Sintesis conceptuales 1
Unidad 1 y 2 Mecanismos moleculares que participan en el mantenimiento y la variabilidad del ADN nuclear
⚫ Mantenimiento: Replicación del ADN: características y fases. Sistemas/Mecanismos de reparación del ADN.
⚫ Variabilidad: Mutaciones. Mutaciones en células somáticas y en células germinales. Reproducción sexual. Reorganización del ADN:
Recombinación homóloga, recombinación sitio específica, transposición (vía intermediarios de ADN/ARN), amplificación génica.
⚫ Replicación de ADN in vitro: PCR (Reacción en cadena de la polimerasa) y RT-qPCR (PCR cuantitativa con retrotranscripción).
Replicación del ADN - Mecanismos de reparación del ADN
Caracteristicas del ADN
⚫ El ADN es un polinucleótido, es decir, un polímero de nucleótidos enlazados covalentemente por puentes fosfodiéster.
⚫ El ADN bicatenario está constituido por dos cadenas (o hebras) complementarias que están estabilizadas por interacciones
intermoleculares conocidadas como puentes de hidrógeno, los cuales se estabelecen entre bases nitrogenadas complementarias
(A-T (2 puentes de hidrógeno) y C-G (3 puentes de hidrógeno)).
⚫ El ADN prensenta dos cadenas antiparalelas. Una de las cadenas discurre en el sentido 5’ → 3’ y la otra 3’ → 5’.
⚫ Componentes de nucleotidos: fosfato + desoxirribosa + base nitrogenada.
Replicación del ADN
⚫ Es el proceso protagónico de la Fase S, en cual también va a replicar a las histonas (proteínas básicas asociadas al ADN).
⚫ Es un proceso semiconservativo, es decir, que cada molécula hija de ADN hereda una hebra complementaria de la molécula
original.
⚫ Es un proceso bireccional debido a que el ADN presenta cadenas antiparalelas, y también, a que las enzimas catalizan la
replicación siempre en sentido 5’ → 3’.
⚫ Es un proceso asimétrico a raíz de que ambas cadenas se sintetizan a diferente velocidad, un de forma continua y la otra en forma
de fragmentos que posteriormente se unen.
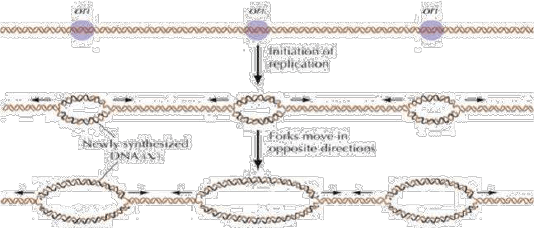
➢ En las células eucariotas, debido a su extenso genoma, la replicación
del ADN tiene inicio cuando una serie de proteínas que componen
un complejo denominado ORC (complejo de reconocimiento de
origen) se unen a varias secuencias, dispersas en el ADN, conocidas
como orígenes de replicación (orí). Los orígenes de replicación contienen
secuencias nucleotídicas denominadas ARS (autonomously replicating
sequence) de alrededor de 100 pares de bases. Existe una
secuencia central de alrededor de 11 pares, común a los ARS, donde
se une el complejo ORC. Las células procariotas, debido un genoma menor, presentan solo un origen de replicación.
Requiere la actividad de una serie de proteínas y enzimas sucesivamente en los pasos de: iniciación, elongación y terminación.
⚫ Iniciación: proceso en el cual ocurre la interacción entre los elementos de iniciación y las secuencias que participan en el
reclutamiento de elementos iniciadores.
⚫ Elongación: la elongación es el proceso donde se extiende en longitud las cadenas que están siendo sintetizadas en la horquilla de
replicación.
⚫ Terminación: es la fase menos comprendida del modelo y se da cunado la molécula ya ha sido completamente replicada.
Las enzimas que se encargan de la polimerización de desoxirribunucleótidos trisfosfato son las ADN polimerasas. Las características
generales de las ADN polimerasas son:
⚫ Requiere de un primer o cebador (es una secuencia corta de ARN que se incorpora en el inicio del proceso de replicación del ADN,
sirve como raíz para que el ADN polimerasa pueda extender la nueva cadena).
⚫ Catalizan las puentes fosfidiéster de forma sucesiva y ordenada en los extremos 3’ de los nucleótidos.
⚫ La polimerización de los desoxirribunucleótidos trifosfato ocurre siempre en dirección 5’ → 3’.
⚫ Requiere la participación de proteínas accesorias para fijarse y mantenerse unida al ADN debido a su alto K
M
(grado de afinidad de
la enzima por su sustrato)
⚫ Participa también en la reparación (lectura de prueba) debido a su actividad 3’ exonucleasa (rompe el ácido nucleico desde sus
extremos).
Pasos de la replicación:
1 - Formación del complejo de pre-replicación: en Fase G1 se asocia el
complejo ORC a las orí; ese se activa recluta MCM helicasa y otras
enzimas, la unión de esos elementos forma el complejo de
pre-replicación.
2 - Formación de la horquilla de replicación: el complejo de pre-replicación
se continua con la separación de las hebras complementarias formando
(abriendo) la horquilla de replicación, acá empieza la iniciación.
3 - Iniciación: síntesis del ARN primer o ARN cebador, hecha por la ADN primasa que se asocia a la horquilla y al ADN y sintetiza una
pequeña cadena de ADN (primer o cebador), posibilitando el inicio de la elongación.
4 - Elongación: síntesis de las nuevas cadenas de ADN. Como las cadenas del ADN son antiparalelas y la ADN polimerasa solo ocurre
en sentido 5’ → 3’ la polimerización de las nuevas cadenas es bidireccional y asimétrica. La cadena adelantada o conductora con
sentido 3’ → 5’ requiere solo un primer, por eso es constante y acelerada. Ya la cadena retrasada con sentido 5’ → 3’ requiere la
presencia de varios fragmentos (Fragmentos de Okazaki) una vez que la ADN polimerasa empiece a elongar cada primer, por eso es
descontinua y lenta.
5 - Eliminación de ARN primer: Una vez sintetizada toda la cadena continua y los fragmentos de Okazaki (a partir de cada orí), los
primers de la cadena retrasa van a ser eliminados por una enzima RNAsa H, la cual va a “digerir” el ARN primer, y va a formar huecos
en la cadena.
6 - Cumplimentación de los huecos: la ADN polimerasa con capacidad de reparación va a rellenar los huecos dejados por los primers,
reemplazándolos por ADN.
7 - Soldadura de fragmentos: la ADN ligasa va a soldar las hendiduras uniendo los nuevos fragmentos de Okazaki, sellando los
huecos.
Existen todavía una serie de enzimas y proteínas accesorias que son extremadamente importantes:
⚫ Helicasas: se encargan de seguir extendiendo la horquilla de replicación para que los fragmentos de ADN neosintetizado puedan
elongar hasta la horquilla de replicación vecina. Su función se debe a su capacidad de romper las interacciones puentes de
hidrógenos entre las bases nitrogenadas complementarias.
⚫ SSB (single-stranded DND binding proteins): son las encargadas de unirse a las hebras de la horquilla para impedir que estas
vuelvan a reasociarse a través del restablecimiento espontáneo de las interacciones puentes de hidrógeno.
⚫ Proteína de enganche de carga (RPC) y abrazadera deslizante (PCNA): son dos proteínas accesorias que auxilian la ADN
polimerasa en su función. Debido a la baja afinidad de la ADN polimerasa por su sustrato, ésta tiende a disociarse de la molécula
de ADN. La proteína de enganche de carga cataliza la unión de la proteína abrazadera deslizante, la cual ayuda a la ADN
polimerasa a mantenerse fija en la hebra de ADN mientras esta está siendo sintetizada.
⚫ Topoisomerasa: se encargan de disminuir la tensión mantenida por la horquilla del ADN. El ADN, al tener sus cadenas separadas
en la formación y separación de la horquilla, gira en los extremos de ésta generando un superenrollamiento de la molécula, lo que
podría detener la replicación por la generación de deformaciones en el ADN. La toisomerasas se encargan de disminuir la torsión
de la molécula generando cortes reversibles en una sola hebra (topoisomerasa I o ADN girasa) o en ambas (topoisomerasa II).
8 - Terminación: es la fase menos comprendida del proceso de replicación y concluimos que ocurre cuando el ADN fue
completamente replicado. Cada origen de replicación nos permite obtener un segmento denominado replicón, al final de la
replicación del ADN, los replicones deben ser ensamblados, constituyendo de esta forma una molécula idéntica a la original en to su
extensión.
ADN polimerasa δ (delta): replicación de la cadena adelantada.
ADN polimerasa α (alfa): elaboración de los fragmentos de Okazaki.
ADN polimerasa β (beta): rellena los huecos dejados por la eliminación de los primers.
Telomerasa
⚫ Como la ADN plimerasa solo extienden los cebadores en de 5’ → 3’ son incapaces de copiar los extremos 5’. Estas secuencias
terminales, los telómeros, se componen de repeticiones en tándem de secuencias simples de ADN. Talles secuencias son
replicadas por la acción de una enzina, la telomerasa, que es capaz de mantener a los telómeros catalizando de su síntesis en
ausencia de una hebra molde de ADN.
⚫ La telomerasa es una transcriptasa inversa, que sintetiza ADN a partir de un molde de ARN. Ella porta su proprio molde de ARN,
que es complementario a las secuencias repetidas de los telómeros.
⚫ El uso de este ARN como molde permite a la telomerasa generar múltiples copias de las secuencias repetidas, manteniendo por lo
tanto a los telómeros en la secuencia de un molde de ADN convencional para dirigir su síntesis.
⚫ La utilización de este ARN como molde permite a la telomerasa extender al extremo 3’ del ADN cromosómico una unidad de
repetición detrás de su longitud original. La hebra complementaria puede ser entonces sintetizada por el complejo polimerasa α-
primasa utilizando como iniciador al ARN convencional.
⚫ La eliminación de los ARN cebadores deja un extremo 3’ del ADN cromosómico suelto, que puede formar lazos al final de los
cromosomas eucariotas.
⚫ La mayoría de las células somáticas no poseen niveles suficientemente elevados de telomerasa para mantener la longitud de sus
telómeros durante las incontables divisiones celulares, consecuentemente los telómeros se acortan a medida que las células
envejecen, y este acortamiento final desencadena la muerte celular o senescencia.
Mecanismos de reparación del ADN
⚫ El ADN posee una serie de mecanismos de reparación para evitar que las células hijas hereden ADN mutado en secuencias de alta
relevancia o que el ADN sea dañado.
⚫ Cuando hay un daño en el ADN la célula lo identifica detiene el ciclo celular y pone en marcha los mecanismos de reparación,
cuando la reparación sucede correctamente se reanuda el ciclo celular y la célula puede proliferarse.
⚫ Cuando la reparación no sucede el adecuado o en general la célula pone en marcha mecanismos pro-apoptóticos, que son
irreversibles.
⚫ La célula puede fallar en el reconocimiento y no activar su mecanismos que evitan la proliferación anormal, lo que precede el
cáncer.
⚫ El ADN puede sufrir daños constantemente que pueden clasificarse en líneas generales en:
⚫ Espontáneos
⚫ Desaminación: perdía de grupos amino de las bases nitrogenadas del ADN
⚫ Depurinación: consiste en la eliminación del enlace N-glucosídico entre la base nitrogenada y el azúcar, con la consiguiente
pérdida de un residuo de adenina o guanina.
⚫ Inducios
⚫ Factores químicos:
⚫ Factores físicos:
⚫ Los mecanismos de reparación del ADN pueden dividirse en dos grupos principales:
⚫ Inversión directa de la alteración o reacción química que desencadenó la mutación
⚫ La “doble lectura” de la ADN polimerasa: la capacidad de la ADN polimerasa de corregir el agregado de bases incorrecatas
durante la replicaión del ADN por su actividad correctora 3’ exonucleasa.
⚫ La fotorreactivación: en este proceso se utiliza la energía de la luz visible para romper el anillo de ciclobutano formado en las
pirimidinas (T o C o U) adyacentes.
⚫ Transferencia de grupos alquilo: en los procesos de alquilación del ADN, donde ciertos agentes alquilantes transfieren a
bases, como la guanina, el grupo metilo o etilo. Algunas enzimas, como las con actividad metiltransferasas, pueden retirar el
metilo o transferirlo a otra molécula restableciendo la base original.
⚫ Eliminación de las bases dañadas seguida de su reposición con ADN recién sintetizado
⚫ Reparación por escisión de bases nitrogenadas: consiste en la elimanación de la base alterada del esqueleto del ADN (azúcar
+ fosfato). Un mecanismo de reparación por escisión de bases es la retirada de residuos dañados que se forman en el ADN.
Lo mejor ejemplo es el uracilo, que puede incorporarse en el ADN de dos formas: (1) El uracilo se incorpora ocasionalmente
en lugar de la timina durante la síntesis del ADN; (2) El uracilo se puede formar en el ADN por la desaminación de una
citosina (ese altera el patrón de pares complementarios). La escisión del uracilo es catalizada por la enzima ADN glicolisada,
la cual rompe la unión del uracilo con la desoxirribosa, está reacción produce un uracilo libre y un sitio apirimidínico o
apurínico (sitio AP)(azúcar sin base). La ADN glucosilasa también reconoce y elimina otras bases anómalas. Luego por acción
de las enzimas AP endonucleasa y desoxirribosafosfodiesterasa se retira el azúcar y el fosfato presente en el sitio AP y se
rellena el espacio por la ADN polimerasa y la ligasa.
⚫ Reparación por escisión de nucleótidos: se requiere cuando es más extenso que una base nitrogenada, como un dímero de
pirimidina inducidos por UV, que afecta al menos dos bases adyacentes. Consiste en la retirada de un oligonucleótido que
contienen una lesión, que tiene como objetivo eliminar un daño, como un dímero de timina. En éstos casos, el ADN dañado
es reconocido y después se abre la doble hebra por acción de la helicasa. A continuación, se escinde un segmento de la
cadena con el daño, por nucleasas 3’ y 5’ que rompen los extremos, produciendo la retirada del segmento dañado y sus
porciones aledañas. Posteriormente, el espacio desocupado es rellenado por la ADN polimerasa y fijado por la ligasa.
⚫ Reparación por eliminación de nucleótidos acoplada a la transcripción: ocurre durante el proceso de transcripción, el
mecanismo implica el reconocimiento de la ARN polimerasa detenida en una lesión de la hebra de ADN que se está
transcribiendo. La ARN polimerasa detenida es reconocida por una proteína denominada facto de acoplamiento de la
reparación a la transcripción, que desplaza a la ARN polimerasa y recluta a la escinucleasa a la zona de la lesión. En las células
de los mamíferos ese reconocimiento a la ARN polimerasa ocurre por las proteínas CSB y CSA. Tras el reconocimiento de la
polimerasa de ARN detenida, las proteínas CSA y CSB reclutan a XPA, RPA y TFIIH hacia la lesión de ADN y se produce una
reparación por escisión de nucleótidos.
⚫ Reparación no complementaria: es un mecanismo que barre de novo el ADN recién replicado, si se encuentra una no
complementariedad, las enzimas de este sistema de reparación son capaces de identificar y escindir la base no
complementaria específicamente de la hebra de ADN recién replicada, permitiendo que se corrija el error y la secuencia
original sea reemplazada.Las proteínas MSH y MLH reconocen la no-complementariedad, se unen a la base no apareada y
dirigen la escisión del ADN, que se encuentra entre la rotura de la hebra y apareamiento erróneo. La especificidad de la
hebra para la reparación no complementaria se puede identificar por la presencia de roturas en ambos extremos de los
fragmentos de Okazaki de la hebra recién replicada, mientras que la hebra conductora se puede identificar por su extremo 3’
en crecimiento.
⚫ Reparación de roturas de doble hebra: la rotura en ambas las hebras del ADN interrumpe la continuidad de dicha molécula,
talles roturas se producen de manera natural en el transcurso de la replicación, o, por otra parte, la radiación ionizante y
algunos compuestos químicos pueden producir roturas de doble hebra. Esa variable no es suscetible de reparación mediante
a los mecanismo anteriores. Los mecanismos de reparación para ese caso son: reparación recombinatoria y recombinación
homóloga. En la reparación recombinatoria las roturas de las hebras pueden repararse simplemente volviendo a unir los
extremos rotos de una sola molécula de ADN, esta forma de reparación introduce errores y hay pérdida de bases e punto de
unión. Mientras en la recombinación homóloga las roturas se reparen mediante una recombinación con secuencias de ADN
de un cromosoma ileso, lo que constituye una mecanismo para reparar esas alteraciones y restaurar la secuencia normal de
ADN. En las eucariotas ese mecanismo solo ocurre después de la replicación del ADN, cuando las cromátidas hermanas recién
replicadas siguen unidas.
Replicación de ADN in vitro: PCR y RT-qPCR
PCR (Reacción en cadena polimerasa)
⚫ Es una técnica de amplificación de ácidos nucleicos in vitro desarrollada por Kary B. Mullis en 1983.
⚫ Es una técnica molecular versátil, rápida, sensible y específica.
⚫ Componentes necesários para la realización: cadena que va a ser amplificada, primers, ADN polimerasa termoestable, sustratos
de ADN polimerasa (desoxidorribonucleotidos (A;G;T;C)) y cofactores.
⚫ Los pasos de la PCR son: desnaturalización, Hibridación y Elongación.
⚫ Desnaturalización: se eleva la temperatura a entre 93° y 95°C, bajo la cual los puentes de hidrógeno entre bases
complementarias se rompen.
⚫ Hibridación: se reduce la temperatura a entre 50° y 70°C permitiendo la formación de puentes de hidrógeno con la
consecuente asociación (annealing) de los cebadores a las cadenas templado.
⚫ Elongación: se eleva la temperatura nuevamente, pero ahora a alrededor de 70° y 75°, temperatura óptima para la ADN
polimerasa termoestable, facilitando la biosíntesis de ADN.
⚫ Las variaciones de temperatura son generadas por un aparato conocido como termociclador.
⚫ La enzima polimerasa (Taq polimerasa) se obtuvo inicialmente de bacterias termófilas del género Thermus aquaticus. Se trata de
una ezima polimerasa termoestable. La Taq polimerasa no tiene actividad 3’ exonucleasa.
⚫ La amplificación es exponencial, luego de 25 ciclos se pueden obtener 10
5
moléculas amplificadas.
⚫ Los productos se visualizan por medio de la electroforesis (en agarosa) de los amplicones (fragmentos amplificados), separándolos,
como consecuencia, según su peso molecular (cuanto menor su peso molecular más lejos de su origen).
⚫ Luego de la electroforesis, se colorea el ADN con bromuro de etidio, se realizan fotografías del gel mientras este es iluminado por
luz UV y un procesador de imágenes se encarga del análisis de las bandas observadas.
⚫ El proceso es rápido, el clonado por PCR puede realizarse en unas pocas horas, con equipos relativamente no sofisticados.
⚫ Es un proceso sencillo, la amplificación puede llevarse a cabo partiendo de cantidades muy pequeñas de ADN, incluso de una
única célula.
⚫ Una de sus limitaciones es que se necesita información previa de la secuencia que se quiere amplificar, para poder sintetizar los
primers. La otra es que la secuencia a amplificar no puede ser muy larga.
RT-qPCR (PCR cuantitativa por retrotranscripción)
⚫ Es una variación de la PCR, conocida como PCR cuantitativa (tiempo real) precedida por retrotranscripción. Se trata de una técnica
molecular que sigue los mismos principios que la PCR, pero que difiere de esta en dos puntos principales:
⚫ Puede amplificar muestras de ARN;
⚫ La observación de los resultados se hace a través de gráficos en una pantalla.
⚫ Los pasos son los mismos que la técnica de PCR, pero antes de los 3 pasos cíclicos, se requiere de un paso único adicional, una vez
aislada la muestra, que es la Retrotranscripción.
⚫ Retrotranscripción: es la síntesis de una cadena de ADN a partir de una cadena molde de ARN. Un primer especial se une a la
cadena molde de ARN, lo que posibilita la enzima transcriptase reversa hacer la transcripción inversa para sintetizar la cadena
de ADN. La cadena de ADN permanece asociada a la cadena molde y empiezan los mismos pasos del proceso PCR.
⚫ La análisis de los productos para allá de los componentes utilizados en la PCR, requiere un componente especial que emita
florescencia, como un fluorocromo.
⚫ El fluorocromo va a asociarse con los primers. Esa sustancia está inicialmente inactiva, pero luego del proceso de replicación el
ciclador posee un tiene un componente que va a emitir una luz UV la cual va a excitar la sustancia haciéndola emitir luz.
⚫ Los termocicladores, en este caso, tienen un aparato de detección de fluorescencia emitida por la reacción química ocurrida en el
recipiente. Según el grado de fluorescencia se produce patrones que se grafican y aparecen en pantalla.
⚫ Custa más de 20 ciclos para lograr analizar se obtuvo o no una amplificación en ese proceso.

Sintesis conceptuales 1.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.