Fiorella Luchini
RESUMEN EMBRIOLOGÍA PRIMER PARCIAL
Gametas y fecundación
Gametas:
• Óvulo: gameta femenina madura con un número haploide de cromosomas (1n:23). Las células
foliculares que rodean al ovocito secundario se denominan corona radiada. Las microvellosidades
que se encuentran en la superficie del ovocito son la zona pelúcida que presenta glucoproteínas
como son la ZP1, ZP2 (encargada del control de la poliespermia) y ZP3 (quien interacciona con el
espermatozoide).
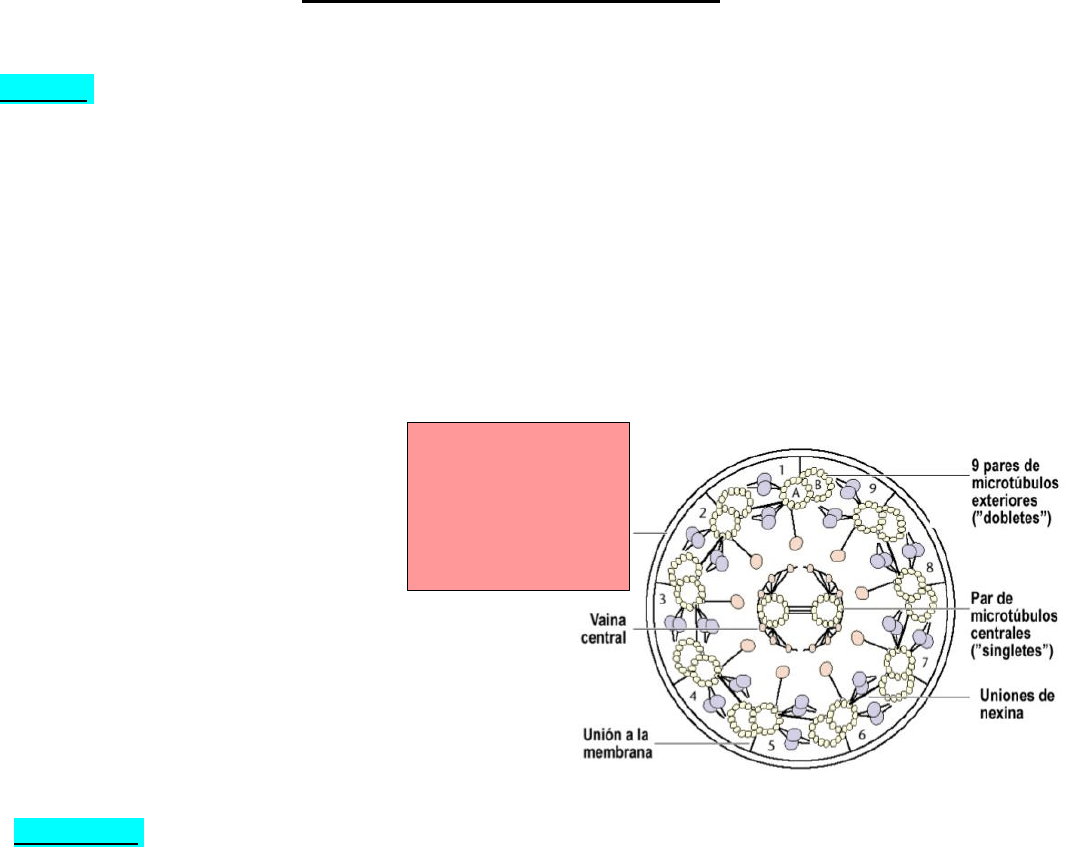
• Espermatozoide: célula altamente especializada que presenta un número haploide de
cromosomas y es la gameta masculina; se genera en los túbulos seminíferos testiculares y hay más
de 300 millones de espermatozoides por eyaculación. Formado por una cabeza con acrosoma y
núcleo, un cuello y una cola que presenta una parte intermedia, una principal y una terminal, la
cola está formada por axonema.
Los espermatozoides deben sufrir
una maduración que se produce en
el epidídimo y son cambios
químicos, morfológicos y
funcionales (como movilidad
anterógrada y cambios en la fluidez de la
membrana) y una capacitación que se da en el
útero y es una incubación de aprox 7hs en donde
se produce una hiperactivación del
espermatozoide, es decir, adquiere mayor
movilidad.
Fecundación: proceso mediante el cual se unen o fusionan el ovulo y el espermatozoide, para formar
el cigoto o célula huevo.
- Transporte de gametas: el ovocito II es captado por la trompa de Falopio que lo transporta
hacia la cavidad uterina. Por otra parte, los espermatozoides avanzan en sentido contrario
desde la vagina hasta el tercio externo de la trompa de Falopio. Solo 200 espermatozoides
llegan a nivel tubárico debido a las barreras mecánicas (como el moco) o el pH acido de la
vagina. El desplazamiento de las gametas está favorecido por las contracciones rítmicas del
aparato reproductor femenino y el movimiento ciliar tubárico.
El ovocito puede ser fecundado 24 hs después de la ovulación, los espermatozoides
sobreviven en el tracto femenino entre 24 y 72 hs.
- Penetración de la Corona Radiada: los espermatozoides tratan de llegar al ovocito, formando
un túnel por acción de la hialuronidasa y por la fuerza mecánica de la hiperactivación en la
corona radiada.
- Reconocimiento y adhesión: existe solo adhesión de la Membrana Pelúcida.
- Reacción acrosómica: el espermatozoide que toma contacto con la Membrana Pelúcida
determina la interacción entre la membrana del espermatozoide y la ZP3.
Estructura interna
axial de los cilios y
flagelos que ayuda
en la movilidad de
la célula.

Fiorella Luchini
Da lugar a múltiples puntos de fusión entre la membrana plasmática del espermatozoide y la
membrana externa del acrosoma. Esta forma poros por los que salen las enzimas acrosómicas
con lisis de ambas membranas, queda expuesta la membrana acrosómica interna.
- Denudación: se produce la separación de las células foliculares por acción de la hialuronidasa
y se desprende la corona radiante.
- Penetración de la membrana pelúcida
- Fusión: solo un espermatozoide establece intimo contacto con la membrana plasmática del
ovocito ll.
- Bloqueo de la Poliespermia: se neutraliza la entrada de otros espermatozoides,
desencadenándose cambios en el ovocito.
Reacción cortical: inmovilización y expulsión de los espermatozoides atrapados en la
Membrana Pelúcida.
Transformación de la estructura de la membrana plasmática: pierde la capacidad de
fusionarse con las membranas plasmáticas de nuevos espermatozoides.
- Reinicio de la Meiosis ll: la presencia del espermatozoide determina el reinicio de la meiosis
ll, con la formación del Segundo Cuerpo Polar.
- Formación de Pronúcleos femenino y masculino: Se tornan esféricos, se dirigen a la región
central del cigoto y duplican su material cromosómico, preparándose para la primera división
de la segmentación.
- Singamia: los pronúcleos se acercan y pierden sus cariotecas.
- Anfimixis: los cromosomas duplicados vuelven a condensarse y se ubican en el plano
ecuatorial (metafase). Es el fin de la fecundación.
Ciclo ovárico y endometrial
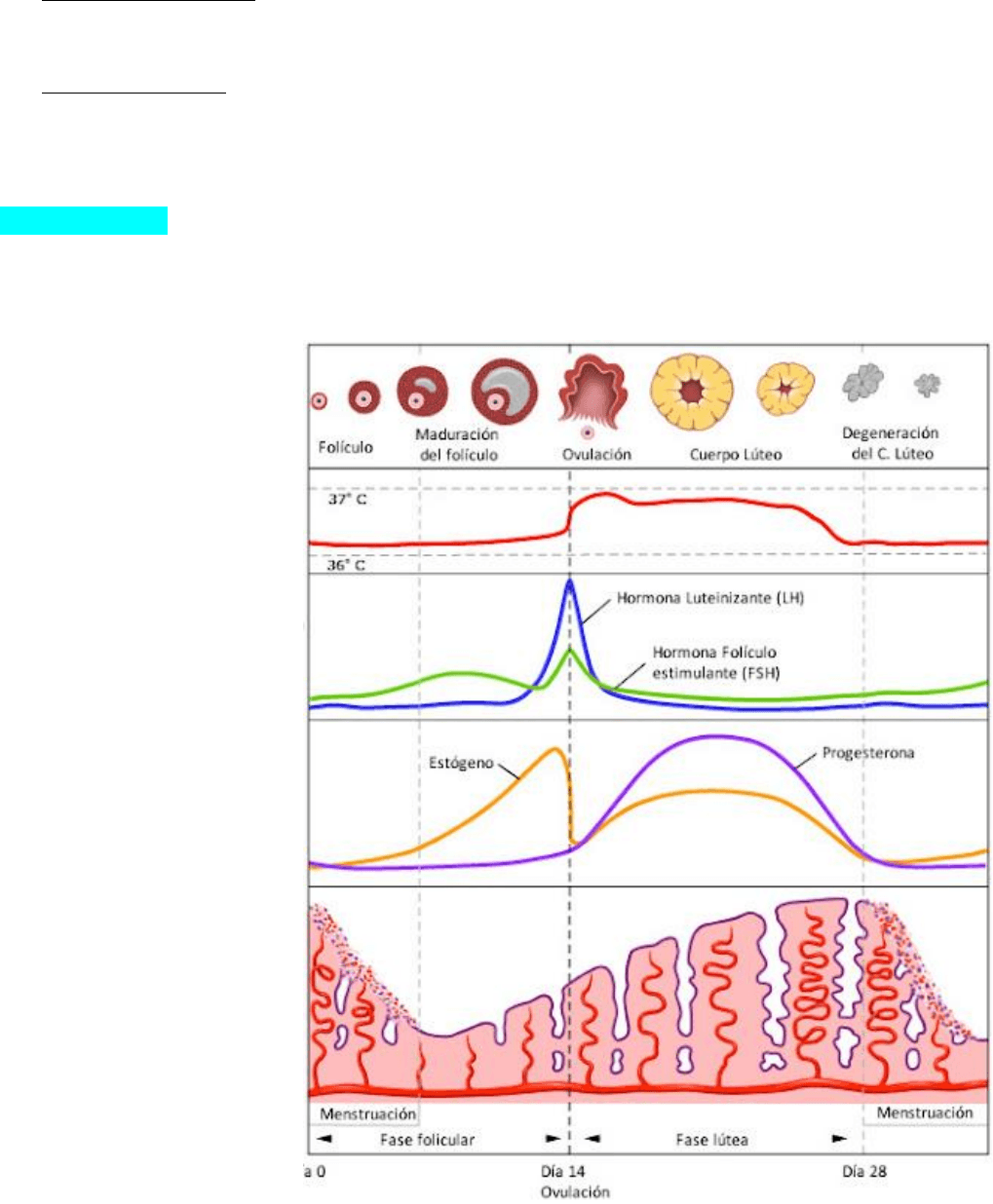
Ciclo Ovárico
Son los cambios cíclicos que ocurren en el ovario, por acción de las hormonas hipofisarias
gonadotróficas: folículo estimulante (FSH) y luteinizante (LH), con una duración de aprox 28 días.
Etapas:
1. Fase Folicular: crecimiento y desarrollo de los folículos ováricos qué por acción de la FSH,
promueve el crecimiento de varios folículos, pero solo uno se desarrolla a maduro y se rompe en
la superficie del ovario, por lo que se expulsa el ovocito.
Folículo primordial: está constituido por un ovocito primario, de aproximadamente 25 m de
diámetro, rodeado por una capa de células foliculares planas.
Folículo primario: las células foliculares se tornan cúbicas o cilíndricas y proliferan formando varias
capas alrededor del ovocito. Las células foliculares se denominan células de la granulosa. Aparece
la zona pelúcida, membrana glucoproteica ubicada entre el ovocito y las células foliculares. Las
células del estroma se organizan alrededor del folículo y se distribuyen en una capa concéntrica, la
teca folicular.
Folículo secundario (antral): Se desarrollan varias capas de células foliculares, entre las cuales
aparecen espacios intercelulares con líquido folicular. Al coale5ser estos espacios, se forma una
cavidad llena de líquido, denominada antro folicular. La capa única de células granulosas que
rodea de manera inmediata al ovocito, se denomina corona radiada. Las células de la granulosa se
redistribuyen, un pequeño grupo que rodean al ovocito se proyecta desde la pared hacia el antro y
de denomina cúmulo ovóforo (disco prolífero). La teca folicular se diferencia en una teca interna y
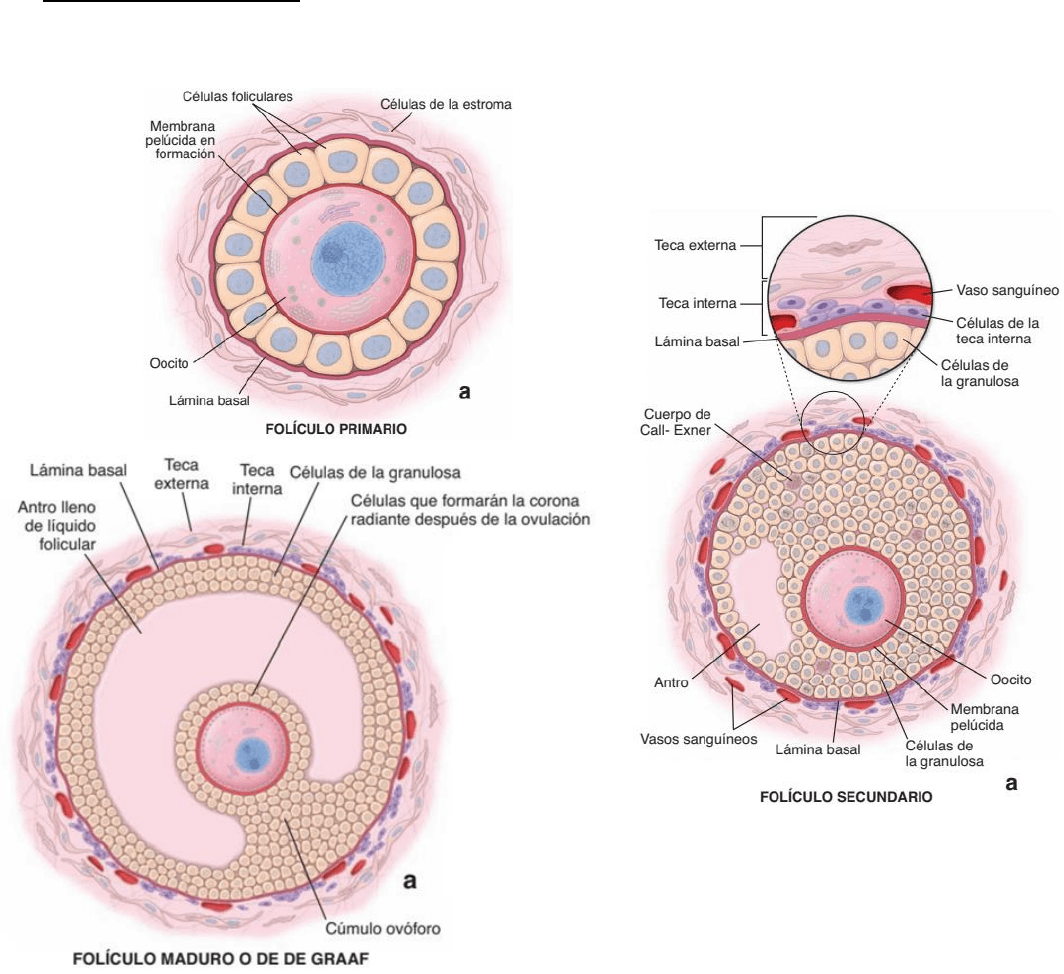
Fiorella Luchini
una teca externa. Las células del estroma de la teca interna se diferencian a células poliédricas
epitelioides, y adquieren una rica vascularización. Esta capa presenta actividad endocrina, ya que,
junto con las células de la granulosa, sintetizan estrógenos. La teca externa, mantiene su
naturaleza de tejido conectivo fibroso (haces de fibras colágenas) y células musculares lisas.
Folículo maduro (de Graff): el ovocito fluye libremente en el líquido folicular, rodeado por la zona
pelúcida y la corona radiada. Está listo para ser fecundado.
2. Ovulación/ Ovocitación: se da aproximadamente a la mitad del ciclo (14dias). Por influencia de la
FSH y FH, el folículo ovárico sufre un crecimiento súbito, produciendo un abultamiento en la
superficie del ovario. Una marcada liberación de LH induce a la ovulación. Se libera es un ovocito
II, que se transforma en ovulo solo si es fecundado.
3. Fase Luteínica: formación y evolución del cuerpo amarillo/lúteo. Después de la ovulación, el
folículo se transforma en cuerpo lúteo, debido a que se colapsan las paredes, la capa de células
granulosas es invadida por tejido conectivo vascularizado. Las células de la granulosa y las de la
teca interna crece, se transforman en poliédricas y en su citoplasma aparece un pigmento
amarillo: la luteína, esta transformación se denomina luteinización y es causada por la LH, cuya
Fiorella Luchini
producción de hormonas esteroides está dada por la síntesis y secreción de progesterona en
mayor cantidad y de estrógenos en menor cantidad.
Ovocito No fecundado: se forma el Cuerpo Lúteo de la Menstruación, que alcanza su máximo
desarrollo a los 9 días después y luego involuciona, se transforma en Cuerpo Albicans, luego es
invadido por tejido conectivo fibroso. Desaparece a los 14 días posteriores a la ovulación.
Ovocito Fecundado: se habla de Cuerpo Lúteo del Embarazo/ Gestación y comienza el embarazo,
produce hormonas esteroideas. El cuerpo lúteo del embarazo se mantiene gracias a la acción de la
hormona Gonadotrofina Coriónica Humana (HCG), secretada por el Sincitiotrofoblasto del corion.
Ciclo endometrial
Cambios cíclicos que ocurren en el endometrio por acción de las hormonas ováricas estrógenos y
progesterona. Tiene una duración de 28 días. Etapas:
1. Fase menstrual: es
cuando se produce la
hemorragia uterina, que
ocurre cuando no se
produce la fecundación
del ovocito. El cuerpo
lúteo pierde su función
por lo que se reducen los
noveles de estrógenos y
progesterona. Se da
desde 1-4 días.
2. Fase proliferativa:
cambios que sufre la
mucosa endometrial,
comenzando a proliferar
luego de la perdida de la
capa funcional durante la
fase menstrual.
3. Fase secretora: cambios
secretores en glándulas
endometriales, durante
esta fase crece aún más
el espesor del
endometrio hasta 6-
7mm, por acción de la
progesterona y
estrógenos, secretados
por el cuerpo lúteo.
-Fecundación: se produce la segmentación del cigoto y formación de blastocisto.
-No fecundación: el cuerpo lúteo degenera y se transforma en cuerpo albicans, ocurre la
menstruación.
Fiorella Luchini
Primera semana del desarrollo
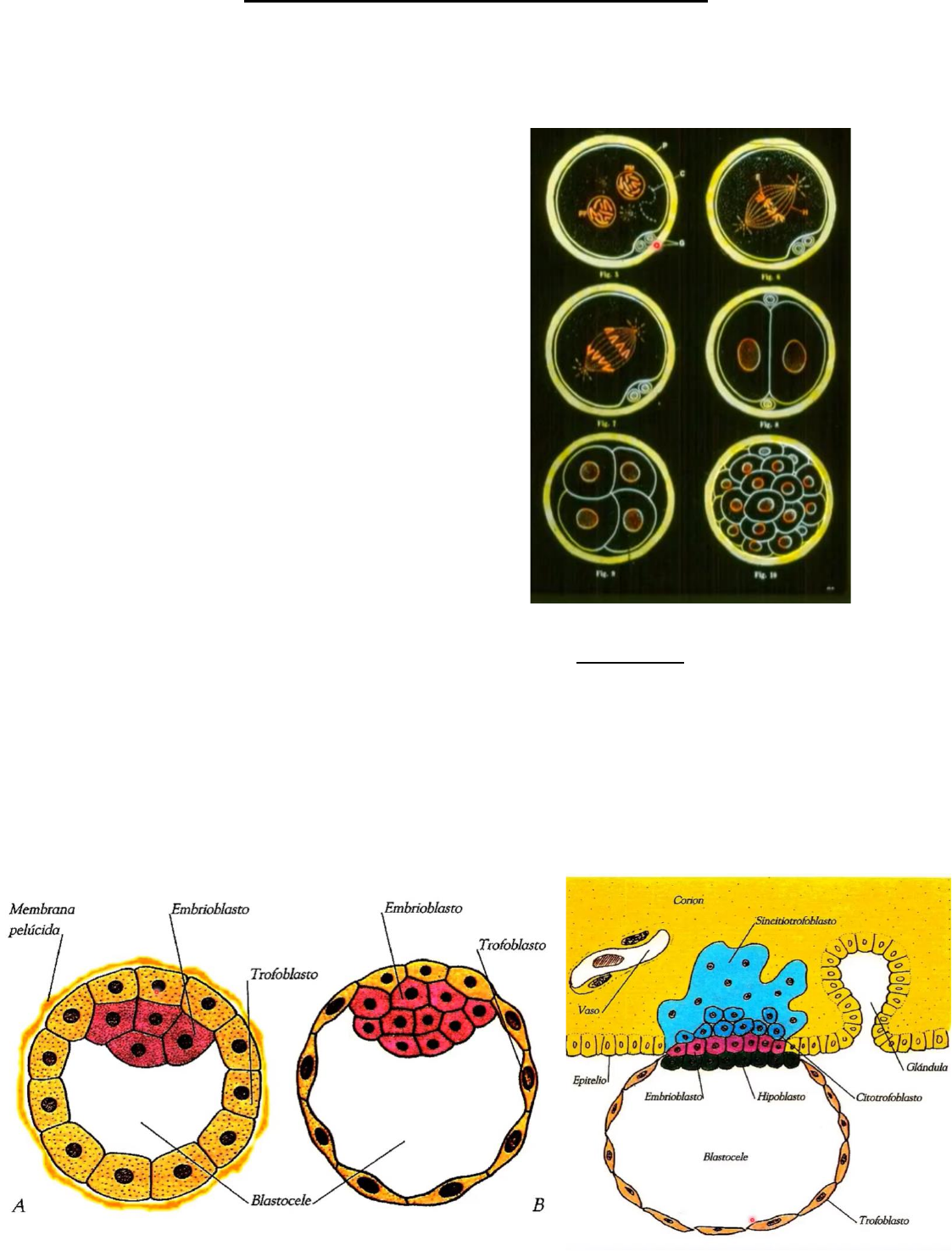
Segmentación: sucesión de divisiones mitóticas que se producen con autoduplicación del
ADN entre una y otra. Origina células hijas denominadas blastómeras. Las características
son:
- Total u holoblástica
- Igual o subigual
- Asincrónica
1° día -> 2 blastómeras;
2° día -> 4 blastómeras;
2 ½° -> 8 blastómeras (compactación);
3° día -> 12 a 16 blastómeras (mórula).
Las blastómeras externas forman el macizo
celular externo, mientras que las internas
forman el macizo celular interno.
4° día: la mórula cae en la cavidad uterina y se
penetra líquido en ella dejando asi un
embrión hueco con una cabidad central ->
blastocele. El MCE origina el trofoblasto, y el
MCI el embrioblasto. A este embrión se lo
conoce como blastocisto.
6° día: se pierde la membrana pelúsida y se produce la implantación. Entre el 6° y 7° día a
partir de la fecundación, el Blastocisto, se implanta en la mucosa uterina (en el endometrio
secretor) por el polo ejmbrionario, el lugar habitual es el 1/3 superior de la pared posterior
del útero.
El trofoblasto comienza a dividirse en dos capas: citotrofoblasto y sincitiotrofoblasto.
Aparece la primera hoja germinativa por delaminación -> el hipoblasto.
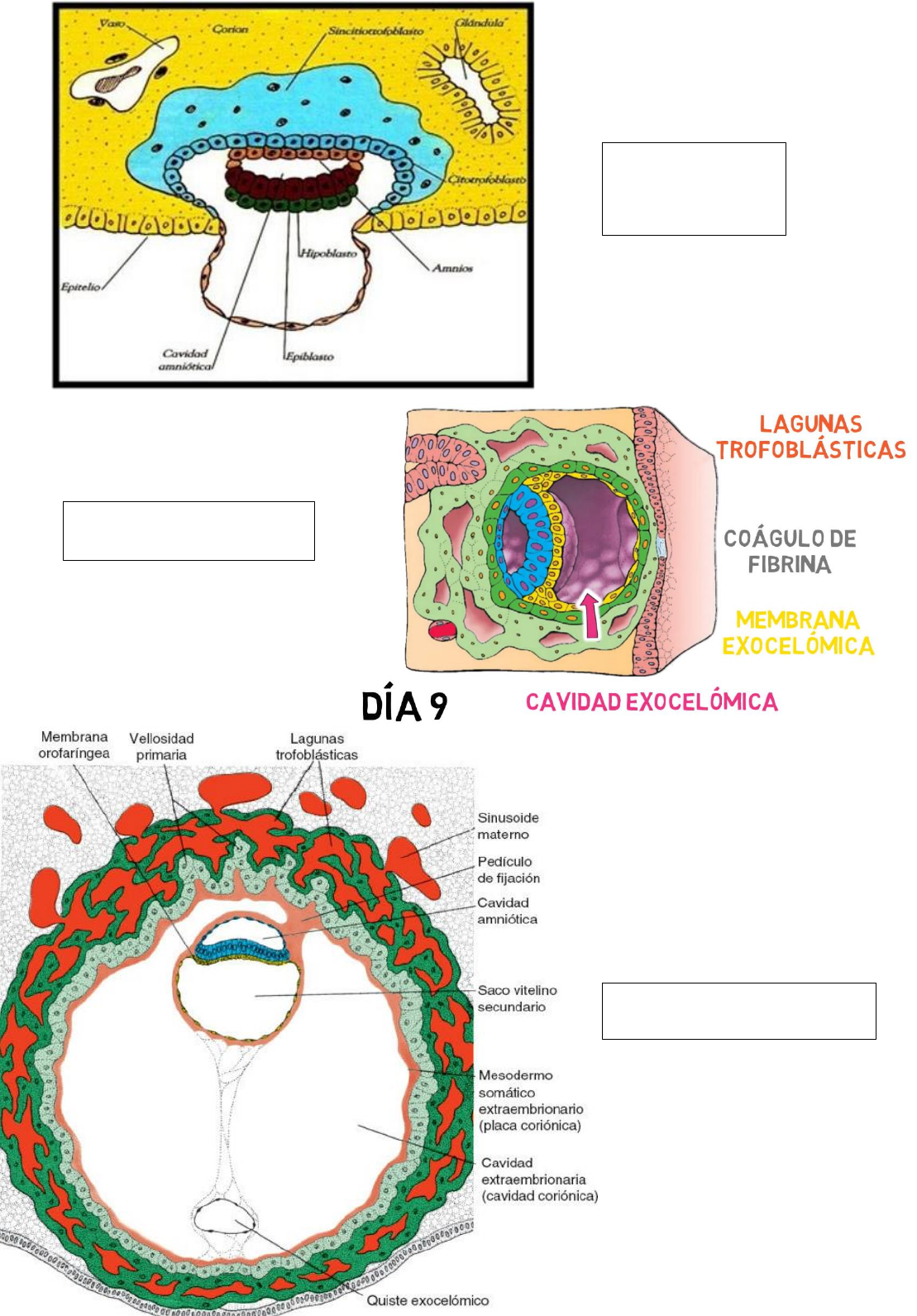
Al día 9 el embrión está completamente ubicado dentro de la mucosa uterina.

Fiorella Luchini
Segunda semana del desarrollo
8° día: el embrioblasto se dividió en dos capas: hipoblasto y epiblasto, originando así el embrión
biliaminar. En el epiblasto aparece una pequeña cavidad, que aumenta de tamaño y se convierte
en la cavidad amniótica.
El epiblasto contiene células que forman el embrión y tejidos extraembrionarios.
Al 9° día, el embrión está ubicado completamente dentro de la mucosa uterina, en el orificio de
entrada aparece el tapón de fibrina. En el sincitiotrofoblasto aparecen lagunas que erosionan
vasos y glándulas por lo que contienen sangre y secreciones glandulares, del endometrio materno.
Los nutrientes pasan por difusión hacia el embrión (nutrición histotrófica).
En la superficie interna del citotrofoblasto comienza a formarse una membrana delgada ->
membrana de heusser. Esta membrana junto con el hipoblasto genera el recubrimiento de la
cavidad exocelómica/saco vitelino primitivo.
Aparece un tejido conectivo laxo, entre el saco vitelino primitivo, la cavidad amniótica y
citotrofoblasto, denominado mesodermo extraembrionario.
Al día 11, se da la interconexión de lagunas trofoblásticas (red lacunar): es el comienzo de la
circulación uteroplacentaria primitiva, donde la sangre materna comienza a circular lentamente
por la red.
En el mesodermo extraembrionario aparecen espacios lacunares, que al coalescer forman al
celoma extraembrionario.
El hipoblasto avanza sobre la membrana de Heusser y comienza a cubrir el exoceloma (saco
vitelino primitivo).
Al día 13, se origina el saco vitelino definitivo. Las células del citotrofoblasto proliferan localmente
y penetran en el sincitiotrofoblasto donde forman columnas celulares rodeadas de sincitio. Las
columnas con cobertura sincitial reciben el nombre de vellosidades primarias
Al 15 día, el celoma extraembrionario se halla tapizado por mesodermo extraembrionario, que se
divide en dos hojas: somatopleura (dorsal) y esplacnopleura (ventral). Se forma el Corion,
constituido por: mesodermo extraembrionario (somatopleura), citotrofoblasto y
sincitiotrofoblasto.
El saco vitelino y la cavidad amniótica quedan suspendidos por el pedículo de fijación, este, con el
desarrollo de los vasos sanguíneos se convierte en el cordón umbilical.
Disco bilaminar:
La placa precordal nos informa la
localización de la boca, y es un
elemento organizador de la
cabeza del embrión. Son células
hipoblásticas cilíndricas.
Fiorella Luchini
Embrión de
Hertig y Rock
Embrión de 9 días
Embrión de 13 días
Fiorella Luchini
Tercera semana del desarrollo
Gastrulación: conjunto de movimientos celulares que llevaran a la formación de tres capas
germinales primitivas del embrión a partir de las células del epiblasto, en la superficie dorsal
del disco embrionario bilaminar.
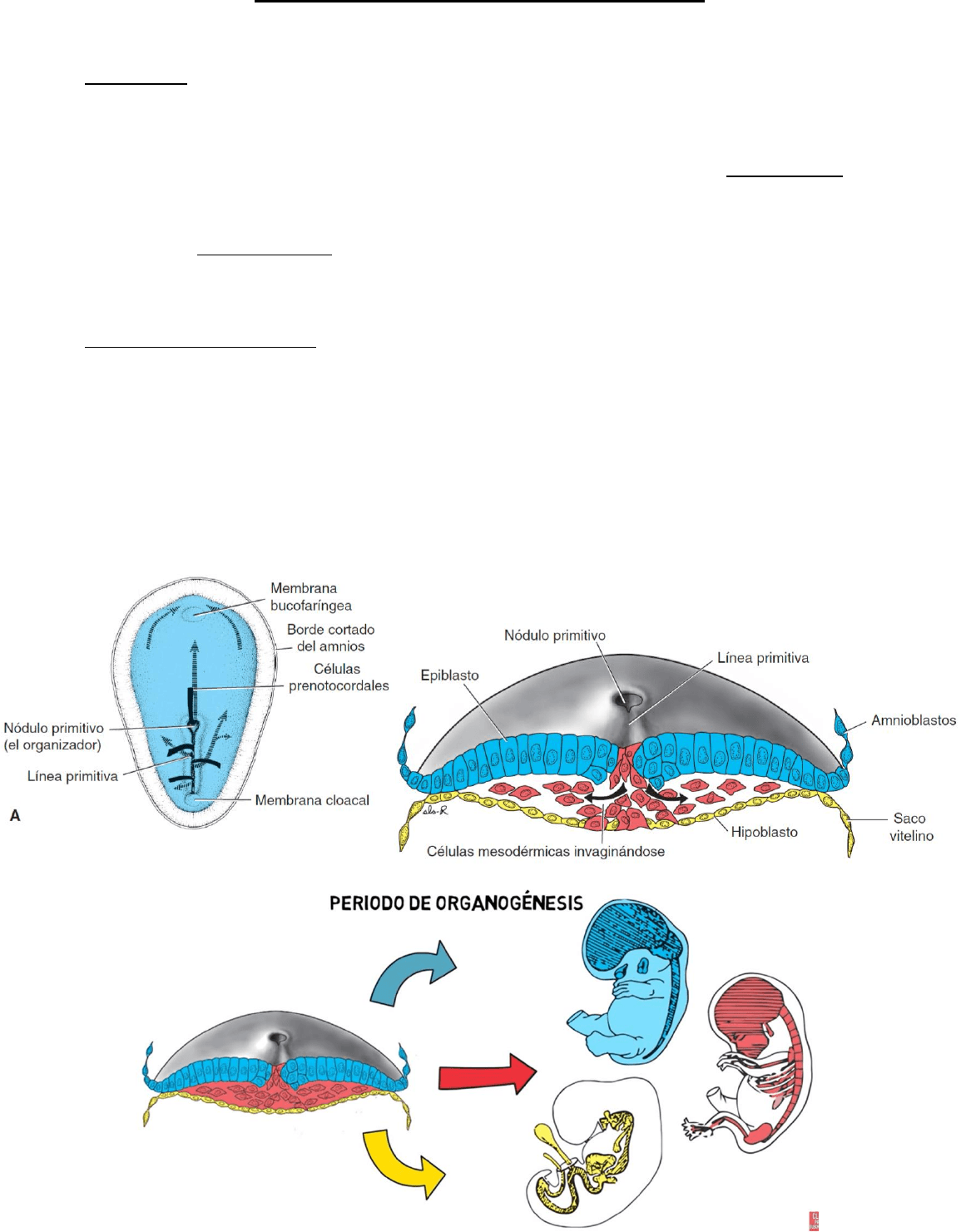
La primera evidencia que marca el inicio de la gastrulación es la aparición de la línea primitiva,
en la región dorsal y caudal del disco embrionario. Se da en este lugar porque se comienzan a
formar los ejes corporales y por la relación directa con el eje céfalo-caudal. El extremo cefálico
de la línea (el nódulo primitivo) consta de un área ligeramente elevada que rodea a la fosa
primitiva.
Se originan las 3 hojas embrionarias: ectodermo, mesodermo y endodermo, convirtiéndose el
disco embrionario trilaminar.
• Las células del epiblasto que se invaginan en la fosita del nodulo, desplazan al hipoblasto
y lo reemplazan por endodermo embrionario.
• Las células epiblasticas al invaginarse adquiere características mesenquimáticas, así se
forma el mesodermo embrionario.
• Las células del epiblasto que no se han invaginado persisten en la superficie dorsal
formando el ectodermo embrionario.
Fiorella Luchini
Formación de la notocorda: Las
células prenotocordales, que se
invaginan en el nódulo primitivo
avanzan cranealmente en la línea
media hasta llegar a la placa
precordal. Después se intercalan
en el hipoblasto, de manera que
por un corto plazo la línea media
del embrión consta de dos capas
celulares que constituyen la placa
notocordal. Al ser el hipoblasto
reemplazado por las células
endodérmicas que entran en la
línea primitiva, las de la placa
notocordal proliferan y se separan
del endodermo. Forman luego un
cordón sólido de células, la
notocorda definitiva que pasa por
debajo del tubo neural y es la
base para inducir el esqueleto
axial.
Las células prenotocordal migran
a través de la línea primitiva, se intercalan en el endodermo para producir la placa notocordal
y finalmente se desprenden del endodermo para formar la notocorda definitiva. Dado que
estos procesos ocurren en una secuencia craneal a caudal, los primeros fragmentos de la
notocorda definitiva se establecen primero en la región de la cabeza.
La membrana cloacal se forma en el extremo caudal del disco embrionario. Esta membrana,
de estructura similar a la membrana bucofaríngea, consta de células ectodérmicas
fuertemente adheridas y de células endodérmicas sin mesodermo entre ellas. Cuando aparece
la membrana cloacal, la pared posterior del saco vitelino produce un divertículo pequeño que
se extiende hasta el pedículo de fijación. Este divertículo, divertículo alantoentérico o
alantoides, aparece hacia el día 16 de desarrollo
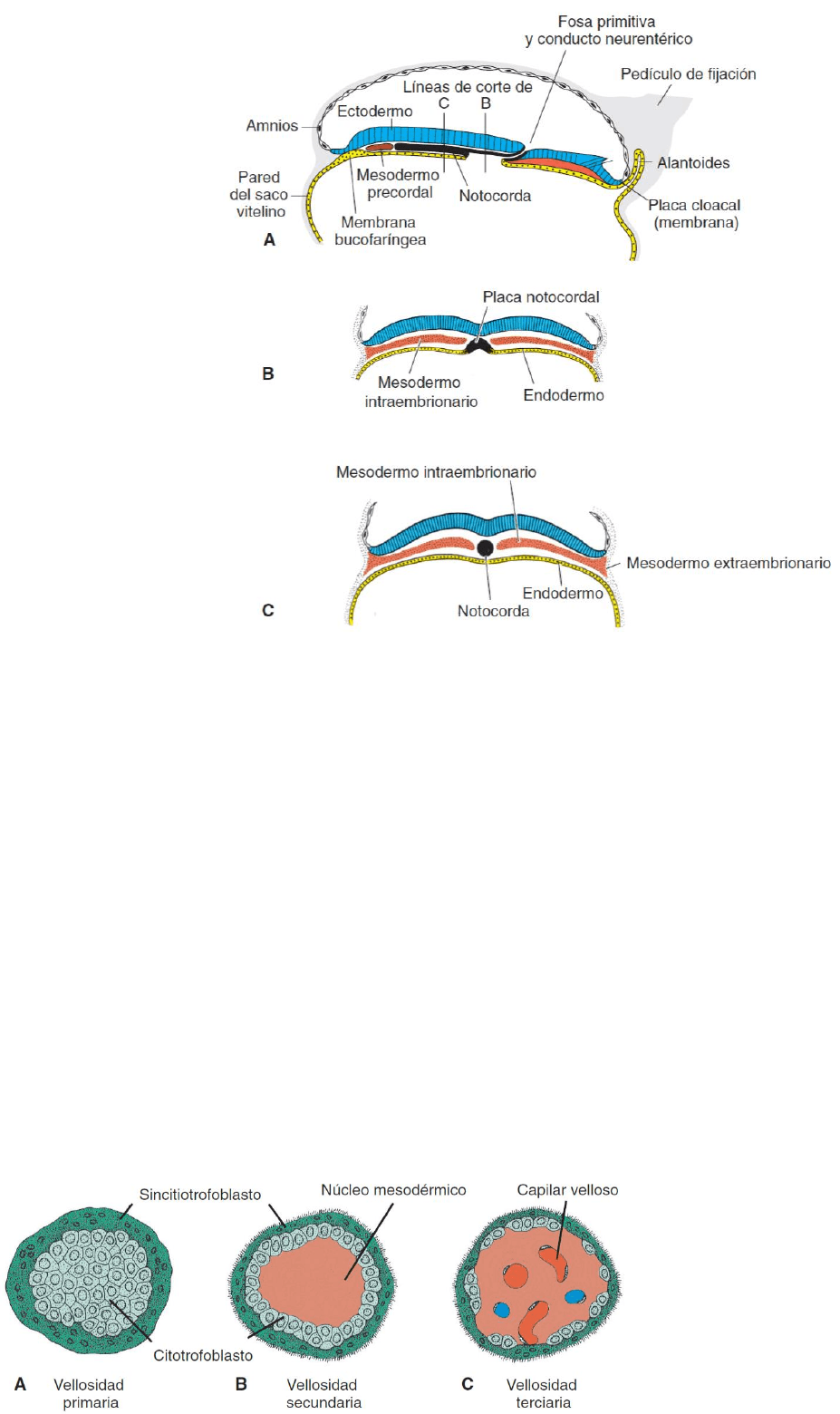
Desarrollo del trofoblasto. Al inicio de la tercera semana el trofoblasto se caracteriza por las
vellosidades primarias que constan de un núcleo citotrofoblástico recubierta por una capa
sincitial. Durante las fases posteriores del desarrollo las células mesodérmicas penetran en el
núcleo de las vellosidades primarias creciendo hacia la decidua (vellosidad secundaria). Al final
de la tercera semana, en el núcleo de la vellosidad, algunas células mesodérmicas empiezan a
diferenciarse convirtiéndose en hepatocitos y en pequeños vasos sanguíneos, lo que da origen
al sistema capilar velloso. Ahora a la vellosidad se conoce como vellosidad terciaria.
El endodermo queda
en contacto con el
ectodermo en la
membrana
bucofaríngea y en la
membrana cloacal.
Fiorella Luchini
Ejes corporales: El establecimiento de los ejes corporales, anteriorposterior (craneocaudal),
dorsoventral y derecho-izquierdo se lleva a cabo al inicio de la embriogénesis. Migran a la futura
región craneal las células que darán origen al endodermo visceral anterior (EVA) en el extremo
craneal de la capa endodérmica del disco laminar. Durante esta etapa del disco bilaminar las células
en el endodermo EVA expresan genes esenciales para formar el encéfalo, entre ellos los factores de
transcripción OTX2, LIM1 y HESX1, lo mismo que los factores secretados cerberus y lefty1
(miembros de la familia TGF-β). Estos inhiben la actividad nodal, con lo cual establecen el extremo
craneal del embrión. La ausencia de cerberus y de lefty1 en el extremo caudal del embrión permite
que prosiga la expresión nodal, señal que establece y mantiene la línea primitiva. Una vez formada
la línea, el gen NODAL regula varios genes que forman el mesodermo dorsal y ventral, así como las
estructuras de la cabeza y de la cola.
Otro miembro de la familia TGF-β, la proteína morfogénica ósea 4 (BMP4), se secreta en todo el
disco embrionario. En presencia de esta proteína y del factor de crecimiento de los fibroblastos, el
mesodermo se dirige a la parte ventral para contribuir a la formación de riñones (mesodermo
intermedio), de la sangre y el mesodermo de la pared corporal (mesodermo de la placa lateral).
Los genes de la cordina junto con nogina y folistatina antagonizan la actividad de BMP-4. Así que, el
mesodermo se desplaza dorsalmente hacia la notocorda, los somitas y los somitómeros. Más tarde,
estos tres genes se expresan en la notocorda y son importantes en la inducción neural dentro de la
región craneal.
Como ya se mencionó, el gen Nodal participa en la iniciación y mantenimiento de la línea primitiva.
De modo análogo, HNF-3β conserva el nódulo y más tarde induce la especificidad regional en las
áreas del prosencéfalo y del mesencéfalo.
La regulación de la formación del mesodermo dorsal en las regiones central y caudal está
controlada por el gen BRACHYURY(T) expresado en el nódulo, en las células precursoras de la
notocorda y en ésta. El gen es esencial para la migración celular a través de la línea primitiva
También la lateralidad (simetría derecha-izquierda) se establece al inicio del desarrollo. En
condiciones normales muchos órganos muestran asimetrías, entre ellos: corazón, pulmones,
intestino, bazo, estómago, hígado. La posición de estos órganos y sus asimetrías dependen de
multitud de moléculas y genes señalizadores.
Cuando aparece la línea primitiva, el factor de crecimiento de los fibroblastos (FGF8) es secretado
por las células en el nódulo y en la línea primitiva. Este factor induce la expresión de gen Nodal.
Luego la expresión queda restringida al lado izquierdo del embrión al acumularse allí serotonina (5-
HT). Las elevadas concentraciones de serotonina en ese lado activa la expresión del factor de
transcripción MAD3 que limita la expresión del gen Nodal a la cara izquierda del nódulo primitivo.
Los genes mediales como SONIC HEDGEHOG (SHH), LEFTY 1 y ZIC3 ayudan a establecer la línea
medial pero al mismo tiempo impiden que la expresión del gen Nodal llegue al lado derecho. En el
mesodermo de la placa lateral izquierda la proteína Nodal inicia una cascada de señales que incluye
LEFTY2 para regular PITX2, factor de transcripción que es un “gen maestro” con una
homeosecuencia encargada de establecer la lateralidad izquierda. Su expresión se repite en el lado
izquierdo del corazón, del estómago y los primordios intestinales, pues estos órganos adoptan sus
posiciones corporales asimétricas normales.
Los genes que regulan el desarrollo del lado derecho no están tan bien definidos, aunque la
expresión del factor de transcripción SNAIL está restringida al mesodermo de la placa lateral
derecha y probablemente regule los genes efectores que establecen el lado derecho.
Fiorella Luchini
Derivados endodermicos
Derivados: revestimiento epitelial de vías respiratorias, tubo digestivo, vejiga urinaria y uraco;
componentes epiteliales de la glándula tiroides, cavidad timpánica, trompa auditiva, amigdalas,
glándulas paratiroides, hígado y páncreas.
A raíz del crecimiento cefalocaudal y del cierre de los pliegues de la pared lateral del cuerpo, una
porción cada vez más grande de la capa germinal endodérmica se incorpora al cuerpo del embrión
para constituir el tubo intestinal. Éste se divide en tres regiones: intestino anterior, intestino
medio e intestino posterior. El intestino medio se comunica con el saco vitelino mediante un
pedículo ancho llamado saco vitelino. En el extremo cefálico, el intestino anterior está delimitado
temporalmente por una membrana ectoendodérmica llamada membrana bucofaríngea. Esta
membrana separa el estomodeo (cavidad bucal primitiva derivada del ectodermo) de la faringe,
una parte del intestino posterior originada en el endodermo. El intestino posterior también
termina temporalmente en una membrana ectoendodérmica: la membrana cloacal. Ésta separa la
parte superior del conducto anal proveniente del endodermo y la parte inferior llamada
proctodeo, que se forma con una invaginación recubierta de endodermo. La membrana se rompe
en la séptima semana para crear el orificio del ano. Otro resultado importante del crecimiento
cefalocaudal y del plegamiento lateral es la incorporación parcial del alantoides al cuerpo del
embrión, donde forma la cloaca.
Fiorella Luchini
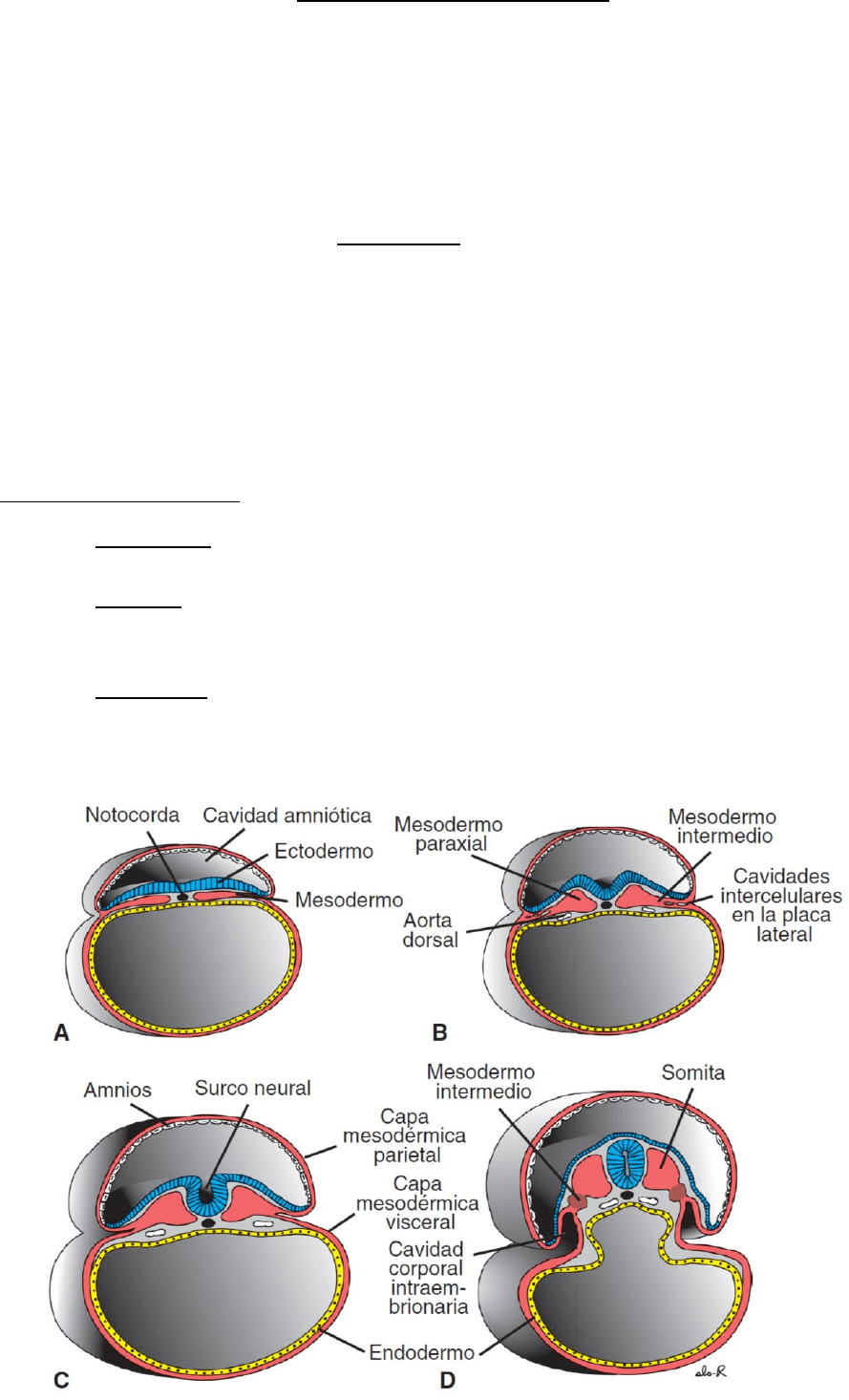
Derivados mesodermicos
En un principio la células de la capa germinal mesodérmica forman a ambos lados de la línea
media una lámina delgada de tejido laxo. Hacia el día 17 las células cercanas a la línea media
proliferan dando origen a una placa engrosada de tejido conocido como mesodermo paraxial.
Más hacia los lados la capa mesodérmica sigue siendo delgada y se llama placa lateral. El
mesodermo intermedio conecta el mesodermo paraxial al mesodermo de placa lateral.
Mesodermo paraxial: al inicio de la tercera semana el mesodermo paraxial empieza a
organizarse en segmentos llamados somitómeros. En la región cefálica, los somitómeros se
convierten en neurómeros en asociación con la segmentación de la placa neural,
contribuyendo al mesénquima en la cabeza; desde la región occipital en sentido caudal, los
somitómeros continúan organizándose hasta convertirse en somitas. El primer par de somitas
aparece en la región occipital del embrión hacia el día 20 del desarrollo, los siguientes
aparecen a un ritmo aproximado de tres pares diarios, hasta que hay de 42 a 44 pares al final
de la quinta semana. Más tarde desaparecen el primer par occipital y los últimos somitas
coccígeos de 5 a 7, mientras los restantes somitas constituyen el esqueleto axial.
Diferenciación de somitas:
- Esclerotoma: deriva de las células de la pared ventral y medial del somita. Origina
las vértebras y las costillas (huesos y cartílagos).
- Miotoma: deriva de las células de los margenes dorsomedial y ventrolateral. Origina
los músculos de la pared y extremidades (hipómero) y los músculos episomíticos
(epímero).
- Dermatoma: se encuentra entre el esclerotoma y el miotoma. De allí deriva la
dermis y el tejido celular subcutáneo de la espalda.
Fiorella Luchini
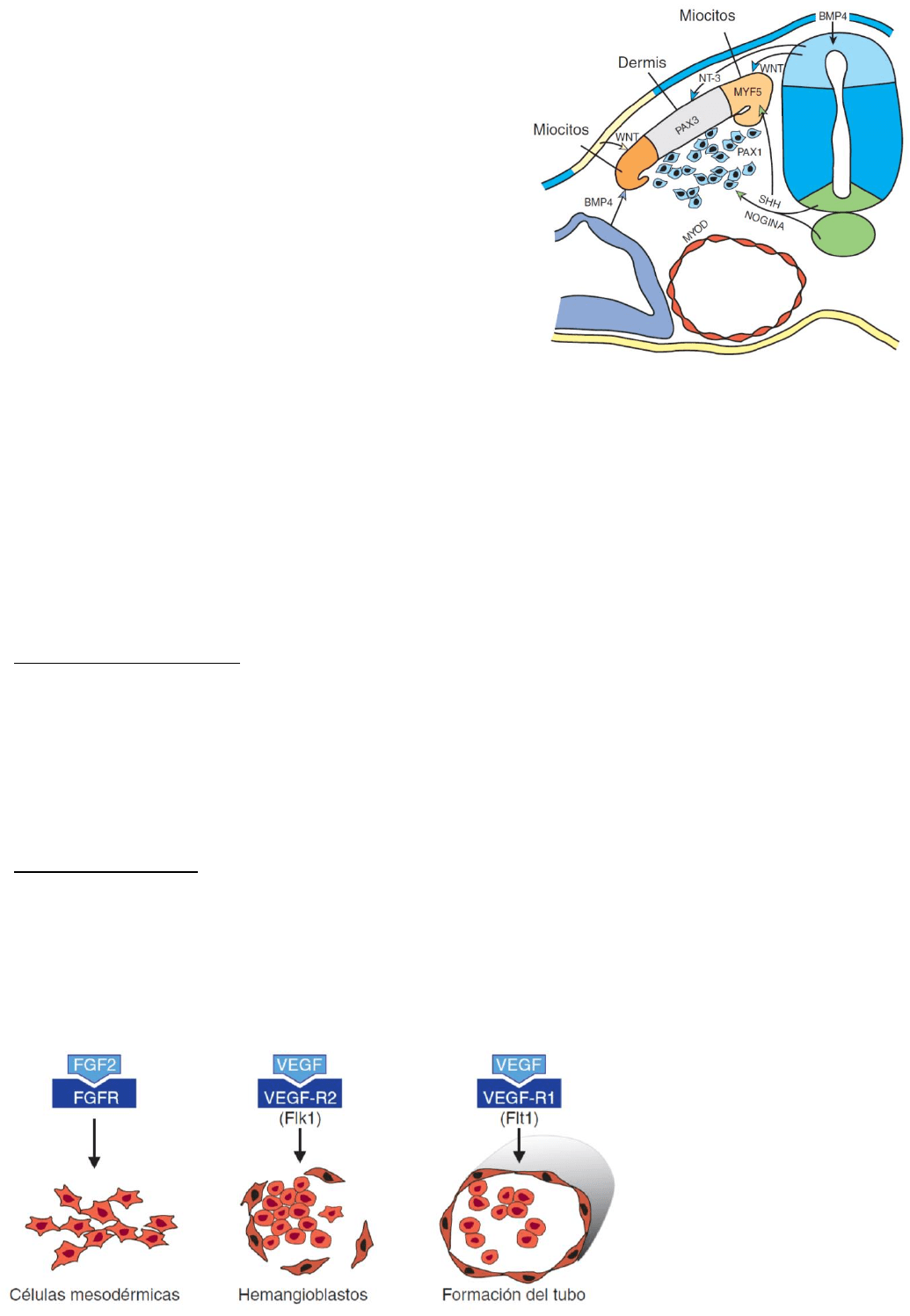
Las proteínas SHH y nogina, secretadas por la notocorda
y la placa basal del tubo neural, hacen que la porción
ventral del somita produzca esclerotoma y exprese PAX1,
que a su vez controla la condrogénesis y la formación de
las vértebras. Las proteínas WNT provenientes del tubo
neural dorsal activan PAX3, que marca el
dermomiotoma. Las proteínas WNT dirigen además la
porción dorsomedial del somita para que se diferencie
en las células precursoras
de la musculatura y exprese el gen MYF5 específico del
músculo. La neurotrofina 3 (NT-3), expresada por el tubo
neural dorsal, dirige la porción dorsomedial del somita
para que se convierta en dermis. Más células precursoras
de la musculatura aparecen en la porción dorsolateral del somita bajo el influjo combinado de las
proteínas activadoras WNT y la proteína morfogenética ósea 4 (BMP4) que juntas activan la
expresión de MyoD.
Mesodermo intermedio: se diferencia en las estructuras urogenitales.
Mesodermo de la placa lateral: se divide en las capas parietal (somática) y visceral (esplácnica)
que revisten la cavidad intraembrionaria y rodean los órganos, respectivamente. Da orígen al
tejido cenctivo y muscular de las visceras, la membrana serosa de la pleura, pericardio y peritoneo,
células sanguíneas y linfáticas, bazo y corteza suprarrenal.
Sangre y vasos sanguíneos: los hematocitos y los vasos sanguíneos también se originan en el
mesodermo. Los vasos sanguíneos lo hacen mediante dos mecanismos: vasculogénesis, los que
provienen de los islotes sanguíneos, y angiogénesis, los que se forman de los vasos ya existentes.
Los primeros islotes aparecen en el mesodermo rodeando la pared del saco vitelino durante la
tercera semana del desarrollo, poco después en el mesodermo de la placa lateral y en otras
regiones. Los islotes proceden de las células mesodérmicas cuya inducción produce
hemangioblastos, un precursor común en la formación de hematocitos y de vasos sanguíneos.
Regulacion molecular: El factor FGF2 induce el desarrollo de los islotes sanguíneos a partir de las
células competentes del mesodermo que constituyen hemangioblastos. Los hemangioblastos son
estimulados para formar hematocitos y vasos sanguíneos mediante el factor de crecimiento
vascular endotelial (VEGF). En el centro de los islotes de sangre, los hemangioblastos producen
células madre hematopoyéticas, precursoras de todos los hematocitos; en cambio, los
hemangioblastos periféricos se diferencian en angioblastos, precursores de los vasos sanguíneos.
Los angioblastos proliferan y con el tiempo se los induce para que generen células endoteliales
mediante el VEGF; luego
ese mismo factor controla
la coalescencia de las
células endoteliales en los
primeros vasos
sanguíneos.

Fiorella Luchini
Derivados ectodermicos
Al inicio de la tercera semana de desarrollo esta capa adopta la forma de un disco más ancho en la
región cefálica que en la caudal. El aspecto de la notocorda y del mesodermo precordal hacen que
el ectodermo suprayacente se engruese para formar la placa neural. Las células de la placa
constituyen el neuroectodermo y esta inducción representa el primer eslabón en el proceso de
neurulación. La inducción de la placa neural se debe a la regulación de señalización del factor de
crecimiento de los fibroblastos (FGF), junto con nogina, cordina y folistatina, la formacion del
prosencéfalo y de la médula espinal requieren ademas del factor WNT3a.
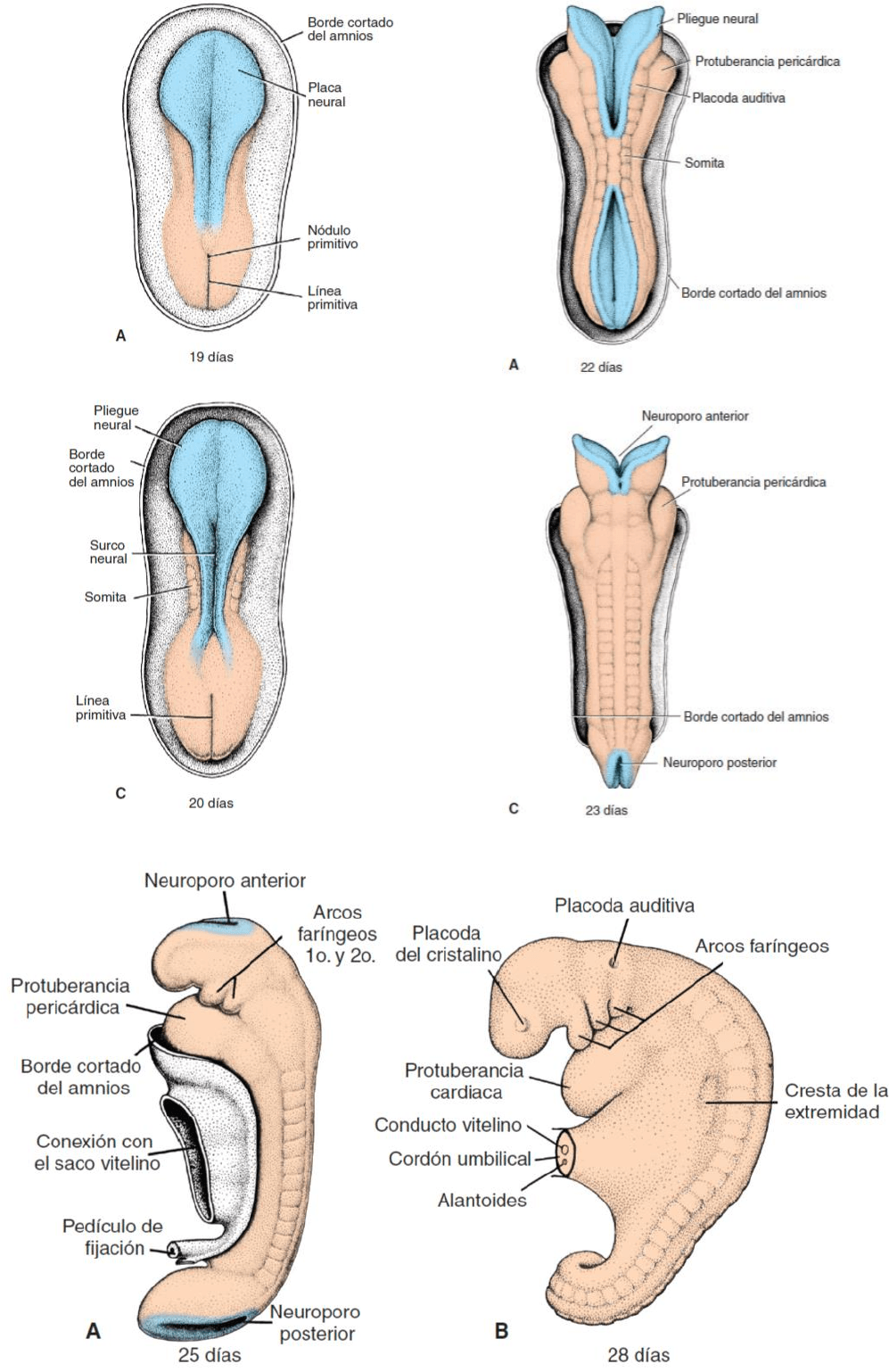
Neurulación: La neurulación es el proceso mediante el cual la placa neural produce el tubo neural.
Uno de los pasos más importantes de este proceso consiste en alargar la placa neural y el eje
corporal con el fenómeno de extensión convergente. Conforme la placa neural va alargándose, sus
bordes laterales se elevan para producir pliegues neurales y la región medial deprimida da origen
al surco neural. En forma gradual los pliegues neurales se acercan uno a otro en la línea media
donde se fusionan. La fusión empieza en la región caudal (quinto somita) y avanza en dirección
craneal y caudal. Así se forma el tubo neural. Mientras la fusión no esté completa, los extremos
cefálico y caudal del tubo neural se comunican con la cavidad amniótica a través de los
neuroporos anterior (craneal) y posterior (caudal), respectivamente. El neuroporo craneal se cierra
aproximadamente en el día 25, en tanto que el neuroporo posterior lo hace en el día 28. En ese
momento la neurulación ha terminado y el sistema nervioso central está representado por una
estructura tubular cerrada con una parte caudal estrecha, la médula espinal, y una parte cefálica
mucho más ancha caracterizada por varias dilataciones: las vesículas encefálicas.
Crestas neurales: son células que sufren una trnsicion epiteliomesenquimática, se originan en el
borde lateral de los pliegues neurales y migran a diferentes partes del mesodermo subyacente. Las
células de la cresta provenientes de la región troncal salen del neuroectodermo después del cierre
del tubo neural y migran por una vía dorsal para producir melanocitos en la piel y en los folículos
pilosos de la piel o por una vía ventral para convertirse en ganglios sensoriales, en neuronas
simpáticas y entéricas, en células de Schwann y en células de la médula suprarrenal. Las células de
la cresta neural también forman pliegues craneales y salen de ellos dejando el tubo neural antes
del cierre en esta región. Estas células contribuyen al esqueleto craneofacial, lo mismo que a las
neuronas de los ganglios craneales, las células gliales, los melanocitos y otros tipos de célula.
En términos generales la capa germinal ectodérmica da origen a los órganos y estructuras que
mantienen contacto con el mundo exterior:
> Sistema nervioso central
> Sistema nervioso periférico
> Epitelio sensorial del oído, de la nariz y ojos
> Epidermis incluidos el cabello y las uñas.
Además da origen a lo siguiente:
> Glándulas subcutáneas
> Glándulas mamarias
> Hipófisis
> Esmalte de los dientes
Fiorella Luchini
Este documento contiene más páginas...
Descargar Completo
RESUMEN EMBRIOLOGÍA PRIMER PARCIAL .pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.