
38
El sistema nervioso
Estudio de caso
¿Cómo te amo?
“¡Espera! ¿Qué es esa luz que asoma por la ventana?
Es el Este y Julieta es el Sol.”
—Romeo y Julieta, Acto II, escena II
EN LA OBRA ROMEO Y JULIETA de Shakespeare, dos
adolescentes se enamoran a primera vista. Unas
horas más tarde, mientras Romeo observa a Julieta
asomada por la ventana, la ve como si fuera el Sol que
ilumina su vida. Esta obra de Shakespeare ilustra con
dramatismo el poder del amor romántico, por el cual
los amantes desafían a su familia, arriesgan su fortuna
y su futuro y, por último, sacrifican su vida.
Desde luego, el romance no es la única
manifestación del amor. El amor de una madre por su
hijo es igual de poderoso. Incluso algunas personas
han definido su vida y han estado dispuestas a morir
por amor a Dios y a su país. Pero, ¿qué es el amor?
¿Todos estos tipos de amor son diferentes o están
relacionados? ¿Qué pasa en el cerebro cuando dos
amantes se encuentran o una madre arrulla a su
bebé?
Nadie lo sabe con seguridad; en todo caso,
no en la gente. Quizá lo sorprendente es que los
neurocientíficos saben mucho del amor, o por lo
menos de la unión en parejas y el sexo, en lo que
respecta a un pequeño roedor llamado ratón de las
praderas. Si Julieta hubiera sido uno de estos roedores,
su primer encuentro con Romeo habría liberado gran
cantidad de oxitocina, la misma hormona que
provoca las contracciones uterinas durante el parto.
La oxitocina se habría unido a receptores en algunas
pequeñas áreas del cerebro, haciendo que las células
nerviosas liberaran dopamina, conocida a menudo
como el químico del placer del cerebro. Se habría
sentido maravillosamente. Es más, habría asociado
esa sensación de euforia con Romeo. Asimismo, en
caso de que Romeo hubiera sido un ratón de las
praderas, algunas de las moléculas y regiones del
cerebro habrían diferido, pero el resultado final habría
sido similar: la liberación de un torrente de dopamina, y
habría pensado que la sensación que ello le provocaba
sólo podía estar asociada con Julieta. De modo que es
muy probable que ambos ratones de las praderas se
aparearan y unieran sus vidas, aunado a ello el hecho
de construir un nido, vivir juntos y criar a sus hijos.
¿Cómo perciben su mundo los seres humanos y
otros animales? ¿Cómo evalúan lo que perciben, y se
tranquilizan o emocionan, temen o se entusiasman?
¿Cómo responden con comportamientos apropiados
como descansar, comer o aparearse? Aunque no se ha
entendido por completo la mayoría de las percepciones
y comportamientos, las respuestas a estas preguntas se
encuentran en el sistema nervioso.
Capítulo
Amor: “¿Un fuego que brilla
en los ojos de los amantes…
una locura prácticamente
discreta” o simplemente
la mezcla correcta de
sustancias químicas en lo
más profundo del cerebro de
los enamorados? El Microtus
ochrogaster proporciona
algunos conocimientos sobre
la base neuroquímica del
amor emocional. (detalle)

734 UNIDAD 5 Anatomía y fisiología animal
38.1
¿CUÁLES SON LAS ESTRUCTURAS Y LAS
FUNCIONES DE LAS CÉLULAS NERVIOSAS?
El sistema nervioso tiene dos tipos de células principales: neuro-
nas, llamadas a menudo células nerviosas, y células gliales. Como
se verá más adelante, las neuronas reciben, procesan y transmiten
la información. Las células gliales ayudan a la función neuronal de
varias formas, como proveer nutrientes, regular la composición del
líquido extracelular en el cerebro y la médula espinal, modular la
comunicación entre las neuronas, y acelerar el movimiento de las
señales eléctricas dentro de las neuronas. Aunque las células gliales
son muy importantes —pues el sistema nervioso no podría funcio-
nar sin ellas—, este capítulo se enfoca en la estructura y función de
las neuronas.
Las funciones de una neurona se localizan
en partes separadas de la célula
Una neurona debe realizar cuatro funciones:
1. Recibir información del ambiente interno o externo, o de
otras neuronas.
2. Procesar esta información, a menudo junto con la de otras
fuentes, y producir una señal eléctrica.
3. Transportar la señal eléctrica, en ocasiones a una distancia
considerable, hasta la unión con otra célula.
4. Transmitir la información a otras células, ya sea a otras
neuronas o a las células de músculos o glándulas.
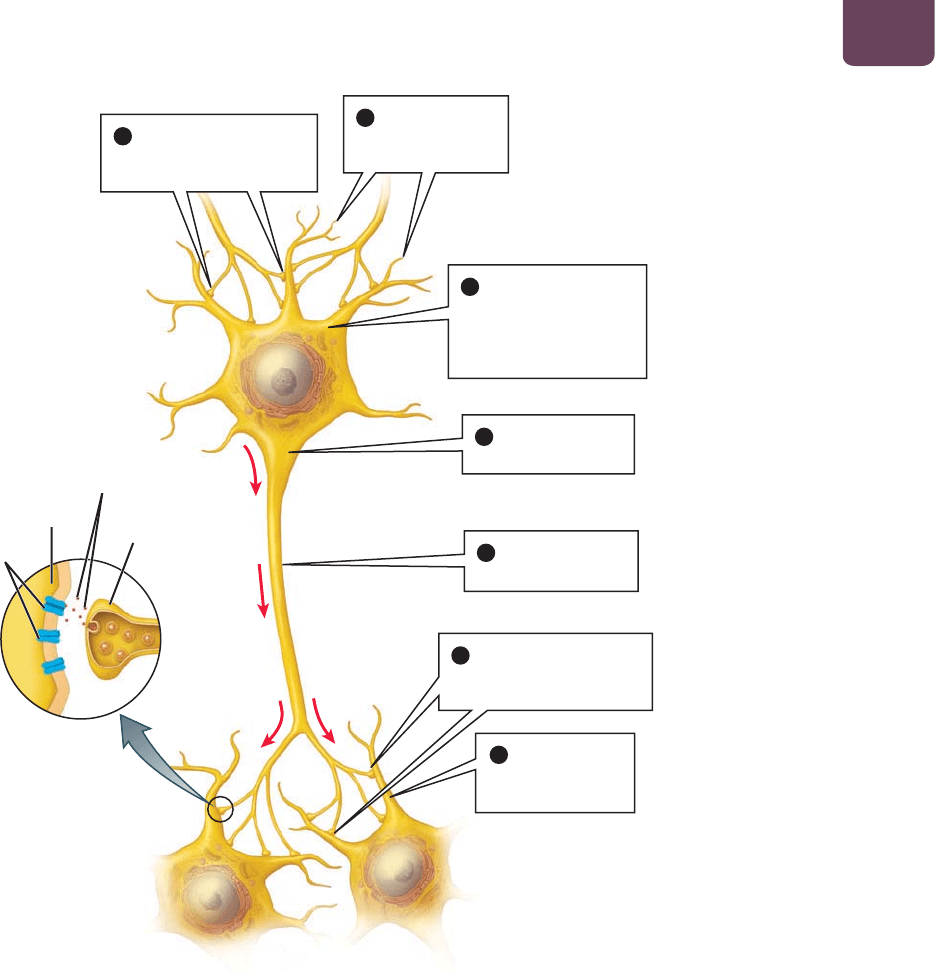
Aunque las neuronas varían en gran medida en su estruc-
tura, en la mayoría de los vertebrados éstas se encuentran confor-
madas por cuatro partes diferentes: dendritas, un cuerpo celular,
un axón y terminales sinápticas (
FIGURA 38-1), que llevan a cabo
las cuatro funciones mencionadas.
Las dendritas responden a los estímulos
Las dendritas, formadas por tallos ramificados que sobresalen del
cuerpo celular, realizan la función de “recibir la información” (
FI-
GURA 38-1
❷ y ❼). Sus ramas ofrecen un área de superficie exten-
sa para recibir las señales, ya sea del ambiente o de otras neuronas.
Las dendritas de las neuronas sensoriales tienen adaptaciones en la
membrana que les permiten producir señales eléctricas en respues-
ta a estímulos específicos del ambiente externo (como presión,
olor o luz) o interno (como temperatura corporal, pH sanguíneo
o la posición de una articulación). Las dendritas de las neuronas
en el cerebro y la médula espinal por lo regular responden a com-
puestos químicos conocidos como neurotransmisores, liberados
por otras neuronas.
El cuerpo celular procesa las señales de las dendritas
Las señales eléctricas viajan hacia las dendritas y convergen en el
cuerpo celular de las neuronas, que llevan a cabo la función de
“procesar la información” (
FIGURA 38-1 ❸). El cuerpo celular
“acumula” o integra las señales eléctricas que recibe de las dendri-
tas. Como se verá en breve, algunas de estas señales son positivas y
otras son negativas. Si su suma es lo bastante positiva, la neurona
produce una señal eléctrica extensa y rápida conocida como po-
tencial de acción (
FIGURA 38-1 ❹). El cuerpo celular contiene
también los organelos encontrados en la mayoría de las células,
como núcleo, retículo endoplasmático y aparato de Golgi, y realiza
actividades celulares típicas como la síntesis de moléculas comple-
jas y la coordinación del metabolismo celular.
Estudio de caso ¿Cómo te amo?
38.1 ¿Cuáles son las estructuras y las funciones
de las células nerviosas?
Las funciones de una neurona se localizan en partes
separadas de la célula
38.2 ¿De qué manera las neuronas producen
y transmiten la información?
Las señales eléctricas transportan la información dentro
de una sola neurona
Las neuronas utilizan sustancias químicas para comunicarse
entre sí en las sinapsis
De cerca Las señales eléctricas en las neuronas
How Neurons Work (disponible en inglés)
De cerca La transmisión sináptica
Estudio de caso continuación ¿Cómo te amo?
How Synapses Work (disponible en inglés)
38.3 ¿Cómo procesan la información los sistemas
nerviosos?
El flujo de información en el sistema nervioso requiere cuatro
operaciones básicas
De un vistazo
38.4 ¿Cómo están organizados los sistemas
nerviosos?
Los sistemas nerviosos complejos están centralizados
38.5 ¿Cuáles son las estructuras y funciones
del sistema nervioso en los seres humanos?
El sistema nervioso periférico une al sistema nervioso
central con el resto del cuerpo
El sistema nervioso central está conformado por la médula
espinal y el cerebro
La médula espinal controla muchos reflejos y conduce la
información hacia y desde el cerebro
El encéfalo se conforma de varias estructuras que realizan
funciones específicas
Guardián de la salud: Drogas, neurotransmisores
y adicción
Estudio de caso continuación ¿Cómo te amo?
El “hemisferio izquierdo” y el “hemisferio derecho”
del cerebro se especializan en distintas funciones
El aprendizaje y la memoria implican cambios bioquímicos
y estructurales en partes específicas del cerebro
Investigación científica: Neuroimagenología:
observar al cerebro en acción
Estudio de caso otro vistazo ¿Cómo te amo?
El sistema nervioso Capítulo 38 735
El axón conduce los potenciales de acción
por largas distancias
En una neurona típica, una fibra larga llamada axón se extiende
hacia fuera del cuerpo celular. El axón conduce los potenciales de
acción (
FIGURA 38-1 ❺) del cuerpo celular a las terminales sináp-
ticas en el extremo del axón, donde entra en contacto con otras
células (
FIGURA 38-1 ❶ y ❻). Los axones individuales pueden
extenderse desde tu médula espinal hasta tus dedos del pie, una
distancia aproximada de un metro, por lo que las neuronas son
las células más largas del cuerpo. Por lo regular, los axones están
unidos en los nervios, de modo muy similar a los alambres uni-
dos en un cable eléctrico. En los vertebrados, los axones unidos en
los nervios surgen del cerebro y la médula espinal y se extienden a
todas las regiones del cuerpo.
En las sinapsis, las señales son transmitidas
de una célula a otra
El sitio donde una neurona se comunica con otra célula se conoce
como sinapsis. Una sinapsis típica consiste en (1) la terminal si-
náptica, que es un ensanchamiento en el extremo de un axón de
la neurona “transmisora”; (2) una dendrita o cuerpo celular de una
neurona “receptora”, célula muscular o glandular y (3) una peque-
ña brecha que separa ambas células (
FIGURA 38-1 ❻); véase tam-
bién la figura 38-4 más adelante en este capítulo). La mayoría de
las terminales sinápticas contiene neurotransmisores liberados en
respuesta a un potencial de acción que llega a la terminal. La mem-
brana plasmática de estas neuronas receptoras porta los receptores
que se unen a los neurotransmisores y estimulan una respuesta en
esta célula. Por tanto, en una sinapsis, la salida de la primera célula
se convierte en la entrada de la segunda.
38.2
¿DE QUÉ MANERA LAS NEURONAS
PRODUCEN Y TRANSMITEN LA
INFORMACIÓN?
Aunque hay muchas excepciones, como regla general, la informa-
ción se transporta dentro de una neurona por señales eléctricas, y la
información se transmite entre las neuronas mediante neurotrans-
misores que son liberados por una neurona y recibidos por una
segunda neurona.
Dendritas:
reciben las señales
de otras neuronas
2
Cuerpo celular:
integra las señales;
coordina las actividades
metabólicas de
la neurona
3
Aquí se inicia un
potencial de acción
4
Axón: conduce el
potencial de acción
5
Dendritas (de
otras neuronas):
reciben las señales
sinapsis
dendrita
receptores
terminal
sináptica
7
Terminales sinápticas:
transmiten las señales a
otras neuronas
6
Terminales sinápticas:
transmiten las señales de
otras neuronas
1
neurotransmisores
FIGURA 38-1 Partes especializadas
y funciones de una neurona Las
flechas de color rojo indican los potenciales
de acción que pasan del cuerpo celular al
axón y las terminales sinápticas.
736 UNIDAD 5 Anatomía y fisiología animal
Las señales eléctricas transportan la información
dentro de una sola neurona
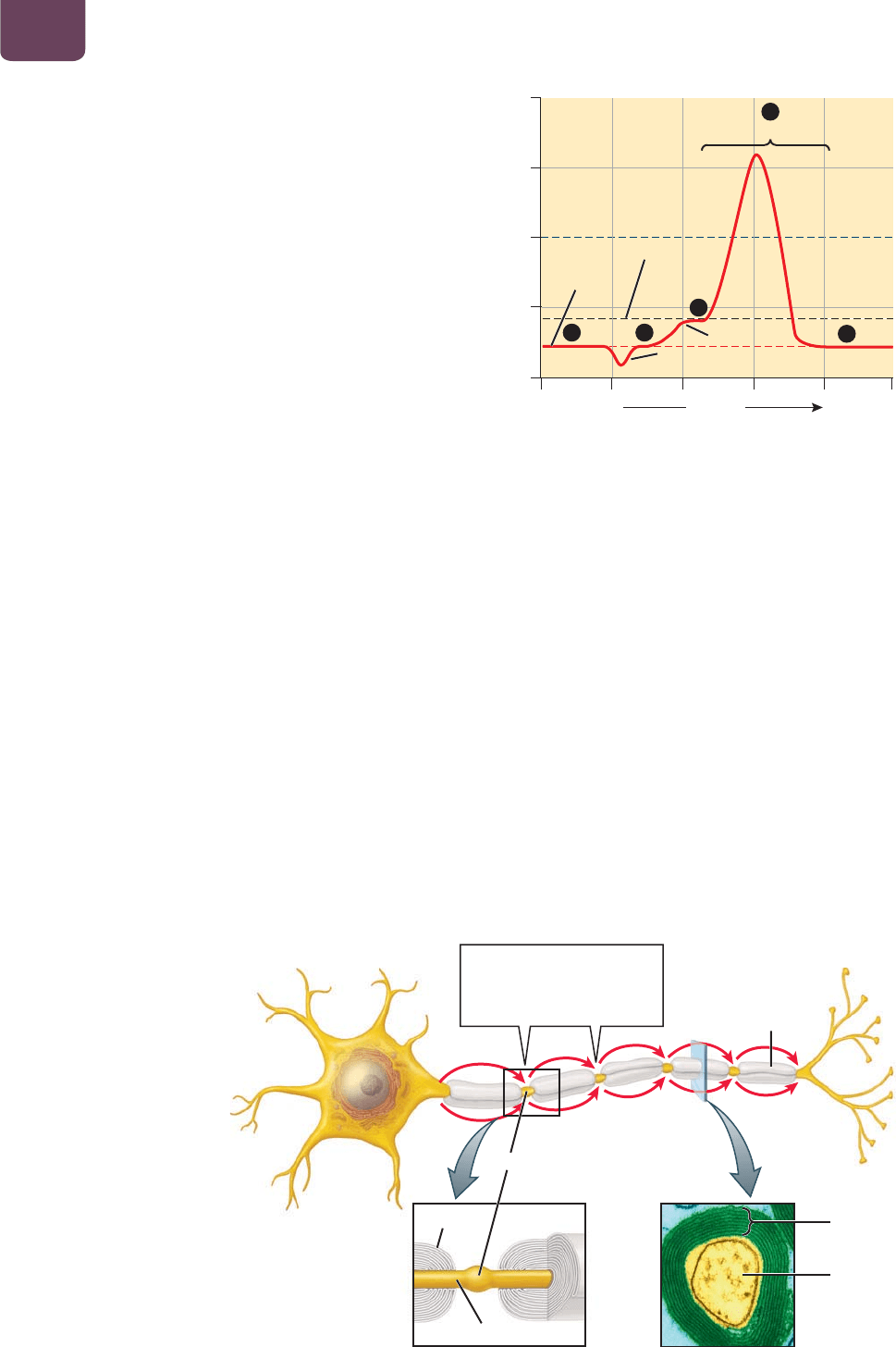
En la década de 1930, los biólogos desarrollaron formas de regis-
trar los eventos eléctricos dentro de neuronas individuales. Des-
cubrieron que una neurona inactiva no estimulada mantiene una
diferencia de voltaje eléctrico constante, o potencial, en toda su
membrana plasmática, similar al voltaje a través de los polos de
una batería. Este voltaje, llamado potencial de reposo, siempre
es negativo dentro de la célula y varía entre 40 a 90 milivoltios
(mV; milésimas de voltio) aproximadamente.
Si la neurona recibe un estímulo, ya sea natural o por una
corriente eléctrica aplicada por un investigador, el potencial en su
interior puede volverse más o menos negativo (
FIGURA 38-2). Si
el potencial se vuelve suficientemente menos negativo, alcanza
un nivel llamado umbral y produce un potencial de acción. Du-
rante un potencial de acción, el voltaje de la neurona aumenta
con rapidez a alrededor de 50 mV en el interior de la célula. Los
potenciales de acción duran unas cuantas milésimas de segundo
antes de la restauración del potencial de reposo negativo de la
célula. Las membranas plasmáticas de los axones se especializan
en la conducción de los potenciales de acción desde el cuerpo
celular de una neurona hasta las terminales sinápticas del axón.
A diferencia de los voltajes eléctricos en los alambres de metal,
que disminuyen con la distancia, los potenciales de acción son
conducidos del cuerpo celular a la terminal en el axón sin ningún
cambio en el voltaje. En la sección “De cerca: Las señales eléctri-
cas en las neuronas”, páginas 738 y 739, se estudian los mecanis-
mos celulares de los potenciales de reposo y de acción.
La mielina acelera la conducción
de los potenciales de acción
La velocidad a la que el potencial de acción viaja varía en gran me-
dida entre los axones. En general, cuanto más grueso sea el axón, el
potencial de acción se mueve con mayor rapidez. Una forma mu-
cho más eficaz de acelerar la conducción es cubrir el axón con un
aislante constituido de lípidos conocido como mielina (
FIGURA
38-3
). La mielina está formada por células gliales, oligodendrocitos
en el cerebro y la médula espinal, y células de Schwann en el resto
del cuerpo, que envuelven el axón cubriéndolo con varias capas
de membrana plasmática aislante, con muy poco citoplasma entre
sí. Cada capa de mielina cubre alrededor de 0.2 a 2 mm del axón,
dejando segmentos cortos de axón descubiertos, conocidos como
nodos. En vez de viajar de manera continua pero muy lenta por el
axón —casi siempre alrededor de 1 a 2 metros por segundo—, los
potenciales de acción en los axones cubiertos con mielina “saltan”
con rapidez de un nodo a otro, viajando a una velocidad de 3 a 100
metros por segundo (véase la figura 38-3).
FIGURA 38-2 Eventos eléctricos durante un potencial
de acción ❶ Una neurona mantiene un voltaje a través de la
membrana plasmática, conocido como potencial de reposo, de
alrededor de –60 mV en relación con el exterior. ❷ La estimulación
del ambiente o de otras células puede hacer que la neurona sea
más negativa (deflexión hacia abajo) o menos negativa (deflexión
hacia arriba). ❸ Si el potencial se vuelve alrededor de 10 a 20 mV
menos negativo, la neurona alcanza el umbral y ❹ produce un breve
potencial positivo conocido como potencial de acción. ❺ Después
de una o dos milésimas de segundo, el voltaje a través de la
membrana plasmática de la neurona regresa al potencial de reposo.
5
3
4
12
tiempo
(
milésimas de se
g
undo
)
potencial
de reposo
potencial de acción
umbral
menos
negativo
potencial
(milivoltios)
80
40
0
_
80
_
40
más
negativo
Un potencial de acción
salta de un nodo a otro,
acelerando en gran medida
la conducción en el axón
nodo
axón
mielina
cubierta
de mielina
axón
célula de Schwann
FIGURA 38-3 Un axón
recubierto con mielina Los axones
de muchos vertebrados están cubiertos
con mielina, que consiste en capas
aislantes de membranas de células
gliales especializadas. Los potenciales
de acción ocurren sólo en los nodos
entre cada capa de mielina, saltando
de un nodo a otro (flechas de color
rojo), sin tardar casi nada en viajar por
debajo de la mielina.
El sistema nervioso Capítulo 38 737
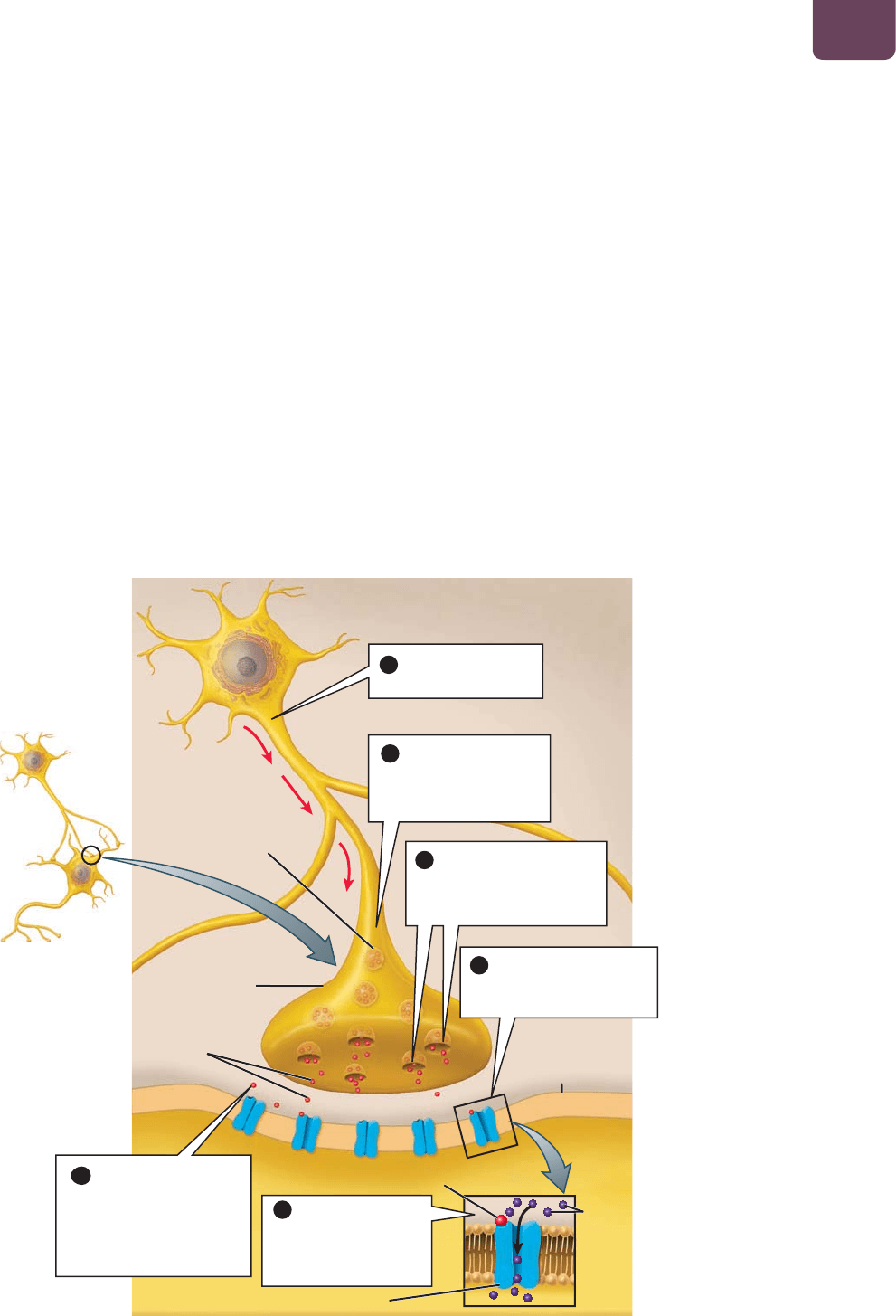
Las neuronas utilizan sustancias químicas
para comunicarse entre sí en las sinapsis
Piensa en el potencial de acción como un paquete de información
que se mueve hacia abajo del axón. Una vez que llega a la terminal
sináptica, esta información debe transmitirse a otra célula, ya sea
otra neurona o una célula en un músculo o glándula. En las que se
conocen como sinapsis eléctricas, la actividad eléctrica puede pasar
directamente de una neurona a otra a través de espacios que conec-
tan el interior de las células (véase la página 93). En el corazón, las
sinapsis eléctricas interconectan las células del músculo cardiaco,
ayudando a producir actividad eléctrica coordinada y, por tanto, la
contracción del corazón. Aunque las sinapsis eléctricas ocurren en
muchos lugares del cerebro de los mamíferos, todavía no se com-
prende bien su importancia en la función cerebral. Con mucha fre-
cuencia, las neuronas utilizan sustancias químicas para comunicar-
se entre sí o con células de músculos o glándulas. Aquí, el análisis se
limitará a estas sinapsis químicas.
En el lenguaje común, la palabra transmitir significa “enviar
algo”, y eso es exactamente lo que sucede en una sinapsis, donde
la terminal sináptica de una neurona se une a la dendrita de otra
(
FIGURA 38-4). Las dos neuronas en realidad no se tocan en una
sinapsis: un espacio diminuto, la hendidura sináptica, separa
a la primera (neurona presináptica) de la segunda (neurona
postsináptica). La neurona presináptica envía moléculas neuro-
transmisoras a través de este espacio hasta la neurona postsináp-
tica. Se sintetizan distintas neuronas y responden a una amplia
variedad de neurotransmisores. La
Tabla 38-1 enlista algunos
neurotransmisores importantes y sus funciones.
Una terminal sináptica contiene numerosas vesículas, cada
una llena de moléculas neurotransmisoras. Al iniciarse un poten-
cial de acción (
FIGURA 38-4 ❶), éste viaja a través de un axón
hasta llegar a la terminal sináptica (
FIGURA 38-4 ❷). El interior
de la terminal se carga positivamente, lo que inicia una casca-
da de cambios que hace que algunas de estas vesículas liberen
neurotransmisores en la hendidura sináptica (
FIGURA 38-4 ❸).
(En realidad, la liberación de neurotransmisores es un caso espe-
cializado de exocitosis; véanse las páginas 90 y 91). La superficie
exterior de la membrana plasmática de la neurona postsináptica,
justo del otro lado de la hendidura sináptica, está repleta de pro-
teínas receptoras que se especializan en unirse a los neurotrans-
misores liberados por la neurona presináptica. Las moléculas
neurotransmisoras se extienden por toda la hendidura y se unen
a estos receptores (
FIGURA 38-4 ❹).
FIGURA 38-4 Estructura y función
de la sinapsis
PREGUNTA Imagina un experimento en
el que las neuronas que aquí ilustramos
se bañan en una solución que contiene
veneno para los nervios. La neurona
presináptica es estimulada y produce
un potencial de acción, pero esto no da
como resultado un PPS en la neurona
presináptica. Cuando el investigador
agrega algún neurotransmisor a
la sinapsis, la neurona postsináptica sigue
sin producir ningún PPS. ¿Cómo actúa
el veneno para interrumpir la función
nerviosa?
Los neurotransmisores
se unen a los receptores en
la neurona postsináptica
dendrita de
la neurona
postsináptica
receptor
neurotransmisor
iones
4
vesícula
sináptica
hendidura sináptica
La carga positiva del
potencial de acción hace
que las vesículas sinápticas
liberen neurotransmisores
Se inicia un
potencial de acción
El potencial de
acción llega a la terminal
sináptica de la neurona
presináptica
1
2
3
Los neurotransmisores
son regresados a la
terminal sináptica, se
degradan o se extienden
hacia fuera de la
hendidura sináptica
6
terminal sináptica
de la neurona
presináptica
neurotransmisores
El neurotransmisor
hace que los canales
de iones se abran y los
iones fluyan hacia
dentro o hacia fuera
5

738 UNIDAD 5 Anatomía y fisiología animal
Las sinapsis producen potenciales postsinápticos
de excitación o de inhibición
Cabe recordar que la información por lo regular se transporta den-
tro de una neurona por medio de señales eléctricas. Estas señales
que recorren con rapidez grandes distancias desde el cuerpo celular
de una neurona hasta sus terminales presinápticas, se transportan
por potenciales de acción. Por tanto, la actividad sináptica casi
siempre altera la actividad eléctrica de la neurona postsináptica,
haciendo que la neurona sea más o menos propensa a disparar
potenciales de acción.
En casi todas las sinapsis, las proteínas receptoras en la neu-
rona postsináptica se encuentran físicamente conectadas a canales
de iones que se extienden por toda la membrana plasmática de la
neurona. Cuando las moléculas neurotransmisoras se unen a es-
tas proteínas receptoras, abren los canales de iones. Dependiendo
de qué canales estén asociados con un receptor específico, se pue-
den mover a través de ellos iones de Na
, K
, Ca
2
o Cl
(FIGURA
38-4
❺), lo cual provoca un pequeño y breve cambio en el voltaje,
conocido como potencial postsináptico (PPS). Si la neurona
postsináptica se vuelve más negativa (deflexión hacia abajo en la
figura 38-2 ❷), su potencial de reposo se aleja más del umbral, lo
que reduce la probabilidad de iniciar un potencial de acción. Este
cambio en el voltaje se conoce como potencial postsináptico de
inhibición (PPSI). Si la neurona postsináptica se vuelve menos
negativa (deflexión hacia arriba en la figura 38-2 ❷), su potencial
de reposo se acerca más al umbral y es más probable que inicie
un potencial de acción. Como consecuencia, este cambio en el
voltaje se llama potencial postsináptico de excitación (PPSE).
De cerca Las señales eléctricas en las neuronas
La permeabilidad del potasio produce
el potencial de reposo
El potencial de reposo se basa en un equilibrio entre los
gradientes químicos y eléctricos, y se mantiene mediante el
transporte activo y una membrana selectivamente permeable a
iones específicos. Los iones del citoplasma consisten sobre todo
de iones potasio con carga positiva (K
) y grandes moléculas
orgánicas con carga negativa como ATP y proteínas, que no
pueden salir de la célula (
FIGURA E38-1a, abajo). Fuera de la
célula, el líquido extracelular contiene más que nada iones sodio
con carga positiva (Na
) y iones cloruro con carga negativa
(Cl
). Las gradientes de concentración de Na
y K
se conservan
mediante una proteína de transporte activo en la membrana
plasmática llamada bomba de sodio-potasio
(Na
-K
), que bombea simultáneamente K
hacia dentro de la
célula y Na
hacia el exterior de la misma.
En una neurona no estimulada, sólo K
puede cruzar la
membrana plasmática, viajando a través de proteínas de membrana
específicas llamadas canales de K
en reposo (véase figura E38-
1a, abajo). En la membrana también hay canales de Na
y K
con
“puertas de voltaje”; como su nombre lo dice, tienen “puertas” en
sus poros que se abren o se cierran debido al voltaje que cruza
la membrana plasmática. En una neurona no estimulada, estos
canales de Na
y K
con puertas de voltaje se cierran. Más adelante
se describe la función de los canales con puertas de voltaje.
Como la concentración de K
es más alta en el interior de la
célula que en su exterior, el K
se difunde fuera de la célula a
través de los canales de K
en reposo, dejando atrás los iones
orgánicos con carga negativa (véase la figura E38-1a, abajo).
Conforme el interior de la célula adquiere una carga cada vez
más negativa, el K
es atraído eléctricamente de regreso a la
célula. Con el tiempo, el voltaje negativo dentro de la célula se
vuelve lo suficientemente alto para que el índice de K
que sale
quede exactamente equilibrado con el índice de K
que regresa
mediante la atracción eléctrica (no se muestra en la figura
E38-1a). Este voltaje negativo es el potencial de reposo.
Los cambios en la permeabilidad al sodio
y al potasio producen el potencial de acción
Los potenciales de acción ocurren cuando el potencial de reposo
cambia, volviéndose menos negativo y alcanzando el voltaje del
umbral (casi siempre alrededor de 10 a 20 mV menos negativo
que el potencial de reposo). En el umbral, los canales de Na
con
puertas de voltaje se abren, lo que permite un influjo rápido de Na
y hace que el interior de la neurona sea positivo (
FIGURA E38-1b).
Los canales de Na
con puertas de voltaje permanecen abiertos
muy poco tiempo y luego se cierran en forma espontánea.
Mientras tanto, los canales de K
con puertas de voltaje se abren,
permitiendo que el K
fluya hacia fuera de la célula, restaurando el
potencial de reposo negativo (
FIGURA E38-1c).
Los potenciales de acción son conducidos a través
de los axones sin cambiar la amplitud
Los potenciales de acción son todo o nada: si la neurona no llega
al umbral, no habrá potencial de acción; si llega al umbral, ocurrirá
un potencial de acción completo y viajará a todo lo largo del axón.
Por lo general, un potencial de acción se inicia en el punto
en que el axón sale del cuerpo celular de una neurona. Cuando
el Na
entra en el axón, su carga positiva repele otros iones
con carga positiva en el citoplasma del axón. Imagina una mesa
de billar con una docena de bolas alineadas en fila, tocándose
entre sí. Si golpeas una bola en uno de los extremos con el
taco, la bola en el extremo opuesto sale disparada al instante,
mientras que las bolas en el centro permanecen en su lugar.
De modo similar, cuando el Na
entra durante un potencial de
acción, su carga positiva repele otros iones con carga positiva
a lo largo del axón, provocando casi al instante que el potencial
de las áreas cercanas se vuelva más positivo y supere el umbral.
Esto hace que los canales de Na
en estas áreas cercanas se
abran, iniciando un nuevo potencial de acción. El Na
entra en
estos nuevos lugares ligeramente más allá de ese punto en el
axón (véase la figura E38-1c, arriba), iniciando otra vez todo
el proceso. Como un nuevo potencial de acción completo se
produce una y otra vez a lo largo de todo el axón, éste viaja
hasta su extremo sin perder voltaje.
Cuando la onda de cargas positivas pasa por un punto
determinado a lo largo del axón, se restaura el potencial de
reposo mientras los canales de K
con puertas de voltaje se
abren y el K
fluye hacia el exterior (véase la figura E38-1c).
Sólo se intercambia una fracción diminuta del total de K
y
Na
dentro y alrededor de la neurona durante cada potencial de
acción, de modo que los gradientes de concentración de K
y
Na
no cambian de manera notoria. A largo plazo (de minutos
a horas), la actividad de la bomba de Na
-K
mantiene la
concentración de gradientes de ambos iones.
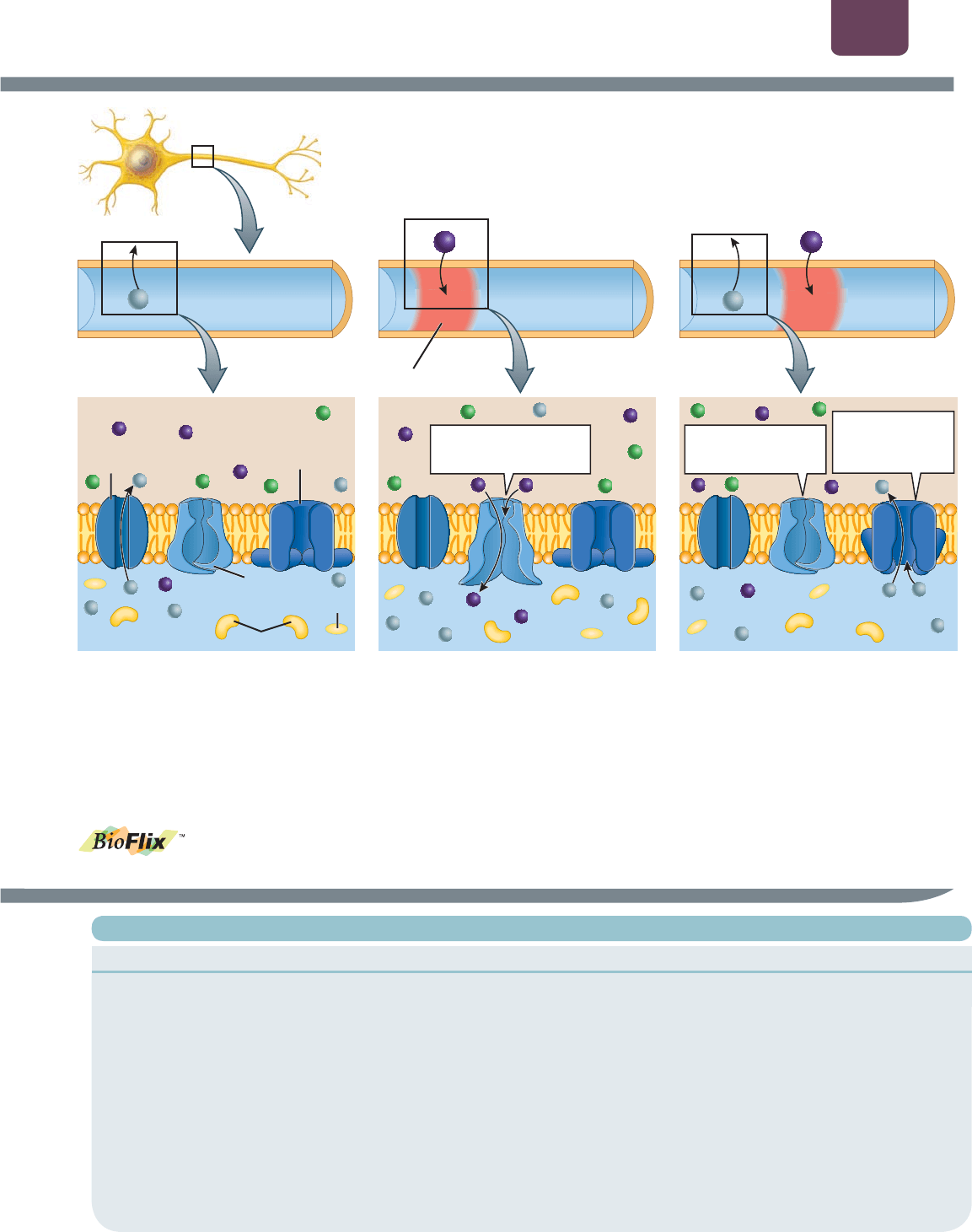
El sistema nervioso Capítulo 38 739
canal de K
ⴙ
en reposo
(siempre abierto)
canal de Na
ⴙ
con
puertas de voltaje
(cerrado)
canal de K
ⴙ
con
puertas de voltaje
(cerrado)
ⴙⴙⴚⴚ ⴚⴚⴚⴚⴚⴚⴚ ⴚ ⴚⴚⴚ ⴙⴙ ⴚⴚⴚ ⴚⴚ
Cl
Cl
Cl
Cl
Cl
Cl
Cl
Cl
Cl
Cl
Cl
K
K
K
K
K
K
K
K
K
K
K
K
K
K
K
K
K
Na
Na
Na
Na
Na
Na
Na
Na
Na
Na
Na
Na
Na
Na
(a) El potencial de reposo
(b) El potencial de acción
K
K
(c) Se restaura el potencial de reposo
Se abre un canal de Na
ⴙ
con puertas de voltaje
Se cierra el canal de
Na
ⴙ
con puertas de
voltaje
Se abre el canal
de K
ⴙ
con
puertas de voltaje
(citoplasma de la neurona)
(líquido extracelular)
ATP
(interior de
la neurona)
proteínas
potencial de acción
Na Na
FIGURA E38-1 Los mecanismos iónicos subyacentes a los potenciales de reposo y acción Las ilustraciones
superiores en cada parte de la figura muestran una sección de un axón, con los movimientos iónicos importantes a través de
la membrana plasmática durante (a) el potencial de reposo, (b) la fase en aumento de un potencial de acción y (c) la fase en
disminución de un potencial de acción que ocurre en la parte del axón en el cuadro. En la ilustración superior de la parte (c),
se inició un nuevo potencial de acción, en una parte más alejada del axón (el Na
entra en el axón). Las ilustraciones inferiores
muestran la distribución de los iones dentro y fuera del axón, los canales iónicos importantes que controlan los potenciales de
reposo y acción, y los movimientos de los iones a través de los canales en la parte del axón en el cuadro.
Tabla 38-1 Algunos neurotransmisores importantes
Neurotransmisor Ubicación en el sistema nervioso Algunas funciones importantes
acetilcolina Sinapsis entre neuronas motoras y músculos; sistema
nervioso autónomo, muchas áreas del cerebro
Activa los músculos esqueléticos; activa los órganos
blanco del sistema nervioso parasimpático
Dopamina Mesencéfalo Importante en las emociones, los placeres y el
control del movimiento
Norepinefrina (noradrenalina) Sistema nervioso simpático Activa los órganos blanco del sistema nervioso
simpático
Serotonina Mesencéfalo, puente de Varolio y bulbo raquídeo Influye en el humor y en el sueño
Glutamato Muchas áreas del cerebro y médula espinal Principal neurotransmisor de excitación en el
sistema nervioso central
Glicina Médula espinal Principal neurotransmisor de inhibición en la médula
espinal
GABA (ácido gamma aminobutírico) Muchas áreas del cerebro y médula espinal Principal neurotransmisor de inhibición en el cerebro
Endorfinas Muchas áreas del cerebro y médula espinal Influye en el humor, reduce las sensaciones de dolor
Óxido nítrico Muchas áreas del cerebro Importante en la formación de recuerdos
How Neurons Work (disponible en
inglés)
740 UNIDAD 5 Anatomía y fisiología animal
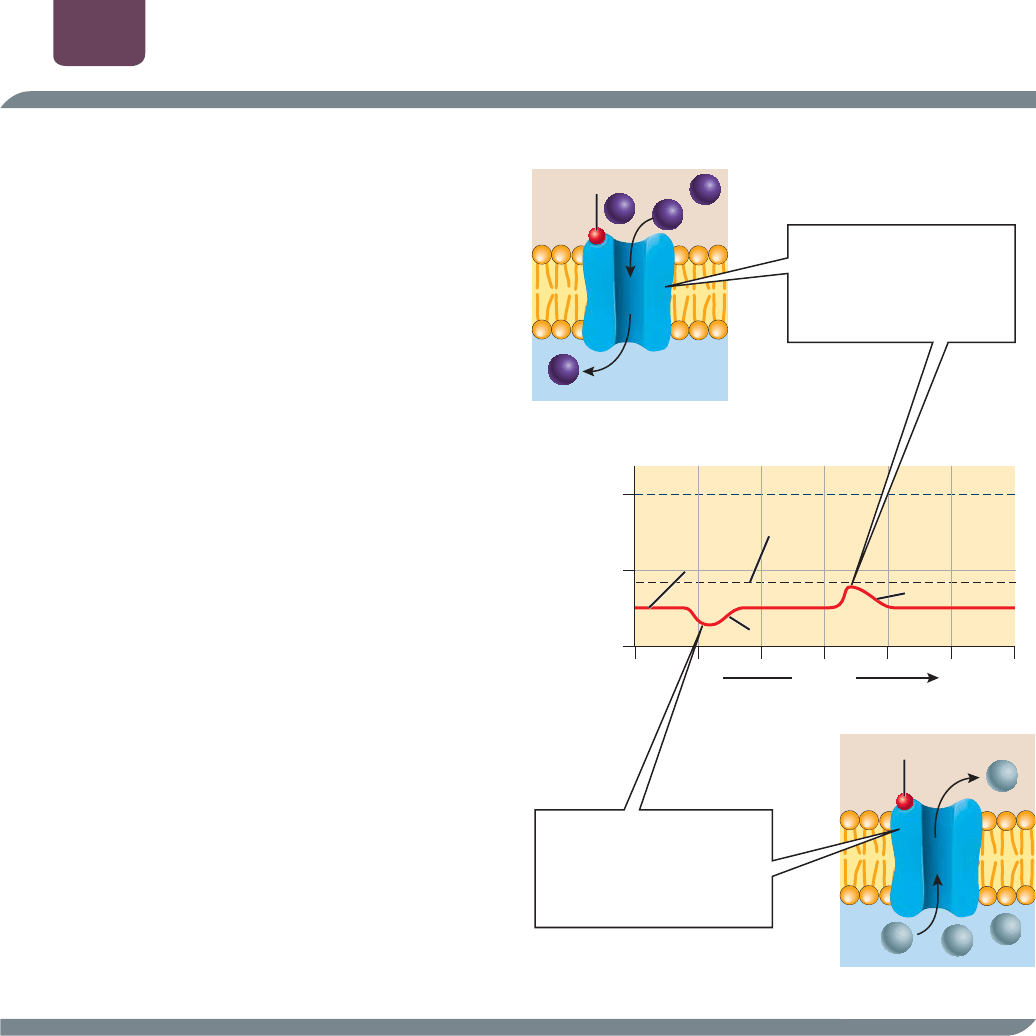
De cerca La transmisión sináptica
Cuando un potencial de acción llega a una terminal
presináptica, la carga positiva dentro de la terminal abre
un nuevo grupo de canales de iones con puertas de
voltaje, los cuales son selectivamente permeables al calcio
(Ca
2
). La concentración de Ca
2
fuera de la terminal es
aproximadamente 10 mil veces más alta que la concentración
en el interior. Por tanto, el Ca
2
entra en la terminal,
donde activa una serie de proteínas que provocan que los
neurotransmisores que contienen vesículas se fundan con la
membrana presináptica y liberen su neurotransmisor en la
hendidura sináptica.
Los neurotransmisores se dispersan a través de la
hendidura y se unen a las proteínas receptoras en la célula
postsináptica. Por lo regular, lo anterior tiene uno de
dos efectos. En algunas sinapsis, el resultado de la unión
neurotransmisor-receptor es similar a lo que sucede cuando
una hormona péptida se une a su receptor (véase la figura
37-2): se sintetizan los mensajeros intracelulares, y cambia
el metabolismo de la célula postsináptica. Sin embargo, en la
mayoría de las sinapsis, las proteínas receptoras se encuentran
unidas a canales de iones, y la unión de neurotransmisores
abre los canales (
FIGURA E38-2).
Si los canales son permeables al Na
(FIGURA E38-2a), el Na
se difunde a lo largo de su gradiente de concentración hacia
la neurona postsináptica, haciendo que la célula sea menos
negativa. Si la neurona postsináptica se vuelve suficientemente
menos negativa, puede llegar al umbral y producir un potencial
de acción. Como “excitan” a la célula postsináptica, estos
cambios de voltaje se conocen como potenciales postsinápticos
de excitación (PPSE).
Si los canales son permeables al K
(FIGURA E38-2b), el
K
se difunde fuera de la célula, volviéndola más negativa. El
hecho de que la célula sea más negativa inhibe la producción
de potenciales de acción en la célula postsináptica, de modo
que el cambio de voltaje se llama potencial postsináptico de
inhibición (PPSI).
neurotransmisor
K
ⴙ
K
ⴙ
K
ⴙ
K
ⴙ
Na
ⴙ
Na
ⴙ
neurotransmisor
(a) Potencial postsináptico de excitación (PPSE)
(
b
)
Potencial
p
ostsiná
p
tico de inhibición
(
PPSI
)
La unión del neurotransmisor
abre un canal iónico
permeable al Na
ⴙ
; el Na
ⴙ
entra en la neurona
postsináptica, motivando
un PPSE
La unión del neurotransmisor
abre un canal iónico
permeable al K
ⴙ
; el K
ⴙ
sale
de la neurona postsináptica,
activando un PPSI
tiempo
(milésimas de segundo)
potencial
de reposo
umbral
potencial
(milivoltios)
0
_
80
_
40
PPSI
PPSE
Na
ⴙNa
ⴙ
FIGURA E38-2 La unión del neurotransmisor a las
proteínas receptoras abre los canales de iones (a) El
mecanismo iónico de un PPSE. (b) El mecanismo iónico de un PPSI.
En la sección “De cerca: La transmisión sináptica” se explican los
mecanismos mediante los cuales la unión de neurotransmisores a
receptores provoca PPS.
La acción de los neurotransmisores
por lo general es breve
Piensa qué sucedería si una neurona presináptica empezara a es-
timular una célula postsináptica y nunca dejara de hacerlo. Por
ejemplo, podrías contraer los bíceps, flexionar el brazo y ¡dejarlo
flexionado para siempre! No resulta sorprendente que el sistema
nervioso tenga varias formas de concluir una acción neurotrans-
misora. Algunos neurotransmisores, sobre todo la acetilcolina —el
transmisor que estimula las células de los músculos esqueléticos—
son degradados en poco tiempo por las enzimas en la hendidu-
ra sináptica. Muchos otros se transportan de regreso a la neurona
presináptica (
FIGURA 38-4 ❻). Para todos los neurotransmisores,
estos mecanismos están complementados por la difusión fuera de
la hendidura sináptica.
La suma de los potenciales postsinápticos
determina la actividad de una neurona
La mayoría de los potenciales postsinápticos son pequeñas señales
que desaparecen en poco tiempo, pero que viajan la distancia sufi-
ciente para llegar al cuerpo celular. Ahí, determinan si se produce
un potencial de acción. ¿Cómo? Las dendritas y el cuerpo celular
de una sola neurona a menudo reciben PPSE y PPSI de las termi-
nales sinápticas de miles de neuronas presinápticas. Los voltajes
de todos los PPS que llegan al cuerpo de la célula postsináptica se
suman simultáneamente, proceso conocido como integración. Si
al sumarse los potenciales postsinápticos de excitación e inhibi-
El sistema nervioso Capítulo 38 741
ción elevan el potencial eléctrico dentro de la neurona por encima
del umbral, la célula postsináptica produce un potencial de acción.
How Synapses Work (disponible en
inglés)
38.3
¿CÓMO PROCESAN LA INFORMACIÓN
LOS SISTEMAS NERVIOSOS?
La neurona individual utiliza un lenguaje de potenciales de acción
sencillo; no obstante, este lenguaje básico permite a los animales
realizar comportamientos complejos. Una clave sobre la versatili-
dad del sistema nervioso es la presencia de redes de neuronas que
van desde docenas hasta millones de células. Como en las compu-
tadoras, elementos sencillos pueden llevar a cabo tareas sorpren-
dentes si se conectan de manera apropiada.
El flujo de información en el sistema nervioso
requiere cuatro operaciones básicas
Como mínimo, un sistema nervioso debe ser capaz de realizar cua-
tro operaciones:
1. Determinar el tipo de estímulo.
2. Determinar e indicar la intensidad de un estímulo.
3. Integrar la información de diversas fuentes.
4. Iniciar y dirigir respuestas apropiadas.
La naturaleza de un estímulo está determinada
por las conexiones entre los sentidos y el cerebro
Si los potenciales de acción son unidades de información de casi
todas las neuronas, y si todos los potenciales de acción son bási-
camente iguales, ¿de qué manera el cerebro determina qué es un
estímulo (luz, sonido, hambre) o qué tan fuerte es el estímulo?
Todos los sistemas nerviosos interpretan qué es un estímu-
lo monitoreando qué neuronas disparan los potenciales de acción.
Por ejemplo, tu cerebro interpreta los potenciales de acción que
ocurren en los axones de tus nervios ópticos (que se originan en
el ojo y viajan hasta las áreas de la visión en el cerebro) como la
sensación de luz. Se dice que un fisiólogo alemán se sentó un día
en una habitación oscura y se picó el ojo, lo que causó un ligero
daño en la retina y produjo potenciales de acción que viajaron
hasta el cerebro. (Como dicen los anuncios en televisión que
muestran automóviles a toda velocidad en pistas de carreras, ¡no
intentes esto en casa!) ¿El resultado? “Vio estrellas” porque su
cerebro interpretó como luz los potenciales de acción en su ner-
vio óptico. De ahí que distingas el sonido de la música del sabor
del café, o el sabor amargo de éste de lo dulce del azúcar, porque
estos estímulos diferentes dan como resultado potenciales de ac-
ción en distintos axones que se conectan a diversas áreas de tu
cerebro.
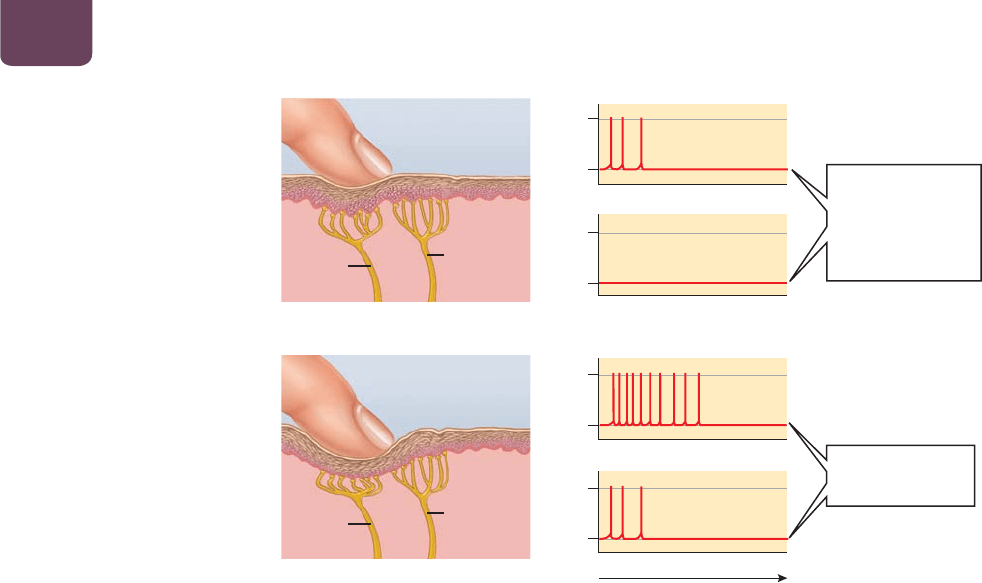
La intensidad de un estímulo se codifica por la frecuencia
de los potenciales de acción
Como todos los potenciales de acción son casi del mismo tamaño
y duración, no es posible codificar la información sobre la fuerza,
o intensidad, de un estímulo (como el volumen de un sonido) en
un solo potencial de acción. En vez de ello, la intensidad se codi-
fica de otras dos maneras (
FIGURA 38-5). Primero, es posible que
la intensidad esté señalada por la frecuencia de los potenciales de
acción en una sola neurona. Cuanto más intenso sea el estímulo, la
neurona disparará con mayor rapidez los potenciales de acción. En
segundo lugar, la mayoría de los sistemas nerviosos tienen muchas
neuronas que pueden responder a la misma información. Los estí-
mulos más fuertes excitan a una mayor cantidad de estas neuronas,
mientras que los estímulos más débiles excitan a menos neuro-
nas. Por tanto, también es posible señalar la intensidad mediante
el número de neuronas similares que se activan al mismo tiempo.
Un toque muy suave puede hacer que un solo receptor en tu piel
dispare potenciales de acción muy lentamente (
FIGURA 38-5a); un
pinchazo fuerte puede hacer que se activen varios receptores del
tacto, algunos de ellos con mucha rapidez (
FIGURA 38-5b).
El sistema nervioso procesa información de varias fuentes
Tu cerebro es bombardeado en forma continua por estímulos
sensoriales provenientes del interior y el exterior del cuerpo. El
cerebro debe evaluar estos datos, determinar cuáles son impor-
tantes y decidir cómo responder. Los sistemas nerviosos integran
información de varias fuentes: muchas neuronas pueden enviar
sus señales a unas cuantas neuronas. Por ejemplo, muchas neu-
ronas sensoriales pueden converger en un número menor de
células cerebrales. Algunas de estas células del cerebro actúan
como “células que toman decisiones”, sumando los potenciales
postsinápticos que resultan de la actividad sináptica de las neu-
ronas sensoriales. Dependiendo de su fuerza relativa (y de otros
factores internos como las hormonas o la actividad metabólica),
producen las acciones apropiadas.
El sistema nervioso produce salidas
a los músculos y las glándulas
Los potenciales de acción de las neuronas que toman decisiones
pueden viajar a otras partes del cerebro, a la médula espinal o a
los sistemas nerviosos simpático y parasimpático (que se describen
más adelante). Con el tiempo, la información del sistema nervioso
estimula la actividad en los músculos o glándulas que producen
comportamientos.
Los mismos principios de conectividad y codificación de
la intensidad que se acaban de mencionar para la información
sensorial se utilizan para los datos procedentes del cerebro. Los
músculos o glándulas activados se determinan por sus conexio-
nes con el cerebro o la médula espinal. Por ejemplo, las neuronas
que activan tus bíceps son diferentes a las que activan los múscu-
los en tu cara. La dureza con la que un músculo se contrae es
determinada por la cantidad de neuronas conectadas a éste y la
rapidez con la que estas neuronas disparan potenciales de acción.
Estudio de caso continuación
¿Cómo te amo?
Diferentes especies animales e incluso distintos individuos
en una especie tienen diferentes números y tipos de
receptores para neurotransmisores. Los Microtus ochrogastes
monógamos, por ejemplo, tienen una densidad muy
alta de receptores de dopamina, el neurotransmisor de
recompensa, en ciertas partes del cerebro. Por el contrario,
Microtus montanus, otra especie de ratón de campo, tiene
concentraciones de receptores de dopamina mucho más bajas
en estas regiones del cerebro. Entre los seres humanos, la
gente con fobias sociales tiene menos receptores de dopamina
en las áreas de recompensa del cerebro que aquellos con una
interacción social adecuada.
742 UNIDAD 5 Anatomía y fisiología animal
38.4
¿CÓMO ESTÁN ORGANIZADOS
LOS SISTEMAS NERVIOSOS?
La mayoría de las conductas están controladas por rutas compues-
tas por cuatro elementos:
1. Las neuronas sensoriales responden a un estímulo, ya sea
interno o externo al cuerpo.
2. Las interneuronas reciben señales de las neuronas senso-
riales, hormonas, neuronas que almacenan recuerdos y mu-
chas otras fuentes. Con base en esta información, a menudo
las interneuronas activan las neuronas motrices.
3. Las neuronas motrices reciben instrucciones de las neuronas
sensoriales o interneuronas y activan músculos o glándulas.
4. Los efectores, que por lo general son músculos o glándulas,
llevan a cabo la respuesta dirigida por el sistema nervioso.
Las conductas sencillas, como los reflejos (véase la sección
38.5), pueden ser controladas por la actividad en tan sólo dos
o tres neuronas (una neurona sensorial, una neurona motriz y
quizá una interneurona intermedia), estimulando posteriormen-
te un solo músculo. Muchos reflejos no utilizan el cerebro para
nada. En los seres humanos, los reflejos simples, como la cono-
cida reacción refleja de la rodilla o los reflejos de retiro ante el
dolor, son producidos por las neuronas en la médula espinal.
Las conductas complejas están organizadas por rutas neu-
rales interconectadas, en las que varios tipos de información sen-
sorial (con recuerdos, hormonas y otros factores) convergen en
un grupo de interneuronas. Mediante la integración de potencia-
les postsinápticos de varias fuentes, las interneuronas “deciden”
qué hacer y estimulan a las neuronas motrices para dirigir la ac-
tividad apropiada en músculos y glándulas. Tal vez se requieran
cientos o incluso millones de neuronas, sobre todo en el cerebro,
para realizar acciones complejas como tocar el piano, pero los
principios siguen siendo los mismos.
Los sistemas nerviosos complejos
están centralizados
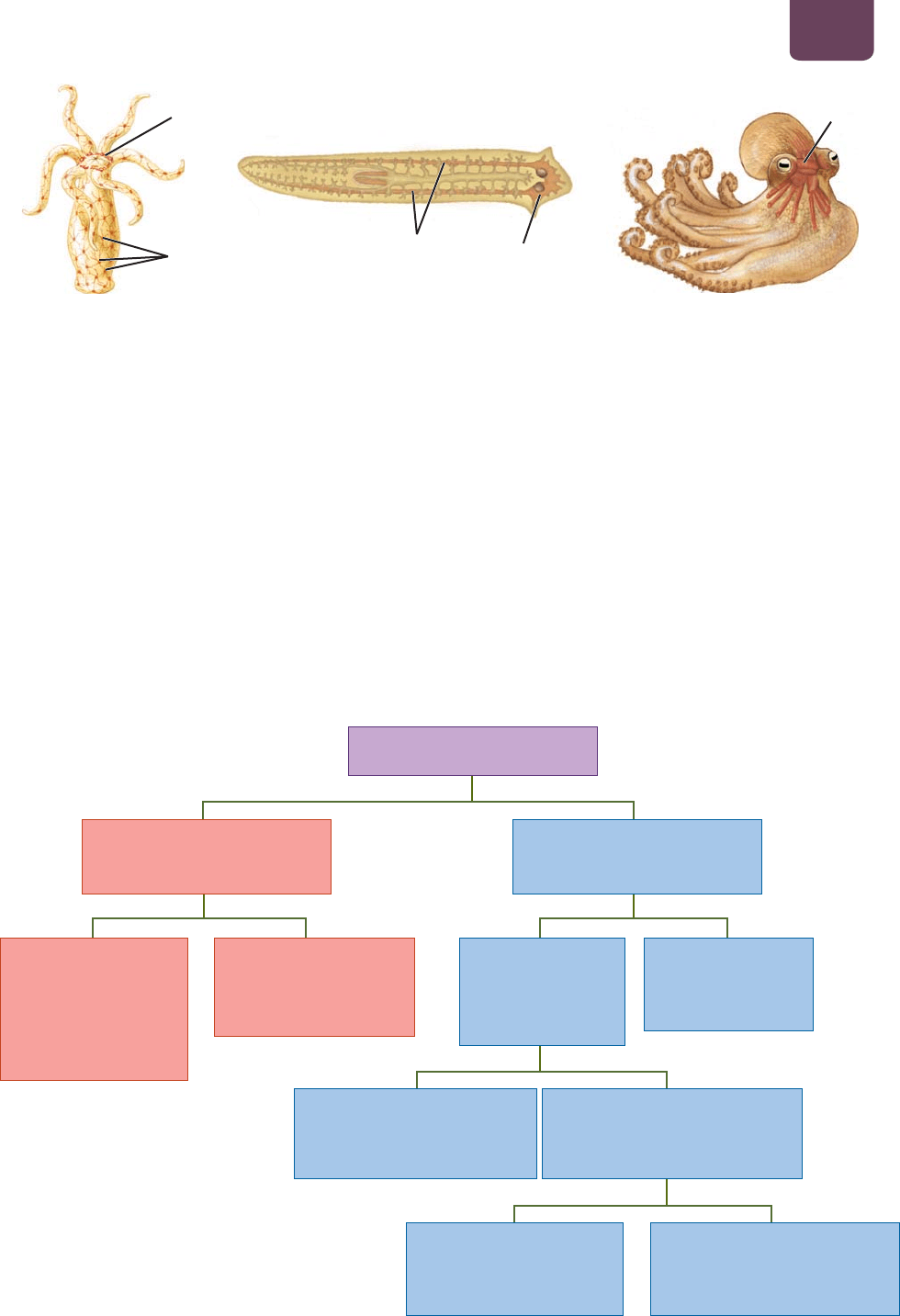
En el reino animal, en realidad hay sólo dos diseños de sistemas
nerviosos: un sistema nervioso difuso, como el de los cnidarios
(Hydra, medusas y demás organismos del filum;
FIGURA 38-6a),
y un sistema nervioso centralizado, encontrado en diversos grados
en organismos más complejos.
No resulta sorprendente que el diseño del sistema nervioso
esté muy relacionado con el estilo de vida de un animal. Los cni-
darios que presentan simetría radial (véase la figura 23-2) no tie-
nen “frente”, de modo que no ha existido una presión evolutiva
para concentrar los sentidos en un lugar. Por ejemplo, una Hydra
se ancla en una roca en el fondo de un estanque, de modo que es
igualmente probable que la presa o los depredadores vengan de
una misma dirección. Los sistemas nerviosos de los cnidarios es-
tán compuestos por una red de neuronas, a menudo llamada red
nerviosa, entretejida a través de los tejidos del animal. Por todas
partes se encuentran neuronas agrupadas, lo que se conoce como
ganglio, pero nada parecido a un verdadero cerebro.
Casi todos los demás animales presentan simetría bilateral
con extremos definidos de cabeza y cola. Como por lo regular la
cabeza es la primera parte del cuerpo que se enfrenta a la comida,
el peligro y los compañeros potenciales, resulta ventajoso tener
los órganos de los sentidos concentrados en ella. Con la evolu-
ción se desarrollaron ganglios grandes con la función de integrar
la información recolectada por los sentidos y dirigir las acciones
apropiadas. A través del tiempo evolutivo, los principales órganos
de los sentidos de los animales con sistemas nerviosos complejos
se localizaron en la cabeza, y los ganglios se centralizaron en un ce-
rebro. Esta tendencia, llamada cefalización, se observa con claridad
en los invertebrados (
FIGURAS 38-6b, c). La cefalización alcanza
su máxima expresión en los vertebrados, en los que casi todos los
cuerpos celulares del sistema nervioso se encuentran en el cerebro
y la médula espinal.
FIGURA 38-5 Intensidad de los
estímulos de las señales La intensidad
de un estímulo está determinada por
la velocidad a la que las neuronas
sensoriales individuales producen
potenciales de acción, así como por
el número de neuronas sensoriales
activadas. (a) En este ejemplo, un ligero
toque activa sólo la neurona sensorial
más cercana, que dispara los potenciales
de acción a baja velocidad. (b) Un fuerte
pinchazo activa también las neuronas
sensoriales cercanas, provocando que la
más próxima se active con rapidez y
las más distantes con mayor lentitud.
PREGUNTA Con base en esta
información, ¿en qué crees que se
diferencian las áreas de la piel que son
especialmente sensibles al tacto con
respecto a las áreas menos sensibles?
⫺70
⫹40
⫺70
⫹40
(a) Toque suave
neurona
sensorial 1
neurona
sensorial 2
neurona
sensorial 1
neurona
sensorial 2
neurona sensorial 1
neurona sensorial 2
tiempo
neurona sensorial 2
(b) Pinchazo fuerte
neurona sensorial 1
⫺70
⫹40
⫺70
⫹40
La neurona
sensorial 1
se activa con
lentitud; la neurona
sensorial 2
es silenciosa
Las neuronas
sensoriales 1 y 2
se activan
El sistema nervioso Capítulo 38 743
38.5
¿CUÁLES SON LAS ESTRUCTURAS
Y FUNCIONES DEL SISTEMA NERVIOSO
EN LOS SERES HUMANOS?
El sistema nervioso de todos los mamíferos, incluidos los seres hu-
manos, se puede dividir en dos partes: central y periférico. Cada
una de las cuales tiene otras subdivisiones (
FIGURA 38-7). El siste-
ma nervioso central (SNC) está conformado por el cerebro y la
médula espinal. El sistema nervioso periférico (SNP) está inte-
grado por neuronas situadas fuera del SNC y axones que conectan
a estas neuronas con el SNC. Los cuerpos celulares de las neuronas
en el SNP se localizan a menudo en los ganglios junto a la médula
espinal (véanse las figuras 38-9 y 38-10 más adelante) o en los gan-
glios cerca de los órganos blanco, como los ganglios en la cabeza y
el cuello que controlan las glándulas salivales (véase la figura 38-8).
El sistema nervioso periférico une al sistema
nervioso central con el resto del cuerpo
Los nervios del sistema nervioso periférico conectan el cerebro y la
médula espinal con el resto del cuerpo, incluidos músculos, glán-
dulas, órganos sensoriales y los sistemas digestivo, respiratorio,
urinario, reproductor y circulatorio. Los nervios periféricos contie-
nen axones de neuronas sensoriales, los cuales llevan información
anillo de ganglios
red de neuronas
difusa
médulas nerviosas
ganglios
cerebrales
(cerebro)
cerebro
(a) Hydra
(b) Gusano plano (c) Pulpo
FIGURA 38-6 Organización del sistema nervioso (a) El sistema nervioso difuso de la Hydra contiene unas
cuantas concentraciones de neuronas en la base de los tentáculos, pero no tiene cerebro. Las señales neurales
se conducen en todas las direcciones por todo el cuerpo. (b) El gusano plano tiene un sistema nervioso menos
difuso, con un grupo de ganglios en la cabeza. (c) El pulpo tiene un cerebro grande y complejo, y sus capacidades
para el aprendizaje rivalizan con aquellas de algunos mamíferos.
Neuronas sensoriales
Sistema nervioso autónomo
El sistema nervioso
transportan las señales
al SNC desde los
órganos sensoriales
controla las respuestas involuntarias
influyendo en los órganos,
las glándulas y los músculos lisos
Sistema nervioso central (SNC)
recibe y procesa la información;
acción iniciada
Cerebro
recibe y procesa la
información sensorial;
inicia las respuestas;
almacena los recuerdos;
genera pensamientos
y emociones
Médula espinal
conduce las señales hacia
y desde el cerebro; controla
las actividades reflejas
Neuronas motrices
transportan las señales
del SNC que controlan
las actividades de
músculos y glándulas
Sistema nervioso periférico (SNP)
transmite las señales entre
el SNC y el resto del cuerpo
Sistema nervioso somático
controla los movimientos
voluntarios mediante la activación
de los músculos esqueléticos
División simpática
prepara el cuerpo para
las actividades estresantes o
energéticas; “pelear o huir”
División parasimpática
domina durante los momentos
de “descansar y digerir”; dirige
las actividades de mantenimiento
FIGURA 38-7 Organización y funciones del sistema nervioso de los vertebrados

744 UNIDAD 5 Anatomía y fisiología animal
sensorial al sistema nervioso central de todas las partes del cuerpo.
(Quizá pienses que estas fibras deberían llamarse dendritas, por-
que transportan información hacia el cuerpo celular. Sin embargo,
los neurobiólogos las llaman axones porque son largas y conducen
potenciales de acción.) Los nervios periféricos contienen también
los axones de las neuronas motrices que transportan señales del
sistema nervioso central a las glándulas y músculos. La porción
motriz del sistema nervioso periférico está formada por dos partes:
el sistema nervioso somático y el sistema nervioso autónomo.
El sistema nervioso somático controla
el movimiento voluntario
Las neuronas motrices del sistema nervioso somático forman
sinapsis con los músculos esqueléticos y controlan el movimiento
voluntario. Cuando tomas notas, levantas una taza con café o ajus-
tas tu reproductor de música, tu sistema nervioso somático está a
cargo. Los cuerpos celulares de las neuronas motrices somáticas se
localizan en la médula espinal (véase la figura 38-10 más adelante).
Sus axones van directo a los músculos que controlan. En el capítulo
40 se estudian los músculos.
El sistema nervioso autónomo controla
las acciones involuntarias
Las neuronas motrices del sistema nervioso autónomo inervan
el corazón, los músculos lisos y las glándulas, y producen más que
nada acciones involuntarias. El sistema nervioso autónomo es con-
trolado sobre todo por el hipotálamo, la médula y el puente de
Varolio, partes del cerebro que se describen más adelante en este
capítulo. Se divide en dos: la división simpática y la división
parasimpática. Las dos divisiones del sistema nervioso autónomo
inervan la mayoría de los mismos órganos, pero casi siempre pro-
ducen efectos opuestos (
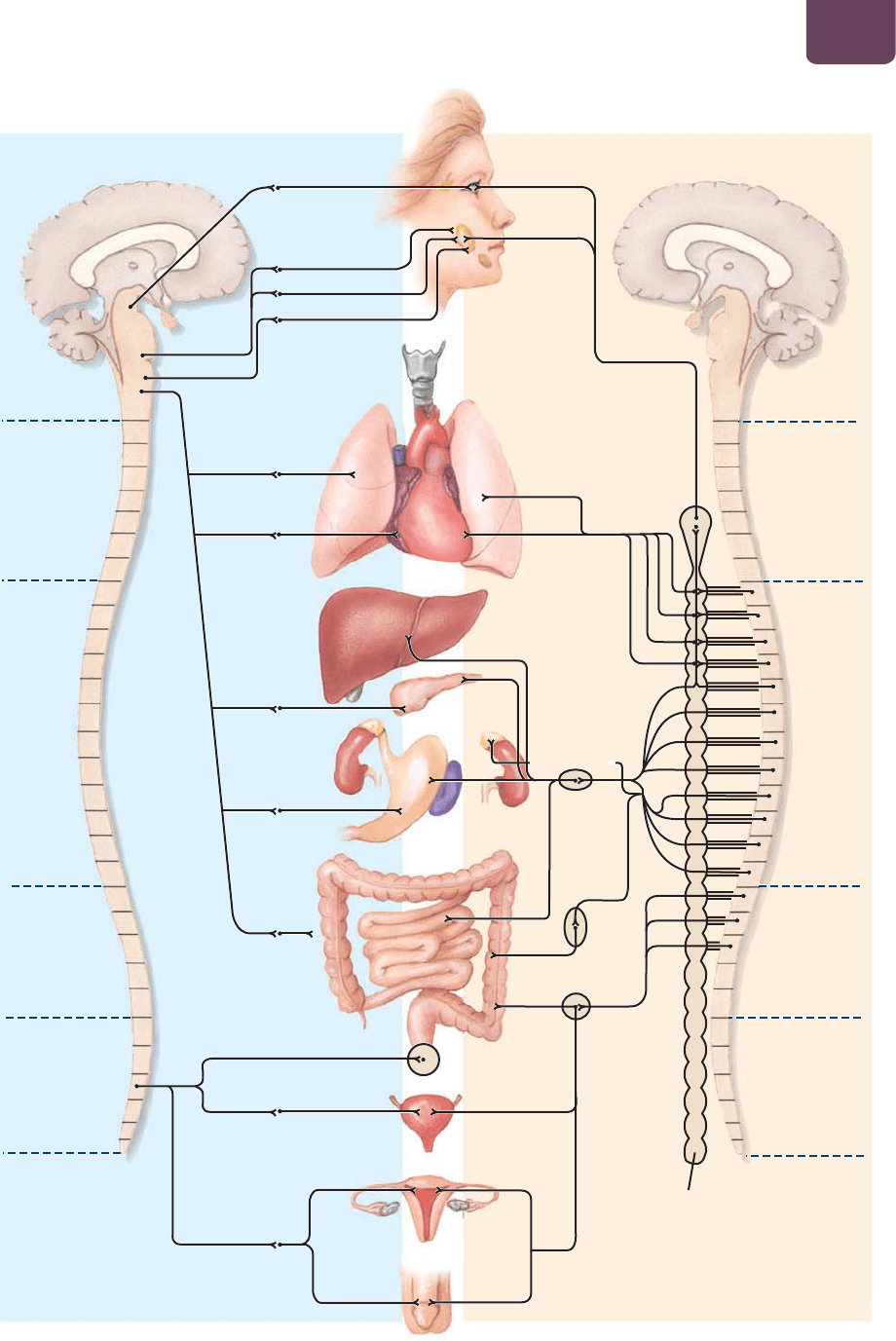
FIGURA 38-8).
Las neuronas de la división simpática liberan la norepinefri-
na (noradrenalina), un neurotransmisor hacia sus órganos blanco,
preparando al cuerpo para actividades estresantes o energéticas,
como pelear, escapar o presentar un examen. Durante estas acti-
vidades de alerta, “pelear o huir”; el sistema nervioso simpático
reduce la actividad en el tracto digestivo, redirigiendo parte del
suministro de sangre a los músculos de brazos y piernas. El ritmo
cardiaco se acelera. Las pupilas de los ojos se agrandan, admitien-
do más luz, y las vías respiratorias en los pulmones se expanden,
dejando entrar más aire. Esto también puede suceder si de repen-
te te piden que respondas una pregunta en clase, ¡sobre todo si
no conoces la respuesta!
Las neuronas de la división parasimpática liberan acetilco-
lina hacia sus órganos blanco. La división parasimpática domina
durante las actividades de mantenimiento que es posible llevar a
cabo en el tiempo libre, conocidas a menudo como “descansar y
digerir”. Bajo el control parasimpático, el tracto digestivo se acti-
va, el ritmo cardiaco se hace más lento y las vías respiratorias en
los pulmones se contraen, porque el cuerpo necesita de un menor
flujo de sangre y menos oxígeno.
El sistema nervioso central está conformado
por la médula espinal y el cerebro
La médula espinal y el cerebro constituyen el sistema nervioso
central (SNC). El SNC recibe y procesa la información sensorial,
genera los pensamientos y dirige las respuestas. El SNC consiste
sobre todo de interneuronas; ¡probablemente alrededor de 100
mil millones!
El cerebro y la médula espinal se encuentran protegidos del
daño físico de tres maneras. La primera línea de defensa es una
armadura ósea, que consiste en el cráneo, que rodea el cerebro,
y una cadena de vértebras que protege la médula espinal. Deba-
jo de los huesos se encuentran tres capas de tejidos conectivos
llamados meninges (véase la figura 38-12a, más adelante en este
capítulo). Entre las capas de las meninges, el líquido cerebroes-
pinal, un líquido claro similar al plasma sanguíneo, protege al
cerebro y la médula espinal y nutre a las células del SNC.
El cerebro también está protegido de químicos dañinos
potenciales en el torrente sanguíneo porque las paredes de los
capilares cerebrales son mucho menos permeables que los capi-
lares en el resto del cuerpo. Esta barrera de sangre cerebral
transporta de manera selectiva los materiales necesarios hacia el
cerebro, al tiempo que deja fuera muchas sustancias peligrosas.
En general, la barrera de sangre cerebral evita que las sustancias
solubles en agua difundan de la sangre hacia el cerebro, pero mu-
chas sustancias solubles en lípidos pueden difundirse a través de
las paredes de los capilares.
La médula espinal controla muchos reflejos y
conduce la información hacia y desde el cerebro
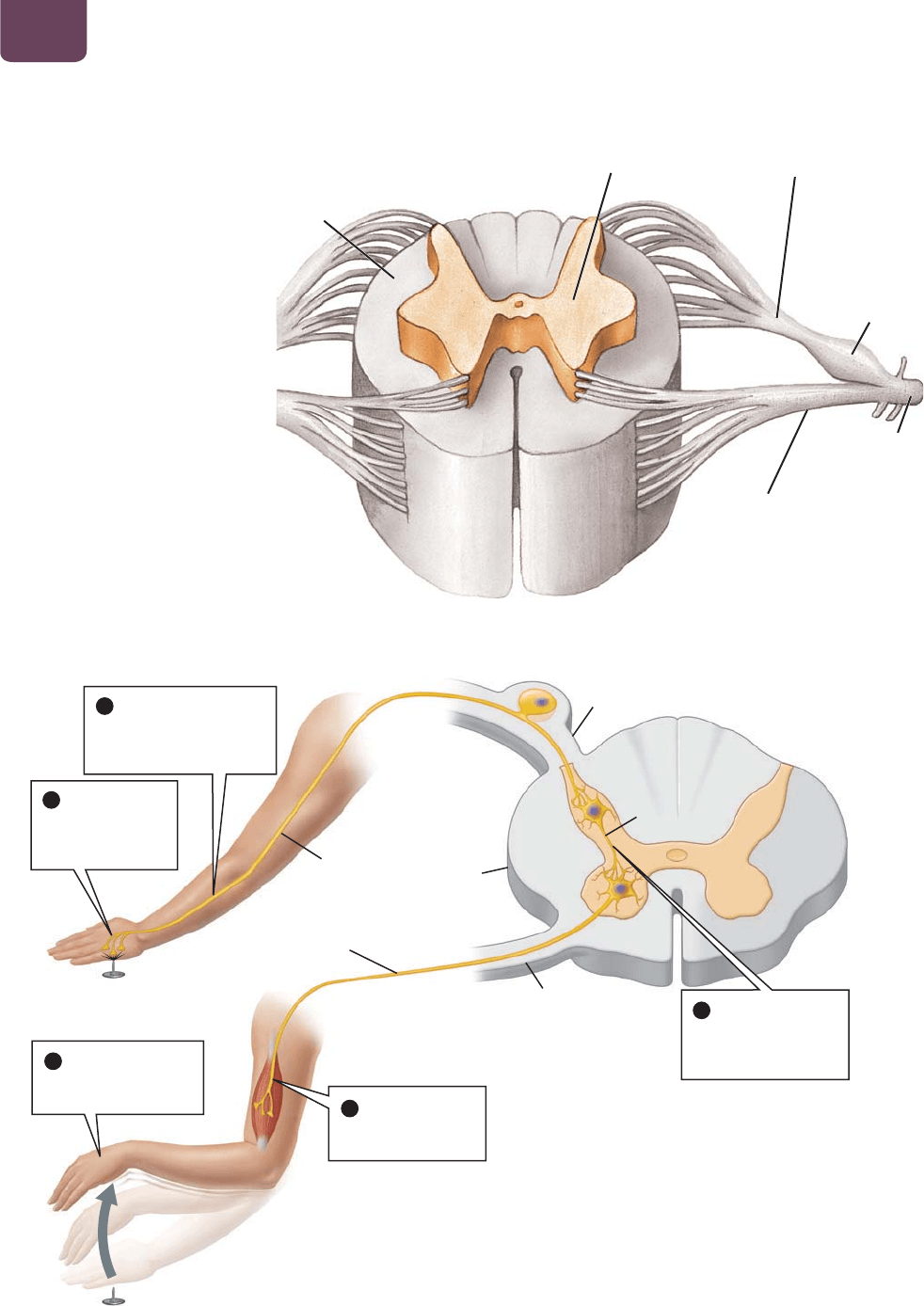
La médula espinal, que tiene más o menos el grosor de tu dedo
meñique (
FIGURA 38-9) se extiende desde la base del cerebro has-
ta la parte baja de la espalda. Los nervios que transportan axones
de neuronas sensoriales surgen de la parte dorsal (posterior) de la
médula espinal, y aquellos que transportan axones de neuronas
motrices surgen de la parte ventral (anterior). Estos nervios salen
para formar los nervios espinales que inervan la mayor parte del
cuerpo. Debido a su parecido con las raíces de un árbol que sur-
gen en un solo tronco, estas ramificaciones se conocen como raíces
dorsales y ventrales de los nervios espinales, respectivamente. Los
ensanchamientos de cada raíz dorsal, conocidos como ganglios
de raíces dorsales, contienen los cuerpos celulares de las neuro-
nas sensoriales.
En el centro de la médula espinal se encuentra un área de
materia gris en forma de mariposa (véase la figura 38-9). (En
realidad, “materia gris” es un término poco apropiado, porque
el tejido nervioso es predominantemente de color café rosado y
se vuelve gris al preservarlo.) En la médula espinal, la materia
gris está conformada por los cuerpos celulares de las neuronas
motrices que controlan los músculos voluntarios y el sistema ner-
vioso autónomo, y por las interneuronas que se comunican con
el cerebro y otras partes de la médula espinal. La materia gris está
rodeada de materia blanca, que contiene axones recubiertos con
mielina de neuronas que se extienden hacia arriba o hacia abajo
de la médula espinal (los recubrimientos de mielina constituida de
lípidos dan un color blanco a estos axones). Los axones transpor-
tan señales sensoriales desde los órganos internos, los músculos y
la piel hasta el cerebro. Asimismo, los axones se extienden hacia
abajo desde el cerebro, transportando señales que dirigen las por-
ciones motrices del sistema nervioso periférico.
Si la médula espinal se corta, la información sensorial
transmitida desde la parte inferior al corte no puede llegar al
cerebro, y el resultado motriz del cerebro no puede alcanzar las
neuronas motrices localizadas debajo del corte. Por tanto, las par-
tes del cuerpo inervadas por las neuronas motrices y sensoriales
localizadas debajo de la lesión se paralizan y se sienten adorme-
cidas, aun cuando las neuronas motrices y sensoriales, los nervios
espinales y los músculos permanezcan intactos.
El sistema nervioso Capítulo 38 745
dilata las pupilas
ojo
inhibe la
salivación
y las
lágrimas
relaja las vías
respiratorias
aumenta el
ritmo cardiaco
estimula la
producción de
glucosa y la libera
inhibe la
digestión
corazón
pulmones
contrae las vías
respiratorias
disminuye el
ritmo cardiaco
hígado
páncreas
estómago
bazo
intestino
delgado
intestino
grueso
vejiga
relaja la
vejiga
estimula el
orgasmo
útero
genitales
externos
recto
riñón
riñón
estimula la
secreción
de epinefrina y
norepinefrina
de la médula
suprarrenal
estimula el
páncreas para
que libere
insulina y
enzimas
digestivas
dilata los vasos
sanguíneos en
los intestinos
estimula la vejiga para
que se contraiga
estimula la
excitación sexual
glándulas salivales
y lagrimales
estimula la
salivación y
las lágrimas
estimula
la digestión
ganglios
simpáticos
craneales
cervicales
torácicos
lumbares
sacros
craneales
cervicales
torácicos
lumbares
sacros
DIVISIÓN PARASIMPÁTICA DEL
SISTEMA NERVIOSO AUTÓNOMO
DIVISIÓN SIMPÁTICA DEL SISTEMA
NERVIOSO AUTÓNOMO
contrae las pupilas
FIGURA 38-8 El sistema nervioso autónomo El sistema nervioso autónomo se divide en dos:
simpático y parasimpático, que abastecen de nervios a muchos de los mismos órganos pero, en general,
producen efectos opuestos. La activación del sistema nervioso autónomo es controlada de forma
involuntaria por las señales del hipotálamo.
746 UNIDAD 5 Anatomía y fisiología animal
materia blanca
contiene
axones
mielinizados
nervio
espinal
raíz dorsal
contiene los
axones de
las neuronas
sensoriales
ganglio de la
raíz dorsal
contiene los
cuerpos celulares
de las neuronas
sensoriales
raíz ventral
contiene los axones
de las neuronas motrices
materia gris
contiene los cuerpos
celulares de las
neuronas motrices
y las interneuronas
FIGURA 38-9 La médula espinal En
un corte transversal, la médula espinal
tiene una región externa de axones
mielinizados (materia blanca) que
viajan hacia y desde el cerebro, y una
región interna en forma de mariposa de
dendritas y los cuerpos celulares de las
interneuronas y las neuronas motrices
(materia gris). Los cuerpos celulares de las
neuronas sensoriales se encuentran fuera
de la médula, en el ganglio de raíz dorsal.
estímulo
neurona
sensorial
médula
espinal
neurona
motriz
raíz dorsal
interneurona
raíz ventral
La neurona
motriz estimula
al músculo efector
El músculo efector
provoca una
respuesta de retiro
Un estímulo
doloroso activa
una neurona
sensorial del dolor
La señal se
transmite a través de la
neurona sensorial del
dolor a la médula espinal
La señal se
transmite a una
interneurona y luego
a una neurona motriz
4
3
2
1
5
FIGURA 38-10 El reflejo de retiro ante el dolor
PREGUNTA ¿Por qué una víctima paralizada debido a una lesión en la médula espinal, al darle un pinchazo en una parte
paralizada del cuerpo, a menudo presenta un reflejo de retiro ante el dolor normal pero no siente ningún dolor?
El sistema nervioso Capítulo 38 747
Los circuitos neuronales para muchos reflejos
residen en la médula espinal
El tipo de conducta más sencillo es el reflejo, un movimiento en
gran medida involuntario de una parte del cuerpo en respuesta a
un estímulo. En los vertebrados, muchos reflejos son producidos
por la médula espinal y las neuronas periféricas, sin que interven-
ga el cerebro.
Enseguida se examina el reflejo de retiro ante el dolor, en
el que están involucradas neuronas tanto del sistema nervioso
central como del periférico (
FIGURA 38-10). Si recargas la mano
sobre una tachuela, el daño al tejido activa las neuronas senso-
riales del dolor (
FIGURA 38-10 ❶). Los potenciales de acción en
los axones de estas neuronas sensoriales del dolor viajan por el
nervio espinal y entran en la médula espinal a través de una raíz
dorsal (
FIGURA 38-10 ❷). Dentro de la materia gris de la médula,
la neurona sensorial del dolor estimula a una interneurona, que
a su vez estimula a una neurona motriz (
FIGURA 38-10 ❸); las
interneuronas se encuentran literalmente “entre otras neuronas”,
en este caso, entre una neurona sensorial y una motriz. Los po-
tenciales de acción en el axón de la neurona motriz salen de la
médula espinal a través de un músculo esquelético. El potencial
de acción estimula el músculo (
FIGURA 38-10 ❹), que se contrae,
alejando tu mano de la tachuela (
FIGURA 38-10 ❺).
Muchas interneuronas de la médula espinal tienen tam-
bién axones que se extienden hasta el cerebro. Los potenciales de
acción en estos axones informan al cerebro sobre la mano lasti-
mada y pueden disparar conductas más complejas, como gritos
de dolor y un aprendizaje sobre los peligros de las tachuelas. El
cerebro a su vez envía potenciales de acción a través de los axones
en la materia blanca de la médula espinal a las interneuronas y
las neuronas motrices en la materia gris. Estas señales del cerebro
pueden modificar los reflejos espinales. Con el aprendizaje o la
motivación suficientes, puedes suprimir el reflejo de retiro ante el
dolor. Para rescatar a un niño de una cuna en llamas, por ejem-
plo, podrías pasar a través del fuego.
Algunas acciones complejas son coordinadas
en la médula espinal
El cableado para algunas actividades muy complejas también re-
side dentro de la médula espinal. Todas las neuronas y las inter-
conexiones necesarias para los movimientos básicos al caminar
y correr, por ejemplo, se encuentran en la médula espinal. La ven-
taja de esta organización parcialmente independiente entre el cere-
bro y la médula espinal tal vez sea un incremento en la velocidad
y la coordinación, porque los mensajes no tienen que viajar hasta
el cerebro y de regreso sólo para mover la pierna hacia delante al
caminar. La función del cerebro en estas conductas semiautomá-
ticas consiste en iniciar, guiar y modificar la actividad de las neuro-
nas motrices espinales con base en decisiones conscientes (hacia
dónde vas; qué tan rápido debes caminar). Para mantener el equi-
librio, el cerebro utiliza también la información sensorial de los
músculos para ordenar a las neuronas motrices que ajusten la for-
ma en que se mueven los músculos.
El encéfalo se conforma de varias estructuras
que realizan funciones específicas
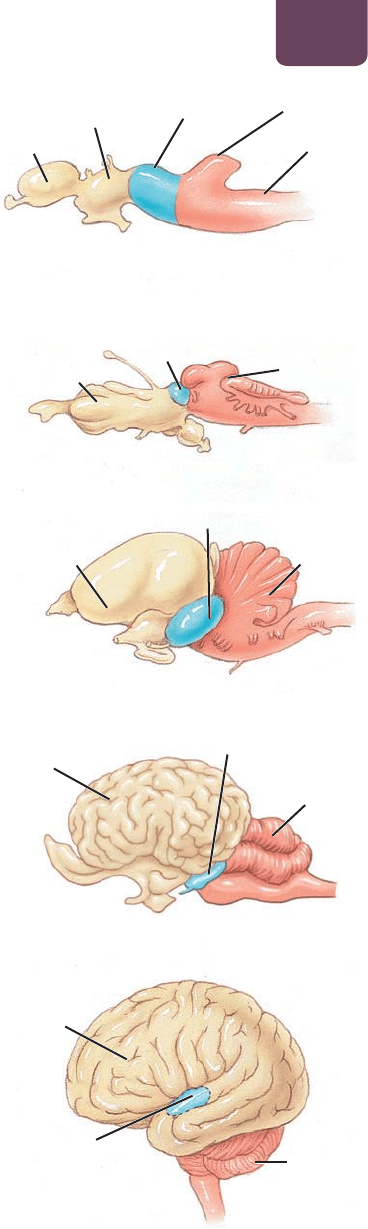
El cerebro de todos los vertebrados tiene tres partes principales: el
romboencéfalo, el mesencéfalo y el prosencéfalo (
FIGURA 38-
11a
). Los científicos creen que en los primeros vertebrados estas tres
FIGURA 38-11 Comparación de los cerebros de varios
vertebrados (a) El cerebro de los embriones vertebrados
modernos, el cual se pensaba que era similar al de los ancestros
distantes de los vertebrados actuales, está conformado de
tres regiones diferentes: el prosencéfalo, el mesencéfalo y el
romboencéfalo. (b) El cerebro de un tiburón adulto conserva su
organización básica. (c) En el ganso, el mesencéfalo se reduce y
el cerebro y el cerebelo son más grandes. (d, e) En los mamíferos,
en especial en los seres humanos, el cerebro es muy grande en
comparación con otras regiones cerebrales.
(a) Cerebro de un embrión vertebrado
cerebro
tálamo
mesencéfalo
mesencéfalo
lóbulo óptico
cerebelo
médula
prosencéfalo mesencéfalo romboencéfalo
(b) Cerebro de un tiburón
(c) Cerebro de un ganso
(e) Cerebro humano
cerebro
cerebelo
cerebro
cerebro
mesencéfalo
(interior)
cerebelo
cerebelo
(d) Cerebro de un caballo
cerebro
mesencéfalo
cerebelo
Este documento contiene más páginas...
Descargar Completo
Transmision sináptica215.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.