Sistema de endomembranas
El sistema de endomembranas se distribuye por todo el citoplasma y está
formado por varios subcompartimientos (cisternas, sacos, túbulos) y organoides
(RER, REL, complejo de Golgi, endosomas y lisosomas).
Las membranas de los organoides y subcompartimientos se comunican
directamente o por vesículas transportadoras. Las vesículas transportadoras
brotan en la menbrana de un compartimiento llamado donante y viajan por el
citosol hasta otro compartimiento llamado receptor donde se fusiona con su
membrana.
Las membranas de los organoides y de las vesículas están constituidas por una
bicapa lipídica similar a de la membrana plasmática. Una de las capas de la
membrana se relaciona con el citosol (cara citosólica) y la otra se relaciona con
la cavidad interna de los organoides denominada cara luminal. Las membranas
poseen glicolípidos y glicoproteínas intrínsecas y periféricas, los hidratos de
carbono se orientan siempre hacia la cavidad de los organoides
Retículo endoplasmatico
Se distribuye por todo el citoesqueleto desde el núcleo hasta la membrana
plasmática, está compuesto de una red tridimensional de túbulos y sacos
aplanados totalmente interconectados. Se divide en dos sectores que se
diferencian por la presencia o no de ribosomas sobre su cara citosólica.
Retículo endoplasmatico liso (REL)
Está formado por túbulos anastomosados ramificados que son continuación del
retículo endoplasmatico rugoso con el cual comparten el lumen. Es responsable
por:
• Síntesis de lípidos: todos los lípidos son sintetizados en el REL,
triglicéridos, fosfolípidos, glucolípidos, hormonas esteroideas, etc.
o Síntesis de fosfolípidos: ocurre en la cara citosólica del REL, luego
mediante una flipasa (enzima) estos fosfolípidos originados en la
monocapa citosólica de la bicapa se transfieren a la otra cara de la
bicapa para un crecimiento simétrico de la bicapa lipídica. También
se reciclan membranas de la célula.
o Síntesis de hormonas esteroideas: para producir hormonas
esteroideas el colesterol primero pasa por las mitocondrias donde
es transformado en Pregnenolona, y esta molécula pasa al REL
para ser transformada en la hormona esteroidea específica de
cada célula.
• Detoxificación: es la inactividad y el aumento de la hidrosolubilidad de
sustancias toxicas endógenas (producidas pelo cuerpo) y exógenas (que
ingerimos). Posee enzimas detoxificantes como la familia enzimática de
la Citocromo P450.
• Almacenamiento de calcio: el calcio ingresa desde el citosol a través de
combas de calcio en la menbrana del REL.

• Metabolismo de la glucosa: en el REL de los hepatocitos hay una enzima
llamada glucosa 6 – fosfatasa que participa en la vía metabólica
transforma glucógeno en glucosa que vuelve a la sangre.
Retículo endoplasmatico rugoso (RER)
Es una continuación de la carioteca y se continua como REL, está recubierto de
ribosomas y por lo tanto su principal función es la síntesis de proteínas no
citosólica (de la membrana plasmática, las extracelulares, las de exportación y
las del sistema de endomembranas).
El péptido señal
Todas las proteínas comienzan a sintetizarse en ribosomas libres, sin embargo,
dependiendo del destino pueden ir o no al RER. El péptido señal son los primeros
10 a 14 aminoácidos de la proteína que está sintetizando que es altamente
hidrofóbico y de secuencia precisa y conocida que permiten saber cuál va ser el
destino de cada proteína.
En el RER el péptido señal es reconocido por la partícula de reconocimiento de
señal (PRS) que es una ribonucleoproteína formada por ARNpc y 6 proteínas.
La PRS tiene las siguientes funciones secuenciales:
• Reconoce al péptido señal cuando se asoma por el ribosoma y se une a
él.
• Detiene la síntesis proteica.
• Arrastra al polirribosoma hacia la membrana del RER.
• Se une al receptor para PRS que se encuentra en la membrana del RER.
El receptor para la PRS se encuentra muy cerca de del canal proteico Translocón
que permite que la proteína ingrese al lumen del RER. Una vez dentro del lumen
del RER pasan dos cosos claves:
• Se corta el péptido señal por medio de la enzima péptidas señal presente
en la cara no citosólica de la membrana del RER.
• Se hace n-glicolización o glicosilación inicial de la proteína por medio de
un conjunto de enzimas llamadas Glucosil Transferasa.
La Glucosil Transferasa obtiene glúcidos de un lípido especial llamado Dolicol
Fosfato presente en la membrana del RER, este lípido tiene un mismo
oligosacárido de 12 azucares el cual lo transfieren a la proteína.
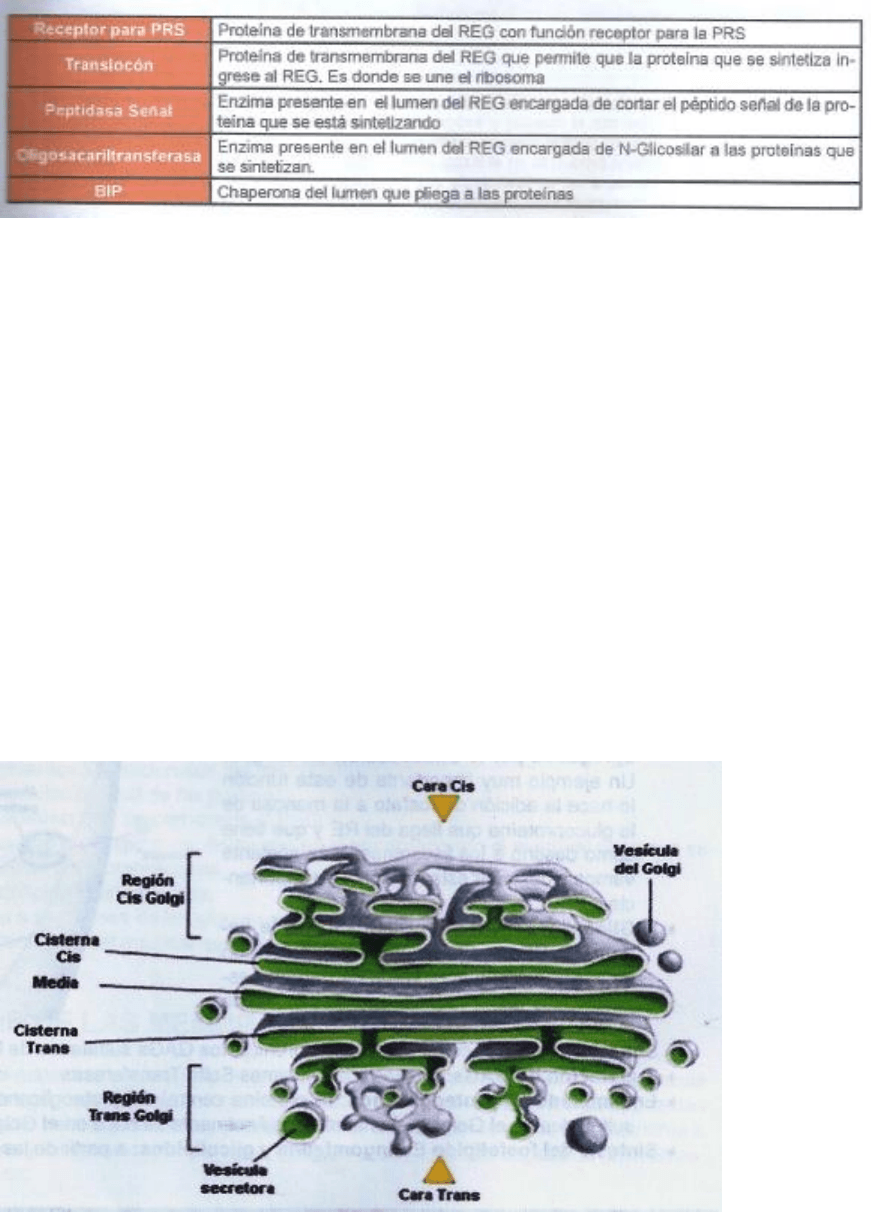
Complejo de Golgi
El aparato de Golgi se ubica entre el retículo endoplasmatico y la membrana
plasmática de la célula. Se trata de sáculos aplanados anastomosados y
polarizados o sea poseen una cara de entrada y una de salida.
El complejo de Golgi está integrado por una o varias unidades funcionales
llamadas dictiosomas que poseen una forma curvada con la cara convexa
mirando hacia el núcleo y la convexa hacia la membrana plasmática. Cada
dictiosoma está integrado por:
• Red cis: cara de entrada formada por numerosos sacos y túbulos
interconectados.
• Cisterna cis: conectada a la red cis
• Cisterna media: puede ser una o más, no están conectadas entre sí o con
lo restante del dictiosoma.
• Cisterna trans: conectada a la red trans
• Red trans: cara de salida, similar a la red cis.
Las funciones del aparato de Golgi son:
• Selección del destino de las proteínas que llegan del RE
• Modificación de los hidratos de carbono agregados por N-Glicosilación en
el REG
• O-Glicosilación de las proteínas que llegan del RE
• Síntesis de GAGs
• Sulfatación de GAGs
• Ensamblado de proteoglicanos
• Síntesis del fosfolípido esfingomielína y glicolípidos
La envoltura nuclear
La envoltura nuclear está formada por dos membranas una membrana nuclear
externa y una interna que delimitan un espacio perinuclear, estas membranas se
continúan con las membranas del retículo endoplasmatico mientras que el
espacio perinuclear se continua con el lumen del RE.
Esta interrumpida por poros nucleares que es por donde entran y salen todas las
moléculas. Cada poro está formado por muchas unidades proteicas diferentes
entre ellas 8 columnas proteicas y fibrillas que asociadas forman una estructura
similar a una jaula. Posee también proteínas radiales que llenan el centro del
canal y permiten el transporte selectivo a través de la envoltura nuclear. Las
moléculas pequeñas e hidrosolubles pasan libremente mientras que las
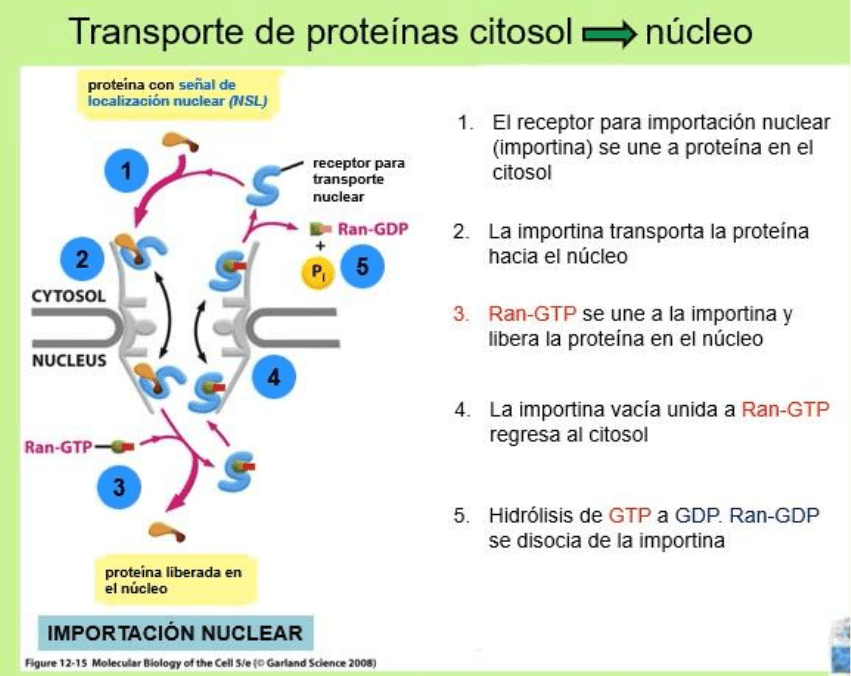
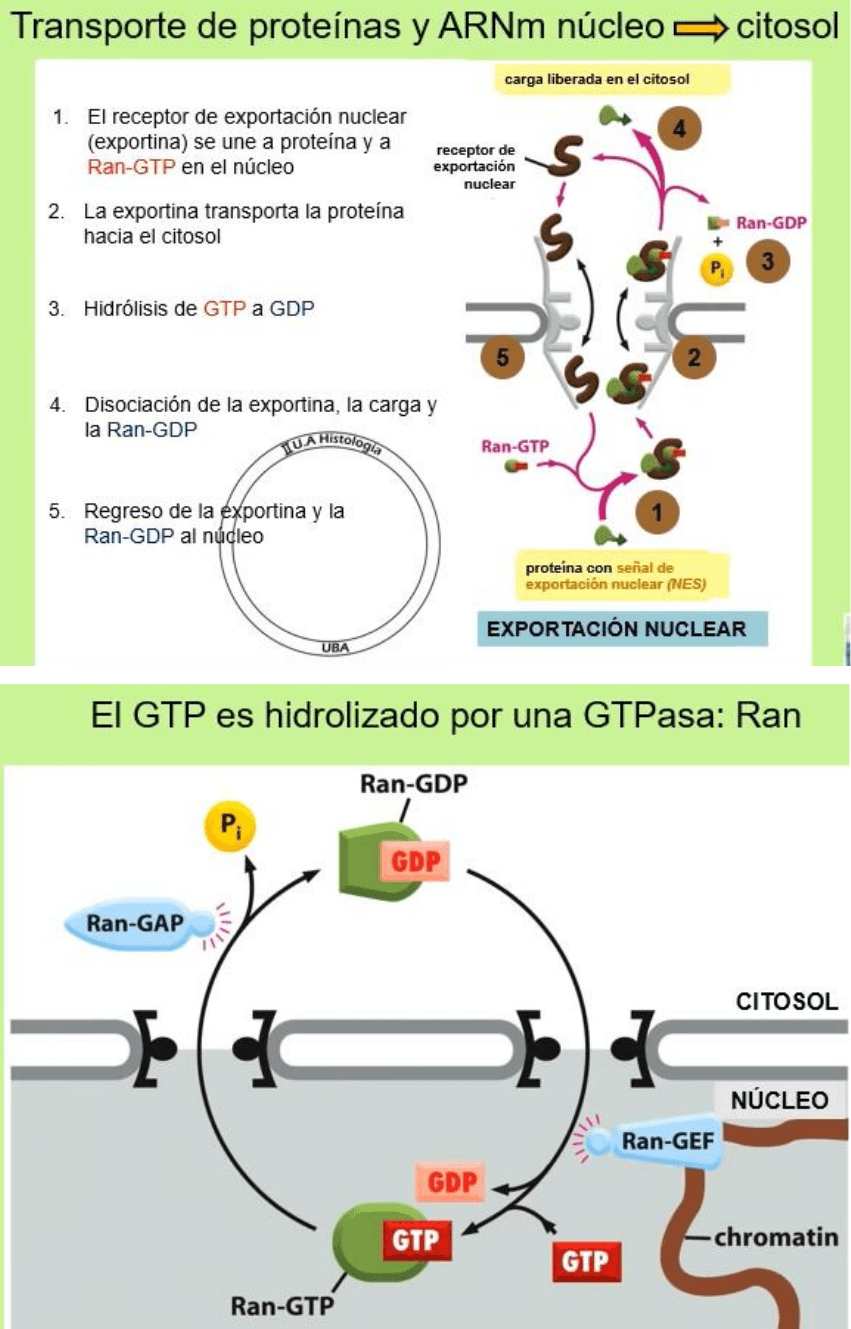
macromoléculas atraviesan por transporte activo. El transporte de proteínas se
lleva de la siguiente forma
Transporte vesicular
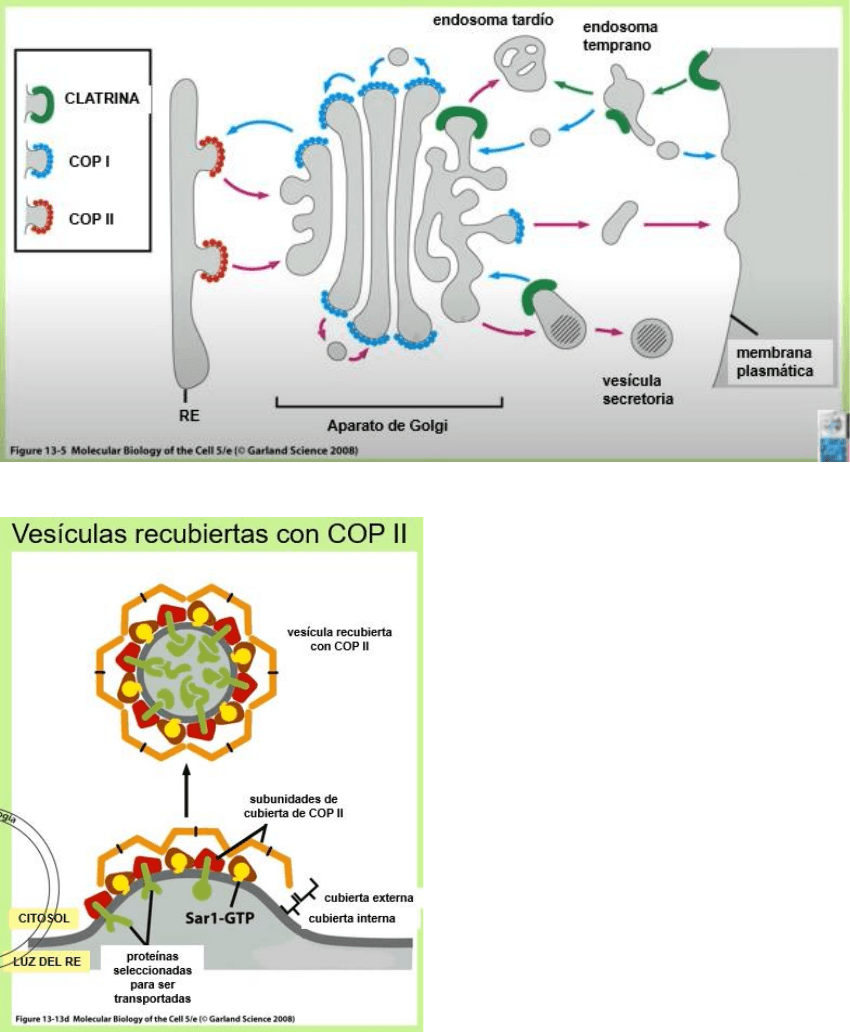
Toda vez que dos organelas separadas por citosol deban comunicarse lo harán
por vesículas, que forman un sistema de transporte muy preciso, es decir, la
vesícula que brota de un compartimiento debe tener las proteínas específicas
para unirse solamente a su destino adecuado. Hoy se conocen 3 tipos de
cobertura proteica:
• Clatrina: corresponde a las vesículas del endocitosis. Las del Golgi a los
lisosomas, Golgi a endosomas, endosomas de reciclaje a membrana y las
de exocitosis.
• COP 1: las del Golgi a RE, las del Golgi entre sí, las del endosoma
temprano al tardío.
• COP 2: las del RE al Golgi, y del Golgi entre sí pero en sentido
anterógrado
Vesículas recubiertas de coatómero
COP es una sigla que significa cubierta
de coatómero, que es un complejo
proteico de 700 KDa formado por 7
subunidades que forman una rede 3D.
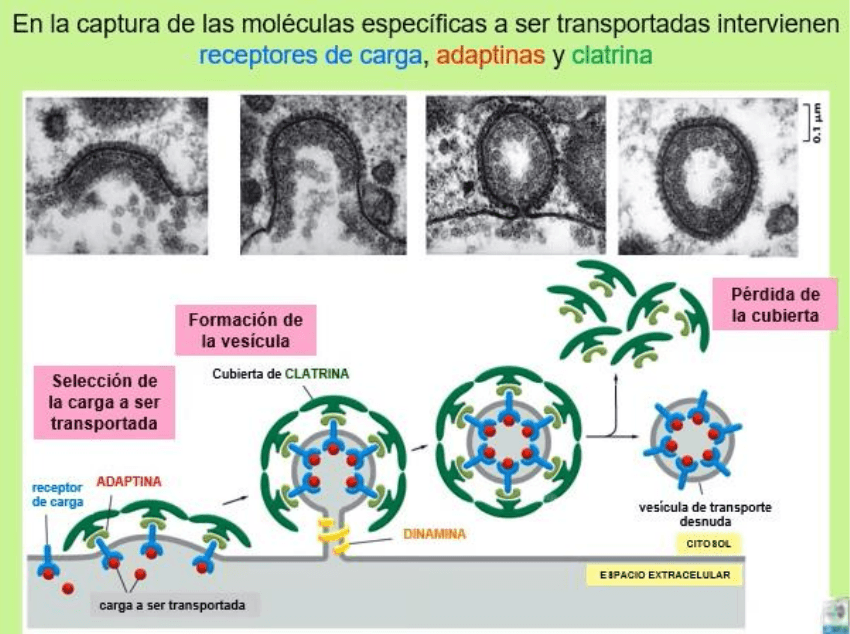
Vesículas recubiertas de Clatrina
Clatrina es un complejo proteico compuesto por 3 grandes cadenas
polipeptídica y tres cadenas polipeptídica pequeñas que conforman una
estructura llamada TRISKELION.
En las vesículas cubierta de Clatrina encontramos las siguientes proteínas:
• Clatrina
• Adaptina: se une de un lado con la Clatrina y con el otro con el receptor
de membrana para la molécula específica a endocitar.
• Receptor: se une específicamente a la molécula que se quiere endocitar.
• Dinamina: proteína motora microtubular con función del cierre del cuello
de las vesículas y la siguiente separación de la membrana plasmática.
Fusión de las vesículas con la membrana destino
El acople de la vesícula con la membrana es un mecanismo regulado por una
familia de proteínas llamadas SNARE:
• T-SNARE: presente en la membrana de la organela receptora
• V-SNARE: presente en la membrana de la vesícula
Para poder fusionar ambas SNARE deben interactuar y reconocerse. Este
reconocimiento altamente selectivo es regulado por una familia de proteínas
monomerica fijadoras de GTP llamadas RAB que chequean que el
reconocimiento y la unión de las V-SNARE y las T-SNARE sea correcta en
vesículas cubiertas de Clatrina.
Endosomas
Los endosomas son organelas que participan de la vía endocítica, están
revestidos por membranas biológicas, presentan un pH ácido y poseen bomba
de protones en su membrana para mantener su pH.
Existen 4 tipos de endosomas:
• Endosomas tempranos: cercanos de la membrana plasmática, pH 6. No
poseen enzimas hidrolíticas, recibe la vesícula endocítica (vesícula que
viene del endocitosis mediada por receptor).
• Cuerpos multivesiculados: intermedio entre el endosoma temprano y el
tardío. Degrada proteínas de membrana y cuando se fusiona con otros
cuerpos multivesiculares forman el endosoma tardío.
• Endosoma tardío: cerca del aparato de Golgi, con pH 5.6. Tiene enzimas
hidrolíticas porque recibe vesículas del Golgi con enzimas. Se forma de
la fusión de varios cuerpos multivesiculares. Cuando se fusiona a
lisosoma primario da lugar al Heterolisosoma o vesícula digestiva.
• Endosoma de reciclaje: participan en el reciclado de proteínas de
membrana y de proteínas de Golgi. Las proteínas de membrana que se
reciclan son receptores de las Acuoporinas II.
Los EXOSOMAS son cuerpos multivesiculares que captan proteína, lípidos,
ADN, ARNm y ARN no codificante especifico y se fusionan a la membrana
plasmática y sus vesículas son liberadas al exterior de la célula. Las vesículas
suelen en aparecer en fluidos corporales como sangre, saliva, orina, LCR.
Lisosomas

Sistema de endomembranas.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.