CARACTERÍSTICAS DE LOS SERES VIVOS
Sistemas abiertos, intercambian materia y energía con el medio que los rodea.
● Están formados por una o muchas células.
● Son complejos y organizados.
● Realizan
metabolismo, el cual consiste en el conjunto de todas las reacciones químicas para el aprovechamiento de la
materia y la energía.
● Realizan
homeostasis, es decir, que tienen la capacidad de mantener su medio interno relativamente constante a
pesar de los cambios externos.
● Poseen
irritabilidad, o sea que reaccionan ante estímulos internos o externos.
● Se
adaptan al medio en el que habitan. Esto se relaciona con los procesos evolutivos.
● Se
reproducen, o sea que tienen capacidad de generar descendientes.
●
Crecen y se desarrollan: o sea que aumentan de tamaño debido al aumento en el número de células y además se
desarrollan, es decir se especializan y diferencian.
●
Autopoyesis: Generan componentes propios a partir de los componentes que lo formaron (ADN-PROTEINA-ADN).
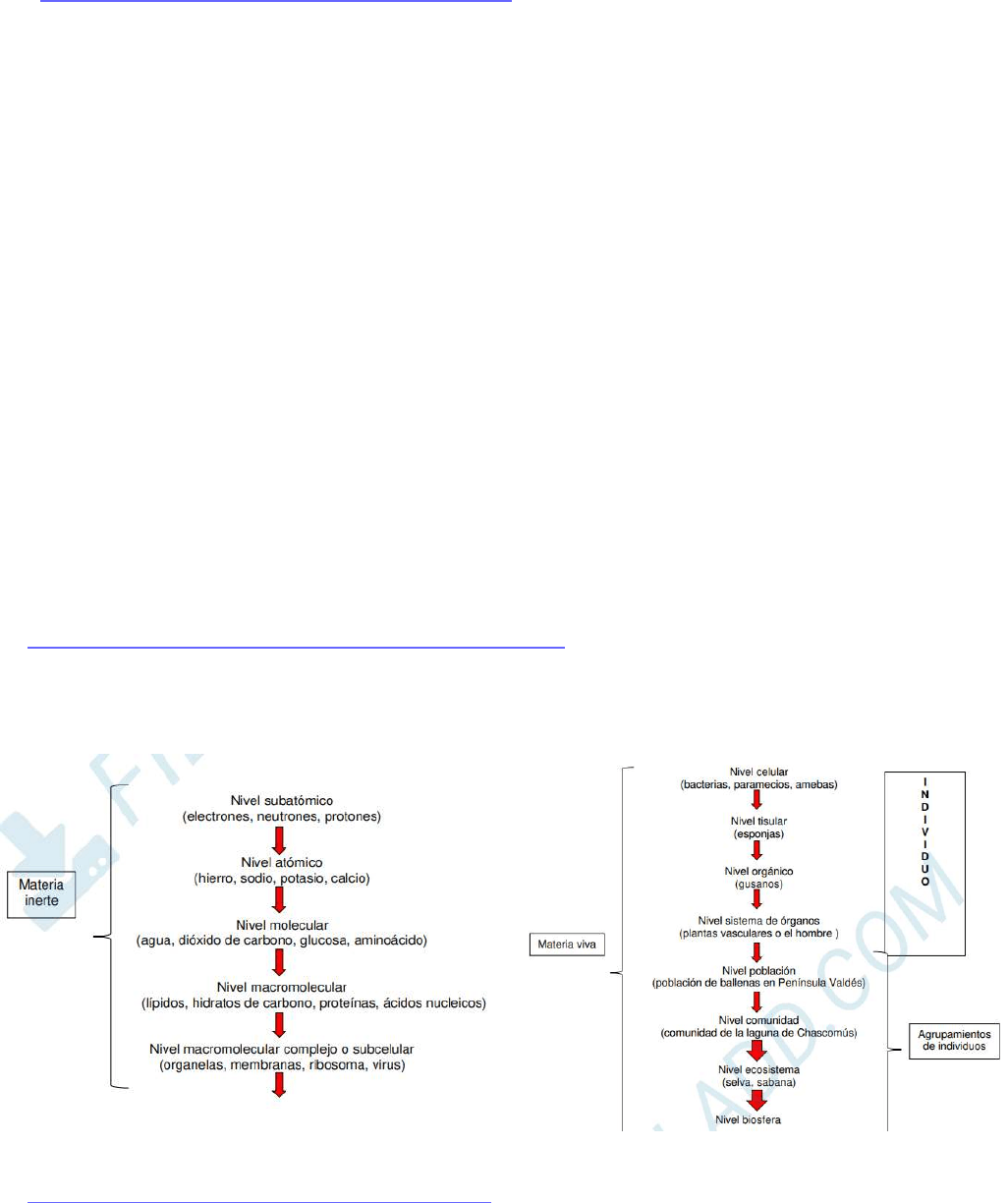
NIVELES DE ORGANIZACIÓN DE LA MATERIA
En cada nivel, la interacción entre sus componentes determina las propiedades de ese nivel.
CLASIFICACIÓN DE LA BIODIVERSIDAD
Se pueden tener en cuenta tres criterios, según las características fundamentales que posee cada individuo:
● El número de células, es decir unicelular o pluricelular
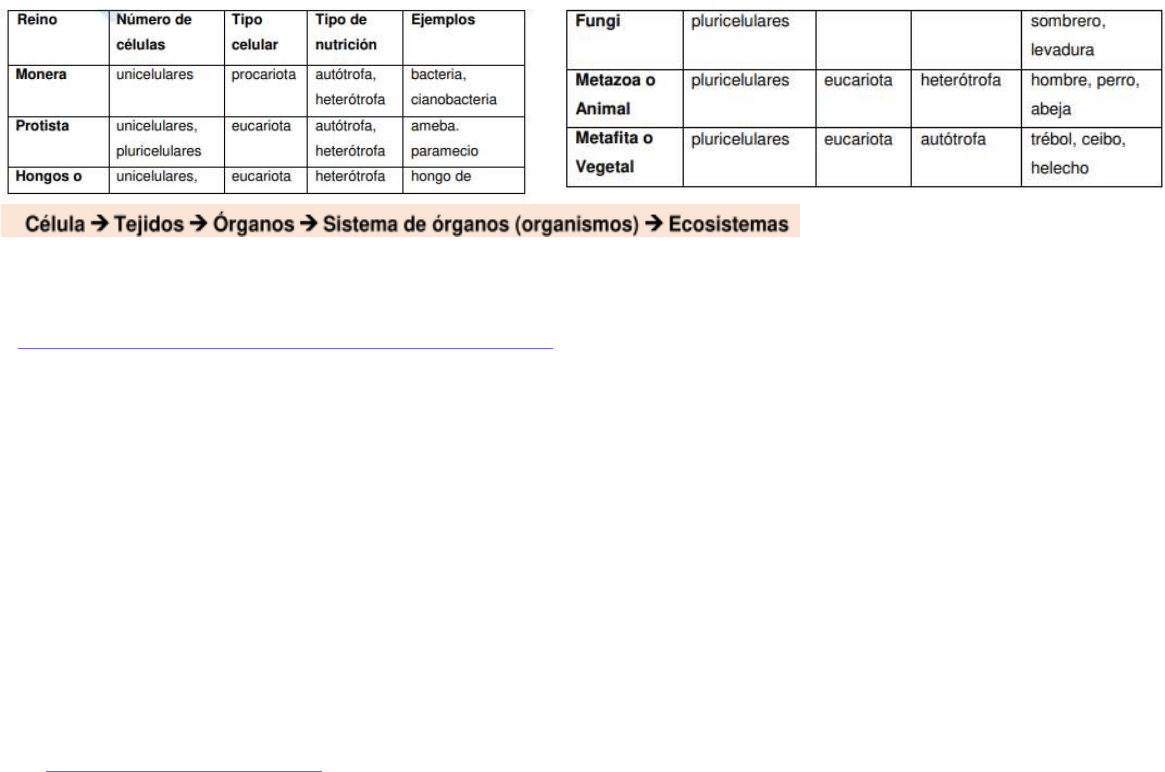
● El tipo de células eucariota (estructura lineal de ADN ligado a proteínas, ubicados en un núcleo bien definido) o procariota
(se caracterizan por tener una estructura grande y circular de ADN asociado débilmente a proteínas. No posee
endomembrana nuclear. El núcleo se encuentra en una zona del citoplasma llamado nucleoide.)
● La forma de nutrición autótrofa (Capaces de generar su propio alimento energético) o heterótrofa (Organismos que
necesitan incorporar moléculas orgánicas del exterior para producir energía y generar sus estructuras).
Se le llama
ECOSISTEMA a las interacciones (intercambio de materia y energía) de los organismos vivos en determinadas
condiciones ambientales.
ORGANIZACIÓN GENERAL DE LAS CÉLULAS
Poseen la capacidad de metabolismo y autoperpetuación. Por lo tanto, tienen vida propia y son responsables de las características
los organismos. La célula es la unidad estructural y funcional de los seres vivos.
● Todos los seres vivos están formados por células.
● El funcionamiento de un organismo es el resultado de la interacción entre las células que lo componen.
● Toda célula proviene de otra preexistente.
● Las células contienen material hereditario.
La forma depende de la disposición de ciertos elementos estructurales internos, como el citoesqueleto, y está generalmente
relacionada con las funciones especiales que esas células cumplen.
CÉLULA PROCARIOTA: en la zona más superficial se encuentra la CÁPSULA que está formada por un material mucoso
y permite adherirse a otros sustratos; en uno de los extremos se encuentra el FLAGELO, con aspecto de un largo cabello el cual le
permite desplazarse; por debajo de la cápsula existe una
PARED CELULAR RÍGIDA o flexible la cual brinda protección; más hacia
el interior la
MEMBRANA PLASMÁTICA que también rodea a toda la célula y formada por una bicapa de fosfolípidos asociados a
proteínas, sin colesterol. Presenta
pliegues hacia el interior que aumentan su superficie con diversas funciones (respiración
celular, fotosíntesis, etc.). Uno de esos pliegues es el
MESOSOMA (punto de unión de la membrana con el ADN). El interior de la
célula está ocupado por una masa tipo gelatinosa que es el
CITOPLASMA donde está incluida la molécula de ADN circular no
asociada a proteínas y ribosomas aislados o agrupados en poliribosomas.
PARED CELULAR: Es rígida, sirve como protección mecánica. Está compuesta por dos capas interna y externa. La interna se
compone de peptidoglicano (carbohidratos + péptidos). La capa
externa se compone de lipoproteínas y lipopolisacáridos. En la
pared celular también posee porinas, que son conductos cilíndricos donde se difunden los solutos.
MEMBRANA PLASMÁTICA: Se encuentra por debajo de la pared celular, es una estructura lipoprotéica que controla la entrada
y salida de solutos. En ella también se encuentran los complejos para realizar la fotosíntesis.

PROTOPLASMA: Equivalente a citoplasma en las células eucariotas. En el protoplasma se encuentran los ribosomas 70s en
grupos, encargados de la síntesis proteica
(los 70s se encuentran en procariotas y algunas eucariotas, son más pequeños que los
80s y están formados por más ARN que proteínas). Otras
sustancias que se encuentran en el protoplasma son el agua, iones y
proteínas.
CROMOSOMA BACTERIANO: Es una molécula única de ADN que se encuentra en el nucleoide. Este se encuentra unido a la
membrana plasmática. Algunas bacterias contienen un ADN pequeño llamado plásmido que otorga resistencia a antibióticos.
CÉLULA EUCARIOTA: Se diferencia de la procariota por la presencia de un núcleo bien definido y por poseer
compartimientos externos. Las poseen algunas bacterias, los animales, plantas y hongos. Las plantas y algunos hongos tienen
pared celular, formada de celulosa y quitina respectivamente.
ENDOMEMBRANAS: son bicapas lipídicas de todos aquellos compartimientos que posee la célula en su interior. Se separan
por bicapa al igual que la membrana plasmática, y se denomina cara citosólica al lado externo y cara luminal al lado interno de
la organela (bicapa de glicolípidos y glicoproteínas). Estas últimas se pueden comunicar directamente o por vesículas
transportadoras (que llevan solutos o señales de uno a otro).
VESÍCULAS TRANSPORTADORAS: Es una organela con bicapa lipídica que almacena, transporta o digiere productos
celulares. Se encargan de comunicar distintos compartimientos. Brotan de la endomembrana donante y viajan a través del
citosol buscando la endomembrana receptora. Poseen
cubiertas protéicas denominadas COP y CLARTINA.
•Cubierta COP: Existen COP I y COP II (diferente estructura). Las I se originan desde el RE al AG y las II se originan en el AG y
van al RE. Está cubierta se ubica del lado citosólico mientras se forma la vesícula, cubriéndola.
•Cubierta de clartina: Se originan en las vesículas de la membrana plasmática durante la endocitosis y también en el
complejo de Golgi dirigidas a la membrana plasmática. Estas cubiertas están formadas por unidades protéicas llamadas
trisqueliones y se originan desde el lado interno de la membrana plasmática. Los trisqueliones se unen con receptores y
porciones de la membrana, que son los que se filtran en el AG durante la endocitosis.
¿Qué es lo que guía a las vesículas transportadoras a su destino? Las guían dos proteínas, V-SNARE (de la vesícula) y T-
SNARE
(de la membrana destino). Dependiendo el camino a seguir, existen distintas v y t Snare, y cuando se encuentran, la
vesícula deposita su contenido allí. Luego vuelve a su lugar de origen porque la membrana emisora posee la SNARE
complementaria de la vesícula. [Cuando la vesícula vuelca el contenido, intervienen proteínas llamadas
fusógenas]
RIBOSOMAS: Se encuentran en el citosol y en el retículo endoplasmático rugoso. Su función es la síntesis de proteínas y su
estructura es un 60% ARN ribosomal y
el resto proteínas. Los que se encuentran en el citosol se encargan de sintetizar
proteínas destinadas al RER, y cuando las crean, lo hacen con una
péptida señal específica que direcciona al ribosoma hacia el
RER. En él,
depositan la proteína y este queda plegado en la superficie del RER. Los ribosomas son reconocidos por una
proteína llamada PRS del retículo
endoplasmático rugoso. Los ribosomas poseen tres sitios distintos en los que ocurren
diferentes
procesos. Está el sitio A, que es donde ingresa el ARNt combinado con los aminoácidos; el sitio P, que es donde se
alojan los anteriores; y el
sitio E que es por donde sale el ARNt.
RETÍCULO ENDOPLASMÁTICO: Se distribuye desde el núcleo celular hasta la membrana plasmática. Se diferencian en dos:
R.E. Liso y R.E. Rugoso. El
rugoso presenta ribosomas en su superficie, mientras que el liso no. Estos últimos poseen mayor
concentración de calcio que el citosol, al igual que el líquido extracelular.
Funciones del Retículo endoplasmático rugoso
*Síntesis de trigliceroles

*Regeneración de las membranas celulares (ya que envejecen). Primero retiene los componentes de la membrana a
regenerar y luego las envía por vesículas. Los lípidos que produce el RE son fosfatildicolina, fosfatildietanoamina,
fosfatildiserina y fosfatidinositol. Las vesículas ubican estos componentes en la capa citosólica de la membrana plasmática (o la
externa de las endomembranas) y luego por flip-flop se van distribuyendo en la bicapa.
*Formación de glicoproteínas con las proteínas que ingresan al RE (traídas por los ribosomas), donde luego, son
transportadas al aparato de Golgi.
*Síntesis de proteoglicanos, que luego son llevados a la membrana plasmática para formar parte del glico cáliz (material
extracelular proteínacarbohidrato).
Funciones del retículo endoplasmático liso
*Síntesis de esteroides y síntesis de lipoproteínas.
APARATO DE GOLGI: Se encuentra entre el retículo endoplasmático y la membrana celular. En este, las moléculas sufren
transformaciones para posterior actividad
biológica. El aparato de Golgi está compuesto por unidades llamadas dictiosomas,
que tienen forma curvada con cara convexa (U) hacia el
núcleo y cóncava (n) hacia la membrana plasmática. La cara de
entrada (U) sólo recibe moléculas de vesículas transportadoras
desde el retículo endoplasmático. La cara de salida envía
mediante
vesículas moléculas a la membrana plasmática o a los endosomas.
Funciones del aparato de Golgi
*Desde el aparato de Golgi ocurre la exocitosis. Proceso en donde las vesículas generadas desde el aparato de Golgi
transportan al exterior de la célula las sustancias a desechar, fusionándose la vesícula con la membrana plasmática. Puede ser:
Constitutiva: Las sustancias se segregan de forma automática desde el AG hasta la membrana plasmática. Regulada: Las
vesículas se mantienen en el citoplasma hasta que llegue una señal de secreción.
*También es el encargado de
sintetizar glucolípidos cerebrósidos y gangliósidos.
CÉLULA EUCARIOTA VEGETAL: hay una estructura muy desarrollada, llamada VACUOLA altamente desarrollada
limitada por una membrana llamada TONOPLASTO. Otras de las estructuras que aparecen como características de la célula
vegetal son estructuras discoidales con repliegues
en su interior, son los CLOROPLASTOS; la PARED CELULAR Y LOS
PLASMODESMOS
, estos últimos son diferenciaciones de la membrana que mantienen unidas a las células y permiten la
circulación de sustancias del citoplasma entre células vecinas
GLIOXISOMAS (VEGETALES): Organelas que se encargan del metabolismo de los triglicéridos. La función es, mediante los
enzimas que posee en su interior, transformar ácidos grasos
de la semilla en carbohidratos.
VACUOLA (VEGETALES): Son orgánulos de las células vegetales que funcionan como lisosomas, aunque también poseen
otras funciones:
• Depósito de nutrientes o desechos metabólicos (reserva energética también).
• Almacén de líquidos que regula el volumen de la célula.
CITOSOL: Contiene componentes muy variados. Su pH es de 7,20.s
Componentes del citosol
* Inclusiones:
Macromoléculas que se acumulan en el citosol parecido a un coloide. Pueden ser de glucógeno o de grasas
(trigliceroles). Se encuentran en las células musculares y funcionan como reserva energética.
* Ribosomas: Una parte de los ribosomas celulares se encuentran en el citosol, y la otra, fue redirigida hacia algun
compartimiento (RER, Mitocondria o Peroxisoma).

* Chaperonas: Son proteínas que se encuentran junto a las proteínas que se están sintetizando y se encargan de que los
plegamientos sean correctos. Existen tres tipos hsp 70, hsp 60 y hsp 90 (llamadas así por el peso molecular).
Las
hsp 70 son estructuras poliméricas de polipéptidos, y se encargan de prevenir los plegamientos prematuros a medida
que la proteína que se está sintetizando sale del ribosoma.
Los
hsp 60 aparecen cuando el hsp70 falla, corrigiendo los plegamientos de las proteínas.
* Proteosomas: Son complejos enzimáticos encargados de destruir las proteínas que se plegaron mal, que son erróneas o
que cumplieron su función. Saben qué proteína destruir ya que estas están marcadas por un conjunto de polipéptidos
llamados ubiquitinas. Consumen energía del ATP
COMPONENTES CELULARES COMUNES EN EUCARIOTAS ANIMALES Y VEGETALES:
Membrana plasmática
Núcleo
Mitocondrias
Ribosomas
sistema de Golgi
lisosomas
peroxisomas
REL y RER.
En las
CÉLULAS EUCARIOTAS existe una variedad de estructuras internas, ellas son las ORGANELAS que son similares o, en
algunos casos, idénticas de una célula a otra, en varios tipos celulares.
Las células están separadas del medio circundante por una
MEMBRANA CELULAR. Esta membrana funciona como la puerta
de entrada y de salida de la célula permitiendo el intercambio entre el medio intracelular y el medio extracelular. Las
membranas son barreras con permeabilidad selectiva.
Las células de las plantas, están además separadas del ambiente por una
PARED CELULAR elaborada por las mismas células.
El CITOPLASMA de la célula es una solución acuosa concentrada que contiene enzimas, moléculas disueltas e iones.
En las
CÉLULAS EUCARIOTAS, el CITOESQUELETO (que es una red compleja de filamentos proteicos) ocupa todo el
citoplasma. El CITOPLASMA está atravesado, por un complejo SISTEMA DE MEMBRANAS, el RETÍCULO ENDOPLÁSMICO que
sirve como superficie de trabajo para muchas actividades bioquímicas, y en algunas áreas está cubierto por
RIBOSOMAS,
que son las estructuras sobre las cuales los aminoácidos se ensamblan en proteínas. También se encuentran ribosomas en
otras partes del citoplasma y son las estructuras más numerosas tanto en las células procariotas como en las eucariotas.
Cuanto más proteínas esté fabricando una célula, más ribosomas tendrá. O sea que, la cantidad de retículo endoplásmico de
una célula no es fija, sino que aumenta o disminuye de acuerdo con la actividad celular. El conjunto de membranas que
forma el retículo y posee ribosomas en su cara externa, se denomina
RETÍCULO ENDOPLASMÁTICO RUGOSO y es
especialmente abundante en células que producen proteínas de exportación.
El RETÍCULO ENDOPLÁSMICO LISO, que carece de ribosomas, es abundante en células especializadas en la síntesis lipídica o
en el metabolismo de lípidos como las células glandulares que producen hormonas esteroides y también se encuentra muy
desarrollado en las células hepáticas, donde parece estar relacionado con varios procesos de desintoxicación.
El
COMPLEJO DE GOLGI son zonas de empaquetamiento o compactación de las moléculas sintetizadas dentro de la célula.
Los
LISOSOMAS Y PEROXISOMAS son vesículas en las cuales diferentes tipos de moléculas se degradan a constituyentes
más simples que pueden ser utilizados por la célula o en el caso de productos de desecho, eliminados fácilmente. Las
MITOCONDRIAS son el asiento de las reacciones químicas que suministran energía para las actividades celulares.
El NÚCLEO es un cuerpo grande, frecuentemente esférico y, por lo común, es la estructura más voluminosa dentro de las
células eucariotas. El núcleo está separado del citoplasma por la
ENVOLTURA NUCLEAR formada por una doble membrana.
Dentro de la envoltura nuclear se encuentra un
NUCLÉOLO, que es el sitio donde se forman las subunidades ribosómicas y
se realiza la síntesis de proteínas ribosomales. Los
POROS de la envoltura nuclear suministran los canales a través de los
cuales, pasan las moléculas (
proteínas ribosomales) desde y hacia el citoplasma. En las células eucariotas, el núcleo contiene
el
MATERIAL GENÉTICO, el ADN que es lineal a diferencia del ADN procariota que es circular, y está fuertemente unido a
proteínas especiales llamadas histonas.
En las células vegetales, como se dijo anteriormente, la
MEMBRANA PLASMÁTICA está rodeada por una pared celular, la
VACUOLA frecuentemente ocupa la mayor parte de la células, mantiene la turgencia de la célula y puede almacenar
nutrientes o productos de desecho.
Los CLOROPLASTOS, son las organelas grandes en las que ocurre la fotosíntesis, generalmente se concentran cerca de la
superficie de la célula.
El
COMPLEJO DE GOLGI es un centro de procesamiento y compactación de materiales que se mueven a través de la célula y
salen de ella. Cada complejo de Golgi recibe vesículas del retículo endoplasmático, modifica sus membranas y sus contenidos
e incorpora los productos terminados en vesículas de transporte que los llevan a otras partes del sistema de
endomembranas, a la superficie celular y al exterior de la célula.
Los ribosomas, el retículo endoplásmico y el complejo de Golgi y sus vesículas cooperan en la síntesis, procesamiento químico,
empaquetamiento y distribución de macromoléculas y nuevo material de membrana.
Los
LISOSOMAS, un tipo de vesícula relativamente grande, formada en el complejo de Golgi, contienen enzimas hidrolíticas a las
que aíslan de la célula y están implicados en las actividades digestivas intracelulares de algunas células. Estas enzimas están
implicadas en la degradación de proteínas, polisacáridos, ácidos nucleicos y lípidos. Para su óptima actividad, las enzimas
hidrolíticas requieren de un medio ácido. Los lisosomas proveen este medio. Las ENZIMAS LISOSOMALES son capaces de
hidrolizar a todos los tipos principales de macromoléculas que se encuentran en una célula viva. Las enzimas hidrolíticas que los
lisosomas liberan en las vacuolas, digieren su contenido. Las enzimas no destruyen la membrana de los lisosomas que las
contienen. Los PEROXISOMAS son otro tipo de vesícula relativamente grande presente en la mayoría de las células eucarióticas;
contienen enzimas oxidativas que remueven el hidrógeno de pequeñas moléculas orgánicas y lo unen a átomos de oxígeno
formando peróxido de hidrógeno, un compuesto que es extremadamente tóxico para las células.
Otras organelas vinculadas con la transformación de energía, son las
MITOCONDRIAS, que son organelas limitadas por
membrana en las cuales las moléculas orgánicas que almacenan energía química son degradadas y la energía liberada es
envasada en unidades más pequeñas. En este proceso, la energía liberada es almacenada en moléculas de ATP que será utilizada
luego en otros procesos celulares.
En general, cuando se trata de una célula eucariota vegetal, se mencionan a los
PLÁSTIDOS y se los clasifica en tres tipos:
leucoplastos, cromoplastos y cloroplastos. Los CLOROPLASTOS son los plástidos que contienen clorofila y en los cuales se
produce energía química a partir de energía lumínica, en el proceso de fotosíntesis. Están rodeados por dos membranas. Los
CILIOS Y FLAGELOS tienen la misma estructura, sólo que, cuando son cortos y aparecen en cantidades grandes se los llama cilios y
cuando son más largos y más escasos se los llama flagelos. Están asociados con el movimiento del organismo.
La principal diferencia entre estos tipos celulares eucariotas y procariotas se debe a la compartimentalización. En las
CÉLULAS
EUCARIOTAS
, cada compartimiento está destinado a una cierta función particular, lo que las hace más eficientes. En los
organismos
PROCARIONTES todas las funciones se realizan en el citoplasma, o bien en la membrana plasmática.

Para el estudio de la célula y sus componentes se utilizan los
MICROSCOPIOS. Existen dos tipos básicos de microscopios: el
microscopio
óptico y el electrónico. Dentro de estos últimos, existen los microscopios electrónicos de transmisión y
microscopios de barrido
.
Con el
MICROSCOPIO ÓPTICO, se puede identificar la presencia o ausencia del núcleo, de las mitocondrias y cloroplastos, la
forma celular. El microscopio electrónico
DE TRANSMISIÓN permite detectar los detalles a escala macromolecular es decir,
la ultraestructura por ejemplo, de la membrana plasmática. El microscopio electrónico
DE BARRIDO, permite reconocer
superficies a través de imagenes tridimensionales.
VIRUS, VIROIDES, PRIONES
Se definen como parásitos intracelulares obligados ya que no son considerados células porque no realizan funciones básicas
de la materia viva como el metabolismo y la reproducción. Para ello, deben infectar a una célula y utilizar de la misma, toda
la maquinaria de síntesis. Si bien esta es la característica común, se diferencian entre ellos por el tipo de moléculas que los
constituyen.
VIRUS: La estructura del VIRUS ENVUELTO está representada por un círculo, rodeado totalmente de pequeñas esferas.
Esa capa externa con la sucesión de pequeñas esferas constituye la ENVOLTURA del virus. Hacia adentro, se encuentra una
CUBIERTA PROTEICA que encierra al material genético, ya sea ADN o ARN. La CÁPSIDE está formada por capsómeros. El
esquema de la izquierda, correspondiente a un virus desnudo, es igual pero
sin la envoltura.
En síntesis, la estructura general de los virus consiste en: Una cubierta
proteica denominada
CÁPSIDE que está formada por capsómeros y que
encierra al ácido nucleico, ya sea ADN o ARN. Algunos
VIRUS ENVUELTOS,
pueden tener otra cubierta más de lípidos, proteínas y glucoproteínas. Por
estar formados por una asociación de macromoléculas, pertenecerían al
nivel de organización macromolecular complejo o subcelular.
VIROIDES: Son agentes infecciosos constituidos exclusivamente por una molécula de ARN. Infectan
fundamentalmente a plantas. Pertenecen al nivel de organización macromolecular por estar formados por tan solo una
molécula de ARN.
PRIONES: Son proteínas infecciosas mal plegadas capaz de transmitir su mal plegamiento a otras. Constituyen
enfermedades degenerativas. Agentes responsables de encefalopatías espongiformes transmisibles, que afectan al sistema
nervioso central. Pertenecen al nivel de organización macromolecular por estar formados por una proteína.
MEMBRANA Y MECANISMOS DE TRANSPORTE DE MEMBRANA
CARACTERÍSTICAS generales de las membranas biológicas
El modelo que describe la estructura básica de toda membrana es el “MODELO DE MOSAICO FLUIDO”.
Son estructuras laminares constituidas por
LÍPIDOS y por PROTEÍNAS. La mayor parte posee también HIDRATOS DE
CARBONO
unidos a las proteínas y a los lípidos.
Son fluídas, lo que permite a los lípidos y proteínas desplazarse en la bicapa por:
MOVIMIENTO LATERAL (dentro de
una misma monocapa),
ROTACIÓN (giro sobre su propio eje) y FLIP-FLOP (entre una monocapa y la otra).
Son
ASIMÉTRICAS en cuanto a la disposición de sus componentes.
Poseen una
PERMEABILIDAD SELECTIVA, es decir que es permeable a algunas sustancias y a otras no. Por eso, las
membranas biológicas son “semipermeables”.
FUNCIONES de las membranas biológicas
SEPARAN COMPARTIMIENTOS de composición química distinta (por ejemplo, la membrana plasmática separa el
medio intracelular del extracelular) y permite que esas diferencias se mantengan.
COMPARTIMENTALIZACIÓN: en eucariontes el trabajo celular se divide en distintos compartimientos
membranosos.
TRANSPORTE de partículas.
INTERACCIÓN entre células y con la matriz extracelular.
ANCLAJE del citoesqueleto.
SOPORTE de enzimas.
Los componentes de la membrana son
LÍPIDOS, como los fosfolípidos y el colesterol, este último sólo presente en las
células eucariotas animales. Otros componentes de las membranas son las PROTEÍNAS INTEGRALES Y PERIFÉRICAS y
además, los
HIDRATOS DE CARBONO asociados a lípidos formando los GLUCOLÍPIDOS o a proteínas conformando las
GLUCOPROTEÍNAS.
1) Lípidos
TRIACILGLICEROLES: ácidos grasos unidos a cada OH de un glicerol mediante una unión ester (OH del ácido graso con
OH del glicerol). Funcionan como reserva de energía para la célula (ya que cuando se oxida un ácido graso libera el
doble de energía que un carbohidrato).
FOSFOLÍPIDOS: Son dos ácidos grasos unidos a dos carbonos del glicerol por enlace éster, mientas que el tercer
carbono posee un grupo fosfato. Dentro de esta clasificación se encuentran los
glicerofosfolípidos (aquellos que el
fosfato posee un alcohol unido). Debido a su naturaleza anfipática espontáneamente en medios acuosos se disponen
formando
bicapas. La fluidez de la membrana depende de las características de las cadenas hidrocarbonadas de los
fosfolípidos (
INSATURADAS aumentan la fluidez; SATURADAS disminuyen la fluidez). La distribución de los
fosfolípidos es
ASIMÉTRICA, algunos fosfolípidos se ubican solamente en una de las caras de la membrana.
ESFINGOLÍPIDOS: Son todos aquellos formados por un aminoalcohol esfingosina con ácidos grasos. Pueden formar lo
que se llaman
ceramidas, esfingosinas + ácido graso unido por enlace amida.
GLUCOLÍPIDOS: Son aquellos formados por un oligosacárido y una ceramida. Se clasifican en cerebrósidos si el
monómero del oligosacárido es único, y
gangliósidos si presenta más de un monosacárido.
ESTEROIDES: Se le denominan esteroides a los lípidos derivados del ciclopentanoperhidrogenantreno. Son
insaponificables, y son los que componen las
hormonas sexuales.
POLIPRENOIDES: Se le denominan así a aquellos derivados del isopreno.
COLESTEROL: aumenta la IMPERMEABILIDAD de la bicapa a moléculas hidrofílicas y regula el grado de FLUIDEZ. Si
aumenta el colesterol, la fluidez disminuye y viceversa.
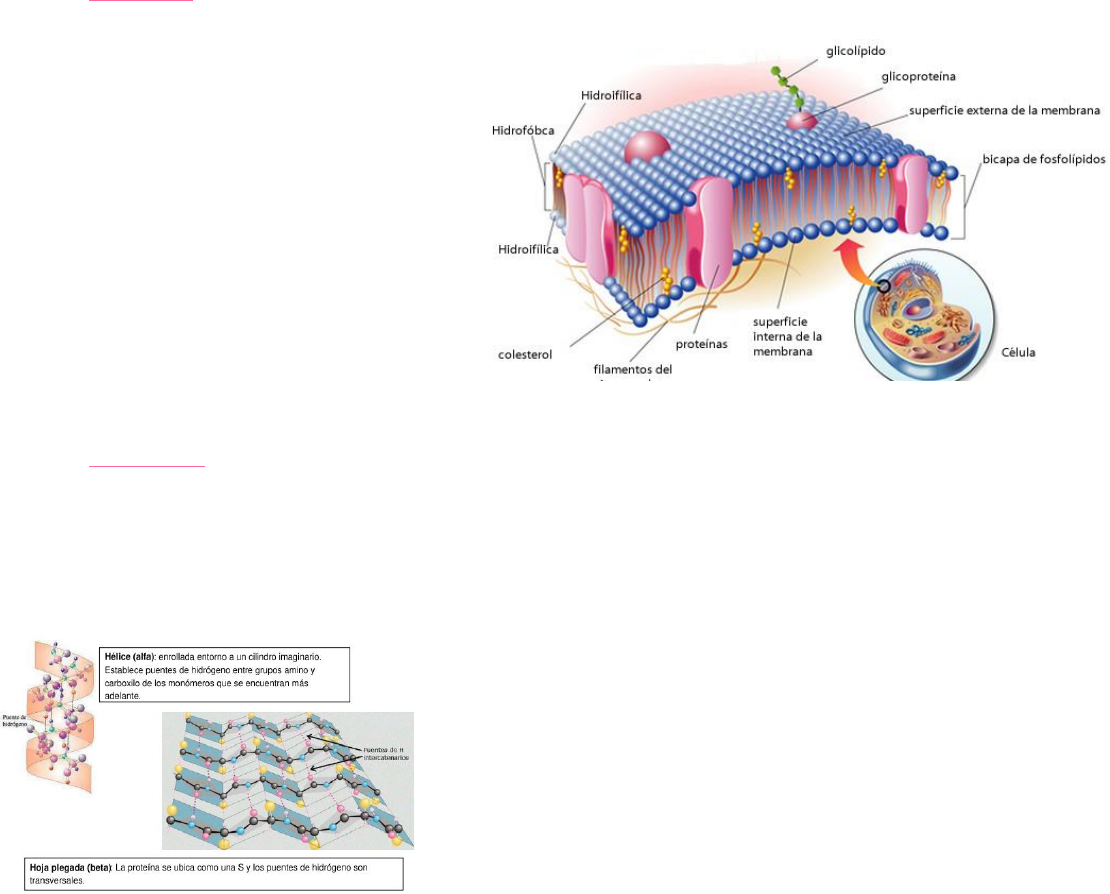
2) Proteínas: Son el COMPONENTE FUNCIONAL de las membranas (proteínas receptoras, canal, enzimas, etc.). Se
clasifican según su ubicación en la bicapa lipídica y si se asocian o no a otras moléculas. Son cadenas de aminoácidos
(ácidos con un grupo amino) que se unen para formar
péptidos (3 amin.), oligopeptidos (3-10 amin), polipéptidos
(10- 100 amin) y finalmente
proteínas (+100 amin). Los aminoácidos se unen mediante uniones peptídicas que se
dan entre el grupo amino de uno y el carboxilo de otro, con la pérdida de una molécula de agua
TIPOS DE PROTEÍNAS
Enzimáticas
Estructurales
Conjugadas (unidas con algo no protéico)
Glicoproteínas (asociadas con carbohidratos)
Lipoproteínas (asociadas con lípidos)
Nucleoproteínas (asociada con ácidos nucleicos)
Cromoproteínas (asociadas con un pigmento)
Según su
UBICACIÓN en la membrana:
INTEGRALES: están insertas dentro de la
bicapa. Interactúan tanto con las cabezas
como con las colas de los fosfolípidos y
atraviesan completamente la membrana, por
eso se las denomina también
proteínas
transmembrana.
Estas se distribuyen
asimétricamente ya que los polipéptidos que
emergen en ambas caras son diferentes.
PERIFÉRICAS: no penetran en el interior
hidrofóbico de la bicapa por lo que se asocian
con la membrana a través de las cabezas de
los fosfolípidos o de las proteínas integrales.
Su distribución es también asimétrica.
Según su ASOCIACIÓN a otras moléculas, las proteínas pueden ser GLICOPROTEÍNAS (glúcidos y proteína) o
LIPOPROTEÍNAS (lípidos y proteínas).
ORGANIZACIÓN ESTRUCTURAL DE LAS PROTEÍNAS
PRIMARIA: es una secuencia de aminoácidos que forma una unidad proteica.
SECUNDARIA: Posee una configuración espacial, puede ser tanto de hélice como de hoja plegada:
TERCIARIA: Son proteínas que toman una posición espacial como consecuencia de las atracciones que sufre la
misma, se plega. Estos plegamentos se originan por la atracción de los aminoácidos. Pueden ser solo en las alfas, solo
en betas, o en la mezcla de ambas. [La estructura terciaria es la que le da la función a la proteína, su pérdida indica la
desnaturalización] Si los plegamientos son solo en proteína alfa, se les denominan fibrosas. Si son betas o mezcla de
ambas, se denominan
globulares (por la forma esférica).
CUATERNARIA: Se origina a partir de la atracción entre estructuras terciarias, que al igual que ellas, puede ser por
puentes de H o puentes disulfuro, Van der Waals, etc.
3) Hidratos de carbono (moléculas orgánicas formada por c, h y o)
Se presentan en forma de OLIGOSACÁRIDOS unidos covalentemente a lípidos (GLUCOLÍPIDOS) o a proteínas
(
GLUCOPROTEÍNAS) de la membrana. Se distribuyen en forma asimétrica, se encuentran en la monocapa que da hacia
el medio extracelular y en el
sistema de endomembranas en la monocapa interior. El conjunto de todos los
oligosacáridos de la membrana plasmática forman una cubierta denominada
GLUCOCÁLIX.
Tienen funciones tanto estructurales como de energía celular a corto plazo.
Se
CLASIFICAN en los siguientes:
MONOSACÁRIDOS: azúcares simples, patrones que se repiten en la macromolécula. Cadenas de 3 a 6 C.
DÍSACÁRIDOS: azúcares formados por dos unidades monómeros unidos por enlace O-glucosídico (grupo OH de uno
con el OH del otro, una unión ‘eter’ liberando agua).
OLIGOSACÁRIDOS: Combinación hasta 9 monosacáridos. Se pueden combinar con lípidos o proteínas para formar
respectivamente glicolípidos y glicoproteínas.
POLISACÁRIDOS: +10 monosacáridos hexosas
Los OLIGOSACÁRIDOS que existen en la membrana de las células de la sangre (de los antígenos) son los que determinan
el grupo sanguíneo (ya que pueden poseer distintos de estos). Los ANTÍGENOS son sustancias que el sistema
inmunológico reconoce y no las trata como agentes externos.
GRUPO SANGUÍNEO 0:
Antígeno 0, es decir, aquél que se puede transformar en A o en B (no posee inicialmente A o
B). Por ello son dadores universales. Como no poseen antígenos ni A ni B, desarollan anticuerpos contra estos (y por
eso no pueden recibir sangre A o B).
GRUPO A:
La membrana celular sanguínea posee antígeno A y desarrollan anticuerpos contra los B.
GRUPO B:
Poseen antígenos B y desarrollan anticuerpos contra los A.
Grupo AB:
Poseen antígenos A y B en la membrana celular, no fabricando anticuerpos contra ninguno.
ORGANIZACIÓN de la membrana plasmática
La organización de las membranas biológicas es explicada por el modelo del “MOSAICO FLUIDO” (Singer y Nicholson).
Este modelo describe una bicapa de lípidos con proteínas dispuestas como un mosaico con fluidez, dinámicas y una
disposición
asimétrica de sus componentes. Además poseen glúcidos asociados a la cara externa.
Las membranas consisten en una
DOBLE CAPA DE FOSFOLÍPIDOS DIFERENTES ENTRE SI, cuyas colas hidrofóbicas están
dispuestas hacia el centro de la bicapa mientras que las cabezas hidrofílicas se disponen hacia el medio acuoso, sea el
citoplasmático o interno como el externo. Esta doble capa posee
COLESTEROL, biomolécula presente únicamente en las
células eucariotas animales.
La membrana limita el medio externo o extracelular del medio interno o intracelular. La doble capa de fosfolípidos está
atravesada por PROTEÍNAS. Además, la cara que da al medio extracelular posee una serie de GLÚCIDOS, generando
prolongaciones a modo de antenas. Dichos oligosacáridos se unen a los lípidos o a las proteínas formando
GLUCOLÍPIDOS O GLUCOPROTEÍNAS.
TRANSPORTE a través de la membrana

Las membranas son barreras de PERMEABILIDAD SELECTIVA. Esta selección, se basa fundamentalmente en
características de la membrana y de las moléculas que atraviesan la membrana: la
POLARIDAD o la presencia de una
CARGA NETA, el TAMAÑO y el GRADIENTE DE CONCENTRACIÓN.
Los mecanismos de transporte se pueden agrupar en pasivos y activos. Dentro del
TRANSPORTE PASIVO, se encuentran:
la
difusión simple, la ósmosis y la difusión facilitada.
Dentro de la difusión facilitada, se encuentran los carriers o permeasas, los canales y el cotransporte.
Dentro del TRANSPORTE ACTIVO, se incluyen a los sistemas bombas y al transporte en masa.
El
transporte en masa, puede ser endocitosis, como por ejemplo la pinocitosis, fagocitosis y endocitosis mediada por
receptor y la
exocitosis.
1) Transporte pasivo:
son transportes que
ocurren sin gasto directo o acoplado de
energía metabólica.
A- DIFUSIÓN SIMPLE: es a favor del gradiente
de concentración. Las moléculas atraviesan
libremente la bicapa. No intervienen
proteínas de membrana. Ejemplos: gases, el
benceno, metanol, etanol y glicerol.
B- DIFUSIÓN FACILITADA: es un transporte
mediado por proteínas de membrana. Por este mecanismo, pueden transportarse
moléculas polares sin carga
(glucosa, por ejemplo) o que tengan carga neta (un ión).
b.1- Carriers o permeasas (pueden dar lugar a transportes pasivos como activos. Existen tres tipos de permeasas:
monotransporte (la que permite el paso de un solo soluto), cotransporte (la que permite el paso de dos solutos en
un mismo sentido) o la
contratransporte (que permite el paso de dos solutos en sentido opuesto.
b.2- Canales
b.3-
COTRANSPORTE: transporte simultáneo de dos partículas, una a favor de su gradiente y la otra en contra del
gradiente. Hay dos casos:
● Simporte: si ambas moléculas son transportadas en el mismo sentido.
● Antiporte: si ambas moléculas son transportadas en sentido contrario.
C- ÓSMOSIS: es el pasaje de agua desde un compartimiento hacia
otro, ambos separados por una
membrana semipermeable .El
pasaje de agua se produce siempre desde las soluciones
más
diluidas
o hipotónicas (que poseen menor presión osmótica) a las
soluciones
más concentradas o hipertónicas (las de mayor
presión).
2) Transporte activo: es un mecanismo de transporte que ocurre con gasto directo o acoplado de energía
metabólica ya que los iones migran desde el lugar en que están menos concentrados al que están más
concentrados.
A-BOMBAS: Permiten el transporte de un ión o dos iones
acoplados, en contra de sus gradientes y con gasto de ATP.
Bomba Sodio-potasio: consiste en un contratransporte
(permeasa). Se encarga de mantener el potencial eléctrico
de la membrana plasmática, expulsando tres cationes sodio
al exterior e ingresando dos cationes potasio al citosol. (Por ello, el lado citosólico es más electronegativo). El
transporte pasivo Na+ Ca2+ depende de la bomba sodio-potasio (ingreso de un sodio y se retira un calcio).
Intercambio hidrógeno-sodio: Es un contratransporte que se encarga de mantener el pH de la célula. En él,
ingresa un catión sodio y se retira un protón.
Bomba potasio-hidrógeno: Ingresa potasio al citosol y libera protones al exterior por contratransporte. Los
protones liberados se segregan hacia el estómago (ácido estomacal).
Bomba protónica: Se da en los endosomas y lisosomas para aumentar el pH y ayudar a la degradación de
sustancias activando las enzimas hidrolíticas. En este caso, aumenta la concentración de protones.
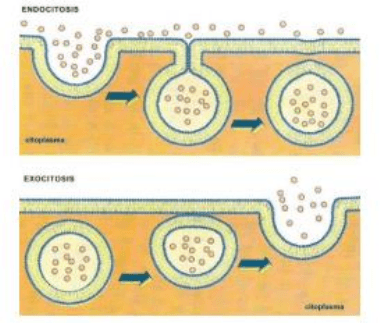
B-TRANSPORTE EN MASA:
Las partículas nunca atraviesan las membranas. Implica la formación de vesículas a partir de la membrana
plasmática (endocitosis) y
fusión secuencial de vesículas membranosas con la membrana plasmática (exocitosis). De
acuerdo a la dirección del transporte de las partículas hay dos tipos de transporte en masa:
B.1- ENDOCITOSIS: es el ingreso a la célula de partículas por
transporte en masa. Hay varios tipos de endocitosis de acuerdo al
material que ingresa:
● PINOCITOSIS: para líquidos o solutos pequeños en fase acuosa.
● FAGOCITOSIS: Se trata del mismo proceso que la pinocitosis pero
para partículas de
mayor tamaño.
● MEDIADA POR RECEPTOR:
es una endocitosis específica ya que
permite el ingreso de moléculas que sean
reconocidas por receptores
específicos que se encuentran en la membrana (por ejemplo el ingreso
de LDL, “lipoproteína de baja densidad” en inglés).
B.2- EXOCITOSIS: es la salida o secreción de sustancias al medio
extracelular por transporte en masa.
SISTEMA DE ENDOMEMBRANAS
Consiste en un conjunto de membranas intracelulares (en eucariontes) que se encuentran relacionadas física y
funcionalmente. Algunas de ellas, tienen continuidad o comunicación directa.
El Sistema de Endomembranas comprende a:
1- envoltura nuclear: está formada por dos membranas concéntricas separadas por un espacio. La membrana externa
tiene ribosomas adheridos.
2- retículo endoplasmático liso: está constituido por túbulos membranosos delgados que carecen de ribosomas.
Desempeña las siguientes funciones:
● Síntesis de lípidos.
● Detoxificación.
● Participa en uno de los pasos de la degradación del glucógeno.
● Almacenamiento y liberación de calcio: posee bombas de calcio.
3- retículo endoplasmático rugoso: se origina a partir de la membrana externa del núcleo. Tiene ribosomas
adheridos. Su función es la
síntesis de algunas proteínas, a saber:
● Proteínas de membrana.
● Proteínas de exportación o secreción.
● Enzimas que pertenecen al sistema vacuolar citoplasmático.
● Enzimas hidrolíticas o lisosomales.
SÍNTESIS DE PROTEÍNAS EN EL RETÍCULO ENDOPLASMÁTICO RUGOSO

A- La síntesis de proteínas, comienza en un RIBOSOMA libre en el citosol. Si la futura proteína será de exportación o de
membrana o lisosomal o del sistema vacuolar citoplasmático, los primeros aminoácidos del péptido que emergen
constituyen la péptida señal que indica que la síntesis deberá continuar en el
REG. La péptida señal es reconocida por la
partícula de reconocimiento de la señal que se une a ella y la síntesis de la proteína se detiene. Todo este complejo
(péptido, ribosoma más la partícula de reconocimiento de la señal) migra hacia el REG.
B- Este complejo se une específicamente a una proteína receptora de la membrana del REG, la RIBOFORINA. La
partícula de reconocimiento de la señal se disocia y se reanuda la síntesis proteica con la particularidad que a medida
que la cadena polipeptídica crece, va ingresando hacia la luz del REG por un proceso conocido como
DESCARGA
VECTORIAL
(es con consumo de energía y siempre en dirección hacia la luz del REG).
C- En la luz o interior del REG la péptida señal es
removido. Dentro del REG las proteínas pueden sufrir
modificaciones. En la mayoría de los casos las
proteínas son glucosiladas en la luz del REG o sea que
se agrega un
OLIGOSACÁRIDO PRESINTETIZADO que
se modificará finalmente en el
GOLGI.
D- La PROTEÍNA GLUCOSILADA sale del REG en una
vesícula y se traslada hacia el complejo de Golgi antes
de dirigirse al destino celular correspondiente.
4- Complejo de Golgi: complejo constituído por varias cisternas formadas por membranas apiladas en forma regular.
Cada una posee dos caras: una convexa o de
formación (en general orientada hacia los retículos) y otra cóncava o de
maduración (generalmente orientada hacia la membrana plasmática).
El Golgi es el principal
DISTRIBUIDOR de macromoléculas en la célula. Muchas de estas moléculas pasan a través del
Golgi para completar así su maduración. Este proceso incluye fundamentalmente la
GLUCOSILACIÓN definitiva de las
glucoproteínas y glucolípidos como así también, la
SEGREGACIÓN Y DIRECCIONAMIENTO de productos hacia sus
destinos finales.
5- Lisosomas: son VESÍCULAS membranosas que en su interior contienen enzimashidrolíticas. Se encargan de la
DIGESTIÓN intracelular de macromoléculas. Se caracterizan por tener un interior con pH 5 que se mantiene gracias a
una bomba de protones en su membrana.
Hay dos tipos de lisosomas:
● LISOSOMAS PRIMARIOS: son aquellos que todavía no tienen en sustrato a digerir
● LISOSOMAS SECUNDARIOS: son los que ya contienen el sustrato a digerir. Hay tres tipos de lisosomas secundarios:
VACUOLA DIGESTIVA: es el que aparece después de una fagocitosis o pinocitosis de material extraño. Este
material es digerido progresivamente por las enzimas hidrolíticas.
CUERPOS RESIDUALES: son los que resultan de una digestión incompleta. Serán eliminados por exocitosis.
VACUOLA AUTOFÁGICA: caso en que el lisosoma contiene partes celulares en vías de digestión.
6- Endosomas: son las organelas encargadas de recibir el
material que ingresa a la célula por
endocitosis. Se transforman
en
lisosomas una vez que incorporan ciertas enzimas hidrolíticas
que provienen del Golgi y que se sintetizaron en el Retículo
Endoplasmático Rugoso. Se los clasifica en
ENDOSOMAS
TEMPRANOS
(los recién formados y que por lo tanto están cerca
de la membrana) y ENDOSOMAS TARDÍOS (ya están alejándose de la membrana).
AUTOFAGIA: Eliminación de organoides envejecidos con ayuda del REL, se transfieren los componentes al endosoma
hasta el lisosoma. Este proceso también ocurre cuando falta alimento en el organismo, y las células activan este
mecanismo de emergencia.
7- Peroxisomas y glioxisomas: estas organelas no tienen ningún tipo de relación con el sistema vacuolar
citoplasmático ya que sus enzimas son
sintetizadas en el CITOSOL.
Peroxisomas: están presentes en todas las células eucariontes. Contienen enzimas oxidativas (oxidan por ejemplo
ácido úrico, oxalacetatos, ácidos grasos, purinas,
aminoácidos, etc.). A diferencia de lo que ocurre en las mitocondrias,
en estas
oxidaciones no se genera ATP. Como resultado las oxidaciones del peroxisoma se produce peróxido de
hidrógeno,
que es una molécula altamente tóxica.
Glioxisomas:
son exclusivas de las células eucariontes vegetales y se relacionan con el metabolismo de los
triglicéridos. Poseen enzimas que transforman los lípidos contenidos en las semillas en hidratos de carbono.
LAS CÉLULAS Y EL MEDIO
En eucariontes se denomina CITOPLASMA a todo lo que queda contenido entre la membrana plasmática y el núcleo.
Está compuesto por:
● CITOSOL: sustancia que ocupa los espacios que quedan entre organelas. Compuesto por agua, proteínas, sales.
● RIBOSOMAS: aislados o en polirribosomas. Pueden estar libres o adheridos al REG.
● SISTEMA DE ENDOMEMBRANAS.
● ORGANELAS:
mitocondrias, cloroplastos, peroxisomas, etc.
● CITOESQUELETO.
CITOESQUELETO:
es el conjunto de filamentos proteicos a modo de red,
formado por
microtúbulos, microfilamentos y filamentos intermedios.
Las FUNCIONES BÁSICAS son:
- da
forma a la célula y permiten el mantenimiento de esa forma
- participa en el
movimiento celular (de apoyo sobre un sustrato o
asociado a un medio acuoso)
- se relaciona con el
transporte intracelular de vesículas.
COMPONENTES del citoesqueleto
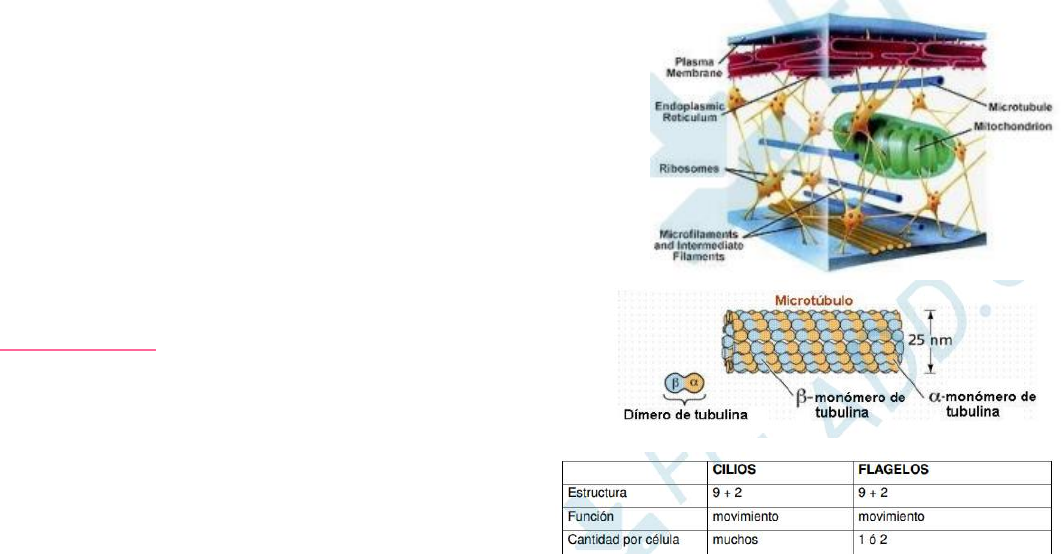
MICROTÚBULOS:
están formados por la proteína tubulina
(proteína globular). Se polimerizan y despolimerizan. Estas
alternancias entre polimerización y despolimerización se llaman
inestabilidad dinámica (crecimiento y acortamiento). Son los
responsables del transporte intracelular de vesículas. Participan
en la
división celular en la formación del huso mitótico y
meiótico. Forman
estructuras estables como cilios y flagelos y
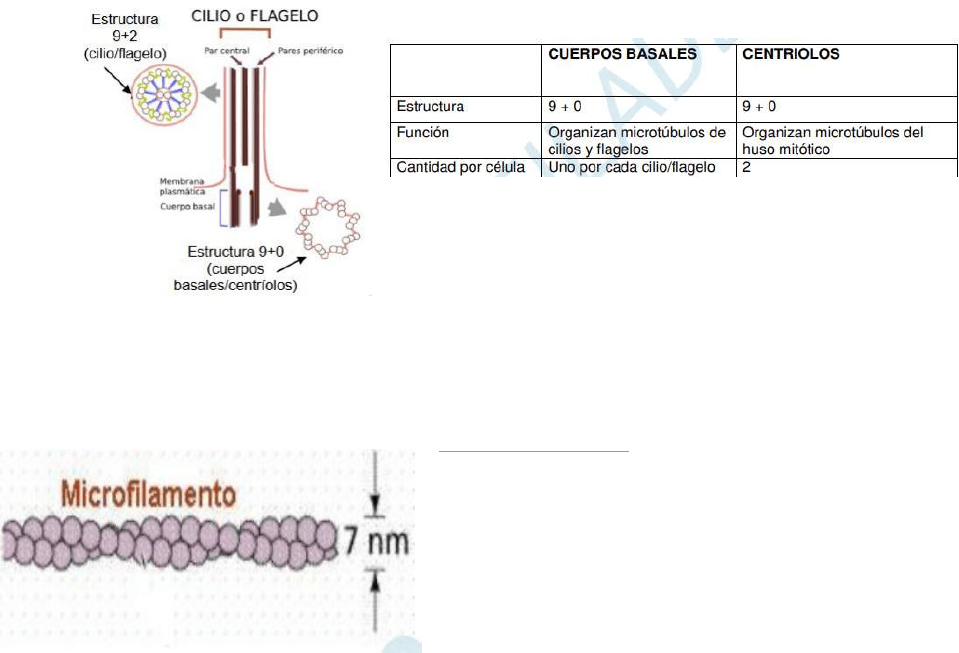
cuerpos basales y centríolos.
(La estructura 9+2 significa 9 pares de microtúbulos periféricos y dos
microtúbulos centrales. Estructura 9+0 significa 9 tripletes de microtúbulos
periféricos y ningún microtúbulo central.)
MOVIMIENTO CILIAR: Los cilios son estructuras que presentan algunas células que ayudan a movilizar fluidos y
partículas con el fin de movilizar a la célula. Se componen de microtúbulos paralelos entre sí llamados axonemas. Nacen
de la lámina basal, que se ubica en la matriz extracelular.
MICROFILAMENTOS: los microfilamentos de actina están formados por la
actina g (proteína globular). Se polimerizan y despolimerizan. Junto con la
miosina son responsables de la
contracción muscular. Participan en la división
celular en la
división del citoplasma. Responsables de la transición gel-sol del
citosol (producción de materiales sólidos a partir de moléculas pequeñas). Se
relacionan con la
emisión de prolongaciones celulares necesarias para
movimientos (de apoyo sobre una superficie), como filopodios, pseudópodos
y lamelipodios.
La
función de los filamentos de actina es contribuir a dar la forma celular, en las células epiteliales forman una capa por
debajo de la membrana plasmática. En estas, los
filamentos de actina son las que mueven las organelas por el
citoplasma, reguladas por las proteínas Miosina I y V.
*Miosina I: Posee una cola que se liga a la membrana de la organela a trasladar. Su cabeza se une a un filamento de
actina vecino y se transporta a base de los polos positivos y negativos del filamento.
Los filamentos de actina se clasifican en dos según donde estén ubicados: corticales (debajo de la membrana
plasmática) y transcelulares (a través del citoplasma).
MICROVELLOSIDADES: Son proyecciones citoplasmáticas nacidas en la superficie celular rodeada por la membrana
plasmática. Esas proyecciones, que las generan los filamentos de actina, incrementan la superficie celular, favoreciendo
la absorción de agua y solutos. Los filamentos intermedios y de actina crean un ‘’enrejado’’ por debajo de la membrana
plasmática. Y los filamentos de actina son los que nacen hacia la superficie de la célula y crean dichas irregularidades.
CÉLULAS MUSCULARES Y FILAMENTOS DE ACTINA: Las células musculares tienen forma cilíndrica y están adaptadas
para contraerse y expandirse. La contracción o expansión está dada por miofibrillas, que están compuestas por un
monómero llamado sarcómero. La expansión o contracción se da por los filamentos de actina que componen la unión
entre los sarcómeros, ubicados en
el disco Z. En el disco Z se juntan (contraen) los filamentos de actina o se expanden,
reguladas por la proteína
MIOSINA II.
MIGRACIÓN CELULAR:
Movimientos celulares hacia lugares específicos. El movimiento se basa en la polimerización y
despolimerización de filamentos de actina. Existen dos tipos de movimiento:
Este documento contiene más páginas...
Descargar Completo
RESUMEN BIOLOGIA.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.