
ROSARIO PEREZ CONTRERAS
1
SISTEMA TEGUMENTARIO
Generalidades
La piel (cutis, tegumento) y sus derivados constituyen el sistema tegumentario. La piel forma la cubierta
externa del cuerpo y es su órgano más grande, ya que constituye del 15 % al 20 % de su masa total. La piel
consta de dos estratos principales:
Epidermis: compuesta por epitelio plano estratificado cornificado. Deriva del ectodermo. Crece
continuamente pero mantiene su espesor.
Dermis: conectivo denso. Deriva del mesodermo. Da sostén mecánico, resistencia y espesor a la
piel.
La hipodermis contiene tejido adiposo organizado en lobulillos separados por tabiques de tejido conectivo.
Se encuentra más profundidad que la dermis, se la conoce como fascia subcutánea.
Los derivados epidérmicos de la piel (anexos cutáneos) son: folículos pilosos y pelo, glándulas
sudoríparas, glándulas sebáceas, uñas y glándulas mamarias.
El sistema tegumentario cumple funciones esenciales relacionadas con su ubicación
en la superficie externa.
Las principales funciones de la piel son:
Barrera, protege contra agentes físicos, químicos y biológicos del medio externo.
Provee información inmunitaria obtenida durante el procesamiento de antígenos a las células
efectoras adecuadas del tejido linfático.
Participa en la homeostasis regulando temperatura corporal y la perdida de agua.
Transmite información sensitiva acerca del medio externo al sistema nervioso.
Desempeña funciones endocrinas mediante la secreción de hormonas.
Interviene en la excreción por la secreción exocrina de las glándulas sudoríparas, sebáceas y
apocrinas.
La piel se clasifica en fina y gruesa, un reflejo de su espesor y su ubicación.
El espesor de la piel varía de 1 a 5mm. La piel gruesa tiene una capa epidérmica mucho más gruesa y sin
pelos, está en las palmas de las manos y las plantas de los pies. La piel fina tiene una epidermis más
delgada y folículos pilosos. Esta clasificación se refiere solo al espesor de la capa epidérmica.
ROSARIO PEREZ CONTRERAS
2
ESTRATOS DE LA PIEL
Epidermis
Está compuesta por un epitelio plano estratificado queratinizado en el que pueden identificarse 4
estratos. En el caso de la piel gruesa hay un quinto estrato.
Estrato basal: llamado también estrato germinativo por la presencia de células con actividad
mitótica, que son las células madre de la epidermis
Estrato espinoso: llamado así por las proyecciones cortas que van de una célula a otra.
Estrato granuloso: sus células contienen gránulos abundantes.
Estrato lucido: solo en la piel gruesa y considerado una subdivisión del estrato corneo.
Estrato corneo: son células queratinizadas.
La diferenciación de células epiteliales constituye una forma especializada de
apoptosis.
La diferenciación comienza en el estrato basal, las células del estrato granuloso tienen la típica morfología
nuclear apoptotica. Sin embargo, la fragmentación celular no se produce, en cambio, las células se llenan
de filamentos de la proteína intracelular queratina y mas tarde se descaman de la superficie cutánea.
El estrato basal tiene a su cargo la renovación de las células epidérmicas.
El estrato basal consiste en una capa celular de una sola célula de espesor que se apoya en la lámina basal.
Las células son las células madre que originan nuevos queratinocitos, son pequeñas y cubicas. El
citoplasma basófilo y los núcleos muy juntos le dan una basofilia marcada a este estrato. Las células
contienen cantidades variables de melanina y también muchas uniones celulares (con la lámina basal y
células contiguas).
A medida que surgen por división mitótica en este estrato, los nuevos queratinocitos se trasladan al
siguiente estrato para comenzar, así, su proceso de migración hacia la superficie. Este proceso termina
cuando la célula se convierte en una célula queratinizada madura, que finalmente se descama en la
superficie de la piel.
Las células del estrato espinoso exhiben proyecciones “espinosas” características.
Este estrato tiene varias células de espesor. Los queratinocitos en esta capa son más grandes que los del
estrato basal y tienen varias evaginaciones o espinas citoplasmáticas que le dan el nombre al estrato. Las
evaginaciones están unidas a otras semejantes de las células contiguas por medio de desmosomas. En el
microscopio el sitio donde está el desmosoma aparece como un engrosamiento leve llamado nodo de
Bizzozero.
Las células del estrato granuloso contienen gránulos de queratohialina conspicuos.
Este estrato es la capa más superficial de la porción no queratinizada de la epidermis. Tiene de una a tres
células de espesor. Los queratinocitos en este estrato contienen muchos gránulos de queratohialina. Estos
gránulos contienen proteínas con cistina e histidina, las cuales son las precursoras de la proteína filagrina,
que aglomera los filamentos de queratina que se hallan dentro de las células cornificadas del estrato
corneo.
ROSARIO PEREZ CONTRERAS
3
El estrato corneo consiste en células escamosas anucleadas repletas de filamentos de
queratina.
Hay una transición brusca del estrato granuloso al estrato corneo. Las células del estrato corneo son las
más diferenciadas de la epidermis. Pierden su núcleo y orgánulos citoplasmáticos y se llenan casi por
completo con filamentos de queratina.
En la porción más profunda de este estrato, la gruesa membrana plasmática de estas células está cubierta
por fuera con una capa extracelular de lípidos que forman parte de la barrera contra el agua en la
epidermis.
El estrato corneo es la capa de espesor más variable y es la de mayor grosor en la piel gruesa.
El estrato lucido solo se observa bien en la piel gruesa. Suele presentar un aspecto refractil y se tiñe poco.
Contiene células eosinofilas en las que el proceso de corinificacion está muy avanzado.
Dermis
La adherencia de la epidermis a la dermis está potenciada por un aumento de la
interfaz entre los tejidos.
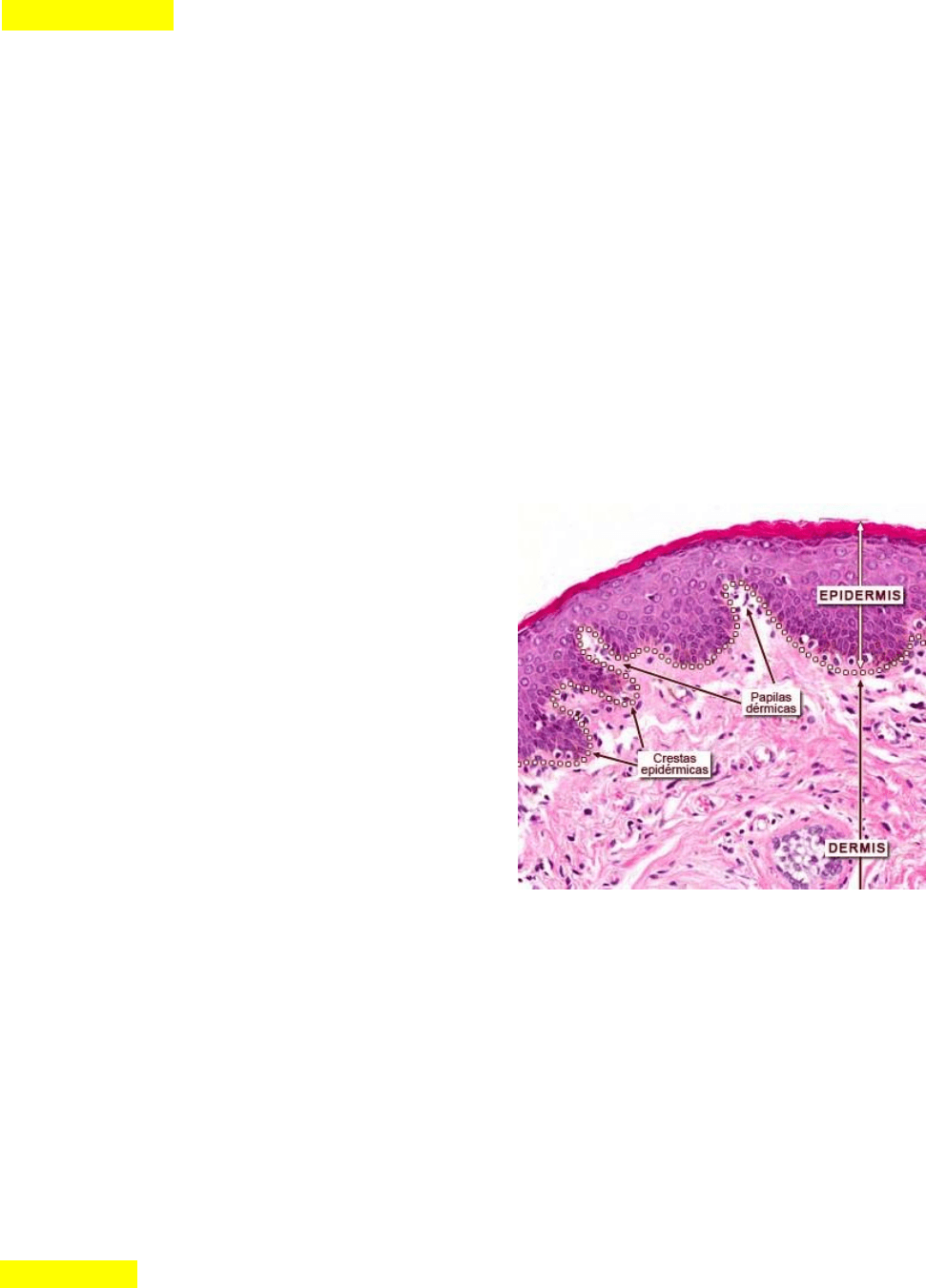
La unión entre la epidermis y la dermis (unión
dermoepidérmica) exhibe un contorno muy irregular,
excepto en la piel más fina. Los cortes de piel
perpendiculares a la superficie permiten observar
abundantes evaginaciones digitiformes del tejido
conjuntivo, llamadas papilas dérmicas, que se extienden
hacia la superficie profunda de la epidermis. Las papilas
se complementan con lo que parecen ser
protuberancias similares a la epidermis, llamadas
crestas epidérmicas o crestas interpapilares, que se
hunden en la dermis.
En la piel gruesa hay crestas dérmicas verdaderas, además de l as papilas dérmicas.
Estas crestas forman un patrón distintivo que es genéticamente singular en cada individuo y se refleja en la
aparición de surcos y pliegues epidérmicos que se ven en la superficie cutánea. Las crestas y papilas
dérmicas son muy prominentes en la piel gruesa de las superficies palmares y plantares.
Aquí la superficie basal de la epidermis supera ampliamente la de su superficie libre. Por lo tanto, el estrato
germinativo está extendido sobre una gran superficie; si se supone que su ritmo de mitosis es casi
constante, entran más células por unidad de tiempo en el estrato córneo de la piel gruesa que en el de la
piel fina. Se cree que estas células adicionales son la causa del espesor mayor del estrato córneo en la piel
gruesa.
La dermis está compuesta por dos capas: la dermis papilar y la dermis reticular.
La dermis papilar la capa más superficial y más delgada, es tejido conectivo laxo. Las fibras colágenas no
son tan gruesas y son sobre todo colágeno tipo I y III. Las fibras elásticas son filiformes y se organizan en

ROSARIO PEREZ CONTRERAS
4
una red irregular. Hay vasos sanguíneos que irrigan la epidermis pero no entran en ella. También hay
nervios pueden introducirse en la epidermis.
La dermis reticular es profunda con respecto a la dermis papilar. Es más gruesa y con menos células que
la dermis papilar. Tiene haces gruesos e irregulares de fibras de colágeno tipo I y fibras elásticas menos
delicadas. Puede haber células de musculo liso en la parte más profunda de esta dermis, por ejemplo en
areola, pene, escroto y periné.
Justo debajo de la dermis reticular pueden encontrarse capas de tejido adiposo,
músculo liso y, en algunos sitios, músculo estriado.
El panículo adiposo es una capa de tejido adiposo de espesor variable, que se ubica en un plano más
profundo que la dermis reticular. Esta capa es un sitio de almacenamiento de energía y funciona como
aislante. Esta capa junto con su tejido conjuntivo laxo asociado constituyen la hipodermis o fascia
subcutánea.
Las células musculares lisas, que se originan en esta capa forman los músculos erectores del pelo que
conectan la parte profunda de los folículos pilosos con la dermis más superficial. La contracción de estos
músculos en los seres humanos produce la erección de los pelos y el fruncimiento en la piel llamada,
conocido como “piel de gallina”.
CÉLULAS DE LA EPIDERMIS
Hay cuatro tipos de células en la epidermis:
Queratinocitos 85% de las células, separan el
organismo del medioambiente
Melanocitos 5% de las células, productoras de
pigmento.
Células de Langerhans 2 a 15%, participan en
respuesta inmunitaria
Células de Merkel 6 a 10%, asociadas a
terminaciones nerviosas sensitivas.
Queratinocitos (
Se originan en el estrato epidérmico basal. Al abandonar este estrato pasan a cumplir dos actividades
esenciales:
Producen queratinas que forman filamentos intermedios que constituyen el 85% de los
queratinocitos.
Participan en la formación de la barrera epidérmica contra el agua.
El citoplasma de los queratinocitos inmaduros aparece basófilo por la gran cantidad de ribosomas que
participan en la síntesis de queratina. A medida que las células avanzan al estrato espinoso la síntesis de
filamentos de queratina continua y estos se agrupan en haces lo bastante gruesos, llamados tonofibrillas,
el citoplasma se torna eosinofilo.
ROSARIO PEREZ CONTRERAS
5
Los gránulos de queratohialina contienen proteínas asociadas con los filamentos
intermedios, que contribuyen a la aglomeración de los filamentos de queratina.
En la parte superior del estrato espinoso, los ribosomas libres dentro de los queratinocitos comienzan a
sintetizar gránulos de queratohialina. Los gránulos contienen filagrina y tricohialina. A medida que la
cantidad de gránulos aumenta, su contenido se libera en el citoplasma de los queratinocitos. La filagrina y
la tricohialina funcionan como promotoras de la aglomeración de los filamentos de queratina en
tonofibrillas, lo cual inicia la conversión de células granulares en las células cornificadas. Este proceso se
denomina cornificación y se produce en 2 h a 6 h. Las fibrillas de queratina que se forman en este proceso
son de queratina blanda, a diferencia de la queratina dura del cabello y de las uñas.
La transformación de una célula granulosa en una queratinizada, también comprende la desintegración del
núcleo y otros orgánulos y el engrosamiento de la membrana plasmática. Esto se acompaña de un cambio
en el pH.
La descamación de los querantocitos superficiales del estrato córneo es regulada por
la degradación proteolítica de los desmosomas de las células.
La exfoliación continua de los queratinocitos superficiales es un proceso proteolítico regulado que
comprende la degradación de los desmosomas de las células. Las serina peptidasas causan la escisión
desmosómica en una manera dependiente de pH.
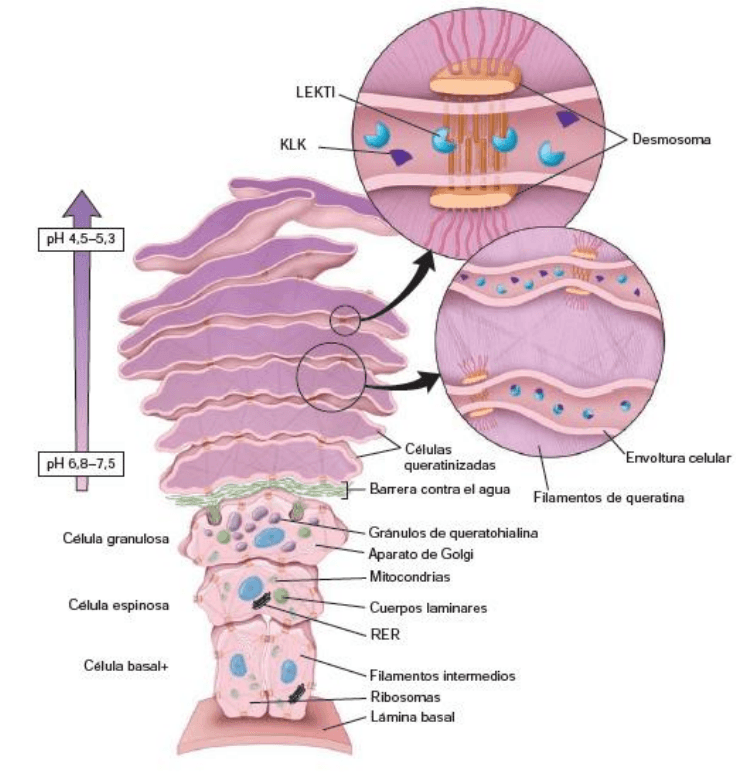
Los cuerpos laminares contribuyen a la formación de la barrera epidérmica
intercelular contra el agua
Una barrera epidérmica contra el agua es esencial para los epitelios “secos” de los mamíferos y es
responsable de mantener la homeostasis corporal. La barrera se establece principalmente por dos factores
en los queratinocitos: 1) el depósito de proteínas insolubles en la superficie interna de la membrana
plasmática y 2) una capa de lípidos que se adhiere a la superficie externa de la membrana plasmática.
Los queratinocitos en el estrato espinoso producen unas vesículas limitadas por membrana que reciben el
nombre de cuerpos laminares. Las células espinosas y granulares sintetizan una mezcla heterogénea de
lípidos probarrera y sus respectivas enzimas procesadoras de lípidos. Esta mezcla pasa al interior de los
cuerpos laminares que se forman en el Golgi. El contenido de los gránulos se secreta por exocitosis hacia el
espacio intercelular entre el estrato corneo y el estrato granuloso. Así la barrera epidérmica contra el agua
se compone de dos elementos estructurales:
La envoltura celular es una capa de proteínas insolubles de 15 nm de espesor, depositada sobre la
superficie interna de la membrana plasmática que contribuye a las propiedades mecánicas de
resistencia de la barrera.
La envoltura lipídica 5nm de espesor son lípidos adheridos a la membrana celular por enlaces éster.
Los componentes principales de los lípidos de la envoltura lipídica son: ceramidas, colesterol, ácidos
grasos libres y acilglucosilceramida.
A medida que las células continúan desplazándose hacia la superficie libre, la barrera se mantiene
constantemente por los queratinocitos que entran en el proceso de diferenciación terminal.
ROSARIO PEREZ CONTRERAS
6
La epidermis está en un estado de equilibrio dinámico, en el que las células
queratinizadas exfoliadas se reemplazan en forma constante, por un flujo continuo
de células terminalmente diferenciadas.
El reemplazo de las células epidérmicas se mantiene por varios procesos que comprenden:
• La mitosis de las células basales en el estrato basal,
• La diferenciación y la muerte celular programada, conforme las células ascienden hacia el estrato córneo
• La pérdida de células por la exfoliación de la superficie cutánea.
Para mantener este equilibrio, cada célula individual en la epidermis tiene una cantidad predeterminada
de tiempo para realizar funciones específicas. El tiempo total de rotación epidérmica es de 47 días.
Melanocitos
Los melanocitos derivan de células de la cresta neural y están dispersos entre las
células del estrato basal.
Los melanocitos establecen unidades melanoepidermicas, en las que un melanocito se mantiene asociado
con una cantidad dada de queratinocitos. Un melanocito está asociado con cerca de 36 queratinocitos,
ROSARIO PEREZ CONTRERAS
7
pero puede variar de 1:4 a 1:40. Esta relación es constante en todos los grupos étnicos, pero esta
influenciada por la edad y los factores ambientales.
El melanocito epidérmico se encuentra entre las células basales del estrato basal, emiten evaginaciones
largas. Ni las evaginaciones ni el cuerpo celular establecen uniones desmosomicas con los queratinocitos
vecinos.
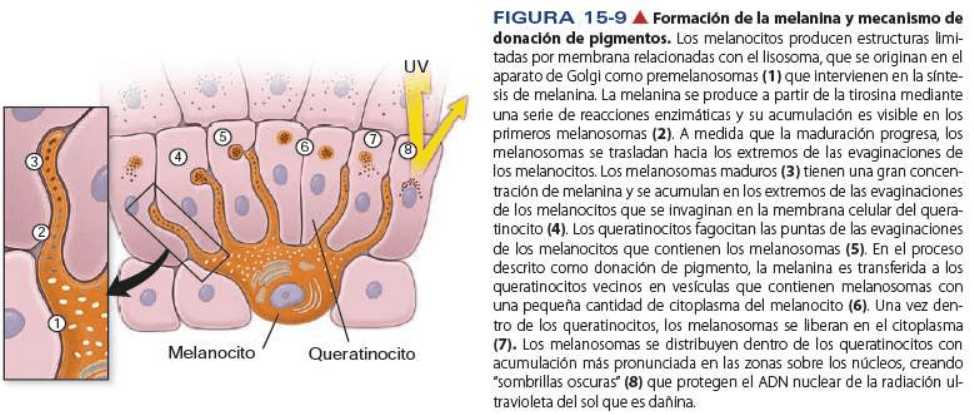
Los melanocitos producen melanina y la distribuyen a los queratinocitos
La función de la melanina es proteger al organismo contra los efectos dañinos de la radiación ultravioleta
no inoizante. La melanina es producida por la oxidación de la tirosina, su síntesis es regulada por la acción
de la hormona melanocito estimulante.
Los premelanosomas y melanosomas tempranos, que tienen poca melanina presentan una estructura
interna ordenada. A medida que se forma más melanina por oxidación de la tirosina, la estructura interna
del premelanosoma se va ocultando hasta que se forma el granulo de melanina maduro, el melanosoma.
Los melanosomas en desarrollo y su contenido de melanina se transfieren a los queratinocitos vecinos por
donación pigmentaria. Este proceso, que consiste en la fagocitosis del extremo de la prolongación
melanocítica por los queratinocitos, es un tipo de secreción citocrina porque también se fagocita una
pequeña cantidad del citoplasma que rodea el melanosoma.
Los melanosomas y su contenido se degradan en el proceso de macroautofagia. En las personas de piel
oscura la melanina se degrada más lentamente, mientras que en las de piel más clara lo hace más
rápidamente.
Células de Langerhans
Las células de Langerhans son células presentadoras de antígenos de la epidermis.
Son células de aspecto dendrítico que se localizan en la epidermis. Se originan a partir de citoblastos
linfoides multipotenciales en la medula ósea. Captan y presentan antígenos que entran a través de la piel,
por tanto forman parte del sistema fagocitico mononuclear. Una vez que el antígeno es fagocitado y
procesado, la célula migra al ganglio linfático regional donde interacciona con linfocitos T.

ROSARIO PEREZ CONTRERAS
8
El núcleo presenta indentaciones o escotaduras en muchos sitios. Además posee gránulos de Birbeck, con
su forma característica de raqueta de tenis. Como es presentadora de antígenos tiene moléculas MHC I y II.
Las células de Langerhans no se pueden distinguir con HE y al igual que los melanocitos no establecen
uniones desmosomicas con los queratinocitos.
Células de Merkel
Las células de Merkel son células epidérmicas que intervienen en la percepción
sensorial cutánea.
Son células dendríticas del estrato basal. Su origen es desconocido, tienen marcadores antigénicos de tipo
epidérmico y nervioso. Son abundantes en la piel en donde la percepción sensible es aguda, como en la
yema de los dedos. A diferencia de los melanocitos y células de Langerhans, están unidas a los
queratinocitos contiguos a través de desmosomas y contienen filamentos intermedios en su citoplasma.
Su núcleo es lobulado y pueden contener algunos melanosomas en su citoplasma, pero se caracterizan
mejor por la presencia de gránulos de neurosecrecion de centro denso de 80nm.
Las células de Merkel están estrechamente asociadas con los bulbos terminales expandidos de las fibras
nerviosas mielinicas aferentes. La terminación nerviosa que llega a la célula de Merkel pierde su cubierta
de células de Schwann y perfora la lámina basal, donde se expande en una estructura en forma de placa
llamada disco receptor, que se encuentra en estrecho contacto con la base de la célula de Merkel.
La combinación de la fibra nerviosa y la célula epidérmica, llamada corpúsculo de Merkel, forma un
mecanoreceptor sensorial.
ESTRUCTURAS DE LA PIEL
Inervación
La piel está dotada de receptores sensoriales de diversos tipos que son terminaciones periféricas de
nervios sensitivos. También está bien inervada con terminaciones nerviosas motoras para los vasos
sanguíneos, los músculos erectores del pelo y las glándulas sudoríparas.
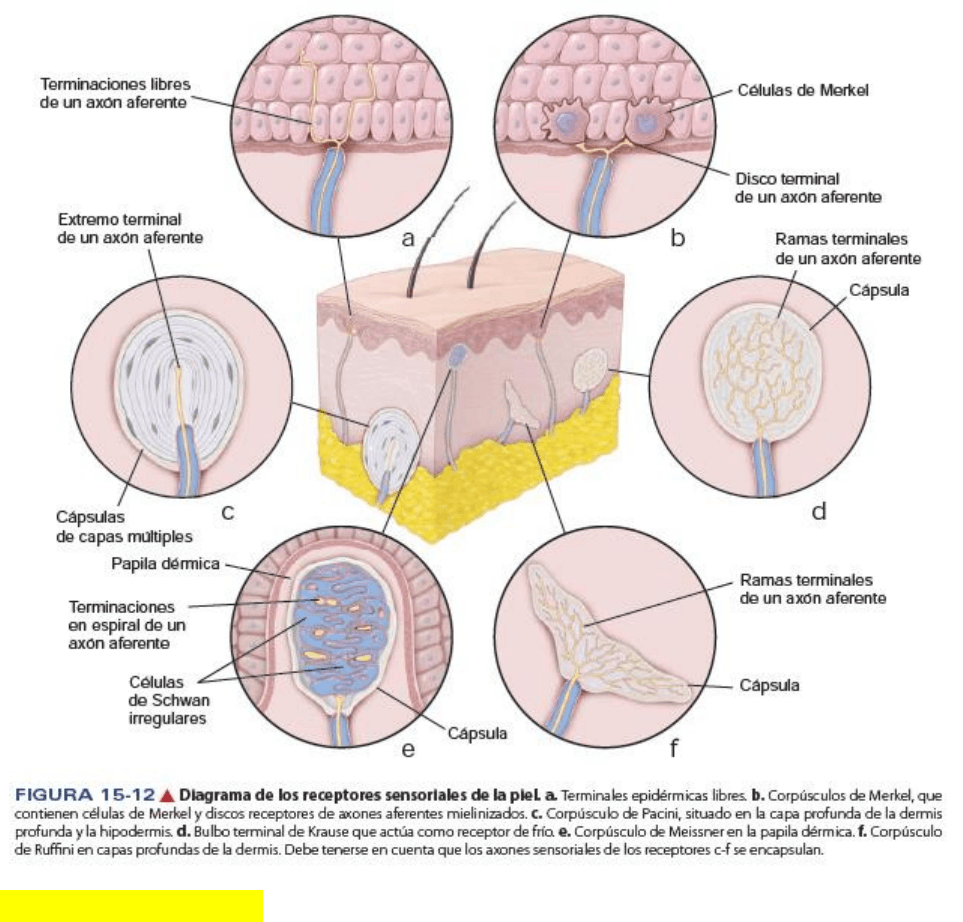
Las terminaciones nerviosas libres son los receptores neuronales más abundantes de
la epidermis
Las terminaciones nerviosas libres en la epidermis finalizan en el estrato granuloso. Son “libres” porque
carecen de cubierta de tejido conectivo o de células de Schwann. Además, tienen modalidades sensoriales
múltiples como: tacto fino, calor, frio y dolor. Estas terminaciones rodean también la mayor parte de los
folículos pilosos y se fijan en su vaina radicular externa.
Otras terminaciones nerviosas de la piel están encerradas en una capsula de tejido conjuntivo. Entre las
terminaciones nerviosas encapsuladas se encuentran:
Corpúsculos de Pacini: detectan cambios de presión y las vibraciones aplicadas a la superficie cutánea.
Corpúsculos de Meissner: perciben las sensaciones táctiles leves
Corpúsculos de Ruffini: sensibles al estiramiento y tensión de la piel
ROSARIO PEREZ CONTRERAS
9
.
Los corpúsculos de Pacini son presoreceptores profundos que captan presiones
mecánicas y vibratorias.
Estos corpúsculos son estructuras ovoides grandes que se encuentran en la dermis e hipodermis (sobre
todo en la yema de los dedos), en el tejido conectivo en general y en asociación con las articulaciones, el
periostio y las vísceras. Miden más de 1mm.
Están compuestos por una terminación nerviosa mielinica rodeada de una estructura capsular. La fibra
nerviosa perfora la cápsula en un polo con su vaina de mielina intacta. La mielina es retenida por uno o dos
nódulos y después desaparece. La porción amielínica del axón está cubierta por una serie de láminas muy
juntas de células de Schwann aplanadas que forman el núcleo interno del corpúsculo.
El núcleo externo está formado por una serie de láminas concéntricas separadas cada una por un espacio
estrecho que contiene líquido semejante a la linfa. Cada lámina está compuesta por células aplanadas.
Los corpúsculos de Pacini responden a la presión y a la vibración a través del desplazamiento de las
láminas capsulares. Esto provoca la despolarización efectiva del axón.

ROSARIO PEREZ CONTRERAS
10
Los corpúsculos de Meissner están situados en las papilas dérmicas y funcionan como
receptores de tacto
Responden en particular a los estímulos de baja frecuencia en la dermis papilar de la piel lampiña (labios,
superficies palmares, etc.). Son cilindros de extremos adelgazados, ubicados en las pailas dérmicas justo
debajo de la lámina basal epidérmica.
En estos receptores, una o dos terminaciones amielínicas de fibras nerviosas mielinicas describen trayectos
en espiral dentro del corpúsculo. El componente celular consiste en células de Schwann aplanadas que
forman varias láminas irregulares entre las cuales transcurren los axones hasta el polo del corpúsculo.
Los corpúsculos de Ruffini responden al desplazamiento mecánico de las fibras
colágenas adyacentes.
Son los mecanoreceptores encapsulados más simples. Tienen forma alargada fusiforme y miden 1 a 2
micras de longitud. Consisten en una delgada capsula de conectivo que encierra un espacio lleno de
líquido. Las fibras colágenas del tejido conectivo circundante atraviesan la capsula. El elemento nervioso
consiste en una sola fibra mielinica que perfora la capsula, pierde su vaina de mielina y se ramifica para
formar una arborización densa de terminaciones axonicas delgadas, que finalizan en una pequeña
dilatación bulbosa.
Responden al desplazamiento de las fibras colágenas inducido por la tensión mecánica sostenida, por lo
tanto responden al estiramiento o torsión. Pertenecen a la familia de los receptores de adaptación rápida
que generan potenciales de acción breves al principio y al final del estímulo.
Anexos cutáneos
Derivan de brotes en profundidad del epitelio epidérmico durante el desarrollo embrionario. Comprende
las siguientes estructuras: folículos pilosos y su producto el pelo; glándulas sebáceas y el unto sebáceo;
glándulas sudoríparas ecrinas y el sudor; y glándulas sudoríparas apocrinas y su producto mixto que
consiste en una forma de sudor con una concentración elevada de hidratos de carbono, lípidos y proteínas.
Pelos y glándulas sudoríparas desempeñan funciones específicas en la regulación de la temperatura
corporal. Las glándulas sebáceas secretan una sustancia oleosa que puede tener funciones de protección.
Las gandulas apocrinas producen una secreción serosa que contiene feromonas que actúan como
sustancias de atracción sexual en animales.
Folículos pilosos y pelo
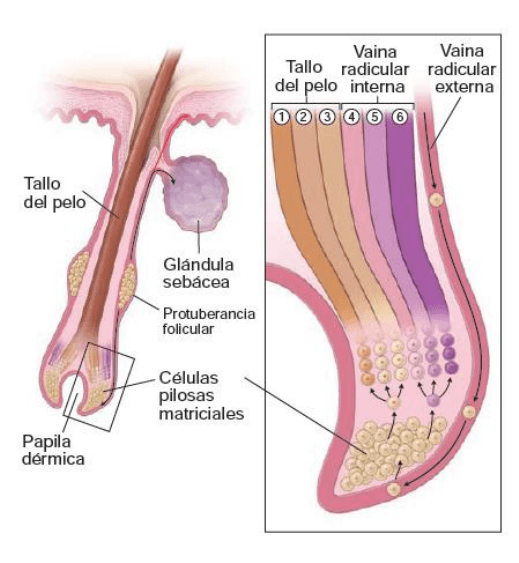
Cada folículo piloso es una invaginación de la epidermis en la que se forma un pelo
Los folículos y el pelo solo están ausentes en las palmas de las manos, las plantas de los pies, los labios y la
piel periorificial de los sistemas urinario y genital. La distribución del pelo esta muy influenciada por las
hormonas sexuales. El cabello se adelgaza con la edad debido a la menor secreción de estrógenos u
hormonas relacionadas.
ROSARIO PEREZ CONTRERAS
11
El folículo piloso se encarga de la producción y el crecimiento de un pelo. La coloración del pelo está dada
por el contenido y el tipo de melanina que posee. El aspecto histológico del folículo varía según esté en
fase de crecimiento o en fase de reposo. El folículo en crecimiento muestra una estructura más compleja,
que es la que se describe aquí. El folículo piloso se divide
en cuatro regiones:
Infundíbulo: desde el orificio superficial del
folículo hasta la altura del orificio de su glándula
sebácea. El infundíbulo es una parte del
conducto pilosebaceo.
Istmo: desde el infundíbulo hasta la altura de la
inserción del musculo erector del pelo.
Protuberancia folicular: sobresale del folículo
piloso cerca de la inserción del musculo erector y
contiene las células madre.
Segmento inferior: con un diámetro uniforme
salvo en la base donde se expande para formar
el bulbo. La base del bulbo se invagina por un
ovillo de conectivo laxo vascularizado llamado,
papila dérmica.
Las otras células que forman el bulbo reciben la
denominación colectiva de matriz del pelo, que consiste
en células matriciales. Estas células constituyen la población de células en división y diferenciación rápidas,
provenientes de la protuberancia folicular que contiene las células madre. La división y proliferación de
estas células son las responsables del crecimiento del pelo. Los melanocitos se encuentran dispersos en
este estrato germinativo. Proveen melanosomas a las células del pelo en desarrollo.
Las células matriciales se diferencian en las células productoras de queratina del pelo y en la vaina
radicular interna. Esta vaina es una cubierta celular multiestratificada que rodea la parte profunda del
pelo. La vaina radicular interna tiene 3 capas:
Capa de Henle: una sola capa exterior de células cubicas. En contacto directo con la parte más
externa del folículo piloso, que es una invaginación de la epidermis y recibe el nombre de vaina
radicular externa.
Capa de Huxley: capa simple o doble de células aplanadas que forman la placa intermedia de la
vaina radicular interna.
Cutícula de la vaina radicular interna: de células planas o escamosas que están en contacto con el
tallo del pelo.
Un nicho de células madre epidérmicas que se encuentra en la prominencia folicular
de la vaina radicular externa provee las células madre para el crecimiento del pelo.
Las terminaciones nerviosas rodean la vaina radicular externa a la altura de la inserción del musculo
erector. En esta región se halla la prominencia folicular, una aglomeración de células epiteliales
relativamente indiferenciadas. Estas células son las encargadas de proveer células madre para el

ROSARIO PEREZ CONTRERAS
12
crecimiento de los folículos pilosos, así como de las glándulas sebáceas. Aun así, las células de la
prominencia folicular no contribuyen a la población de las células madre basales de la epidermis.
Los pelos están compuestos por células queratinizadas que se desarrollan a partir de
folículos pilosos.
La cornificacion del pelo y de la vaina radicular interna se produce poco después de que las células dejan la
matriz en una región llamada zona queratogena , ubicada en el tercio inferior del folículo. Las células que
pasan por esta zona expulsan sus orgánulos y se compactan con los filamentos intermedios de queratina
reticulados. Cuando el pelo emerge del folículo, ya está cornificado por completo como queratina dura. La
vaina radicular interna, que es de queratina blanda, no emerge del folículo, sino que se desintegra en el
istmo folicular.
Una lámina basal gruesa, la membrana vítrea, separa al folículo piloso de la dermis. Alrededor del folículo
hay una vaina de tejido conectivo denso irregular. El musculo erector del pelo se inserta en la
protuberancia folicular.
Los pelos se componen de queratinas duras fuertemente reticuladas y constan de 3 capas:
Medula: forma la parte central del tallo del pelo. Solo está presente en pelos gruesos. Tiene células
con queratina blanda.
Corteza: es el 80% de la masa total del pelo. Esta fuera de la medula y tiene células corticales llenas
de filamentos de queratina dura. Cada filamento está rodeado por un espacio amorfo lleno de
proteínas asociadas a la queratina
Cutícula del pelo: la capa más externa, son varias capas de células escamosas superpuestas,
semitransparentes y queratinizadas. La cutícula protege el pelo del daño físico y químico, y
determina su porosidad.
Glándulas sebáceas
Las glándulas sebáceas secretan el sebo que cubre la superficie del pelo y la piel
Se originan como brotes en la vaina radicular externa del folículo piloso y suele haber varias glándulas por
folículo. El sebo es el producto de esta secreción holocrina. La célula entera produce y se llena de lípidos
mientras que al mismo tiempo sufre una apoptosis. En última instancia, tanto el producto de secreción
como el detrito celular se eliminan desde la glándula hacia el infundíbulo del folículo piloso, que junto con
el conducto corto de la glándula sebácea, forma el conducto pilosebaceo. La actividad mitótica de las
células basales en la periferia de la glándula, produce células nuevas y las células en la glándula
permanecen unidas entre sí por desmosomas. El proceso de producción del cebo tarda unos 8 días.
Glándulas sudoríparas
Las glándulas sudoríparas se clasifican según su estructura y la índole de su secreción. Hay dos tipos
Glándulas sudoríparas ecrinas: en todo el cuerpo, salvo labios y ciertas áreas de los genitales externos
Glándulas sudoríparas apocrinas: se limitan a la axila, la areola y el pezón, la región perianal y los genitales
externos. Las glándulas ceruminosas del conducto auditivo externo y las glándulas apocrinas de las
pestañas (de Moll) también son de tipo apocrino.

ROSARIO PEREZ CONTRERAS
13
Glándulas sudoríparas ecrinas
Las glándulas sudoríparas ecrinas son glándulas tubulares simples que regulan la
temperatura corporal
Son independientes, no asociadas con el folículo piloso. Tienen estructura tubular simple enrollada y de
fondo ciego. Tiene un segmento secretor en la dermis profunda y un segmento canalicular que se continua
con el anterior y desemboca en la superficie epidérmica.
Estas glándulas regulan la temperatura a través el enfriamiento causado por la evaporación del agua del
sudor sobre la superficie del cuerpo. La secreción es semejante a un ultrafiltrado de la sangre. Es de tipo
hipotónico debido a la resorción de sodio y agua en el conducto excretor, además la secreción es baja en
proteínas y contiene cantidades variables de cloruro de sodio, urea, ácido úrico y amonio. Por lo tanto la
glándula de sudor ecrino actúa en parte como un órgano excretor.
El segmento secretor de la glándula contiene tres tipos celulares
Hay células claras y células oscuras, ambas células epiteliales secretoras y células mioepiteliales que son
células epiteliales contráctiles. Se distribuyen como un epitelio pseudoestratificado.
Las células claras tienen abundancia de glucógeno (se tiñen con PAS). Su citoplasma con HE se tiñe muy
poco. Su morfología indica que producen el componente acuoso del sudor
Las células oscuras tienen un RER y gránulos de secreción abundantes. El citoplasma apical contiene
gránulos de secreción maduros y ocupa la mayor parte de la superficie luminal. Tienen actividad secretora
de glucoproteínas.
Las células mioepiteliales están entre las células secretoras, su citoplasma tiene muchos filamentos
contráctiles de actina que permite su fácil identificación.
El segmento canalicular está revestido por un epitelio biestratificado cubico, que consiste en una capa
basal y una capa luminal de células. Carece de células mioepiteliales.
Glándulas sudoríparas apocrinas
Las glándulas apocrinas son glándulas tubulares enrolladas (a veces se ramifican) de luz amplia que están
asociadas con los folículos pilosos.
La porción secretora (adenomero) de las estas glándulas tiene una luz más amplia
que la de las glándulas ecrinas y está compuesta por un solo tipo celular.
Las apocrinas almacenan su producto de secreción en la luz. Los adenomeros tienen un epitelio simple, con
un solo tipo celular y citoplasma esosinofilo. Su secreción es de tipo merocrino, el citoplasma apical tiene
abundantes gránulos de secreción.
Las células mioepiteliales también están presentes en la porción secretora y se encuentran entre las
células secretoras y la lámina basal contigua. La contracción de estas células facilita la expulsión del
producto de secreción.

ROSARIO PEREZ CONTRERAS
14
El conducto excretor de las glándulas apocrinas está revestido por un epitelio
estratificado cubico y carece de células mioepiteliales
No hay resorción de sustancias en el conducto apocrino. El producto de secreción no se altera en su paso a
través del conducto.
El epitelio del conducto es estratificado cubico, en general de dos capas de espesor.
Las glándulas apocrinas producen una secreción co n proteínas abundantes que
contiene feromonas.
La secreción contiene proteínas, hidratos de carbono, amonio, lípidos y ciertos compuestos orgánicos que
le dan color.
Uñas
Las uñas son placas de células queratinizadas que contienen queratina dura
Las uñas se encuentran levemente arqueadas y se denominan placas ungulares, las cuales descansan sobre
los lechos ungulares. El lecho es de células epiteliales que son continuas con el estrato basal y espinoso de
la epidermis.
La parte proximal de la uña, la raíz ungular esta oculta por un pliegue de la epidermis y cubre las células de
la zona germinativa o matriz.
Las células madre de la matriz se dividen con regularidad, migran a la raíz y allí se dividen para producir la
queratina de la uña. La queratina de la uña es queratina dura como la de la corteza del pelo. Esta queratina
no se descama, a diferencia de la queratina blanda de la epidermis. Y tiene un elevado contenido de azufre
responsable de la dureza de la uña.
A medida que la superficie de la uña crece, se desliza sobre el lecho ungular. Desde el punto de vista
microscópico, la superficie de la uña contiene corneocitos interdigitados muy juntos, que carecen de
núcleo y orgánulos.
El área blanca en forma de media luna ubicada cerca de la raíz de la uña, la lúnula tiene ese aspecto claro a
causa de la gruesa capa opaca de células de la matriz con cornificacion parcial que hay en ese sitio. Cuando
la superficie de la uña se queratiniza por completo, es más transparente y adquiere el color del lecho
vascular subyacente.
La cutícula también se compone de queratina dura; por lo tanto no se descama.
DIGESTIVO III Higado y pancreas- RPC.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.