneales mantienen la vejiga en su posición. Cuando se distiende un poco por
la acumulación de orina, adopta una forma esférica. Cuando está vacía, se colapsa. A medida que el volumen
de orina aumenta, toma forma de pera y asciende a la cavidad abdominal. La capacidad de la vejiga oscila en
promedio entre 700 y 800 mL. Es más pequeña en las mujeres porque el útero ocupa el espacio por encima de
la vejiga.
Anatomía e histología de la vejiga
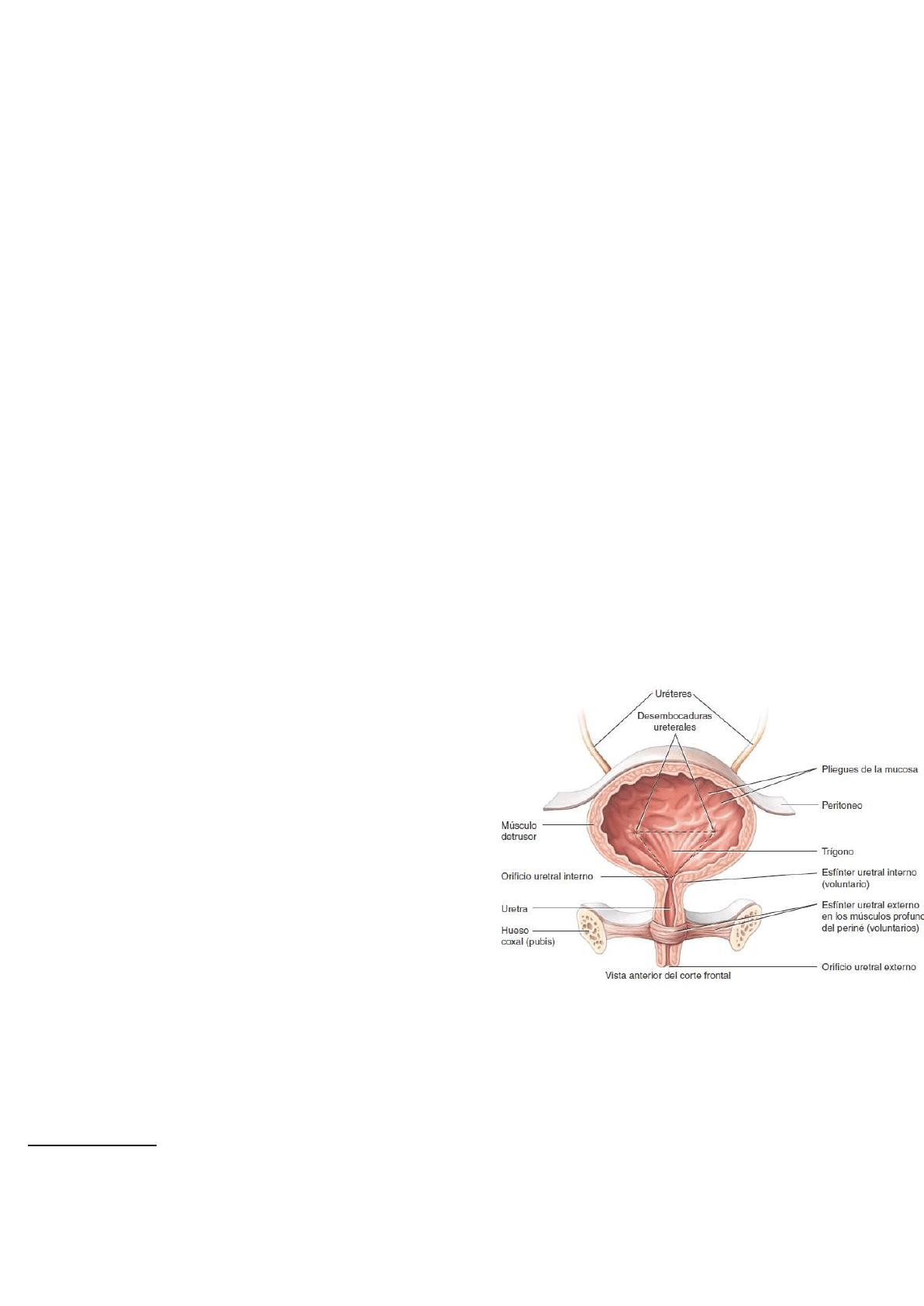
En el piso de la vejiga, se encuentra un área triangular pequeña denominada trígono (triángulo). Los dos
ángulos posteriores del trígono contienen los dos orificios ureterales. La desembocadura en la
uretra, que es el orificio uretral interno, se encuentra en el ángulo
anterior (Figura 26.21). Como la mucosa está adherida con
firmeza a
la capa muscular, el trígono tiene aspecto liso.
Tres capas forman la pared de la vejiga.
★ La más profunda es la mucosa, una membrana
compuesta por epitelio de transición y una lámina
propia subyacente, similar a la de los uréteres. También
hay pliegues que permiten la expansión de la vejiga.
★ Alrededor de la mucosa, se encuentra la túnica
muscular, también llamada músculo detrusor (detrusor,
que impulsa), formada por tres capas de fibras
musculares lisas: la longitudinal interna, la circular
media y la longitudinal externa. Alrededor del orificio
uretral, las fibras circulares forman el esfínter uretral
interno, y más abajo se encuentra el esfínter uretral
externo, constituido por músculo esquelético, que se
considera una modificación de los músculos profundos del periné.
★ La capa más superficial de la vejiga en las paredes posterior e inferior es la adventicia, una capa de tejido
conectivo areolar continuo con la de los uréteres. En la región superior de la vejiga está la serosa, que es
una capa de peritoneo visceral.
Reflejo miccional
La micción se produce debido a una combinación de contracciones musculares voluntarias e involuntarias.
Cuando el volumen de orina en la vejiga excede los 200-400 mL, la presión en su interior aumenta en forma
Este documento contiene más páginas...
Descargar Completo
sistema urinario- fisio (1).pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.