Resumen Renal Pablo. C Página 1
Renal
Agua corporal
El agua corporal total supone un 45-50% del peso corporal en mujeres y un 55-60% en hombres adultos. Las diferencias
entre hombres y mujeres se deben a las variaciones de la composición corporal, sobre todo de masa muscular, tejido
adiposo y hueso. El músculo contiene aproximadamente la mitad del agua corporal.
El porcentaje de agua corporal varía a lo largo de la vida. Es máximo en el recién nacido (aproximadamente 75%) y
disminuye a lo largo de la infancia. Se mantiene estable en los adultos jóvenes y vuelve a disminuir a partir de los 50
años de edad.
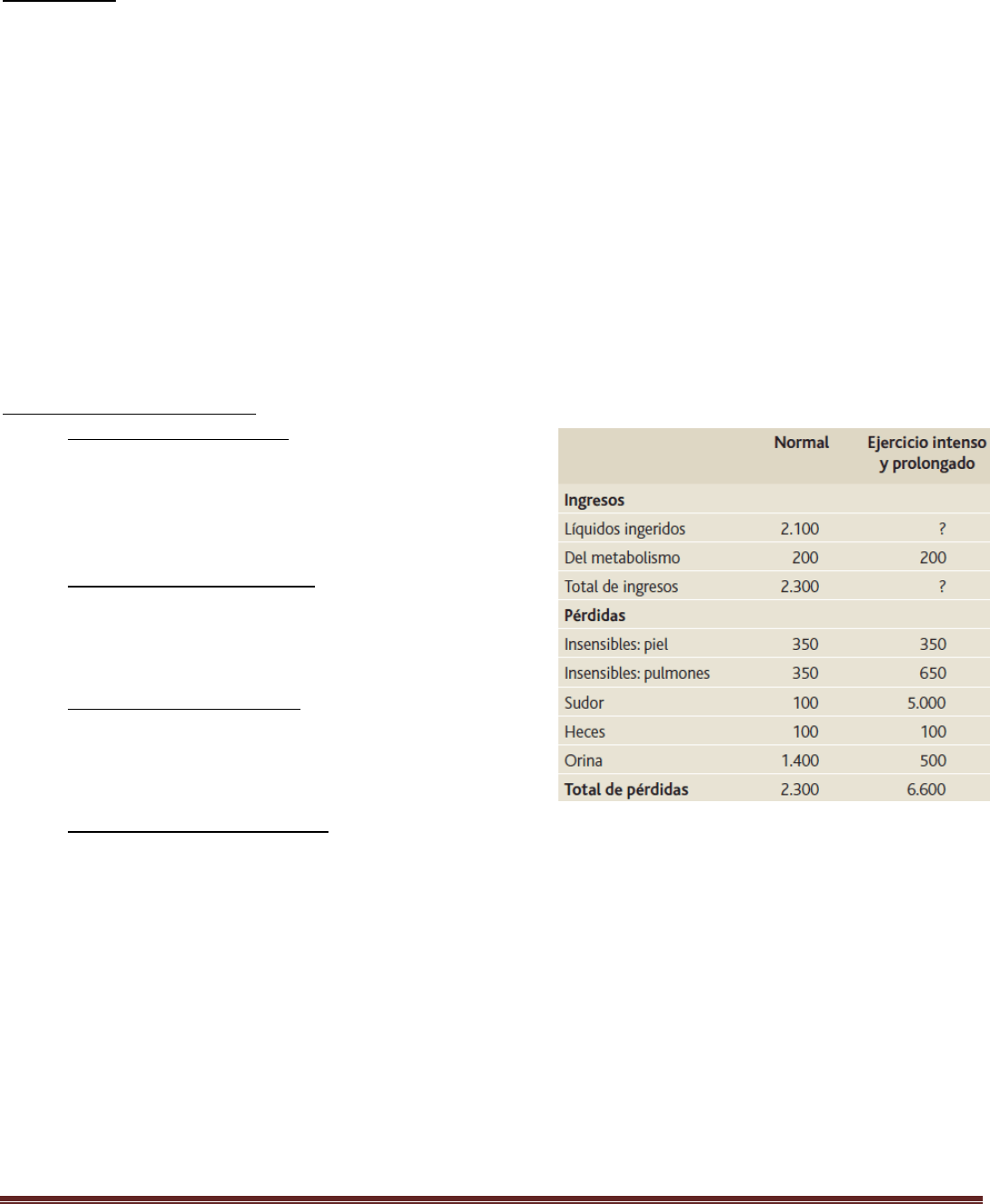
El agua ingresa en el cuerpo a través de dos fuentes principales:
1) se ingiere en forma de líquidos o de agua de los alimentos, que juntos suponen alrededor de 2.100 ml/día de
líquidos corporales,
2) se sintetiza en el cuerpo por la oxidación de los hidratos de carbono, en una cantidad de unos 200 ml/día.
Estos mecanismos proporcionan un ingreso total de agua de unos 2.300 ml/día. Sin embargo, la ingestión de agua es
muy variable entre las diferentes personas e incluso en la misma persona en diferentes días en función del clima, los
hábitos e incluso el grado de actividad física.
Pérdida diaria de agua corporal
Pérdida insensible de agua: Parte de las pérdidas de agua
no puede regularse de manera precisa. Por ejemplo, los
seres humanos experimentan una pérdida continua de
agua por evaporación de las vías aéreas y por difusión a
través de la piel.
Pérdida de líquido en el sudor: El volumen de sudor es
normalmente de unos 100 ml/día, pero en un clima muy
cálido o durante el ejercicio intenso, la pérdida de líquidos
en el sudor aumenta en ocasiones a 1-2 l/h.
Pérdida de agua en las heces: Solo se pierde normalmente
una pequeña cantidad de agua (100 ml/día) en las heces.
Esta pérdida puede aumentar a varios litros al día en
personas con diarrea intensa.
Pérdida de agua por los riñones: El resto del agua perdida se excreta en la orina por los riñones. Múltiples
mecanismos controlan la intensidad de la producción de orina. De hecho, el medio más importante por el que el
cuerpo mantiene un equilibrio entre los ingresos y las pérdidas, así como el equilibrio entre el ingreso y la salida
de la mayoría de los electrólitos en el cuerpo, es controlando la intensidad con la que los riñones excretan estas
sustancias. Por ejemplo, el volumen de orina puede ser tan solo de 0,5 l/día en una persona deshidratada o tan
alta como de 20 l/día en una persona que ha bebido cantidades enormes de agua.
Resumen Renal Pablo. C Página 2
Ionograma
Sodio:
Ingreso: los electrólitos del cuerpo, como el sodio, el
cloro y el potasio. En algunas personas, la ingestión
de sodio puede ser tan solo de 20 mEq/día, mientras
que en otros puede ser tan alta como de 300-500
mEq/día.
Egreso: Por heces, orina y transpiración
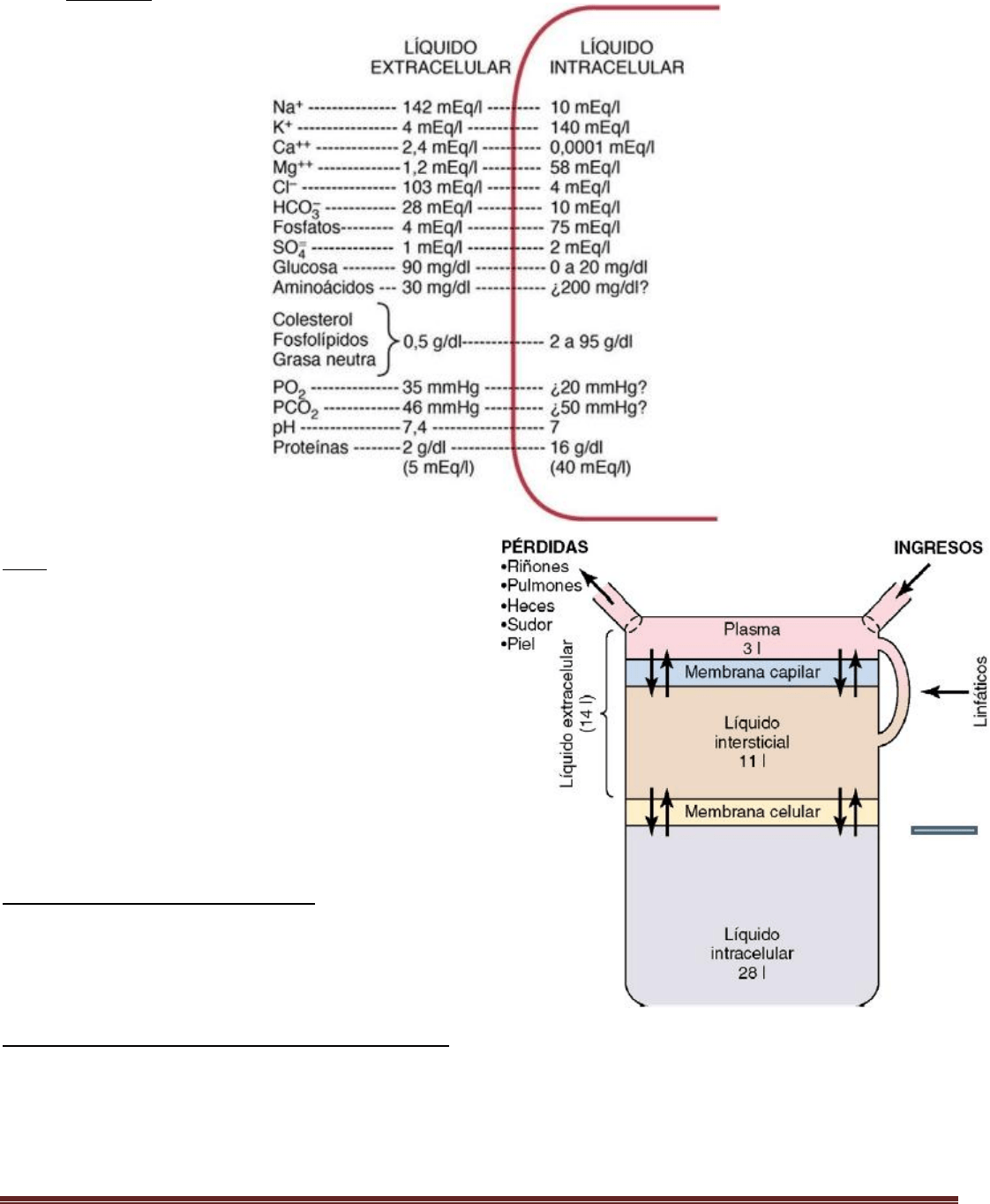
Compartimientos del líquido corporal
El líquido corporal total se distribuye sobre todo entre dos
compartimientos: el líquido extracelular y el líquido
intracelular. El líquido extracelular se divide en el líquido
intersticial y el plasma sanguíneo.
Compartimiento del líquido intracelular Unos 28 de los 42 l
de líquido corporal están contenidos de los 100 billones de
células y se les denomina en conjunto líquido intracelular.
Por tanto, el líquido intracelular constituye alrededor del
40% del peso corporal total en una persona «media».
Compartimiento del líquido extracelular (medio interno)
Todos los líquidos del exterior de las células se denominan en conjunto líquido extracelular y constituyen alrededor del
20% del peso corporal, o unos 14 l en un hombre adulto de 70 kg. Los dos compartimientos más grandes del líquido
extracelular son el líquido intersticial, que supone hasta más de tres cuartas partes (11 l) del líquido extracelular, y el
plasma, que supone casi una cuarta parte del líquido extracelular o unos 3 l. El plasma es la parte no celular de la sangre;
intercambia sustancias continuamente con el líquido intersticial a través de poros de las membranas capilares. Estos
poros son muy permeables a casi todos los solutos del líquido extracelular excepto a las proteínas, por lo que los líquidos
Resumen Renal Pablo. C Página 3
extracelulares se están mezclando constantemente, de manera que el plasma y el líquido intersticial tienen casi la
misma composición excepto en lo referente a las proteínas, que están más concentradas en el plasma.
Volumen sanguíneo
La sangre contiene líquido extracelular (el líquido del plasma) y líquido intracelular (el líquido de los eritrocitos). Sin
embargo, la sangre se considera un compartimiento líquido separado porque está contenida en su propia cámara, el
aparato circulatorio.
Ósmosis
En ciertas condiciones se puede producir una diferencia de concentración del agua a través de la membrana. Cuando se
produce esto, tiene lugar un movimiento neto de agua a través de la membrana celular, haciendo que la célula se hinche
o que se contraiga, dependiendo de la dirección del movimiento del agua. Este proceso de movimiento neto del agua
que se debe a la producción de una diferencia de la concentración del agua se denomina ósmosis.
El término «osmolaridad»
Osmolaridad es la concentración molar expresada en moles por litro.
«Osmolalidad»:
El osmol Para expresar la concentración de una solución en función del número de partículas se utiliza la unidad
denominada osmol en lugar de los gramos. Un osmol es el peso molecular-gramo de un soluto osmóticamente activo.
Por tanto, 180 g de glucosa, que es el peso molecular-gramos de la glucosa, son equivalentes a un osmol de glucosa
porque la glucosa no se disocia en iones. Si un soluto se disocia en dos iones, un peso molecular-gramo del soluto se
convertirá en dos osmoles porque el número de partículas osmóticamente activas es ahora el doble que en el caso del
soluto no disociado. Por tanto, cuando está totalmente disociado, un peso molecular-gramo de cloruro sódico, 58,5 g, es
igual a dos osmoles.
Los riñones
Tienen una función importante: eliminar del cuerpo los materiales de desecho que se han ingerido o que ha producido el
metabolismo. Una segunda función que es especialmente crítica es controlar el volumen y la composición de los
electrólitos de los líquidos corporales. En lo que respecta al agua y casi todos los electrólitos del cuerpo, el equilibrio
entre los ingresos (debidos a la ingestión y a la producción metabólica) y las salidas (debidas a la excreción o al consumo
metabólico) lo mantienen en gran medida los riñones. Esta función reguladora de los riñones mantiene el ambiente
interno estable necesario para que las células desempeñen sus diversas actividades.
Los riñones ejercen numerosas funciones homeostáticas, entre
ellas las siguientes:
• Excreción de productos metabólicos de desecho y sustancias
químicas extrañas:
Eliminación de los productos de desecho del metabolismo que ya
no necesita el cuerpo. Estos productos son la urea (del
metabolismo de los aminoácidos), la creatinina (de la creatina
muscular), el ácido úrico (de los ácidos nucleicos), los productos
finales del metabolismo de la hemoglobina (como la bilirrubina) y
los metabolitos de varias hormonas. Estos productos de desecho
deben eliminarse del cuerpo tan rápidamente como se producen.
Los riñones también eliminan la mayoría de las toxinas y otras
sustancias extrañas que el cuerpo produce o ingiere, como los
pesticidas, los fármacos y los aditivos alimentarios.
Resumen Renal Pablo. C Página 4
• Regulación de los equilibrios hídrico y electrolítico.
Para el mantenimiento de la homeostasis, la excreción de agua y electrólitos debe corresponderse de forma precisa con
su ingreso. Si los ingresos superan a la excreción, la cantidad de esa sustancia en el cuerpo aumentará. Si la ingestión es
menor que la excreción, la cantidad de esa sustancia en el cuerpo se reducirá. El mantenimiento de la vida depende de
la restauración del equilibrio de agua y electrólitos.
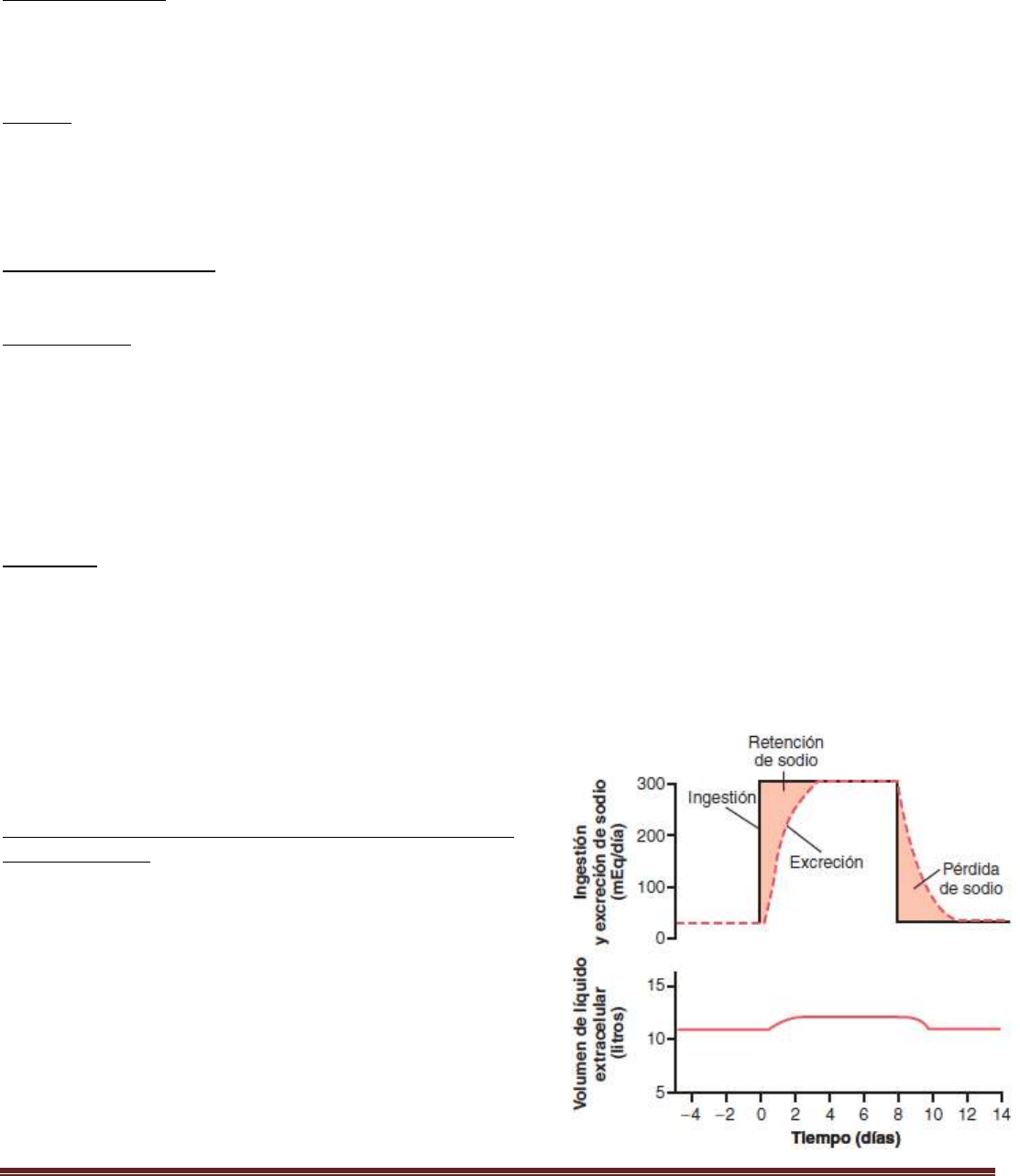
La ingestión de agua y de muchos electrólitos está gobernada sobre todo por los hábitos de bebida y comida de la
persona, y los riñones deben ajustar su excreción a su ingestión. La figura muestra la respuesta de los riñones a un
incremento brusco de 10 veces de la ingestión de sodio, desde un valor bajo de 30 mEq/día a un valor alto de 300
mEq/día. En los 2 a 3 días siguientes del aumento de la ingestión de sodio, la excreción renal también aumenta hasta
alrededor de 300 mEq/día, de manera que se restablece rápidamente el equilibrio entre la ingestión y la salida. Pero
durante la adaptación renal en esos 2 a 3 días a la elevada ingestión de sodio se produce una acumulación modesta de
sodio que incrementa ligeramente el volumen de líquido extracelular y desencadena cambios hormonales y otras
respuestas compensadoras que indican a los riñones que aumenten la excreción de sodio.
• Regulación de la osmolalidad del líquido corporal y de las concentraciones de electrólitos.
• Regulación de la presión arterial.
La regulación a largo plazo de la presión arterial al excretar cantidades variables de sodio y agua. Los riñones también
contribuyen a la regulación a corto plazo de la presión arterial mediante la secreción de hormonas y factores o
sustancias vasoactivos, como la renina, que dan lugar a la formación de productos vasoactivos (p. ej., la angiotensina II).
• Regulación del equilibrio acido básico.
Los riñones contribuyen a la regulación acidobásica junto a los pulmones y los amortiguadores del líquido corporal
mediante la excreción de ácidos y la regulación de los depósitos de amortiguadores en el líquido corporal. Los riñones
son el único medio de eliminar ciertos tipos de ácidos, como el ácido sulfúrico y el ácido fosfórico, que genera el
metabolismo de las proteínas.
• Regulación de la producción de eritrocitos.
Los riñones secretan eritropoyetina, que estimula la producción de eritrocitos en las células madre hematopoyéticas de
la médula ósea. Un estímulo importante para la secreción de eritropoyetina por los riñones es la hipoxia. Los riñones son
responsables normalmente de la mayor parte de la eritropoyetina secretada a la circulación.
• Secreción, metabolismo y excreción de hormonas.
Los riñones producen la forma activa de la vitamina D, 1,25-
dihidroxivitamina D 3 (calcitriol), mediante la hidroxilación de
esta vitamina.
• Gluconeogenia.
Sintetizan glucosa a partir de los aminoácidos y otros precursores
durante el ayuno prolongado, un proceso denominado
gluconeogenia.
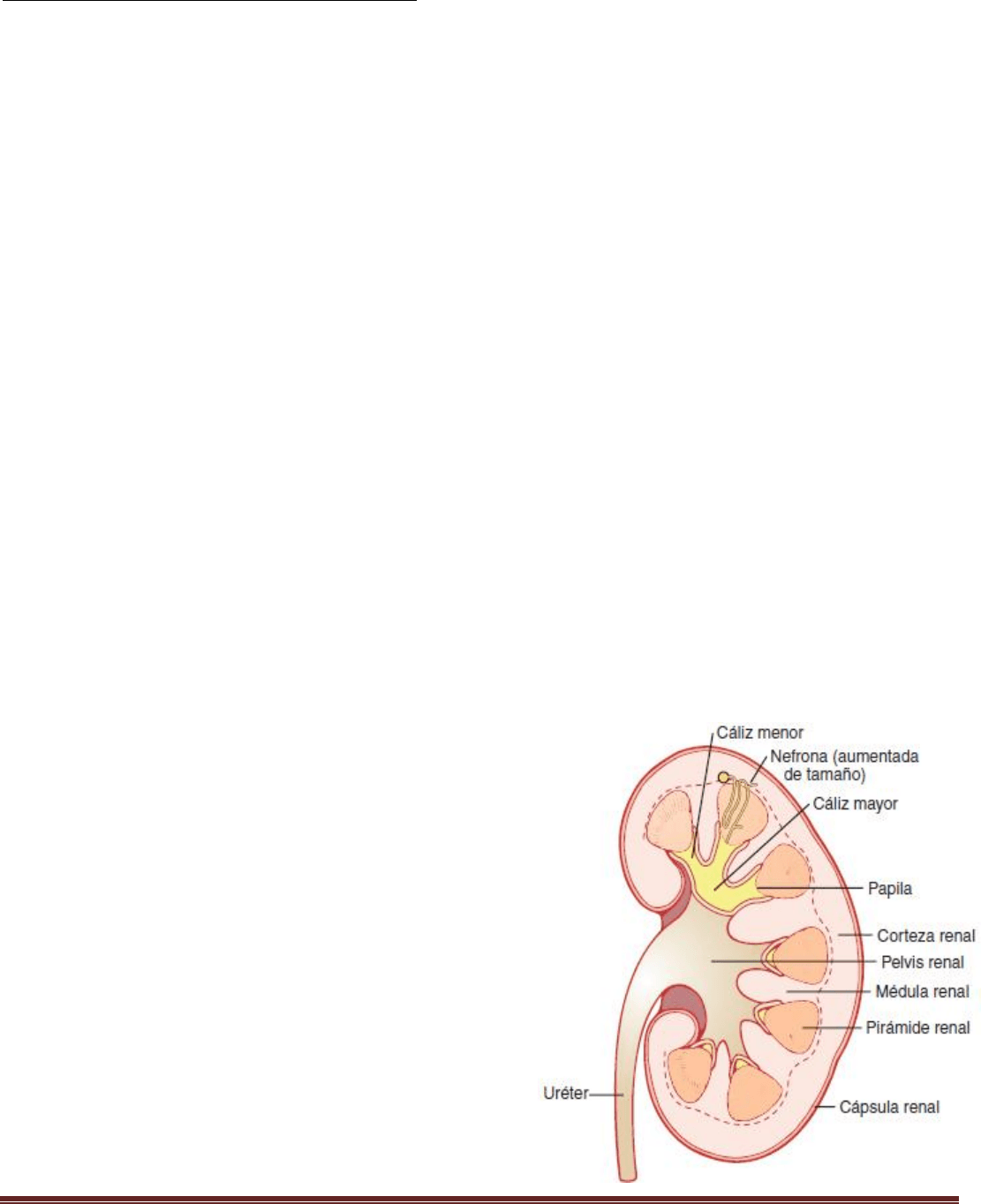
Anatomía fisiológica de los riñones
Cada riñón de un ser humano adulto normal pesa unos 150g y
tiene el tamaño aproximado de un puño cerrado. La cara medial
de cada riñón contiene una región con una muesca, llamada hilio,
por la que pasan la arteria y vena renales, los linfáticos, la
inervación y el uréter, que transporta la orina final desde el riñón
hasta la vejiga, donde se almacena hasta que esta se vacía.
Resumen Renal Pablo. C Página 5
El riñón está rodeado de una cápsula fibrosa y tensa que protege sus delicadas estructuras internas. Si se cortan los
riñones de arriba abajo, las dos regiones principales que pueden verse son la corteza externa y las regiones internas de
la médula. La médula se divide en 8-10 masas de tejido en forma de cono llamadas pirámides renales. La base de cada
pirámide se origina en el borde entre la corteza y la médula y termina en la papila, que se proyecta en el espacio de la
pelvis renal, una continuación en forma de abanico de la porción superior del uréter.
Las paredes de los cálices, la pelvis y el uréter contienen elementos contráctiles que empujan la orina hacia la vejiga,
donde se almacena hasta que se vacía en la micción.
La nefrona (también nefrón)
Es una unidad estructural y funcional
básica del riñón, responsable de la
purificación de la sangre. Su principal
función es filtrar la sangre para
regular el agua y las sustancias
solubles, reabsorbiendo lo que es
necesario y excretando el resto
como orina. Está situada
principalmente en la corteza renal.
Cada riñón humano contiene
alrededor de 800.000 a 1.000.000 de
nefronas, cada una de las cuales es
capaz de formar orina. El riñón no
puede regenerar nefronas nuevas. Por
tanto, en la lesión, la enfermedad o el
envejecimiento renal normal, el
número de nefronas se reduce
gradualmente.
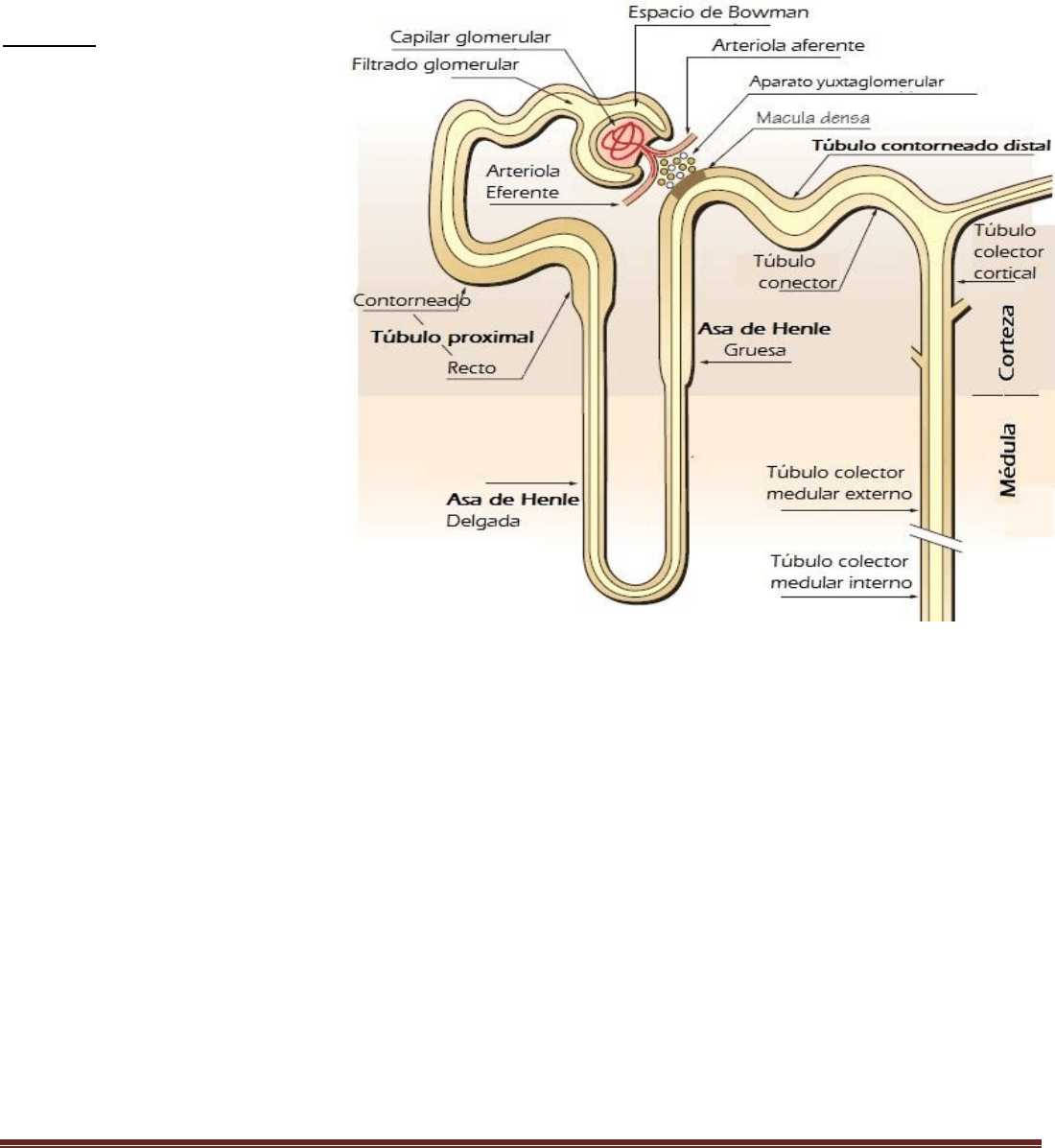
Cada nefrona contiene: 1) un penacho de capilares glomerulares llamado glomérulo, por el que se filtran grandes
cantidades de líquido desde la sangre, y 2) un túbulo largo en el que el líquido filtrado se convierte en orina en su
camino a la pelvis del riñón. El glomérulo contiene una red de capilares glomerulares que se ramifican y anastomosan
(ej: cuando dos venas o arterias o lo que sea anastomosan, quiere decir que confluyen, se unen en algún punto). Los
capilares glomerulares están revestidos de células epiteliales y todo el glomérulo está cubierto por la cápsula de
Bowman. El líquido filtrado desde los capilares glomerulares circula hacia la cápsula de Bowman y después al túbulo
proximal, que se encuentra en la corteza del riñón. Desde el túbulo proximal, el líquido fluye hacia el asa de Henle, que
desciende hasta la médula renal. Cada asa consta de una rama descendente y otra ascendente. Después de que la rama
ascendente del asa ha vuelto a la corteza, su pared se engruesa mucho y se denomina segmento grueso del asa
ascendente.
Al final de la rama ascendente gruesa hay un segmento corto, que tiene en su pared una placa de células epiteliales
especializadas conocida como mácula densa.
Más allá de la mácula densa el líquido entra en el túbulo distal, que, como el túbulo proximal, se dispone en la corteza
renal. Al túbulo distal le siguen el túbulo conector y el túbulo colector cortical, que conduce al conducto colector
cortical. Las partes iniciales de 8 a 10 conductos colectores corticales se unen para formar un solo conducto colector
mayor que discurre hacia abajo al interior de la médula y se convierte en el conducto colector medular. Los conductos
colectores se funden para formar progresivamente conductos cada vez mayores que finalmente se vacían en la pelvis
renal a través de las puntas de las papilas renales.
Resumen Renal Pablo. C Página 6
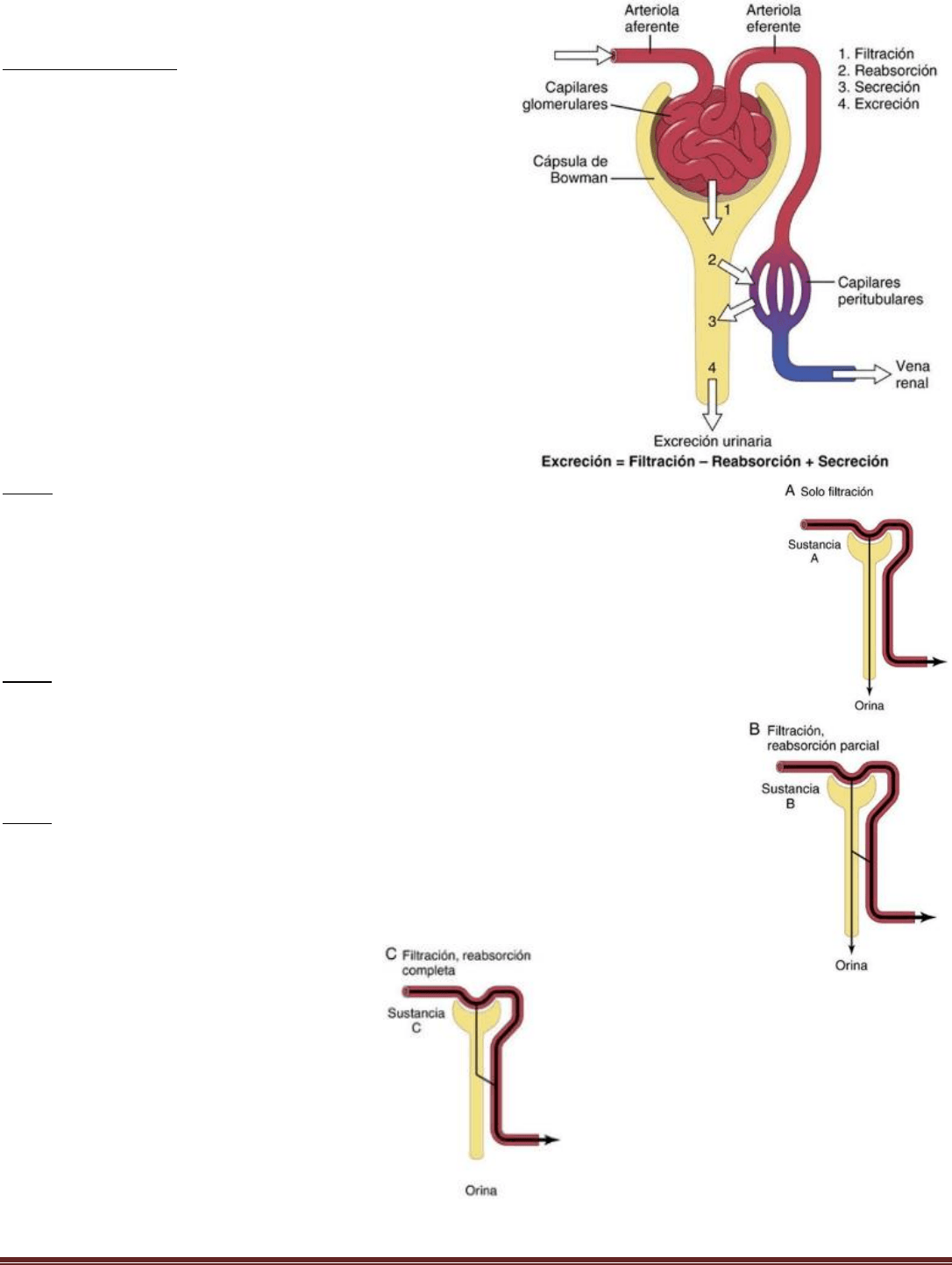
La formación de orina
La intensidad con la que se excretan diferentes sustancias en la
orina representa la suma de tres procesos renales: 1) la
filtración glomerular; 2) la reabsorción de sustancias de los
túbulos renales hacia la sangre, y 3) la secreción de sustancias
desde la sangre hacia los túbulos renales.
La formación de orina comienza cuando una gran cantidad de
líquido que casi no dispone de proteínas se filtra desde los
capilares glomerulares a la cápsula de Bowman. A medida que
el líquido abandona la cápsula de Bowman y pasa a través de
los túbulos, se modifica por la reabsorción de agua y solutos
específicos de nuevo hacia la sangre o por la secreción de otras
sustancias desde los capilares peritubulares hacia los túbulos.
Tipos de formación de la orina
Tipo A
La sustancia mostrada en el panel A se filtra libremente en los capilares glomerulares, pero no
se reabsorbe ni secreta, de forma que su excreción es igual a la intensidad con que se filtra. Los
riñones manejan de esta forma ciertos productos de desecho, como la creatinina, lo que
permite excretar casi todo lo que se filtra.
Tipo B
En el panel B, la sustancia se filtra libremente pero se reabsorbe parcialmente de los túbulos
hacia la sangre. Luego la excreción urinaria es menor que la filtración en los capilares
glomerulares. En este caso, la excreción se calcula como la filtración menos la reabsorción.
Este patrón es típico de muchos electrólitos del cuerpo, como iones sodio y cloruro.
Tipo C
En el panel C, la sustancia se filtra libremente en los capilares glomerulares pero no se
excreta en la orina porque toda la sustancia filtrada se reabsorbe de los túbulos de nuevo a
la sangre. Este patrón aparece en algunas sustancias nutritivas de la sangre, como los
aminoácidos y la glucosa, lo que permite conservarlas en los líquidos corporales.
Resumen Renal Pablo. C Página 7
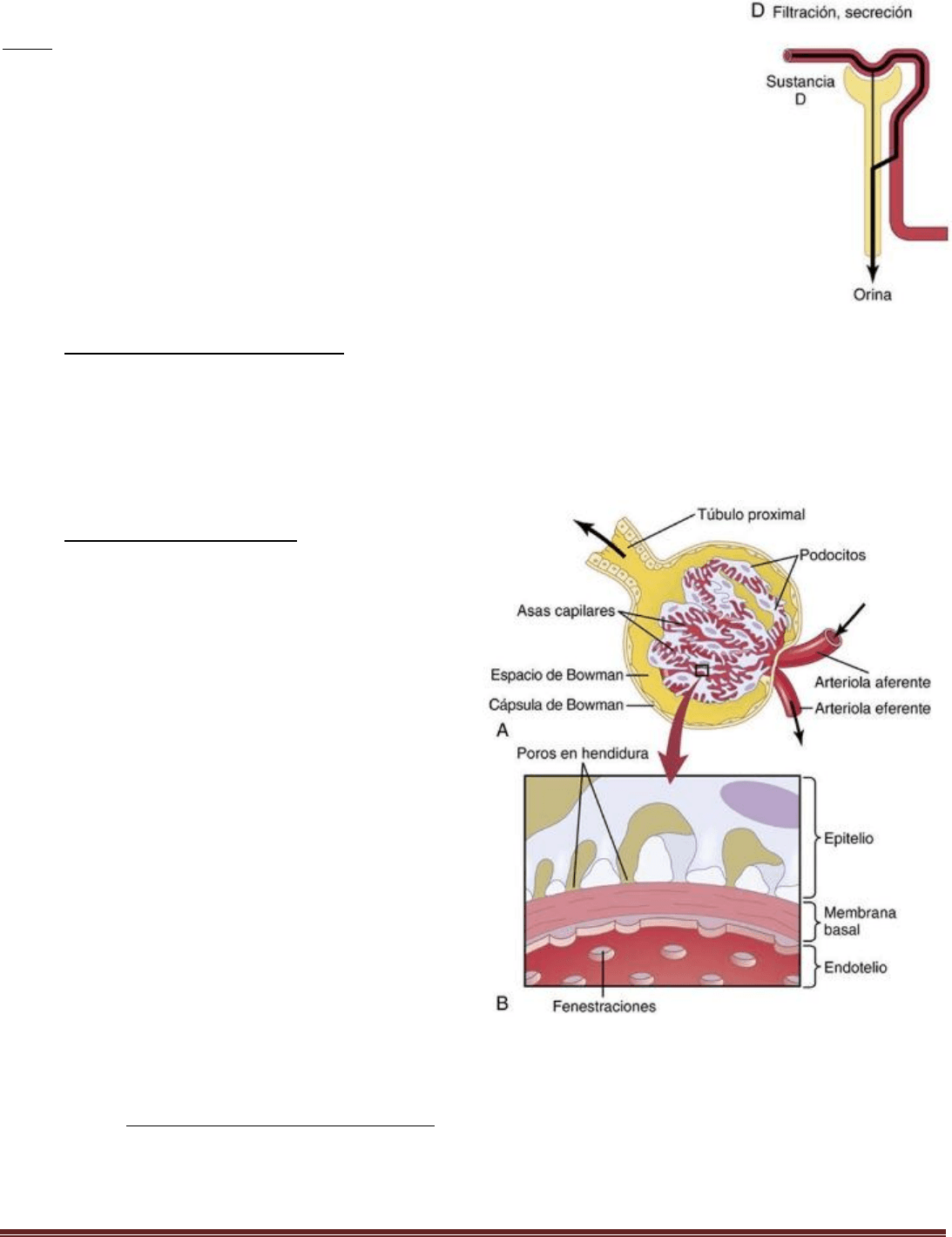
Tipo D
La sustancia del panel D se filtra libremente en los capilares glomerulares y no se reabsorbe,
pero se secretan cantidades adicionales de esta sustancia desde la sangre capilar peritubular
a los túbulos renales. Este patrón es frecuente en los ácidos orgánicos y las bases, lo que
permite eliminarlos rápidamente.
Filtración glomerular:
El primer paso en la formación de orina es la filtración de grandes cantidades de líquidos a
través de los capilares glomerulares en la cápsula de Bowman, casi 180 l al día. La mayor
parte de este filtrado se reabsorbe, lo que deja únicamente 1 l aproximadamente de líquido
para su excreción al día, si bien la tasa de excreción renal de líquidos puede ser muy variable
dependiendo de la ingestión de líquidos.
Composición del filtrado glomerular
Como la mayoría de los capilares, los capilares glomerulares son relativamente impermeables a las proteínas, de manera
que el líquido filtrado (llamado filtrado glomerular) carece prácticamente de proteínas y elementos celulares, incluidos
los eritrocitos. Las concentraciones de otros constituyentes del filtrado glomerular, como la mayoría de las sales y
moléculas orgánicas, son similares a las concentraciones en el plasma. Las excepciones a esta generalización son algunas
sustancias con un peso molecular bajo, como el calcio y los ácidos grasos, que no se filtran libremente porque están
unidas parcialmente a las proteínas plasmáticas.
Membrana capilar glomerular
La membrana capilar glomerular es similar a la de otros
capilares, excepto en que tiene tres capas principales (en
lugar de las dos habituales): 1) el endotelio del capilar; 2)
una membrana basal, y 3) una capa de células epiteliales
(podocitos) rodeando a la superficie externa de la
membrana basal capilar. Juntas, estas capas forman la
barrera de filtración que, a pesar de sus tres capas, filtra
varios cientos de veces más agua y solutos que la
membrana capilar habitual.
La elevada filtración a través de la membrana capilar
glomerular se debe en parte a sus especiales
características. El endotelio capilar está perforado por
cientos de pequeños agujeros, llamados fenestraciones.
Rodeando al endotelio está la membrana basal, que consta
de una red de colágeno y fibrillas de proteoglucanos que
tienen grandes espacios a través de los cuales pueden
filtrarse grandes cantidades de agua y de solutos.
La parte final de la membrana glomerular es una capa de
células epiteliales que recubre la superficie externa del
glomérulo. Estas células no son continuas, sino que tienen
unas prolongaciones largas similares a pies (podocitos) que rodean la superficie externa de los capilares. Los podocitos
están separados por espacios llamados poros en hendidura a través de los cuales se mueve el filtrado glomerular.
La capacidad de filtración de los solutos
La membrana capilar glomerular es más gruesa que la de la mayoría de los otros capilares, pero es también mucho más
porosa y por tanto filtra líquido con mayor intensidad. A pesar de la elevada filtración, la barrera de filtración glomerular
filtra de modo selectivo las moléculas basándose en su tamaño y en su carga eléctrica.
Resumen Renal Pablo. C Página 8
Las moléculas grandes con carga negativa se filtran con menor facilidad que las moléculas con el mismo tamaño
molecular y cargas positivas
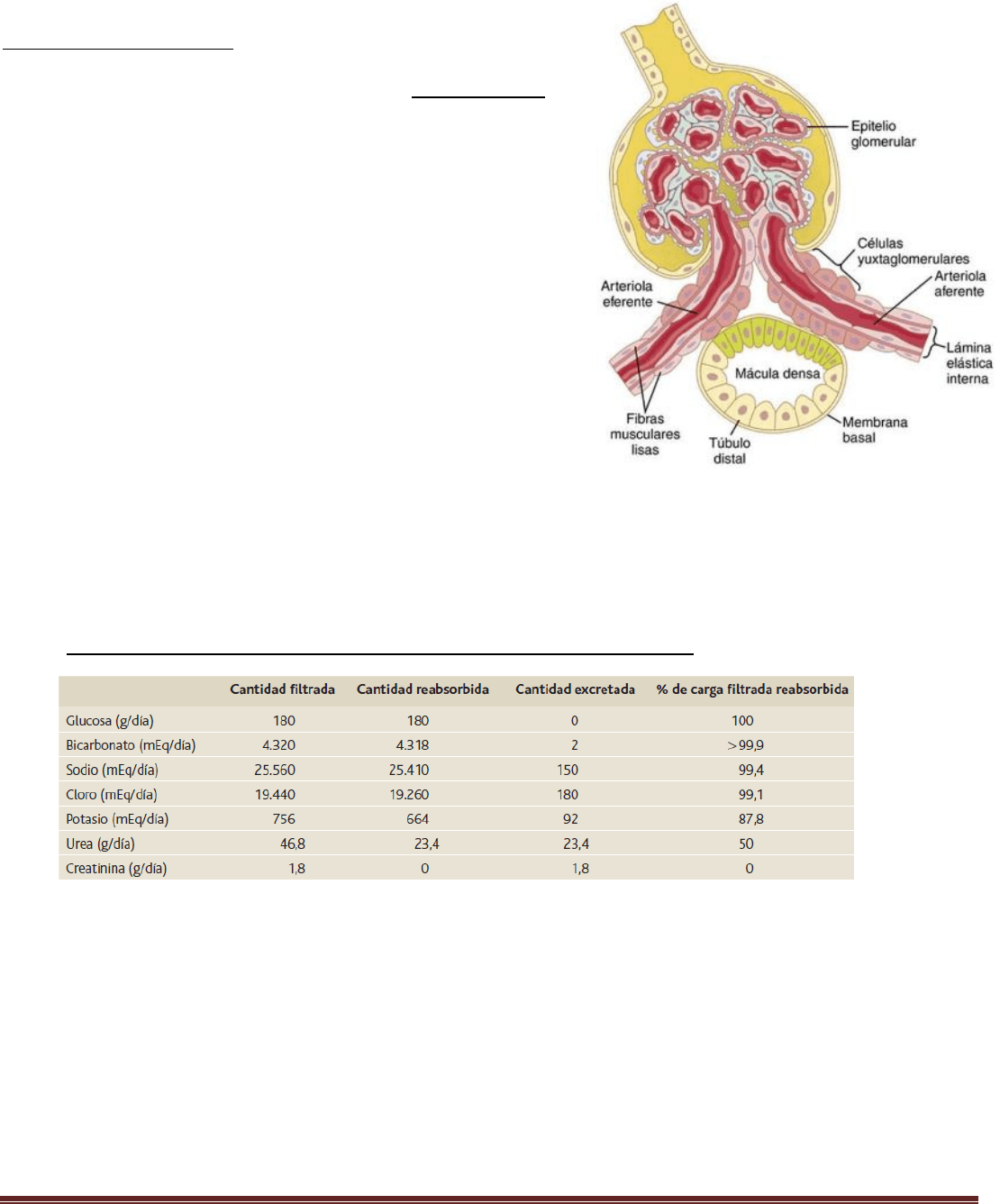
El complejo yuxtaglomerular consta de las células de la mácula densa
en la porción inicial del túbulo distal y las células yuxtaglomerulares en
las paredes de las arteriolas aferentes y eferentes. La mácula densa es
un grupo especializado de células epiteliales en los túbulos distales que
entra en estrecho contacto con las arteriolas aferente y eferente.
Las células de la mácula densa contienen aparato de Golgi, que son
orgánulos secretores intracelulares dirigidos hacia las arteriolas, lo que
indica que estas células pueden estar secretando una sustancia hacia
ellas. La reducción del cloruro de sodio en la mácula densa dilata las
arteriolas aferentes y aumenta la liberación de renina.
Esta retroalimentación ayuda a asegurar una llegada relativamente
constante de cloruro de sodio al túbulo distal y ayuda a evitar las
fluctuaciones falsas en la excreción renal que de otro modo tendrían
lugar.
Reabsorción y secreción tubular renal
A medida que el filtrado glomerular pasa por los túbulos renales, fluye
de forma secuencial a través de sus diferentes partes (el túbulo
proximal, el asa de Henle, el túbulo distal, el túbulo colector y,
finalmente, el conducto colector ) antes de eliminarse por la orina. A lo largo de este recorrido, algunas sustancias se
reabsorben selectivamente en los túbulos y vuelven a la sangre, mientras que otras se secretan desde la sangre a la luz
tubular. Finalmente, la orina ya formada y todas las sustancias que contiene representan la suma de los tres procesos
básicos que se producen en el riñón (la filtración glomerular, la reabsorción tubular y la secreción tubular)
Excreción= Filtración – Reabsorción + Secreción
La reabsorción tubular es cuantitativamente importante y altamente selectiva
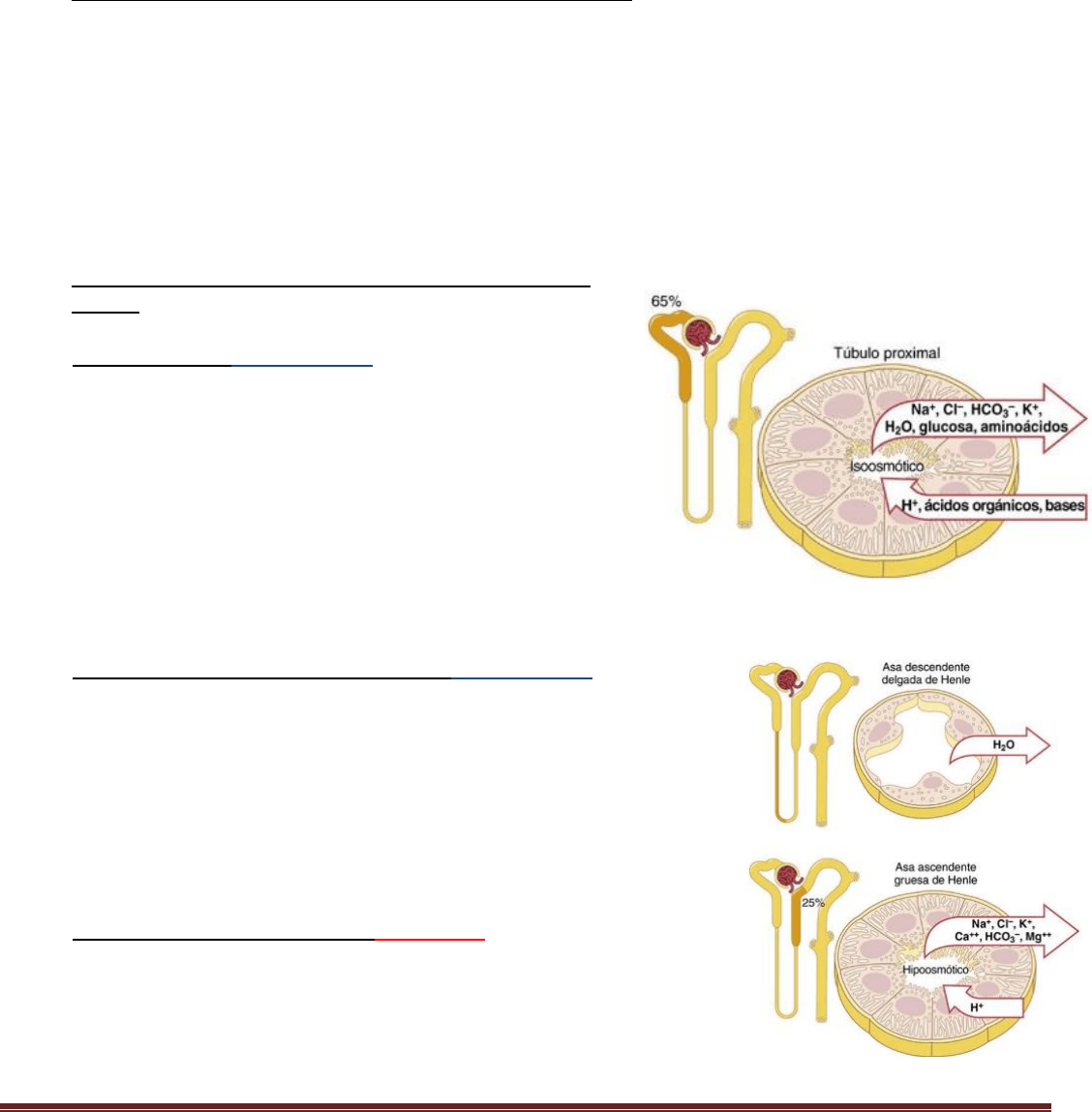
Resumen Renal Pablo. C Página 9
Sustancias, como la glucosa y los aminoácidos, se reabsorben del todo en los túbulos, por lo que su excreción urinaria es
prácticamente nula. Muchos de los iones del plasma, como el sodio, el cloro y el bicarbonato, también se reabsorben en
gran medida, pero su reabsorción y excreción urinarias varían mucho dependiendo de las necesidades del organismo. En
cambio, los productos de desecho, como la urea y la creatinina, se reabsorben mal en los túbulos y se excretan en
cantidades relativamente grandes. Por tanto, al controlar su reabsorción de diversas sustancias, los riñones regulan la
excreción de los solutos de forma independiente entre sí, una facultad que es esencial para el control preciso de la
composición de los líquidos corporales.
La reabsorción tubular comprende mecanismos pasivos y activos
Para que una sustancia se reabsorba, primero debe ser transportada a través de las membranas del epitelio tubular
hasta el líquido intersticial renal y luego a través de la membrana capilar peritubular hasta la sangre. Por tanto, la
reabsorción de agua y de solutos comprende una serie de pasos de transporte. La reabsorción a través del epitelio
tubular hacia el líquido intersticial se efectúa mediante un transporte activo y pasivo.
Luego, una vez producida la reabsorción a través de las células epiteliales tubulares hasta el líquido intersticial, el agua y
los solutos son transportados a través de las paredes de los capilares peritubulares para pasar a la sangre por
ultrafiltración (mayor parte del flujo), que está mediado por fuerzas hidrostáticas y coloidosmóticas. Los capilares
peritubulares se comportan de forma muy parecida a las terminaciones venosas de la mayoría de los demás capilares
porque existe una fuerza de reabsorción neta que mueve el líquido y los solutos desde el intersticio a la sangre.
Reabsorción y secreción a lo largo de diferentes partes de la
nefrona
Reabsorción en el
túbulo proximal
Alrededor del 65% de la carga filtrada de sodio y agua y algo menos del
cloro filtrado se reabsorbe normalmente en el túbulo proximal antes de
que el filtrado alcance el asa de Henle.
Las células epiteliales tubulares proximales tienen un metabolismo alto
y un gran número de mitocondrias para apoyar los potentes procesos
de transporte activo. Además, las células tubulares proximales tienen
un borde en cepillo extenso en el lado luminal (apical) de la membrana,
así como un laberinto extenso de canales intercelulares y basales, todos
los cuales proporcionan juntos una superficie de membrana extensa en
los lados luminal y vaso lateral del epitelio para un transporte rápido de los iones sodio y de otras sustancias.
Secreción de ácidos y bases orgánicos en el
túbulo proximal
El túbulo proximal es también un lugar importante para la secreción de ácidos y
bases orgánicas como las sales biliares, el oxalato, el urato y las catecolaminas.
Muchas de estas sustancias son productos finales del metabolismo y deben
eliminarse rápidamente del organismo. La secreción de estas sustancias en el
túbulo proximal más la filtración en el túbulo proximal por los capilares
glomerulares y la casi total falta de reabsorción por los túbulos contribuyen, todos
combinados, a su excreción rápida en la orina.
Transporte de solutos y agua en el
asa de Henle
El asa de Henle consta de tres segmentos con funciones diferentes: el segmento
descendente fino, el segmento ascendente fino y el segmento ascendente grueso.
Los segmentos descendente fino y ascendente fino, como sus nombres indican,
tienen membranas epiteliales finas sin bordes en cepillo, pocas mitocondrias y
niveles mínimos de actividad metabólica.
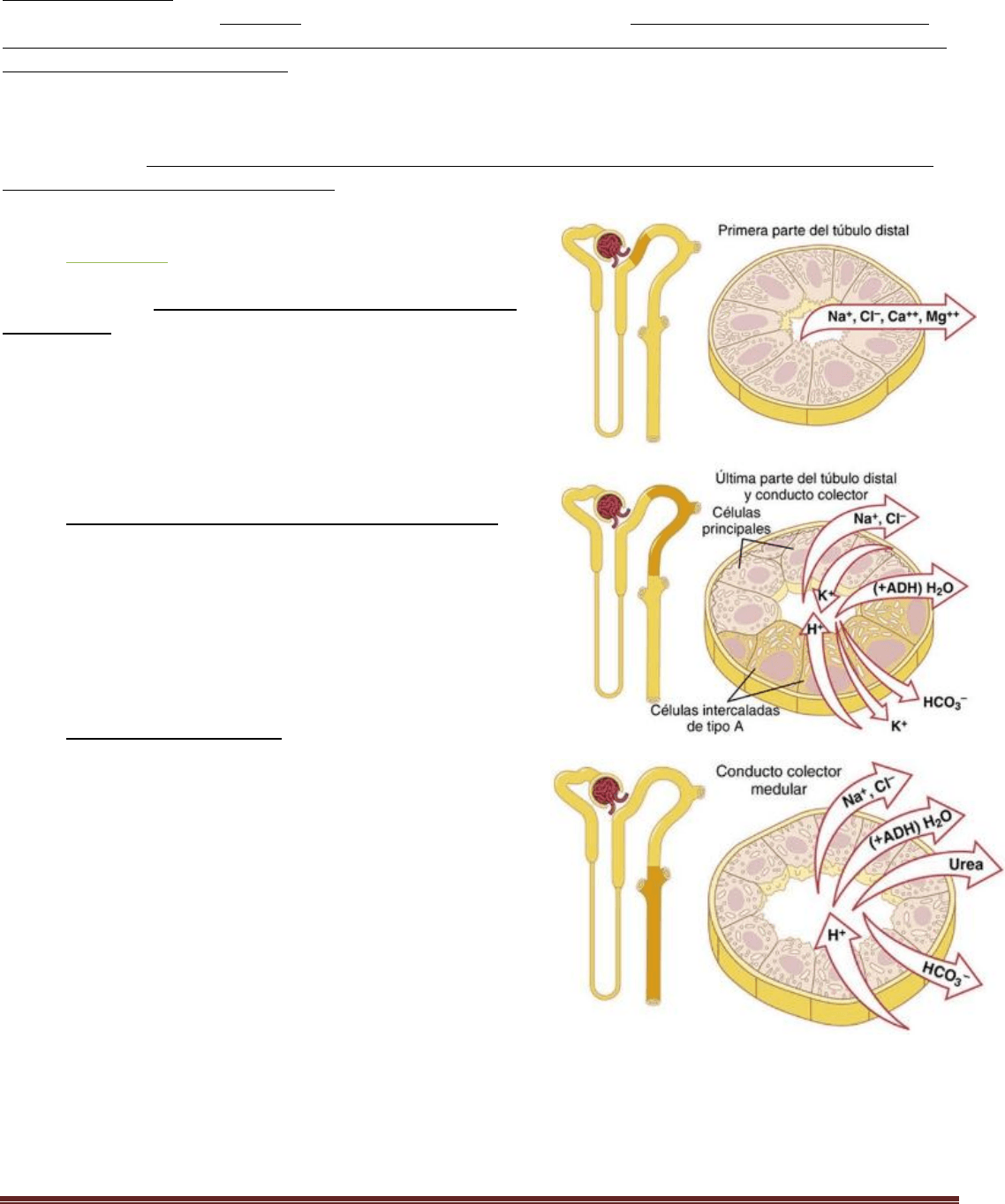
Resumen Renal Pablo. C Página 10
La parte descendente del segmento fino es muy permeable al agua y moderadamente a la mayoría de los solutos,
incluidos la urea y el sodio. La función de este segmento de la nefrona es sobre todo permitir la difusión simple de las
sustancias a través de sus paredes. Alrededor del 20% del agua filtrada se reabsorbe en el asa de Henle, y casi todo esto
ocurre en la rama descendente fina.
La rama ascendente, incluidas las porciones fina y gruesa, es casi impermeable al agua, una característica importante
para concentrar la orina. El segmento grueso del asa de Henle, que comienza en la mitad de la rama ascendente, tiene
células epiteliales gruesas con una elevada actividad metabólica y son capaces de una reabsorción activa del sodio, el
cloro y el potasio. Alrededor del 25% de las cargas filtradas de sodio, cloro y potasio se reabsorben en el asa de Henle,
sobre todo en la rama ascendente gruesa. También se reabsorben cantidades considerables de otros iones, como calcio,
bicarbonato y magnesio, en la rama ascendente gruesa del asa de Henle.
Túbulo distal
El segmento grueso de la rama ascendente del asa de Henle se vacía
en el túbulo distal. La porción inicial del túbulo distal conforma la
mácula densa. La siguiente parte del túbulo distal está muy
contorneada y reabsorbe con avidez la mayoría de los iones,
incluidos el sodio, el potasio y el cloro, pero es casi totalmente
impermeable al agua y a la urea. Por esta razón se le denomina
segmento diluyente, porque también diluye el líquido tubular.
Alrededor del 5% de la carga filtrada de cloruro de sodio se
reabsorbe en la primera parte del túbulo distal.
Porción final del túbulo distal y túbulo colector cortical
La segunda mitad del túbulo distal y el túbulo colector cortical
situado a continuación tienen características funcionales similares.
Están compuestos de dos tipos especiales de células, las células
principales y células intercaladas. Las células principales reabsorben
sodio y agua de la luz y secretan iones potasio a la luz. Las células
intercaladas de tipo A reabsorben iones potasio y secretan iones
hidrógeno a la luz tubular.
Conducto colector medular
Aunque los conductos colectores medulares reabsorben menos del
10% del agua y del sodio filtrados, son el lugar final de
procesamiento de la orina y, por ello, desempeñan una función
muy importante en la determinación de la eliminación final en la
orina de agua y de solutos.
Las características especiales de este segmento tubular son:
1. La permeabilidad al agua del conducto colector medular está
controlada por la concentración de ADH. Con concentraciones altas
de ADH (vasopresina), el agua se reabsorbe ávidamente en el
intersticio medular, lo que reduce el volumen de orina y concentra
la mayoría de los solutos en ella.
2. Al contrario que el túbulo colector cortical, el conducto colector medular es permeable a la urea y existen
transportadores de urea especiales que facilitan la difusión de la urea a través de las membranas luminales y vaso-
laterales. Luego parte de la urea tubular se reabsorbe en el intersticio medular, lo que ayuda a aumentar la osmolalidad
en esta región de los riñones y contribuye a la capacidad global de los riñones de formar una orina concentrada.
Resumen Renal Pablo. C Página 11
3. El conducto colector medular es capaz de secretar iones hidrógeno contra un gran gradiente de concentración, como
ocurre en el túbulo colector cortical. Luego el conducto colector medular también participa en la regulación del
equilibrio acidobásico.
Depuración plasmática o Clearance.
El aclaramiento o depuración renal es un parámetro mediante el cual es posible evaluar algunos aspectos de la función
renal.
El aclaramiento siempre está referido a una determinada sustancia, que de forma genérica se denomina X, donde X
puede ser cualquier sustancia disuelta en la sangre que pueda llegar hasta el riñón. Se observa cómo el volumen
de plasma sanguíneo (en ml), que por efecto de la función renal, queda libre de la sustancia X en la unidad de tiempo (en
minutos)..
Para esto se precisa una sustancia que sea eliminada solamente por filtración y que en su pasaje por los túbulos no sea
reabsorbida ni secretada. Hay sustancias que necesitan que pase mucha sangre para poder depurarla.
Mecanismo contracorriente
Es el mecanismo por el cual los riñones tienen la capacidad de formar orina diluida o concentrada. La cual se debe a la
capacidad de generar un intersticio hiperosmótico.
La hormona antidiurética controla la concentración
de la orina
El organismo cuenta con un sistema de retroalimentación
potente para regular la osmolaridad y la concentración de
sodio en el plasma que actúa modificando la excreción renal
de agua con independencia de la excreción de solutos. Un
efector fundamental de esta retroalimentación es la hormona
antidiurética (ADH), también llamada vasopresina. Cuando la
osmolaridad de los líquidos corporales aumenta por encima
de lo normal (los solutos de los líquidos corporales se
concentran demasiado), el lóbulo posterior de la hipófisis
secreta más ADH, que aumenta la permeabilidad al agua de
los túbulos distales y de los conductos colectores. Este
mecanismo eleva la reabsorción de agua y reduce el volumen
urinario, pero no altera notablemente la excreción renal de
los solutos. Cuando hay un exceso de agua en el organismo y
la osmolaridad del líquido extracelular se reduce, desciende
la secreción de ADH en el lóbulo posterior de la hipófisis, lo
que disminuye la permeabilidad al agua del túbulo distal y los
conductos colectores y conduce a la excreción de mayores
cantidades de orina más diluida. De este modo, la presencia o
falta de ADH determinan, en gran parte, que el riñón excrete
una orina diluida o concentrada.
Funcionamiento del Contracorriente
Cuando el filtrado glomerular está recién formado, su
osmolaridad es aproximadamente la misma que la del plasma (300 mOsm/l). Para excretar el exceso de agua es
necesario diluir el filtrado a medida que circula a lo largo del túbulo. Esta dilución se consigue reabsorbiendo más
solutos que agua, pero esto solo tiene lugar en ciertos segmentos del sistema tubular.
El líquido tubular continúa isoosmótico en el túbulo proximal
A medida que el líquido fluye a través del túbulo proximal, los solutos y el agua se reabsorben en igual proporción, de
forma que se producen pequeños cambios en la osmolaridad; así, el líquido del túbulo proximal permanece isoosmótico

Resumen Renal Pablo. C Página 12
respecto al plasma, con una osmolaridad aproximada de 300 mOsm/l. A medida que el líquido pasa por el asa
descendente de Henle, el agua se reabsorbe por ósmosis y el líquido tubular alcanza el equilibrio con el líquido
intersticial circundante de la médula renal, que es muy hipertónico (alrededor de dos a cuatro veces la osmolaridad del
filtrado glomerular en su origen). Por tanto, el líquido tubular va aumentando su concentración a medida que fluye hacia
la médula interna.
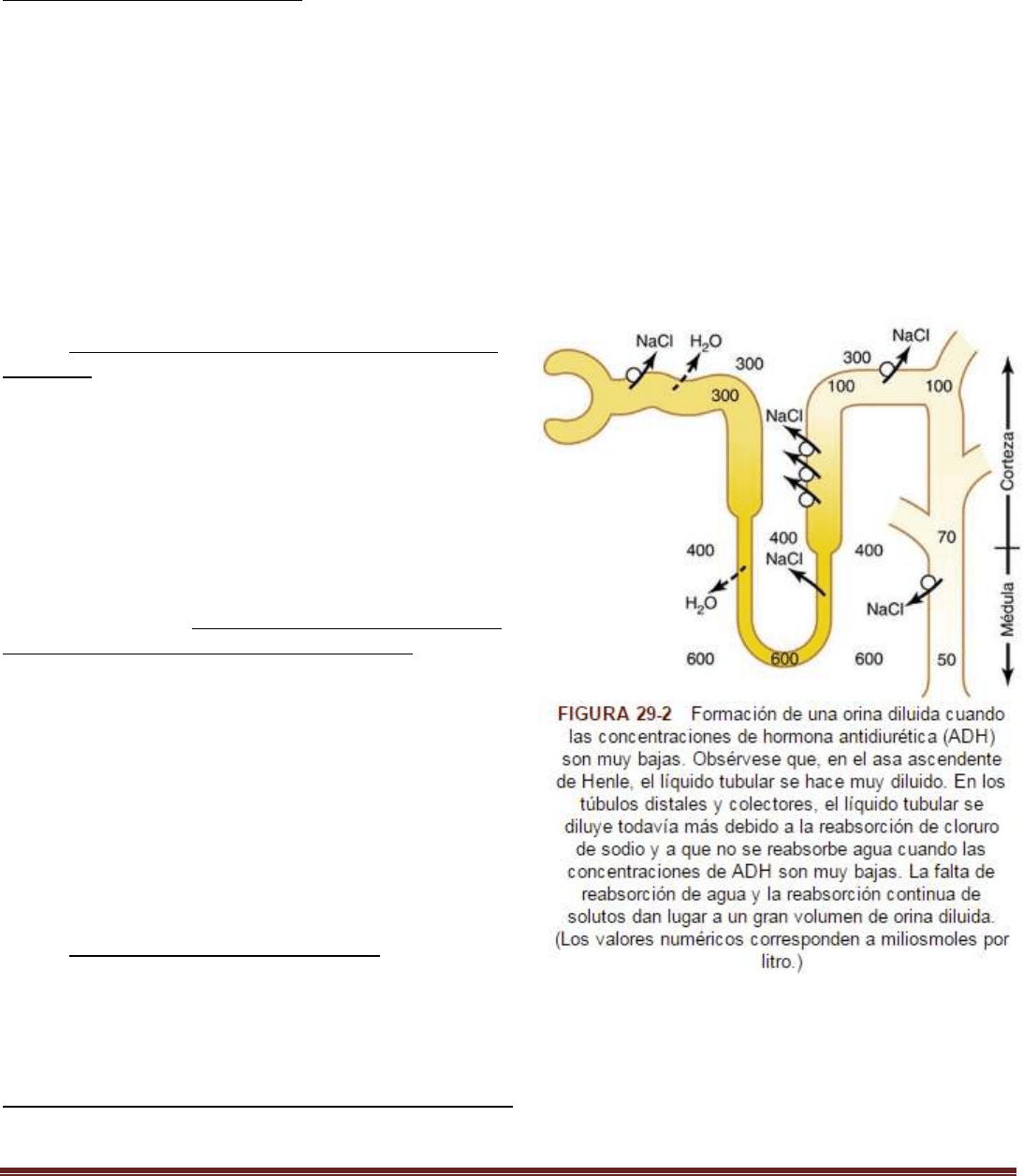
El líquido tubular se diluye en el asa ascendente de Henle
En la rama ascendente del asa de Henle, especialmente en el segmento grueso, se reabsorben con avidez el sodio, el
potasio y el cloro. Pero esta porción del segmento tubular es impermeable al agua incluso en presencia de grandes
cantidades de ADH. Por tanto, el líquido tubular va diluyéndose a medida que fluye por el asa ascendente de Henle hacia
la porción inicial del túbulo distal, con una osmolaridad que disminuye progresivamente hasta llegar a unos 100 mOsm/l
cuando el líquido entra en la porción inicial del segmento tubular distal. De este modo, independientemente de si hay o
no ADH, el líquido que abandona la parte inicial del segmento tubular distal es hipoosmótico, con una osmolaridad que
es tan solo alrededor de la tercera parte de la osmolaridad del plasma.
El líquido tubular se diluye aún más en los túbulos distales y colectores si no hay ADH
Cuando el líquido diluido de la porción inicial del túbulo distal pasa a la porción final del túbulo contorneado distal, el
conducto colector cortical y el conducto colector, se produce una reabsorción adicional de cloruro de sodio. Si no hay
ADH, esta porción del túbulo es también impermeable al agua, con lo que la reabsorción adicional de solutos hace que el
líquido tubular se diluya todavía más, reduciendo su osmolaridad hasta tan solo 50 mOsm/l. El hecho de que no se
reabsorba agua y continúe la reabsorción de solutos lleva a la formación de un gran volumen de orina diluida. En
resumen, el mecanismo de formación de orina diluida consiste en la reabsorción continua de solutos en los segmentos
distales del sistema tubular mientras no se reabsorbe el agua.
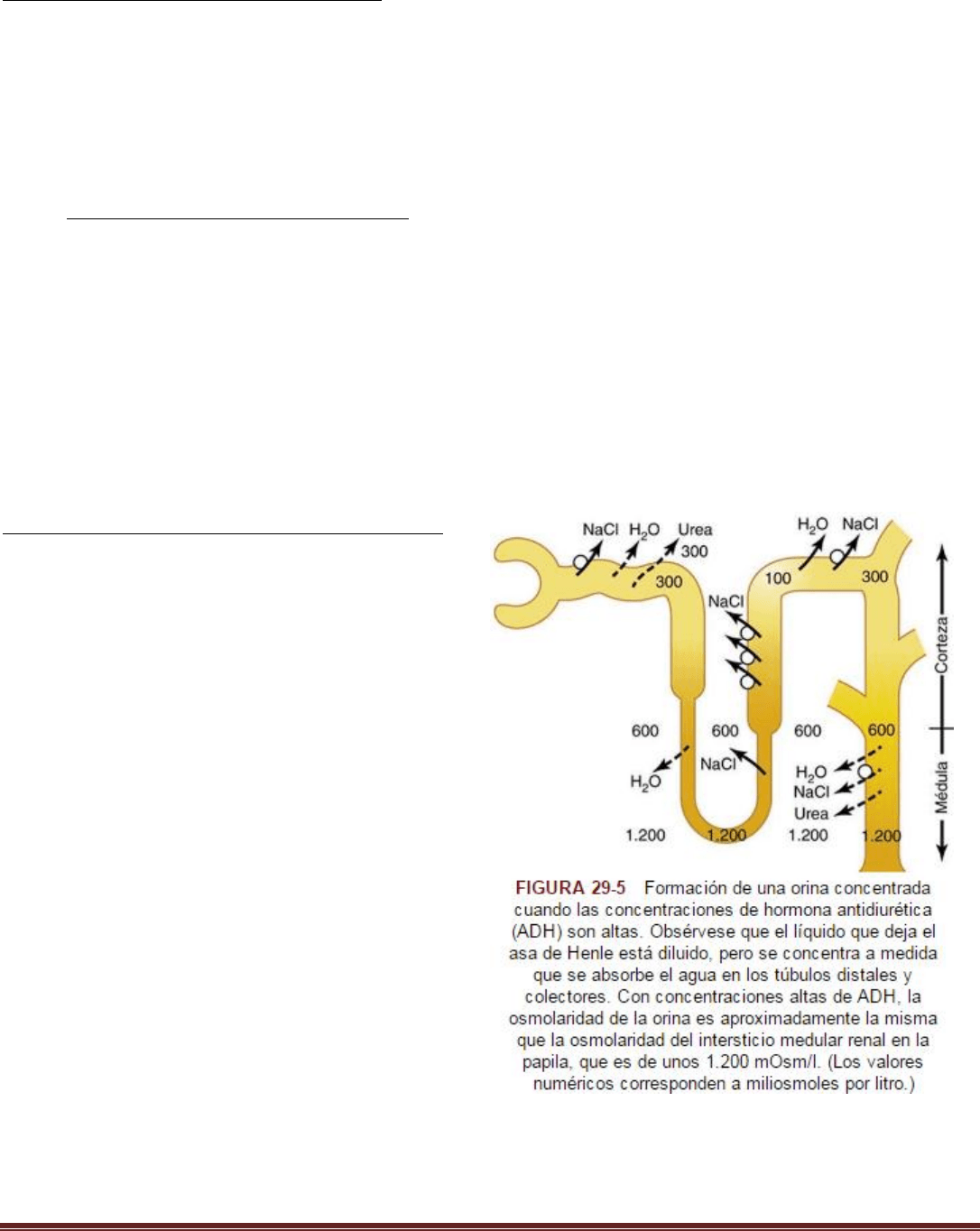
Los riñones conservan agua excretando una orina concentrada
Cuando hay una deficiencia de agua en el organismo, los riñones forman orina concentrada mediante la excreción
continua de solutos mientras aumenta la reabsorción de agua y reduce el volumen de orina formada. El riñón humano
puede lograr una concentración máxima de orina de 1.200-1.400 mOsm/l, cuatro a cinco veces la osmolaridad del
plasma.
Densidad específica de la orina
La densidad específica de la orina se usa a menudo en los centros clínicos para proporcionar una rápida estimación de la
concentración de solutos en orina. Cuanto más concentrada es la orina mayor es su densidad específica. En la mayoría
de los casos, la densidad específica de la orina aumenta linealmente al hacerlo su osmolaridad. No obstante, la densidad
específica de la orina es una medida del peso de solutos en un volumen dado de orina y, por tanto, está determinada
por el número y el tamaño de las moléculas de soluto. En cambio, la osmolaridad está determinada únicamente por el
número de moléculas de soluto en un volumen dado.
Requisitos para excretar una orina concentrada
Los requisitos básicos para formar una orinan concentrada son:
1) una concentración elevada de ADH, lo que aumenta la permeabilidad de los túbulos distales y los conductos
colectores al agua y permite a estos segmentos tubulares reabsorber agua con avidez.
2) una elevada osmolaridad del líquido del intersticio medular renal, que proporciona el gradiente osmótico necesario
para reabsorber el agua en presencia de concentraciones altas de ADH. El intersticio medular renal que rodea los
conductos colectores es normalmente hiperosmótico, de manera que cuando las concentraciones de ADH son altas, el
agua se mueve a través de la membrana tubular por ósmosis hacia el intersticio renal; desde aquí pasa de nuevo a la
sangre a través de los vasos rectos.
Resumen Renal Pablo. C Página 13
El mecanismo multiplicador de contracorriente
Depende de la disposición anatómica especial de las asas de Henle y de los vasos rectos, los capilares peritubulares
especializados de la médula renal. En el ser humano, alrededor del 25% de las nefronas son nefronas yuxtamedulares,
con asas de Henle y vasos rectos que se introducen en profundidad en la médula antes de volver a la corteza.
La osmolaridad del líquido intersticial en la médula renal es mucho mayor, y puede aumentar progresivamente de unos
1.200 a 1.400 mOsm/l en la punta pélvica de la médula. Esto significa que el intersticio medular renal ha acumulado
muchos más solutos que agua. Una vez que se consigue una concentración alta de solutos en la médula, se mantiene
mediante una entrada y salida equilibradas de solutos y de agua.
Características especiales del asa de Henle
Una razón importante de la elevada osmolaridad medular es el transporte activo de sodio y el cotransporte de iones
potasio, cloro y otros desde el asa ascendente gruesa de Henle hacia el intersticio. Esta bomba es capaz de establecer un
gradiente de concentración de unos 200 mOsm entre la luz tubular y el líquido intersticial. Debido a que la rama
ascendente gruesa es casi impermeable al agua, a los solutos bombeados no les sigue un flujo osmótico de agua hacia el
intersticio. De este modo, el transporte activo de sodio y de otros iones fuera del asa ascendente gruesa añade más
solutos que agua al intersticio medular renal. Hay cierta reabsorción pasiva de cloruro de sodio en la rama ascendente
fina del asa de Henle, que también es impermeable al agua, lo que contribuye más a elevar la concentración de solutos
que hay en el intersticio de la médula renal. La rama descendente del asa de Henle, al contrario que la rama ascendente,
es muy permeable al agua, y la osmolaridad del líquido tubular se iguala rápidamente a la osmolaridad de la médula
renal. Luego el agua se difunde fuera de la rama descendente del asa de Henle hacia el intersticio, y la osmolaridad del
líquido tubular aumenta gradualmente a medida que fluye hacia la punta del asa de Henle.
Función del túbulo distal y de los conductos colectores
Cuando el líquido tubular deja el asa de Henle y fluye
hacia el túbulo contorneado distal en la corteza renal, el
líquido se diluye, con una osmolaridad de solo 100
mOsm/l ( fig. 29-5 ). La primera parte del túbulo distal
diluye más el líquido tubular porque este segmento, como
el asa ascendente de Henle, transporta de forma activa
cloruro de sodio fuera del túbulo, pero es relativamente
impermeable al agua.
A medida que el líquido fluye hacia el túbulo colector
cortical, la cantidad de agua reabsorbida depende mucho
de la concentración plasmática de ADH. Si falta la ADH,
este segmento es casi impermeable al agua y no
reabsorbe agua sino que continúa reabsorbiendo solutos
y diluye más la orina. Cuando hay una concentración alta
de ADH, el túbulo colector cortical se hace muy
permeable al agua, de manera que se reabsorben ahora
grandes cantidades de agua desde el túbulo hacia el
intersticio de la corteza, donde es barrida por el flujo
rápido de los capilares peritubulares. El hecho de que
estas grandes cantidades de agua se reabsorban hacia la
corteza, en lugar de hacia la médula renal, ayuda a
conservar la elevada osmolaridad del líquido intersticial
medular. A medida que el líquido tubular fluye a lo largo
de los conductos colectores medulares, hay una mayor reabsorción de agua desde el líquido tubular hacia el intersticio,
pero la cantidad total de agua es relativamente pequeña comparada con la añadida al intersticio cortical. El agua
reabsorbida sale por los vasos rectos hacia la sangre venosa. Cuando hay concentraciones elevadas de ADH, los
conductos colectores se hacen permeables al agua, de manera que el líquido al final de los conductos colectores tiene

Resumen Renal Pablo. C Página 14
prácticamente la misma osmolaridad que el líquido intersticial de la médula renal, unos 1.200 mOsm/l. De este modo,
reabsorbiendo la mayor cantidad de agua posible, los riñones forman una orina muy concentrada, excretando
cantidades normales de solutos en la orina mientras añaden agua al líquido extracelular y compensan las deficiencias de
agua corporal.
La urea contribuye a la hiperosmolaridad del intersticio medular renal y a la formación de una orina concentrada
Hasta ahora hemos considerado solo la contribución del cloruro de sodio a la hiperosmolaridad del intersticio medular
renal. Pero la urea contribuye a alrededor de un 40-50% de la osmolaridad (500-600 mOsm/l) del intersticio medular
renal cuando el riñón está formando una orina concentrada al máximo. Al contrario que el cloruro de sodio, la urea se
reabsorbe de forma pasiva desde el túbulo. Cuando hay una deficiencia de agua y la concentración de ADH es alta, se
reabsorben de forma pasiva grandes cantidades de urea desde los conductos colectores medulares internos hacia el
intersticio. El mecanismo de reabsorción de la urea hacia la médula renal es como sigue. A medida que el agua fluye por
el asa ascendente de Henle y hacia los túbulos distal y colector cortical, se reabsorbe poca urea porque estos segmentos
son impermeables a la misma. En presencia de concentraciones elevadas de ADH, el agua se reabsorbe rápidamente
desde el túbulo colector cortical y la concentración de urea aumenta rápidamente porque la urea no es muy difusible en
esta parte del túbulo. Entonces, a medida que el líquido tubular fluye hacia los conductos colectores medulares internos,
todavía se reabsorbe más agua, lo que da lugar a una concentración de urea en el líquido incluso mayor. Esta elevada
concentración de urea en el líquido tubular del conducto colector medular interno hace que la urea difunda fuera del
túbulo hacia el líquido intersticial renal.
Esta difusión está muy facilitada por transportadores de la urea. Estos transportadores de la urea se activan por la acción
de la ADH, lo que aumenta el transporte de urea fuera del conducto colector medular interno incluso más cuando las
concentraciones de ADH están elevadas. El movimiento simultáneo de agua y urea fuera de los conductos colectores
medulares internos mantiene una elevada concentración de urea en el líquido tubular y, finalmente, en la orina, incluso
aunque la urea se reabsorba. La función fundamental de la urea en la contribución a la capacidad de concentrar la orina
se evidencia por el hecho de que las personas que ingieren una dieta hiperproteica, que origina grandes cantidades de
urea como productos de «desecho» nitrogenados, pueden concentrar la orina mucho mejor que las personas cuya
ingestión de proteínas y producción de urea son bajas. La malnutrición se acompaña de una baja concentración de urea
en el intersticio medular y de un deterioro considerable de la capacidad de concentrar la orina.
Sistema Renina Angiotensina Aldosterona
Esta acción reguladora sobre el medio interno no solo implica modificaciones del volumen de agua corporal y de los
principales iones Na+, K+ y Cl
-
; si no que también es capaz de modificar el valor de la presión arterial.
Frente a un estímulo el sistema yuxtaglomerular responde con la liberación de renina la cual actúa sobre el
angiotensinógeno. Se forma la angiotensina I que gracias a la enzima convertidora de angiotensina se forma
angiotensina II. Esta última es una sustancia es un vasoconstrictor muy importante la cual estimula la zona glomerular de
la corteza suprarrenal para liberar aldosterona.
La aldosterona estimula la reabsorción de sodio, cloro y agua a nivel renal, lo que conduce a un aumento del volumen
del líquido extracelular lo cual genera un aumento de la presión arterial.
Si bien influye sobre la PA su función principal es el control electrolítico del medio interno.
La ANGIOTENSINA II es un potente vasoconstrictor y aumenta la absorción de Na y por lo tanto disminuye el radio, hay
mayor resistencia periférica y aumenta la PA.
Micción
Proceso por el cual se excreta la orina. Primero la vejiga se llena progresivamente hasta que la tensión de sus paredes
aumenta por encima de un umbral desencadenando un segundo paso llamado reflejo miccional, que vacía la vejiga o
provoca el deseo de orinar. Centros presentes en la corteza cerebral pueden inhibirlo o facilitarlo.
Se realiza por la contracción abdominal, entrada extra de orina en el cuello de la vejiga y la uretra posterior, estimulo de
receptores de distensión, se excita el reflejo miccional y se inhibe el esfínter uretral externo.

Resumen Renal Pablo. C Página 15
Regulación: una vez que el reflejo miccional es suficientemente poderoso provoca otro reflejo a través de los
nervios pudendos al esfínter externo para inhibirlo. Si la inhibición es más potente en el encéfalo que las señales
constrictoras voluntarias del esfínter externo se produce la micción.
Regulación de la excreción y concentración de potasio en el líquido extracelular
La concentración de potasio en el líquido extracelular está regulada en unos 4,2 mEq/l, y raramente aumenta o
disminuye más de ±0,3 mEq/l. Este control preciso es necesario porque muchas funciones celulares son muy sensibles a
los cambios en la concentración del potasio en el líquido extracelular. Por ejemplo, un aumento de la concentración de
potasio de solo 3-4 mEq/l puede provocar arritmias cardíacas, y concentraciones mayores una parada cardíaca o una
fibrilación.
El mantenimiento del equilibrio entre la captación y el gasto del potasio depende sobre todo de la excreción renal
porque la excreción fecal es solo del 5-10% de la ingestión de potasio. De este modo, el mantenimiento de un equilibrio
normal del potasio exige que los riñones ajusten la excreción de potasio con rapidez y precisión como respuesta a
amplias variaciones en su ingestión, como es también cierto en la mayoría de los otros electrólitos.
Regulación de la distribución interna del potasio
Tras la ingestión de una comida normal, la concentración de potasio en el líquido extracelular aumentaría hasta un valor
mortal si el potasio ingerido no se moviera rápidamente hacia el interior de las células. Afortunadamente, la mayor
parte del potasio ingerido pasa rápidamente al interior de las células hasta que los riñones pueden eliminar el exceso.
Este documento contiene más páginas...
Descargar Completo
Resumen Renal.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.