REPLICACIÓN, REPARACIÓN Y RECOMBINACIÓN DEL ADN
La capacidad de una célula de mantener el orden en un ambiente caótico depende de la duplicación precisa
de la información genética codificada en su ADN.
Replicación del ADN ---> proceso de duplicación de la información genética.
Este proceso debe ocurrir antes de que una célula produzca dos células hijas genéticamente idénticas.
El mantenimiento del orden en una célula también requiere de la supervisión y la reparación de la info (si
esta es dañada por algún factor).
A pesar de que haya sistemas que protegen a la información genética de errores de copiado o daño, a veces
se producen cambios permanentes o mutaciones (tanto positivas como negativas). De hecho, la
acumulación de cambios en el ADN durante millones de años proporcionó la variedad genética que permite
la existencia de miles de especies diferentes. Además, pequeñas mutaciones también hacen a las pequeñas
diferencias entre individuos de una misma especie. Sin embargo, hay otras que pueden ser perjudiciales
para la salud, y son responsables de enfermedades hereditarias y varios tipos de cáncer.
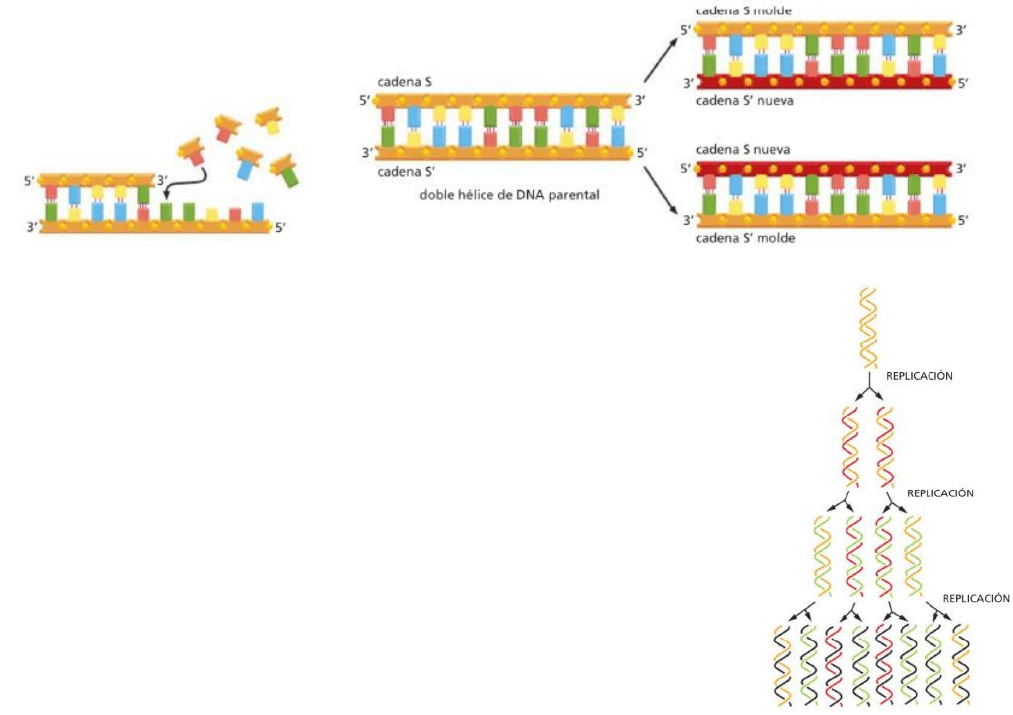
Replicación del ADN
Las cadenas de nucleótidos de la doble hélice de ADN son complementarias entre sí, cada una actuando
como molde de la otra en la síntesis de una nueva cadena complementaria.
El copiado se realiza con precisión y velocidad, con muy pocos errores (en el mejor de los casos). Este
proceso es realizado por una máquina de replicación.
La replicación del ADN produce dos dobles hélices completas a partir de la
molécula de ADN original y cada hélice nueva es idéntica en la secuencia de
nucleótidos a la doble hélice de ADN progenitora. El estilo de replicación es
semiconservadora ya que cada nueva hélice contendrá una de las cadenas
originales (antigua) y una cadena complementaria nueva.
*La cadena original separa sus cadenas, una por un lado y otra por el otro. Se
crea una nueva a partir de ese molde, complementando a la original y creando
una hélice nueva. Por lo tanto, las nuevas hélices tendrán una cadena vieja y
una nueva recién sintetizada.
Las dobles hélices de ADN son muy estables. Las bases de cada cadena están
relacionadas mediante enlaces de hidrógeno. Aunque en conjunto los enlaces
son fuertes, cada uno por separado es bastante débil.
A-T: dos enlaces de H.
G-C: tres enlaces de H.
Proteínas iniciadoras se unen al ADN y separan las dos cadenas, rompiendo los enlaces de hidrógeno
entre las bases (no cuesta mucha energía). Secuencias de nucleótidos marcan los orígenes de replicación,
el lugar por donde el ADN se abre primero. Los tramos de ADN ricos en A-T (pares con menos enlaces)
suelen ser los que se encuentran en los orígenes de la replicación ya que son más fáciles de separar. Una
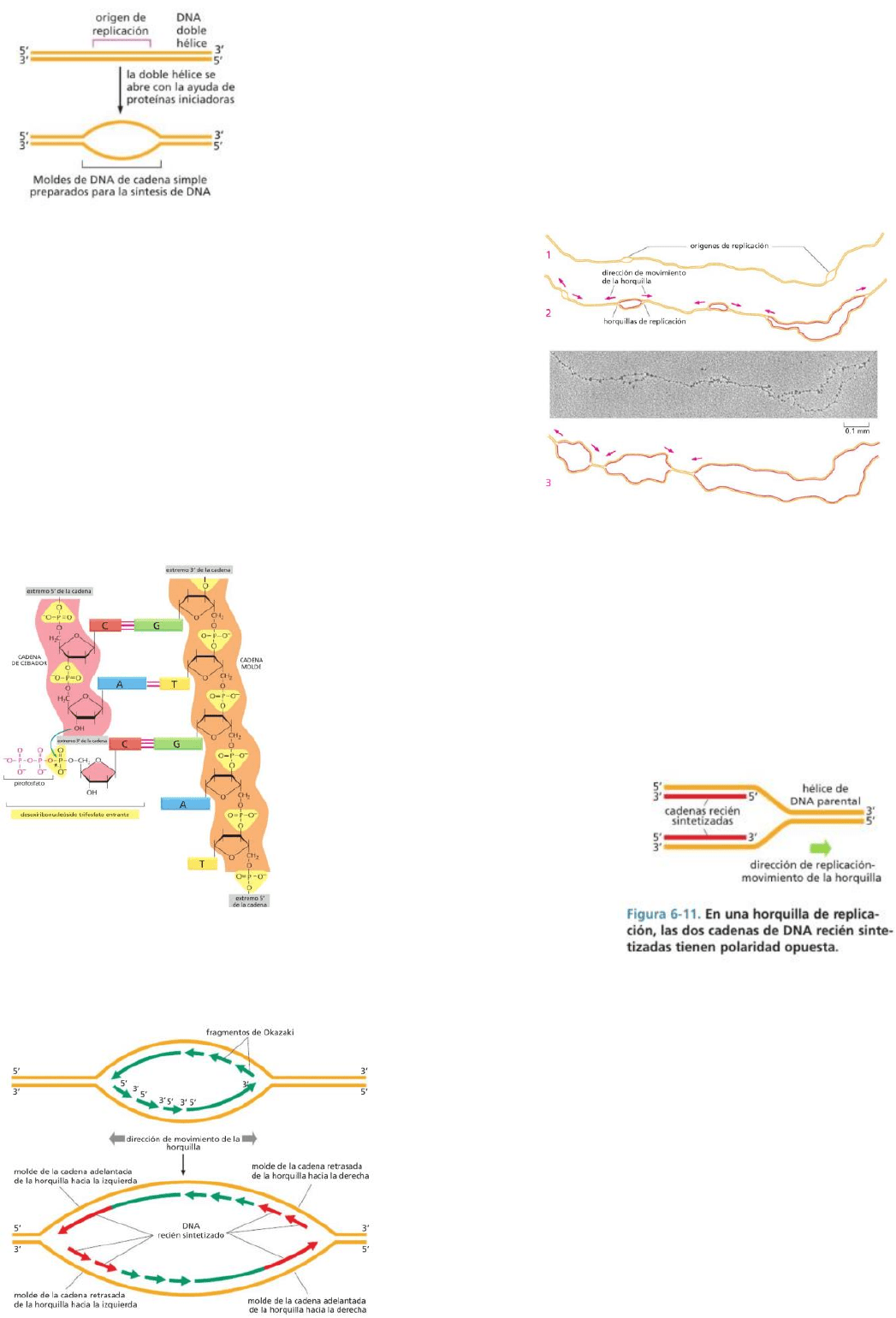
vez unidas las proteínas iniciadoras en el ADN, éstas atraen a un grupo de
proteínas variado que forman una maquinaria proteica de replicación.
El bucle es el espacio donde se encuentran dos horquillas de replicación
(cada Y vendría a ser una horquilla).
*El genoma humano tiene aprox 10mil orígenes. La replicación inicia en
muchos lugares a la vez, permitiendo la replicación con rapidez.
Las moléculas de ADN en proceso de replicación contienen
horquillas de replicación, uniones con forma de Y. En estas
horquillas, la maquinaria de replicación se mueve a lo largo del
ADN abriendo las dos cadenas de la hélice y utilizándolas como
molde para la síntesis de una nueva complementaria. Se
forman dos horquillas de replicación a partir de cada origen, y
se moverán desde allí en direcciones opuestas,
descomprimiendo el ADN a medida que avanzan. Por lo tanto,
la replicación es bidireccional.
En el centro de las maquinarias de replicación se encuentra
una ADN polimerasa, una enzima que sintetiza el nuevo ADN
a partir de la cadena molde. Se cataliza la adición de
nucleótidos al extremo 3’ de la cadena de ADN en crecimiento mediante enlaces fosfodiéster entre
ese extremo y el grupo fosfato 5’ del nucleótido entrante. Los nucleótidos ingresan en la reacción como
nucleósidos trifosfatos, que proporcionan la energía para la
polimerización. La hidrólisis de un enlace de alta energía en el nucleósido
trifosfato impulsa la reacción que une al monómero nucleótido a la
cadena y libera pirofosfato (PPi). El ADN polimerasa acopla la liberación
de esta energía a la reacción de polimerización. El pirofosfato es
hidrolizado después en fosfato inorgánico (Pi), tornando irreversible la
reacción de polimerización. El ADN polimerasa permanece asociado al
ADN durante varios ciclos de reacción de polimerización.
La horquilla de polimerización es
asimétrica, por lo que una nueva
cadena de ADN se forma sobre un
molde que se encuentra en dirección
3’ a 5’ y la otra en dirección opuesta
(5’ a 3’). Sin embargo, el ADN
polimerasa solo puede catalizar el
crecimiento de la cadena en una sola dirección: puede añadir
subunidades únicamente al extremo 3’ de la cadena, por lo que una
cadena solo puede ser sintetizada en dirección 5’ a 3’. Para sintetizar dos cadenas se realiza una maniobra
de pespunte: la cadena que debe crecer desde el extremo 5’ se sintetiza de manera discontinua, en
pequeñas partes sucesivas (fragmentos de Okazaki), y el
ADN polimerasa se mueve hacia atrás con respecto a la
dirección de la horquilla de replicación, a medida que cada
segmento nuevo es sintetizado en dirección 5’ a 3’.
Lee de 3’-5’ y sintetiza de 5’-3’.
Los fragmentos de Okazaki son unidos todos juntos por la
ligasa formando una cadena nueva continua.
Cadena retrasada ---> cadena sintetizada de forma
discontinua.
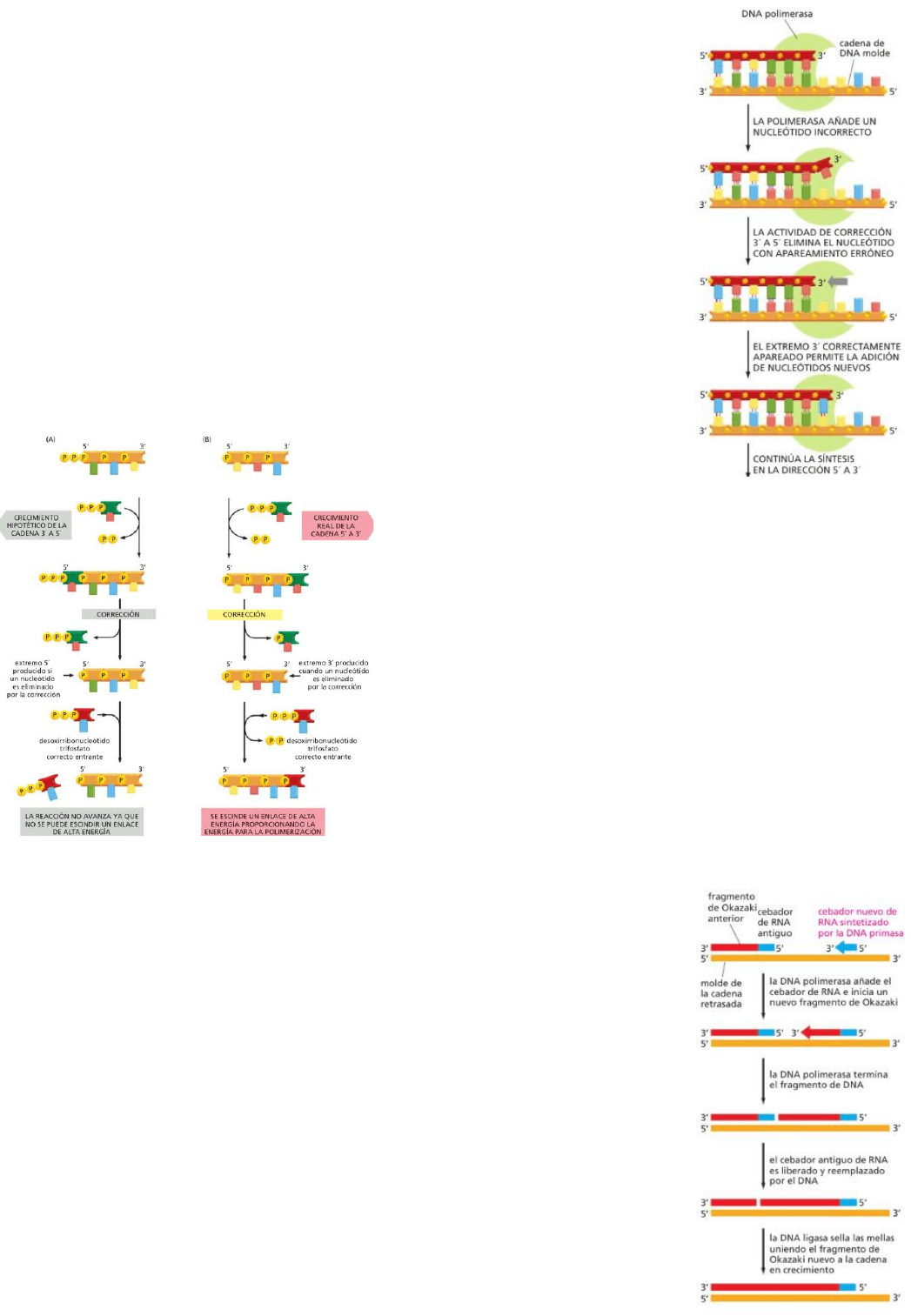
Cadena adelantada ---> cadena sintetizada de forma continua.
En cada horquilla hay una cadena retrasada y otra adelantada.
El ADN polimerasa comete un error cada 10
7
pares de nucleótidos que copia.
Tiene dos cualidades autocorrectoras:
1) La enzima monitoriza el apareamiento de bases entre cada nucleótido
entrante y la cadena molde, solo catalizando la reacción de adición
cuando el apareamiento es correcto.
2) La enzima, cuando comete un error, puede corregirlo mediante la acción
de corrección. Ésta se produce durante la síntesis. Verifica que el
nucleótido agregado esté apareado de manera correcta a la cadena
molde. Si es así, se continúa con la síntesis; de lo contrario, se elimina
el nucleótido mal apareado y se intenta de nuevo. El ADN polimerasa
tiene actividad de polimerización de 5’ a 3’ muy precisa, y de
corrección en 3’ a 5’. La corrección la realiza una nucleasa que corta el
eje fosfodiéster. La polimerización y la corrección son llevadas a cabo
por dos dominios distintos de la molécula polimerasa.
La dirección de polimerización 5’
a 3’ es vital para que exista el
mecanismo de corrección, ya que, si se realizara en otra
dirección, la corrección crearía un extremo químicamente
muerto que no puede ser alargado.
La enzima primasa (un ARN polimerasa) sintetiza el
cebador/primer de ARN, una cadena de ARN corta (de 10
nucleótidos) formada por bases apareadas a la cadena molde
que proporciona el extremo 3’ de la base apareada como
punto de partida para el ADN polimerasa (el cebador es
como un punto de apoyo para la polimerasa). Hay primer en
cada punto que el ADN polimerasa empieza a sintetizar, tanto
en cadenas adelantadas como retrasadas (en las adelantadas
hay solo uno, y en las retrasadas hay un primer en cada
fragmento de Okazaki).
*La primasa tiene una mayor tasa de error de sintetización
porque no tiene función reparadora. Sintetiza y no revisa si hubo error.
Sobre la cadena retrasada, el ADN se sintetiza en fragmentos. Cuando se
expone un nuevo tramo de la cadena, se sintetiza un cebador a intervalos de
200 nucleótidos sobre la cadena retrasada, y cada cebador tiene una longitud
aprox de 10 nucleótidos. El ADN polimerasa agrega un desoxirribonucleico en
el extremo 3’ de este cebador para comenzar con una cadena de ADN y
alargarla hasta encontrar al siguiente (antiguo) cebador.
Una nucleasa reconoce la cadena de ARN en una hélice ADN y la degrada,
dejando un espacio para un ADN polimerasa reparador que completa el lugar
con un fragmento de Okazaki adyacente. Finalmente, un ADN ligasa (enzima)
une el extremo 5’-fosfato del fragmento al extremo 3’-hidroxilo del siguiente,
formando una cadena continua.
La maquinaria de replicación está formada por:
ADN polimerasa ---> cataliza los enlaces entre los nucleótidos.
Primer/cebador ---> punto de apoyo para el ADN polimerasa (para que éste
pueda empezar la sintetización).
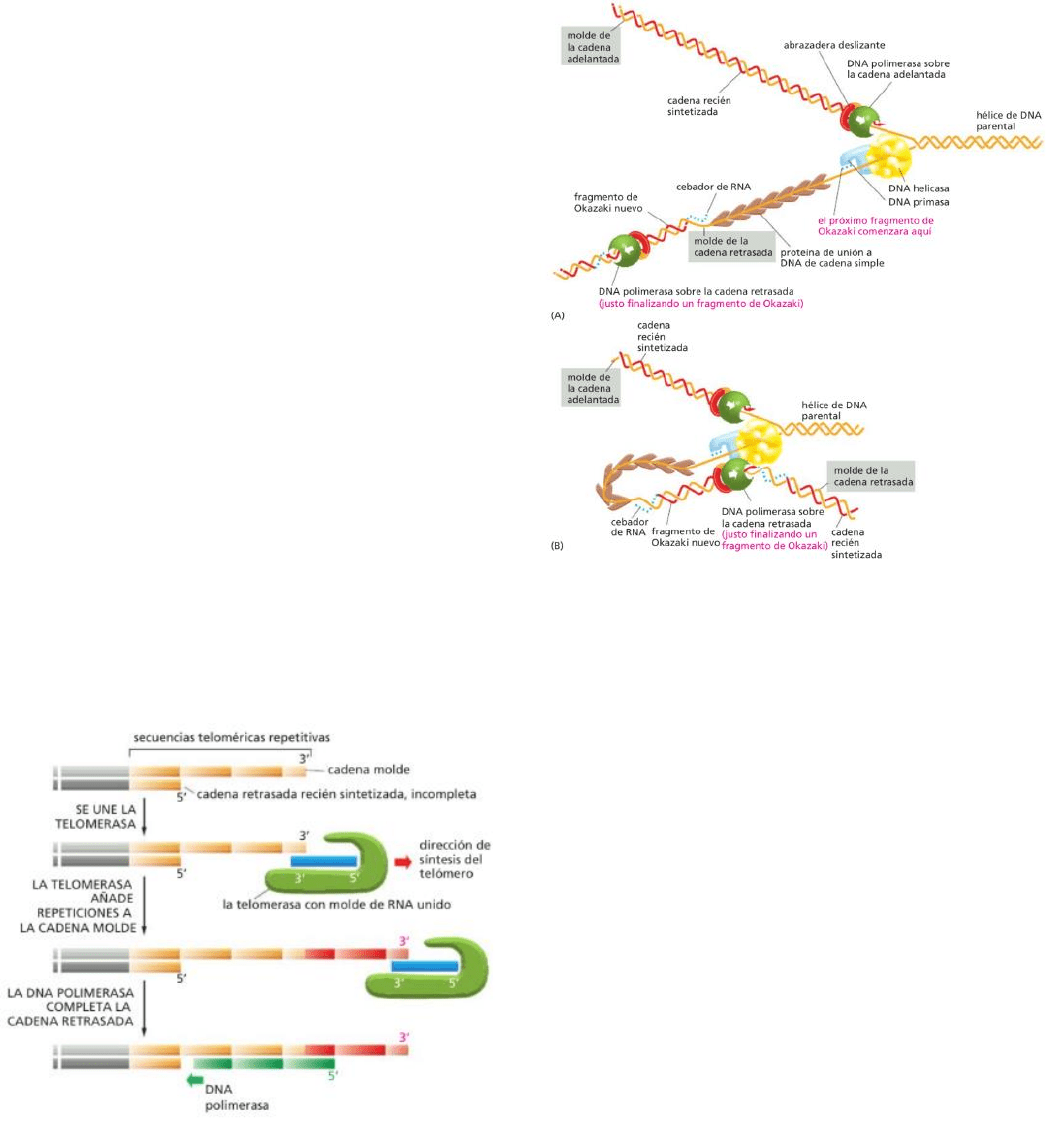
Helicasa ---> proteína que separa la doble hélice de
ADN utilizando energía de la hidrólisis de ATP,
rompiendo los enlaces de H.
Proteína de unión a cadena simple ---> se une a la
cadena simple de ADN expuesta por la helicasa, la
mantiene estirada (evitando reformación de pares de
bases) para que sirva como molde para el ADN
polimerasa.
Abrazadera deslizante ---> mantiene al ADN
polimerasa firmemente adherido al molde de ADN
mientras sintetiza las cadenas nuevas de ADN (forma
como una especie de anillo entre ambos manteniendo
todo ceñido).
Cargador de la abrazadera ---> hidroliza ATP cada
vez que asegura una abrazadera alrededor de ADN.
Esta carga solo ocurre una vez en una cadena
adelantada. En las cadenas retrasadas, la abrazadera
se disocia y se vuelve a adherir cada vez que se
sintetiza un fragmento de Okazaki.
*La mayoría de las proteínas involucradas en la replicación de ADN se mantinen unidas en un gran complejo
enzimático que se mueve como una unidad a lo largo del ADN, permitiendo que éste sea sintetizado en
ambas cadenas de forma coordinada.
Los telómeros permiten que se complete la
síntesis de ADN en los extremos. La telomerasa
(enzima) agrega una serie de repeticiones de una
secuencia de ADN en el extremo 3’ de la cadena
molde, permitiendo que la cadena retrasada sea
completada por el ADN polimerasa. (En seres
humanos la secuencia es GGGGTTA). La enzima
posee una secuencia de ARN complementaria a la
secuencia repetida de ADN. Este ARN actúa como
molde para la síntesis de ADN por la telomerasa.
Además, toda esta estructura de ADN repetido más
las regiones adyacentes a este son reconocidas como
los verdaderos extremos de los cromosomas.
Reparación del ADN
Mutación ---> cambio permanente en el ADN.
Las mutaciones pueden causar graves problemas de salud en un individuo si éste se produce en una
secuencia vital del ADN (afecta la estructura y actividad de una proteína).
*Ej: anemia falciforme. Mutación en la secuencia de nucleótidos que codifica a la hemoglobina. Es una
enfermedad hereditaria.
(Una mutación en las células germinales será transmitida a todas las células del cuerpo del organismo
multicelular que se forme a partir de ésta, incluyendo a las germinales de la siguiente generación. En células
somáticas, la mutación puede dar origen a células variantes que pueden crecer y dividirse
descontroladamente. En casos extremos, evoluciona en un cáncer. Una alta fidelidad en la replicación es
vital).
*(El organismo de un hijo con ADN mutado no va a reconocer a esas mutaciones como tal).
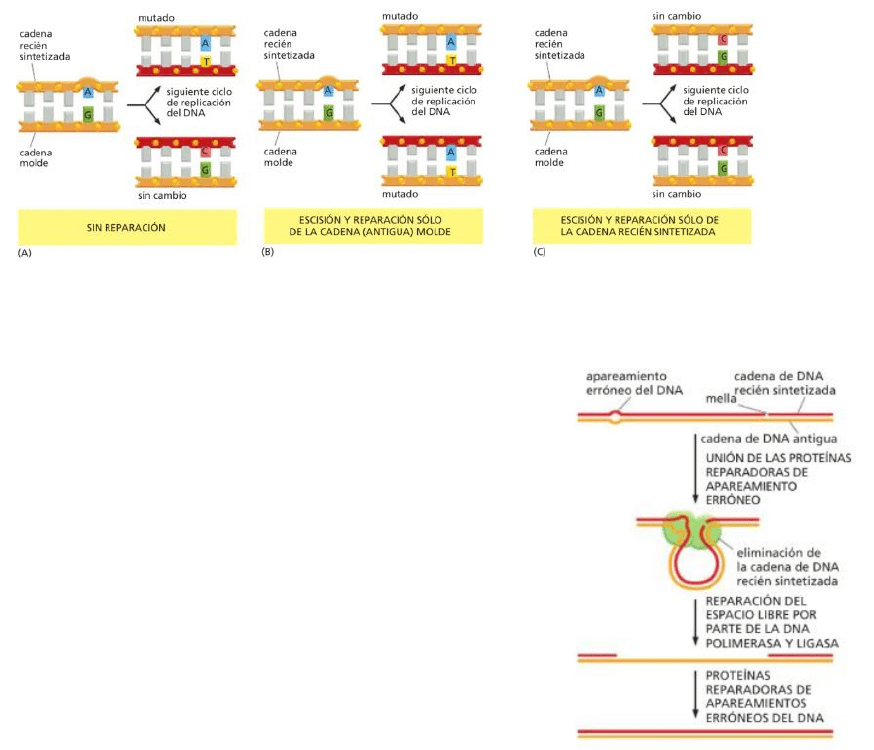
El sistema de reparación de apareamientos erróneos del ADN se dedica a corregir los (poco frecuentes)
errores.
*El mecanismo de replicación comete un error por cada 10
7
nucleótidos copiados. La reparación corrige el
99% y aumenta la precisión de error a 10
9
nucleótidos copiados.
Siempre que la maquinaria de replicación produce un error de copiado, deja por detrás un nucleótido mal
apareado (apareamiento erróneo).
-Si se queda sin corregir, la base mal apareada dará lugar a una mutación permanente en el próximo
ciclo de replicación del ADN (A)
-Un complejo de proteínas de reparación del apareamiento erróneo reconoce a estas bases mal
apareadas, elimina a una de las dos cadenas de ADN involucradas en el apareamiento erróneo y
resintetiza la cadena faltante.
-Si el error es “reparado” utilizando la cadena de ADN recién sintetizada como molde, ambas
moléculas de ADN producidas en el siguiente ciclo contendrán una mutación (B) (Además
que se “corrigió” la cadena molde para que no haya un error de apareamiento con la cadena
nueva, pero esto genera una mutación porque se cambia la información).
-Si el error es corregido utilizando la cadena molde (original) como modelo, se elimina
la posibilidad de una mutación (C). *El esquema C muestra el modo de reparación que
ocurre en las células.
Las proteínas de reparación de apareamientos incorrectos corrigen los errores que se producen
durante la replicación del ADN.
1. Un apareamiento erróneo del ADN distorsiona la
geometría de la doble hélice.
2. La distorsión es reconocida por proteínas reparadoras, las
cuales eliminan el ADN recién sintetizado.
3. El espacio libre en el ADN recién sintetizado es
reemplazado por un ADN polimerasa que corrige a media
que sintetiza y es sellada por el ADN ligasa.
*Una mella en el ADN recién sintetizado es la que le
permite distinguir a las proteínas reparadoras entre la
cadena nueva (con error) y la antigua. Las mellas
generalmente se producen en la cadena retrasada
(aunque también pueden presentarse en las adelantadas).
Solo permanecen pro un breve periodo, por lo que la
reparación debe ocurrir rápidamente. Las mellas son los
espacios que quedan entre los fragmentos de Okazaki antes de ser ligados entre sí. La
mella es la falta del enlace fosfodiester.
*En esta imagen, el proceso de replicación y reparación es simultáneo. Con la mella se
reconoce la cadena recién sintetizada, ya que con eso sabemos que los fragmentos aún no
fueron ligados (por lo que se sigue en el proceso de síntesis).
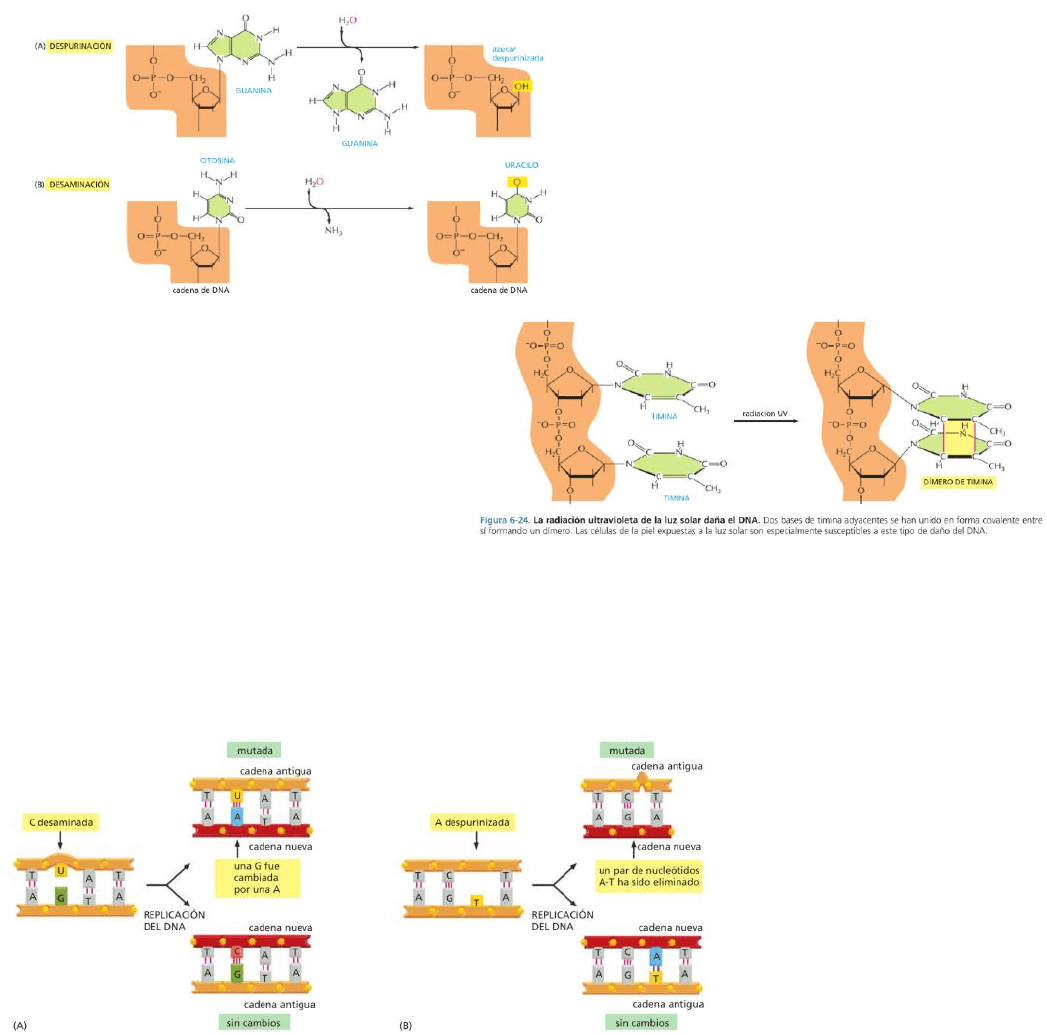
El ADN puede ser dañado de diversas formas.
Por colisiones térmicas con otras moléculas se pueden dar cambios químicos:
-Despurinización: pérdida de bases purina (A y
G). No se rompe el esqueleto fosfodiéster, sino
que origina lesiones similares a la pérdida de
piezas dentales.
-Desaminación: pérdida espontánea de un
grupo amino de la citosina en el ADN, lo que
produce una base uracilo (Una citosina se
convierte en uracilo).
La radiación ultravioleta de la luz solar
promueve enlaces covalentes entre dos bases
pirimidina adyacentes (Ej: dímero de timina),
dañando el ADN.
Si las modificaciones químicas de los nucleótidos no son reparadas, se producen mutaciones. (A)
La desaminación de la citosina, si no es corregida, da como resultado la sustitución de una base por otra
cuando el ADN es replicado (se inserta una adenina cuando se encuentra un uracilo en la cadena molde).
(B) Si no se corrige la despurinización, se puede llevar a la pérdida de un par de nucleótidos. Cuando la
maquinaria de replicación encuentra una purina perdida en la cadena molde puede saltar al siguiente
nucleótido y eliminar un nucleótido de la cadena recién sintetizada (y un par completo de la hélice de ADN).
En otros casos, puede completar
el espacio con un nucleótido
incorrecto, causando una
mutación.
La estabilidad de los genes depende de la reparación de ADN.
Los daños son reparados por una variedad de mecanismos catalizados por un grupo diferentes de enzimas.
Estos mecanismos dependen de la existencia de dos copias de info genética, una de cada cadena de la
doble hélice de ADN. Si una de las cadenas se daña accidentalmente, la info no se pierde forma irreparable
porque existe una “copia de seguridad” de la cadena alterada en la secuencia complementaria. La mayoría
de los daños crean estructuras que nunca se encontradas en un ADN sano (por lo que la mala se distingue
bien de la buena).
Reparación del ADN:
1) ESCISIÓN. El ADN dañado es reconocido y eliminado por una serie de nucleasas, que escinden
enlaces covalentes que unen los nucleótidos dañados al resto de la molécula de ADN y deja un
pequeño espacio en una de las cadenas de la doble hélice de ADN en esa región. *Este paso es
diferente en el tipo de enzima utilizada utilizadas según el tipo de daño que tenga el ADN.
2) NUEVA SÍNTESIS. Un ADN polimerasa de reparación se une al extremo 3’ hidroxilo y corta la cadena
de ADN. Después llena el espacio con nucleótidos complementarios a la cadena inalterada. Aunque
es una enzima diferente del ADN polimerasa que replica el ADN, la polimerasa de reparación sintetiza
las cadenas de ADN del mismo modo. Por ejemplo, sintetiza la cadena en la dirección 5’-3’ y tiene el
mismo tipo de actividad correctora que asegura que la cadena molde sea copiada con precisión. En
muchas células, ésta es la misma enzima que llena el espacio dejado después de que los ARN
cebadores son eliminados durante la replicación normal del ADN.
3) LIGACIÓN. Cuando la polimerasa que repara el ADN llenó el espacio, una rotura se mantiene en el
eje azúcar-fosfato de la cadena reparada. La mella es sellada por ADN ligasa, la misma que une los
fragmentos en la cadena retrasada durante la replicación. El sellado requiere energía que se obtiene
por hidrólisis de ATP, que vuelve a generar el enlace fosfodiéster roto entre nucleótidos adyacentes.
*En la desaminación de timina solo se reemplaza un nucleótido. En los dímeros de timina hace falta
eliminar un largo tramo de entre 10 y 20 nucleótidos de la cadena dañada.
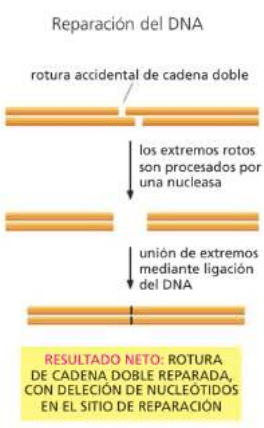
Un tipo de daño peligroso es el que se puede dar en ambas cadenas de la doble hélice al mismo tiempo, ya
que no deja un molde intacto que pueda usarse para la reparación (radiación ionizante, percances en la
horquilla de replicación, agentes oxidantes fuertes, metabolitos, entre otros).
Mecanismo de reparación de rotura bicatenaria ---> unión de extremos no
homólogos.
*Las células pueden usar unión de extremos no homólogos para reparar
roturas de doble cadena. Este mecanismo "rápido y sucio" altera la secuencia
de ADN original durante el proceso de reparación. Las alteraciones suelen ser
deleciones breves.
-Los extremos rotos son aproximados por un grupo de enzimas y vueltos
a unir por ligación del ADN (aunque se repara la ruptura, se suelen
perder nucleótidos en el sitio de reparación y, por lo tanto, alterar la
secuencia general de ADN).
*Apunte: se rompen las dos cadenas accidentalmente, y una nucleasa
elimina los nucleótidos de los extremos (pasando de extremos
puntiagudos a romos). Se unen los extremos no homólogos por ligasa,
pero se pierden nucleótidos porque no hay una resintetización de la info perdida.
Recombinación homóloga
Información genética proporcionada por un ADN bicatenario diferente participa en la reparación de
una doble cadena rota, sin sacrificar la información contenida en el sitio de deleción.
*Un ADN bicatenario intacto participa como molde para la reparación precisa de un ADN bicatenario
roto.
*La recombinación homóloga es el mismo proceso que ocurre en la meiosis, el intercambio de
información genética que genera cromosomas con secuencias de ADN nuevas (sin sentido
reparador).
La recombinación solo ocurre cuando hay regiones extensas de homología/similitud de secuencia. Un par
de moléculas de ADN evalúan la similitud realizando una muestra entre sí de sus secuencias nucleotídicas
cuando una cadena simple de ADN bicatenario se interconecta mediante apareamiento de bases con la
cadena complementaria de la otra molécula bicatenaria en un segmento extenso.
*La coincidencia no tiene que ser perfecta para que se produzca la recombinación homóloga, pero sí
muy similar.
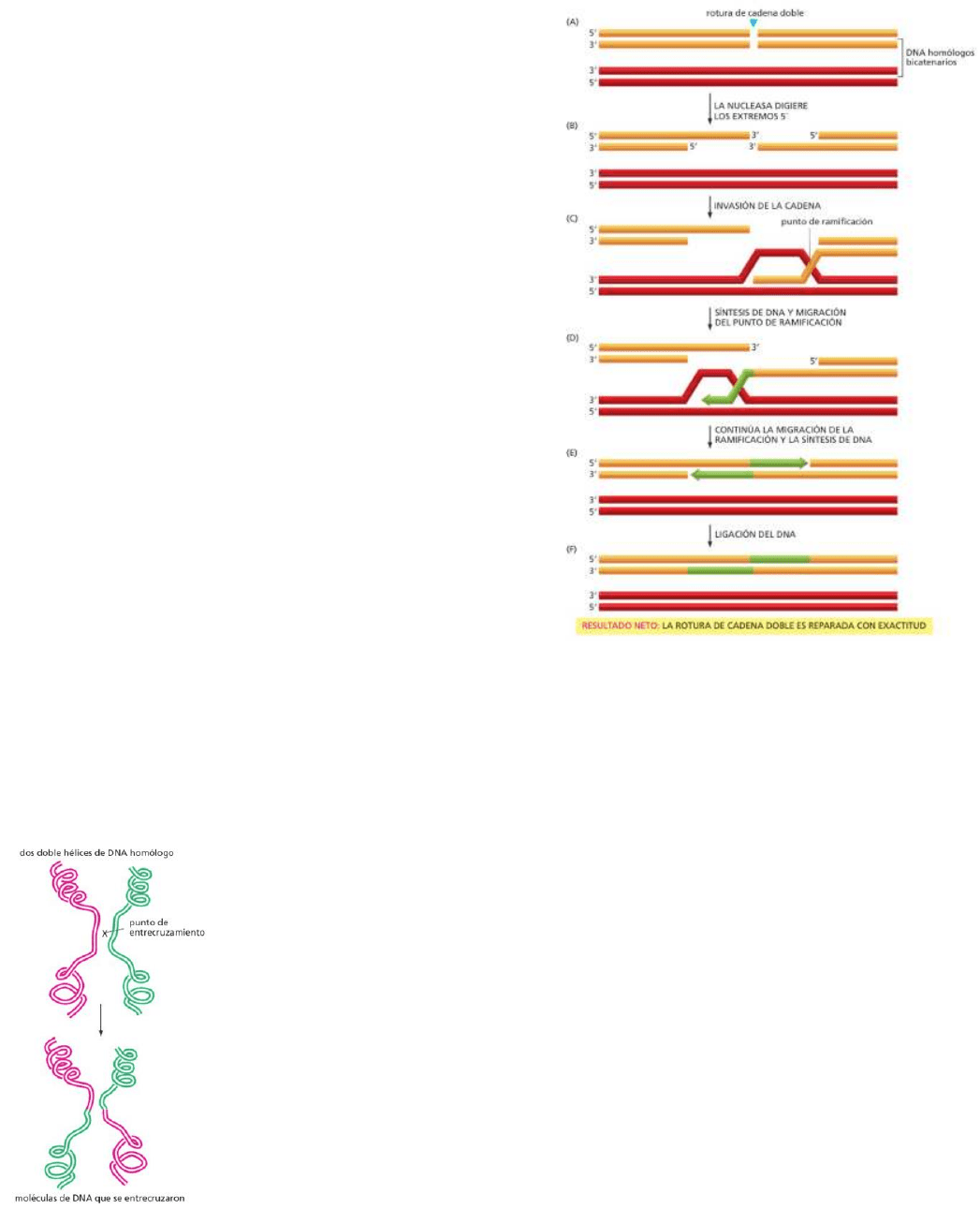
La recombinación usualmente inicia cuando se produce una ruptura en la doble hélice poco tiempo
después de que el ADN se haya replicado, con las hélices replicadas aún próximas entre sí, antes de
que la célula se divida.
A. Una nucleasa genera extremos de cadena simple en
la rotura al digerir hacia atrás una de las cadenas de
ADN complementarias.
B. Una de las cadenas simples “invade”, con ayuda de
enzimas especializadas, el ADN bicatenario
homólogo.
C. Por apareamiento de bases, se crea un punto de
ramificación donde las dos cadenas, una de cada
bicatenario, se cruzan.
D. Un ADN polimerasa reparador alarga la cadena
invasora utilizando la cadena complementaria como
molde.
E. El punto de ramificación “migra” a mediad que los
pares de bases mantienen juntos las brechas de las
cadenas dobles, y se forma una nueva a partir de
ellas.
F. Se completa la reparación por síntesis de ADN
adicional, seguida por la ligación de ADN.
*Como resultado se obtienen dos hélices de ADN
intactas, donde una se utilizó como molde para la
reparar a la otra.
(Notas de clase): RECOMBINACIÓN HOMÓLOGA
-Acá no se pierde info.
-El ADN polimerasa actúa llenando el espacio con un molde (se utilizan las cromátides hermanas).
Ocurre entre fase S y G2.
-Nucleasa genera una rotura que pueda ser detectada por la polimerasa.
La recombinación homóloga intercambia información genética durante la meiosis.
Se produce un entrecruzamiento, dos cromosomas homólogos (uno paterno y otro
materno) se reúnen y sufren intercambio genético. Esto genera combinaciones
nuevas de secuencias de ADN en cada cromosoma.
*El sitio de intercambio puede encontrarse en cualquier lugar de la secuencia de las
dos moléculas de ADN participantes.
*La escisión y reunión que median el intercambio son tan precisos que rara vez se
producen pérdidas o ganancias de nucleótidos.
*La recombinación homóloga tiene lugar entre moléculas de ADN con una
secuencia de nucleótidos similar. La rotura y nueva unión de dobles hélices de
ADN homólogas crean dos moléculas de ADN que se han entrecruzado. Aunque
las dos moléculas de ADN originales deben tener secuencias nucleotídicas
similares para entrecruzarse puede crear moléculas de ADN con una secuencia de
nucleótidos nueva.
A. Una enzima especializada corta ambas cadenas de uno de los cromosomas recombinantes, creando
una rotura en la cadena doble.
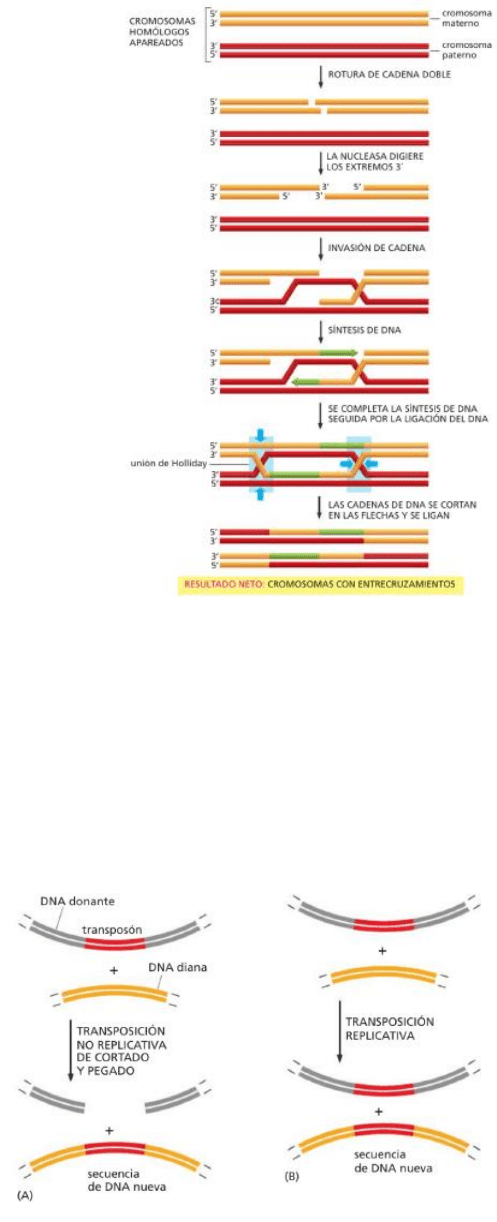
B. Proteínas de reparación convergen en el sitio de “daño”, esta
vez dirigidas por proteínas específicas de la meiosis,
produciendo, por medio de una o más moléculas
entrecruzadas, dos moléculas con secuencias de ADN
nuevas.
C. Se forman uniones de Holliday, sitios donde se producirá el
entrecruzamiento.
*En la meiosis la recombinación se produce de manera
diferencial entre cromos materno y paterno en lugar de
cadenas idénticas, asegurando que cada uno de los
cromosomas hijos tengan una combinación de secuencias
de ambos progenitores.
*La recombinación homóloga en la meiosis genera
entrecruzamientos. Una vez que las proteínas específicas de la
meiosis han cortado el ADN bicatenario y procesado los extremos,
la recombinación homóloga procede a través de la formación de dos
uniones de Holliday (recuadros azules), los sitios donde se cruzan
los ADN bicatenarios.
(Notas de clase): Esto es crossing over. ENTRECRUZAMIENTO. La
info pasa entre cromosómas homólogos, pero no tiene sentido de
reparación. (wow no vayas a iluminar el resumen con tu nota tan clave).
Elementos genéticos móviles y virus
Los elementos genéticos móviles son agentes que producen cambios genéticos drásticos, también
conocidos como genes saltarines. Son segmentos cortos de ADN que pueden moverse desde una posición
del genoma celular a otra, pudiéndose insertar por sí mismos en cualquier secuencia dentro del genoma.
*Un ejemplo drástico son los virus, ADN móviles que pueden escapar de una célula e infectar a otra.
Transposones ---> elementos genéticos móviles.
*Se clasifican de acuerdo con el mecanismo por
el cual se mueven o transportan (Ej:
transposones solo de ADN, de las bacterias,
que durante el movimiento permanece como
ADN sin convertirse en ARN).
*Los transposones de solo ADN se mueven
por dos tipos de mecanismos. (a) En la
transposición de cortar y pegar, el elemento se
corta del ADN donante y se inserta en el ADN
objetivo, dejando una molécula de ADN
donante rota. (El ADN donante se puede
reparar de diversas formas, pero esto a veces da como resultado deleciones o reordenamientos de
la molécula donante). (B) En el curso de la transposición replicativa, el elemento es copiado por la
replicación del ADN. los productos finales son una molécula que parece idéntica al donante original
y una molécula objetivo que tiene insertado un elemento genético móvil. En general, un tipo particular
de transposón se mueve por uno solo de estos mecanismos. Sin embargo, los dos mecanismos
tienen muchas similitudes enzimáticas y algunos transposones pueden moverse por cualquiera de
los dos mecanismos. los ADN donante y diana pueden ser parte de la misma molécula de ADN o
residir en moléculas diferentes.
Casi la mitad del genoma humano está constituido por elementos genéticos móviles. Muchos se
desplazan de un lugar a otro dentro del genoma utilizando un mecanismo de cortado y pegado (como el de
los transposones solo de ADN). Sin embargo, hay otro tipo exclusivo de los eucariontes:
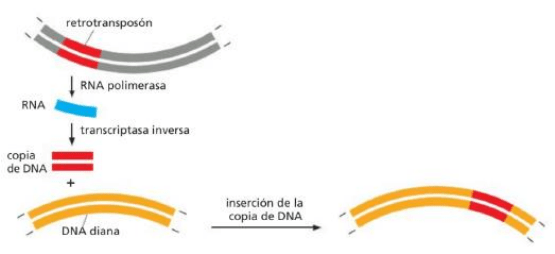
Retrotransposones ---> elementos génicos móviles que se desplazan mediante un ARN intermediario.
(Ej) El elemento L1 (o LINE-1) se transcribe a ARN mediante ARN polimerasa de la célula hospedadora.
Una copia de ADN de este ARN se sintetiza mediante la enzima transcriptasa inversa, que utiliza al ARN
como molde. Esta enzima está codificada por el elemento L1 en sí mismo. La copia de ADN del elemento
después se puede integrar a otro sitio del genoma.
*Los retrotransposones se mueven a través
de un intermediario de ARN. Estos elementos
transponibles se transcriben primero en un
intermedio de ARN. Luego, la enzima
transcriptasa inversa sintetiza una copia de ADN
de este ARN. A continuación, la copia de ADN
del transposón se inserta en la ubicación de
destino, que puede estar en la misma molécula
de ADN o en una diferente. El retrotransposón
donante permanece en su ubicación original, por
lo que cada vez que transpone, se duplica. estos elementos genéticos móviles se denominan
retrotransposones porque en una etapa de su transposición su información genética fluye hacia atrás, desde
el ARN hasta el ADN.

Replicación^J reparación y recombinación ADN - resumen.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.