
SISTEMA SOMATOSENSORIAL Y DOLOR
Procesamiento de la información sensorial
La señal física es captada por el sistema sensorial a través de una estructura especializada en su
recolección, filtración y ampliación. Esta tiene células especializadas que transducen la señal
física en una señal eléctrica. Estas señales se transmiten desde vías nerviosas sensoriales
especificas desde el sistema periférico al SNC donde serán utilizadas para la percepción o para
la acción.
Durante la transducción, el estímulo sensorial activa su receptor produciendo un cambio
conformacional que lleva a la apertura o cierre de canales iónicos produciendo un potencial que
podrá ser excitatorio o inhibitorio. Ejemplos:
● Mecanorreceptor táctil (corpúsculo de Pacini): La deformación de la capsula producida
por el estímulo, estira la membrana del receptor, aumentando la probabilidad de
apertura de canales iónicos asociados a la membrana. Al aumentar la conductancia de
estos cationes se produce la despolarización de la membrana que si alcanza el umbral
producirán potenciales de acción que se enviarán al SNC.
● Fotorreceptores (retina): La membrana del fotorreceptor está normalmente
despolarizada durante la oscuridad gracias a la apertura de unos canales de Na y Ca
controlados por GMPc. Cuando incide la luz sobre el fotorreceptor, la rodopsina cambia
de conformación en presencia de un fotón y activa una serie de segundos mensajeros
que resultaran en la disminución en los niveles de GMPc, por lo que los canales iónicos
se cerraran y se hiperpolarizará la membrana. Por lo cual el potencial será inhibitorio.
Existen dos tipos de receptores sensoriales:
● Axones modificados de la neurona aferente que generará un potencial de acción en
respuesta al estímulo. El soma del receptor se localiza en el ganglio de la raíz dorsal. Es
el caso del sistema somatosensorial.
● Células especializadas que generará un potencial excitatorio o inhibitorio en respuesta
al estímulo. Esta célula hace sinapsis con la neurona aferente que posee canales
dependientes de voltaje y generará potenciales de acción dirigidos al SNC. Este es el
caso de los sentidos especiales (visión, gusto, audición), cuyos receptores se encuentran
en órganos especializados.
Los receptores sensoriales transmiten la señal eléctrica hacia la medula espinal y el tronco
encefálico. Las vías de cada sistema sensorial terminal en diferentes núcleos del tálamo (a
excepción del sistema olfatorio). El tálamo proyecta a la corteza sensorial primaria y secundaria
correspondiente. El nervio olfatorio es el único que proyecta directamente hacia la corteza
olfatoria sin pasar por el tálamo.

El tálamo, el tronco encefálico y la medula reciben a su vez información descendente desde la
corteza que modifica el flujo de información aferente desde los sistemas sensoriales.
Sensación vs Percepción
La sensación resulta de la interacción de receptores específicos con los estímulos. La percepción
resulta de la representación consciente de la experiencia sensorial. No se conoce cuando
termina la sensación y comienza la percepción, sin embargo, estudios indican que las cortezas
sensoriales primarias están relacionadas con la sensación y las cortezas secundarias están
relacionadas más con la percepción.
Características de un estímulo
Los estímulos tienen 4 características fundamentales: Modalidad, intensidad, duración y
localización.
Modalidad
Es el tipo de señal física para la que está especializado un receptor. Por ejemplo, los
fotorreceptores producen información de tipo visual mientras que los mecanorreceptores
especializados en la transducción de ondas sonoras producen información de tipo auditiva. Los
sentidos cuyos receptores se ubican en órganos se denominan sistemas sensoriales especiales;
mientras que aquellos cuyos receptores están localizados en los ganglios anexos a la raíz dorsal
forman parte del sistema somatosensorial.
Intensidad
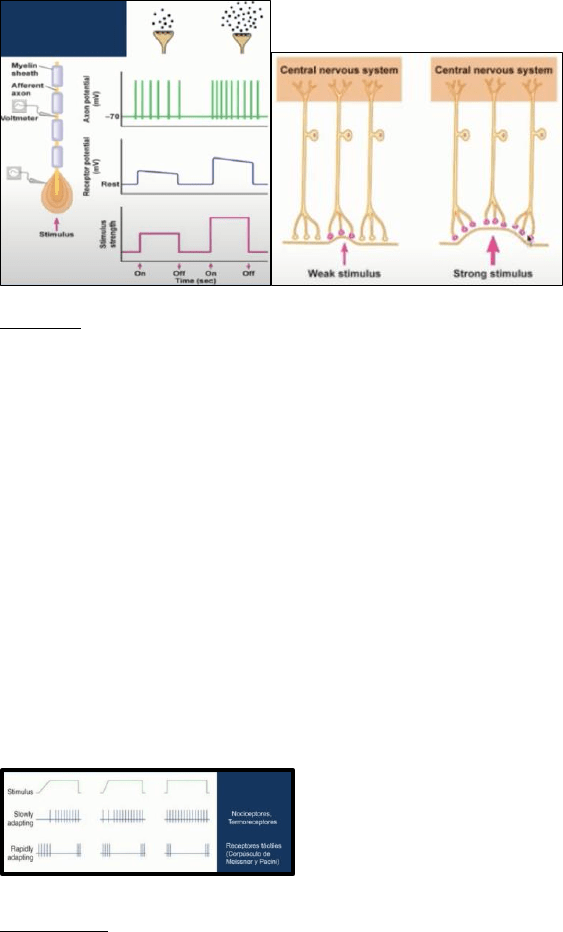
Los receptores codifican la intensidad de un estímulo a través de la amplitud del potencial
receptor. Cuanto mayor el estímulo, mayor la amplitud del potencial receptor y mayor será la
frecuencia de disparo de potenciales de acción de la neurona aferente. A esto se le llama código
de frecuencia.
La intensidad también se codifica en base a la cantidad de receptores activados. Cuanto mayor
sea el estímulo, mayor será la cantidad de receptores reclutados (código poblacional).
Duración
Algunos receptores realizan una descarga sostenida en presencia de un estímulo sostenido
mientras que otros se activan inicialmente y luego se desactivan. La mayoría de los receptores
se adaptan a la duración del estímulo.
La adaptación consiste en la disminución en la respuesta del receptor ante la presencia
prolongada del mismo estímulo. Esta se puede dar tanto a nivel de los receptores sensoriales
como en el SNC. Algunos receptores se adaptan rápidamente y otros lo hacen más lentamente.
Los de adaptación lenta (tónicos) disparan mientras el estímulo está presente (nociceptores,
termorreceptores). En cambio, los receptores de adaptación rápida (fásicos) responden de
manera rápida pero breve a los estímulos, y su respuesta disminuye si se mantiene el estímulo
(corpúsculo de Meissner y Pacini). Estos últimos serán más eficientes en sensar aspectos
dinámicos del estímulo. En contraste, los receptores de adaptación lenta son más eficientes en
la extracción de información estática del estímulo como su forma y su tamaño.
Localización
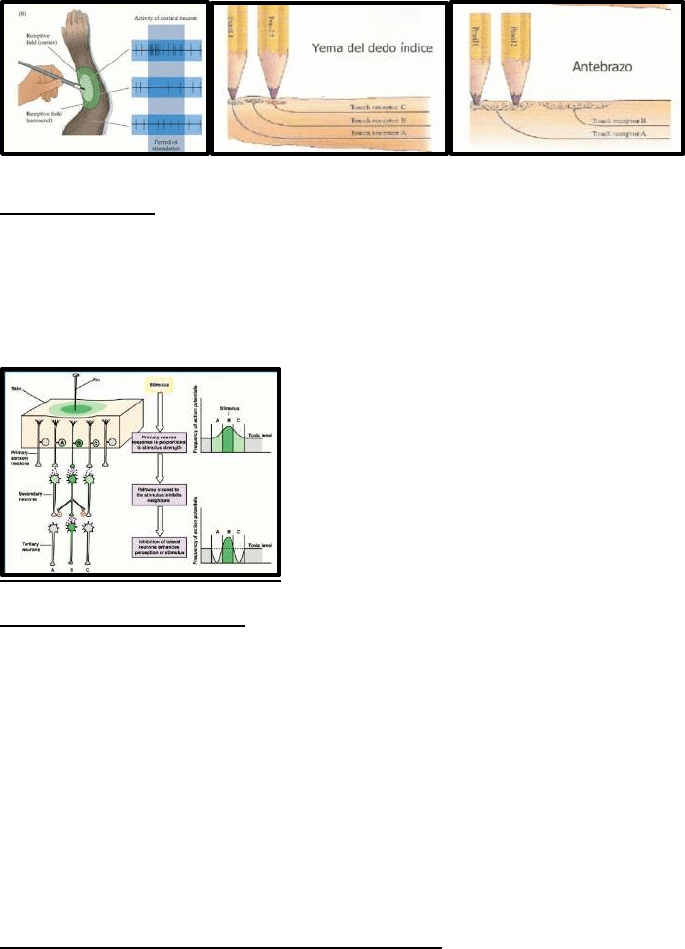
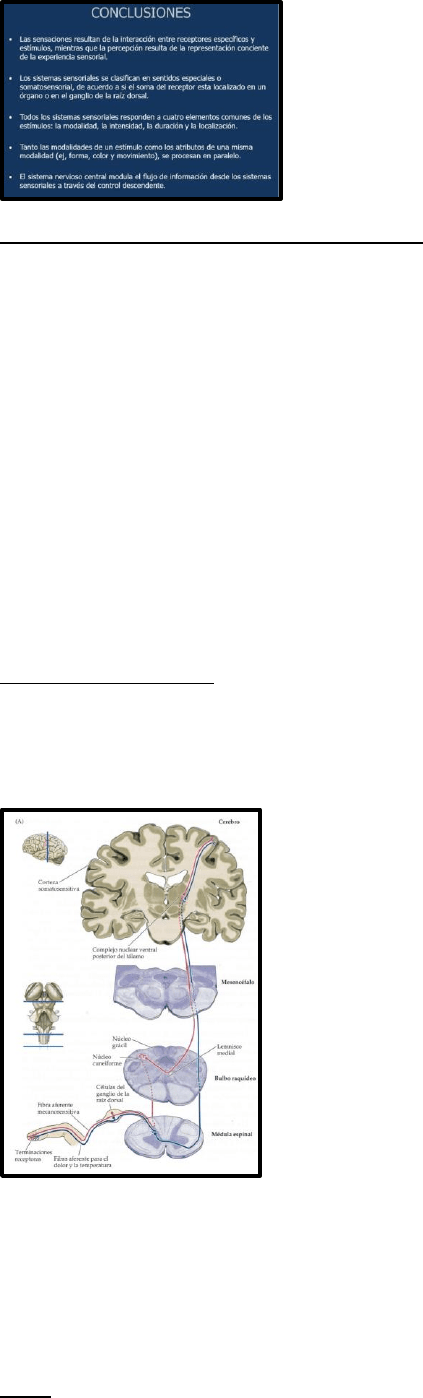
El sistema nervioso codifica la localización espacial del estímulo a partir de su campo receptivo.
El campo receptivo de una neurona sensorial es el área (de la piel, retina, etc.) cuya estimulación
aumenta la frecuencia de disparo de la neurona aferente. La estimulación del centro del campo
receptivo produce un aumento en la frecuencia de disparo, mientras que la estimulación de la
periferia produce una disminución de la frecuencia de disparo. El tamaño del campo receptivo
(resolución espacial) varía para un mismo tipo de receptor dependiendo de la zona del cuerpo
estimulada. En la piel, el umbral de dos puntos (mínima distancia en la que se pueden sentir dos
estímulos como tales) es más bajo en los dedos y en la cara que en otras zonas como los muslos
y los antebrazos. La resolución espacial será mayor en las zonas con campos receptivos
pequeños. La mayor resolución espacial ocurre a expensas de una mayor inervación.
Inhibición lateral
Mecanismo que contribuye a la localización efectiva aumentando la resolución espacial del
estímulo. Es la capacidad de una neurona excitada para reducir la actividad de las neuronas
vecinas en el sentido lateral. Este sistema también funciona a nivel de los sistemas auditivos,
visual, táctil y olfativo.
Procesamiento en paralelo
Vías que operan de forma complementaria y de manera simultánea en la extracción de varios
atributos de la modalidad de un estímulo. Ejemplo: La forma de un objeto está determinado
por la vía parvocelular mientras que el movimiento está determinado por la vía magnocelular,
ambas operan de forma complementaria y en simultáneo para extraer los atributos de forma y
movimiento. El procesamiento en paralelo de estos atributos persiste a nivel de la corteza en
dos grandes vías especializadas para distintos tipos de información. Una vía ventral (corteza
visual 1º hacia la temporal) que se especializa en el proceso de la forma y el detalle, y otra vía
dorsal (desde corteza visual 1º hacia la parietal posterior) que utiliza la información visual acerca
de los objetos y su movimiento para el control de la acción.
Control descendente de la información sensorial
A todos los niveles de la jerarquía sensorial existen proyecciones descendentes desde los centros
superiores. Un ejemplo es el tálamo, que recibe información aferente desde los sistemas
sensoriales hacia sus distintos núcleos, y luego esta información se envía a la corteza. Pero a la
vez recibe retroalimentación desde la corteza hacia el tálamo a través del núcleo reticular. Las
neuronas de la corteza también pueden modular directamente el funcionamiento de los
receptores periféricos. Por ejemplo, la corteza motora puede alterar la sensibilidad de los
receptores sensoriales en el músculo esquelético optimizando su función.
La atención tiene un rol importante en el control de la información sensorial ya que según donde
se ponga el foco de atención, cambia la percepción. La atención tiene un papel clave en la
modulación de la percepción del dolor.
Modalidades del sistema somatosensorial (SS)
Los receptores del sistema somatosensorial procesan estímulos de 4 modalidades distintas.
● Tacto: Mecanorreceptores cutáneos y subcutáneos responsables del tacto fino,
vibración y presión.
● Propiocepción: Mecanorreceptores ubicados en los músculos, tendones y
articulaciones encargados de sensar la posición de los miembros y partes del cuerpo en
el espacio.
● Temperatura: En su mayoría son terminales libres ubicados en la piel. Sensan calor o
frio no nocivo.
● Dolor: En su mayoría son terminales libres ubicados en la piel, mucosas, músculos y
articulaciones que sensan estímulos nociceptivos que producen daño en el tejido.
Los receptores del sistema SS son terminales especializados de la neurona aferente cuyo soma
está ubicado en el ganglio de la raíz dorsal.
Vías somatosensoriales
La información aferente se transmite al SNC por dos cordones principales. La información de
dolor y temperatura viaja por el sistema anterolateral. La información sobre tacto y
propiocepción lo hace a través de la columna dorsal/lemnisco medio.
Los axones aferentes de mayor diámetro del sistema SS son las fibras de los propioceptores que
alcanzan una velocidad de conducción de 80-120 m/s. Luego le siguen los axones de los
receptores táctiles que como son de menor diámetro alcanzan una velocidad de 35-75 m/s.
AMBOS TIPOS DE FIBRAS SON MIELÍNICAS.
Tacto

Tipos de receptores táctiles
Los receptores táctiles generan una representación neural de alta definición de los objetos que
manipulamos. Cuatro tipos de receptores táctiles encapsulados contribuyen al tacto epicrítico:
● Discos de Merkel: Son los únicos ubicados en la epidermis y son abundantes en la punta
de los dedos. 25% de los mecanorreceptores de la mano. Campos receptivos pequeños
y bien definidos, de adaptación lenta. Recepción espacial más alta de todos los
receptores táctiles.
● Corpúsculos de Meissner: Se encuentran en la dermis, justo debajo de la epidermis.
Formados por una capsula de tejido conectivo que contiene un conjunto de células
aplanadas derivadas de las células de Schwann. 40% de la inervación táctil de la mano.
Tienen campos receptivos pequeños, aunque algo más grandes que los de Merkel. Son
de adaptación rápida. Son más sensibles a la deformación de la piel que los de Merkel.
Son eficaces en sensar vibraciones de baja frecuencia.
● Corpúsculos de Pacini: Dermis profunda o tejido subcutáneo. Tiene capas concéntricas
que rodean a la fibra aferente. 10-15%. Responden a la vibración y son responsables del
tacto profundo. Sensan estímulos de alta frecuencia y son de adaptación rápida, esto
hace que generen potenciales de acción en respuesta a desplazamientos muy pequeños
(10nm). Sus campos receptivos son muy grandes y con límites poco definidos. Son
eficaces para detectar vibraciones que surgen del contacto de objetos con la mano.
● Corpúsculos de Ruffini: Consisten en especializaciones capsulares con forma de huso
ubicadas profundamente en la dermis. 20%. Responden al estiramiento de la piel. Son
de adaptación lenta. El eje largo del huso suele ubicarse en paralelo con las líneas de
estiramiento de la piel detectando estiramiento cutáneo causado por el movimiento de
los dedos.
Los receptores táctiles se diferencian en el tamaño de su campo receptivo y en el tipo de
adaptación.
Dermatomas
Es el territorio de la piel inervado por cada nervio espinal. Varían entre individuos y están
superpuestos sustancialmente de modo que una lesión en la raíz dorsal no implica
necesariamente la perdida completa de la sensación en la región de piel inervada.

Propiocepción
Los propioceptores proporcionan información sobre las fuerzas mecánicas que surgen desde el
sistema esquelético. Su función es proveer información detallada y continua sobre la posición
de las extremidades y otras partes del cuerpo en el espacio. Los principales mecanorreceptores
involucrados en la propiocepción son: Huso neuromuscular, órgano tendinoso de Golgi y los
receptores articulares (corpúsculo de Pacini y Ruffini). La posición y movimiento de la cabeza
está censada por propioceptores ubicados en el sistema vestibular.
Receptores propioceptivos
● Huso neuromuscular: Ubicado en el músculo esquelético. Sensa cambios en la longitud
del musculo. Consisten en fibras musculares intrafusales especializadas que se
extienden en paralelo a las fibras musculares. Las fibras aferentes se envuelven
alrededor de la parte central del huso. Cuando el musculo se estira, la tensión activa
canales iónicos en la membrana disparando potenciales de acción. El huso
neuromuscular está inervado por dos tipos de fibras: las fibra 1-a, son los axones
mielínicos más grandes de todos los receptores somatosensoriales. Son de adaptación
rápido y responden velozmente a cambios en la longitud del músculo. Las fibras 2,
también son mielínicas, aunque de menor diámetro y producen respuestas sostenidas
a longitudes musculares constantes. Los músculos involucrados en movimientos finos
tienen alta concentración de husos, los involucrados en movimientos gruesos tienen
baja concentración de husos.
● Órgano tendinoso de Golgi: Informan al SNC sobre cambios en la tensión muscular. Están
dispuestos en serie entre el músculo y el tendón. Las fibras que los inervan son fibras 1-
b, también mielínicas y de gran tamaño.
● Receptores articulares: Se parecen a los táctiles de tipo Ruffini y Pacini. Se ocupan de
sensar la posición y movimiento de la articulación. No parecen contribuir
significativamente en codificar la posición de los miembros en el espacio. Parecen ser
importantes en codificar la posición de los dedos de las manos.
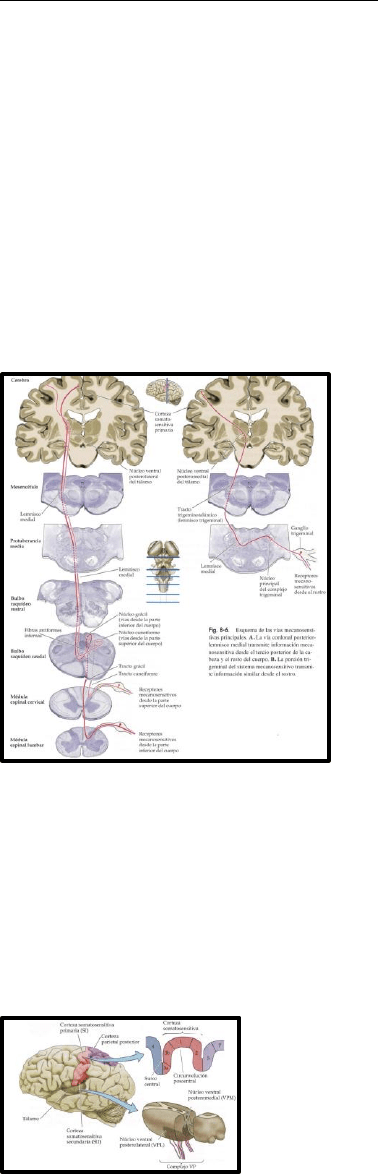
Vía de la columna dorsal/lemnisco medio
Las señales eléctricas producidas en los receptores táctiles y propioceptivos se transmiten en
paralelo al cerebro por la vía de la columna dorsal/lemnisco medio. Las vías somatosensoriales
tienen 2 sinapsis desde el receptor hasta la corteza: Una a nivel del bulbo raquídeo, que es
donde se decusan las fibras al lado contralateral de la médula; la otra está a nivel del tálamo.
Los axones de las neuronas de segundo orden con somas en los núcleos grácil y cuneiforme se
decusan hacia el lado contralateral de la médula y asciende hacia el núcleo ventral posterior del
tálamo. Las neuronas de tercer orden en el núcleo ventral posterior del tálamo proyectan a
través de la cápsula interna hacia la corteza, por lo tanto, la corteza somatosensorial representa
señales táctiles y propioceptivas generadas en la superficie contralateral del cuerpo. Finalmente,
la porción trigémina del sistema mecanosensorial transmite información similar acerca de la
cara.
La lesión de la vía de la columna dorsal/lemnisco medio o el daño a nivel de los receptores puede
provocar ataxia sensorial, que es la falta de coordinación de los movimientos debido al déficit
espacial.
La corteza somatosensorial primaria ocupa el giro poscentral de la corteza parietal ubicado
posteriormente al surco central. La corteza somatosensorial primaria comprende las áreas 3a,
3b, 1 y 2 de Brodmann. El área 3a responde a estímulos propioceptivos, el área 3b y 1 responde
a estímulos cutáneos. El área 2 responde a ambos.
Existe una atenuación sensorial que ocurre en la corteza somatosensorial primaria que, ante un
movimiento generado por uno mismo, se inhibe. El cerebelo parece ser el responsable de esta
inhibición. Es por esto que no podemos hacernos cosquillas a nosotros mismos.
La corteza somatosensorial es plástica y se modifica en función de la experiencia.
Dolor y temperatura
Los somas de los receptores nociceptivos y de temperatura también se alojan en el ganglio
anexo a la raíz dorsal (GARD). La información sobre dolor y temperatura viaja al SNC por el
sistema anterolateral.
El dolor es la experiencia sensorial y emocional asociada al procesamiento de estímulos que
causan daño tisular. Los nociceptores son terminaciones periféricas de las fibras aferentes
primarias (terminaciones libres) que responden al daño tisular por estímulos térmicos, químicos
y mecánicos. Los nociceptores están ubicados en la piel, mucosas, articulaciones, músculos y en
menor medida en las vísceras.
Los axones de los nociceptores pueden ser de dos tipos: fibras Aδ y fibras C. Ambas son mucho
más pequeñas que la de los mecanorreceptores y, por lo tanto, son mucho más lentas.
Las fibras Aδ son mielínicas y de mayor diámetro que las fibras C, que son amielínicas y de menor
diámetro. Las fibras C además de sensar dolor, median también la sensación de picazón y el
tacto protopático (es el tacto grueso con poca resolución espacial).
La percepción del dolor puede descomponerse en dos fases: La fase temprana, caracterizada
por un dolor agudo. Seguida de la fase tardía y duradera de dolor difuso. Ante un estímulo
doloroso se reclutan inicialmente las fibras rápidas Aδ. Si la intensidad se incrementa se reclutan
entonces las fibras amielínicas lentas de tipo C. Los opiáceos actúan eliminando la fase lenta o
tardía.
Nociceptores vs termorreceptores
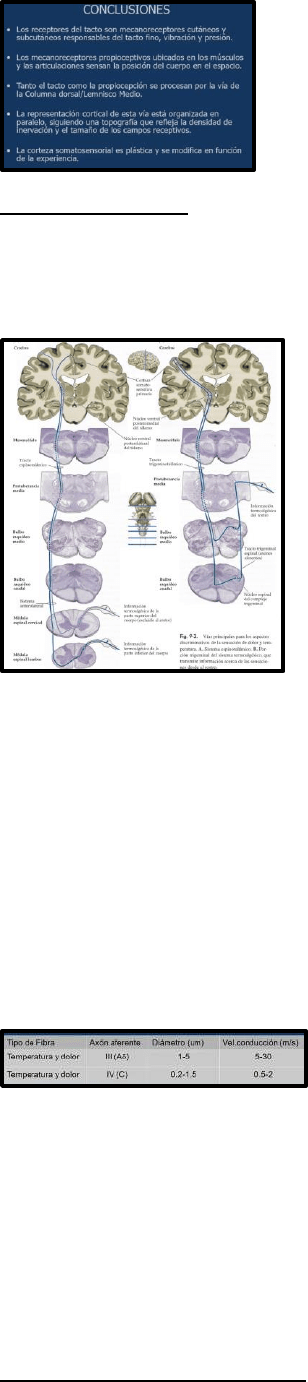
A temperaturas aproximadas de 45º, la frecuencia de disparo de los termorreceptores ya está
saturada, mientras que la de los nociceptores aumenta y sigue aumentando a medida que se
intensifica el estímulo (45º es la temperatura en la cual se considera nocivo el estímulo).
Se cree que las sensaciones de calor y frío no nocivo mediadas por termorreceptores están
procesadas por fibras que responden al calor, y otras que responden al frío.
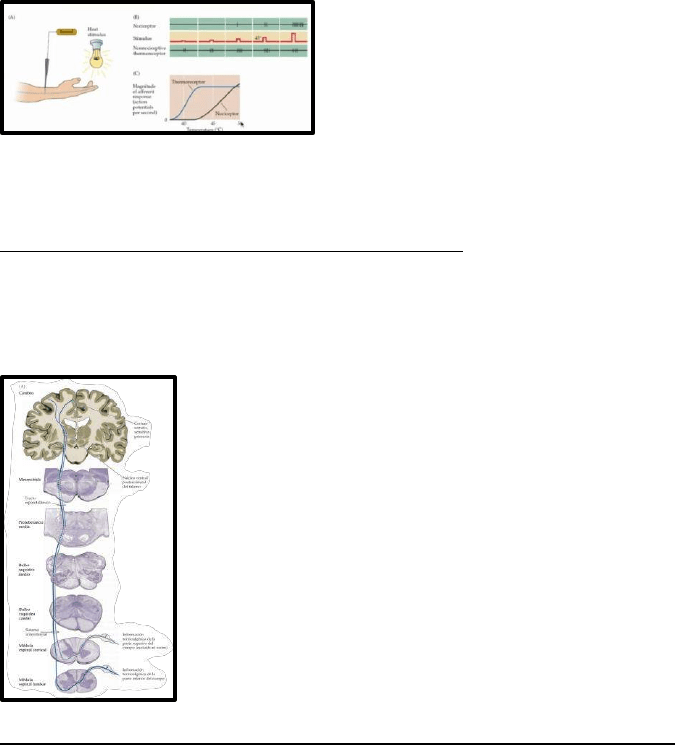
Vía espinotalámica (parte del sistema anterolateral)
Media los aspectos sensoriales discriminativos del dolor, es decir, su localización, su intensidad
y su calidad. Esta vía también media el componente inicial o primera sensación del dolor. Las
neuronas que forman parte de esta vía poseen campos receptivos pequeños y localizados.
Vía espinoretucular y espinomesencefálica (parte del sistema anterolateral)
Procesan información sobre los aspectos afectivos y cognitivos del dolor como puede ser el
sentimiento desagradable, el miedo y la ansiedad que provoca. El espinoreticular proyecta a las
regiones de la formación reticular en el bulbo raquídeo. La vía espino mesencefálica proyecta al
núcleo parabraquial en la protuberancia y al colículo superior y la sustancia gris periacueductal
en el mesencéfalo. Estas regiones proyectan a su vez a la amígdala, el hipotálamo y a la corteza
cingulada anterior y a la ínsula, que en conjunto forman el sistema límbico. El sistema límbico
tendrá un rol clave en el procesamiento de los aspectos motivacionales y afectivos del dolor.

Hiperalgesia periférica
Los receptores táctiles se adaptan ante la presencia de un estímulo continuo, esto no ocurre en
el caso del dolor. La sensación del dolor no solo no se adapta, sino que se intensifica. Este cambio
de sensibilidad genera un estado de amplificación del dolor llamado hiperalgesia. Las
propiedades de los nociceptores cambian en la hiperalgesia periférica como consecuencia de
una noxa. Ante el estímulo nocivo, los nociceptores liberan mediadores como el CGRP y la
sustancia P en la zona inflamada. Las células del tejido conectivo liberan sustancias que
interactúan directamente sobre los terminales nociceptivos aumentando su respuesta a la noxa
(agente cuya presencia causa una alteración).
Hiperalgesia central
Resulta del aumento de la excitabilidad de las neuronas de segundo orden del sistema
anterolateral ubicadas en el asta dorsal de la médula. Esta se desencadena por varios
fenómenos: Liberación masiva de glutamato por parte de los nociceptores, reducción de la
inhibición espinal mediada por GABA y la liberación de citoquinas por parte de la glía que tiene
acción sensibilizante. En conjunto, estos fenómenos aumentan la transmisión glutamatérgica a
nivel de la neurona secundaria mediada por los receptores NMDA. En la hiperalgesia central se
generaliza las señales de mecanorreceptores de bajo umbral, por lo que un estímulo que antes
era inocuo genere una sensación de dolor (alodinia) y puede durar varias horas después de la
desaparición de la noxa.
Lesiones de la medula espinal
El sistema anterolateral cruza y asciende por el lado contralateral de la médula mientras que el
sistema de la columna dorsal asciende por el lado ipsilateral. De acuerdo con los distintos niveles
en los que se decusan estas vías, una lesión espinal tendrá ciertas características en cada una.
Una lesión espinal del lado izquierdo a nivel torácico ocasionará una pérdida de tacto y
propiocepción del mismo lado del cuerpo (izquierdo). Esta misma lesión producirá analgesia y
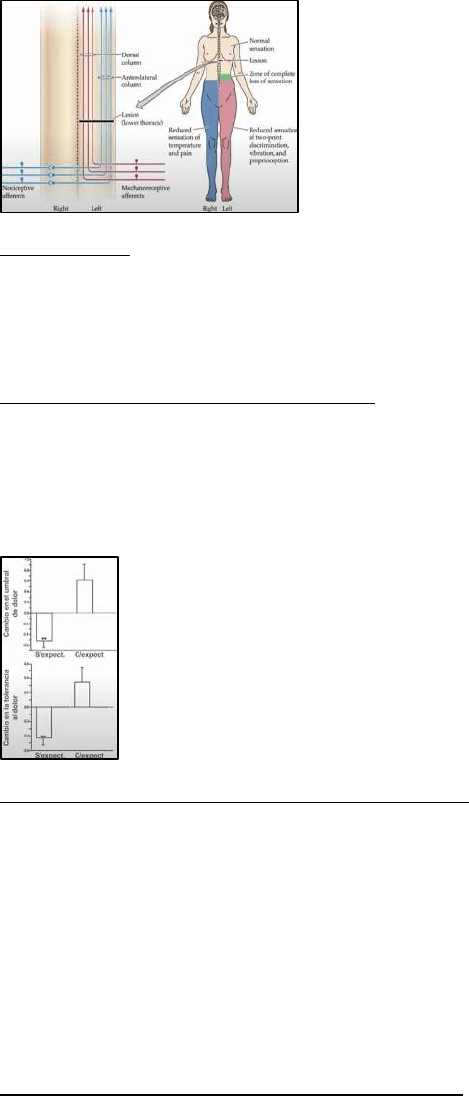
pérdida de sensación de temperatura del lado derecho del cuerpo. Estos déficits ocurrirán en
las regiones del cuerpo por debajo de la lesión.
Dolor referido
Muy pocos receptores nociceptivos están especializados en el sensado del dolor visceral. Como
la información sobre dolor visceral y cutáneo converge a nivel de la neurona de segundo orden,
es común que la percepción del dolor visceral se refiera a nivel cutáneo.
Control descendente del dolor: cognición
La cognición juega un rol importante en la modulación de la percepción de dolor. La atención
como la expectativa pueden afectar significativamente el umbral de dolor, así como su
tolerancia. La percepción de dolor puede ser modulada simplemente por la expectativa creada
(efecto placebo).
Control descendente del dolor: analgesia endógena
El sistema nervioso produce sustancias químicas con propiedades opiáceas. La estimulación de
la sustancia gris periacueductal induce la liberación de opiáceos endógenos a través de la vía
descendente inhibitoria. Esta vía modula la actividad de las neuronas secundarias en el asta
posterior de la médula. Este mecanismo de analgesia endógena está mediado por interneuronas
en el asta posterior que liberan encefalinas que se unen a receptores específicos en el terminal
presináptico (neurona de primer orden) como en los somas de las neuronas de segundo orden.
Las encefalinas tienen un efecto inhibitorio en estas regiones, produciendo analgesia. Los
opiáceos comerciales actúan sobre estos mismos receptores.
Control descendente del dolor: miembro fantasma
Sugiere que la corteza somatosensorial sigue siendo funcional en ausencia del estímulo
periférico. El 75% de los pacientes con miembro fantasma, desarrolla dolor en la extremidad
faltante. No se conoce el origen del dolor del miembro fantasma, se intuye que puede ser
originado por un reordenamiento disfuncional a nivel de la corteza somatosensorial.


Sistema somatosensorial y dolor.doc
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.