
10
Capítulo
Patrones de la herencia
Flo Hyman, ganadora de la medalla de plata de voleibol en
los Juegos Olímpicos, fue víctima del síndrome de Marfan en la
cúspide de su carrera.
Estudio de caso
Muerte súbita en la cancha
FLO HYMAN, de más de 1.80 metros de estatura,
grácil y atlética, era una de las mejores voleibolistas
de todos los tiempos. Fue estrella del equipo
estadounidense de voleibol que obtuvo la medalla
de plata en las Olimpiadas de 1984 y a continuación
se contrató con un equipo profesional japonés. En
1986 salió de un partido por falta de aire y murió en
silencio sentada en la banca, a los 31 años. ¿Cómo
pudo ocurrirle esto a alguien tan joven y en tan buena
condición física?
Flo Hyman sufría un trastorno genético, el
síndrome de Marfan, que afecta a una de cada 5 mil
a 10 mil personas, entre ellas, el presidente de
Estados Unidos Abraham Lincoln, el pianista Sergei
Rachmaninoff y, posiblemente, el faraón egipcio
Akenatón. Quienes sufren el síndrome de Marfan son
altos y esbeltos, con extremidades inusitadamente
largas y manos y pies grandes. A algunas personas
con el síndrome, estas características les reportan
fama y fortuna. Por desgracia, el síndrome de Marfan
también puede ser letal.
Hyman murió por ruptura de la aorta, la gran
arteria que lleva sangre del corazón a la mayor parte
del cuerpo. ¿Por qué se rompió la aorta de Hyman?
¿Qué tienen en común una aorta débil y la estatura
elevada y las manos grandes? El síndrome de Marfan es
causado por una mutación en el gen que codifica una
proteína llamada fibrilina, un componente esencial del
tejido conectivo. Muchas partes del cuerpo contienen
este tipo de tejido, como los tendones que unen los
músculos a los huesos, los ligamentos (por ejemplo, las
fibras que mantienen fijo el cristalino del ojo) y las
fuertes paredes de las arterias. La fibrilina forma
largas fibras que confieren fuerza al tejido conectivo.
La fibrilina normal también “atrapa” ciertos factores de
crecimiento y evita que estimulen excesivamente la
división de, por ejemplo, las células que forman hueso
(véase la página 158). La fibrilina defectuosa no capta
estos factores de crecimiento, con el resultado de que
brazos, piernas, manos y pies de las personas con el
síndrome de Marfan son notablemente grandes. La
combinación de fibrilina defectuosa y concentraciones
elevadas de los factores de crecimiento debilita también
los huesos, cartílagos y paredes de las arterias.
Como dijimos en la página 158, los organismos
diploides, incluidos los seres humanos, tienen
dos copias de cada gen, uno en cada cromosoma
homólogo. Una copia defectuosa del gen de la
fibrilina basta para causar el síndrome de Marfan.
¿Qué nos revela esto sobre la herencia del síndrome?
¿Todas las enfermedades hereditarias son causadas
por una única copia defectuosa de un gen? Para
averiguarlo, tenemos que remontarnos en el tiempo
hasta un monasterio de Moravia y visitar el jardín de
Gregorio Mendel.

174 UNIDAD 2 Herencia
Estudio de caso Muerte súbita en la cancha
10.1 ¿Cuál es la base física de la herencia?
Los genes son secuencias de nucleótidos en lugares
específicos de los cromosomas
Los dos alelos de un organismo pueden ser iguales
o diferentes
10.2 ¿Cómo se descubrieron los principios
de la herencia?
Hacer bien las cosas: los secretos del éxito de Mendel
10.3 ¿Cómo se heredan los rasgos únicos?
La herencia de alelos dominantes y recesivos en los
cromosomas homólogos explica los resultados de las cruzas
de Mendel
Un “registro genético” simple puede predecir genotipos
y fenotipos de las células hijas
La hipótesis de Mendel puede usarse para predecir el
resultado de nuevos tipos de cruzas de rasgos únicos
Estudio de caso continuación Muerte súbita
en la cancha
10.4 ¿Cómo se heredan los rasgos múltiples?
Mendel postuló que los rasgos se heredan de forma
independiente
En un mundo que no está preparado para ellos, los genios
pueden pasar inadvertidos
10.5 ¿Cómo se heredan los genes situados
en el mismo cromosoma?
Los genes del mismo cromosoma tienden a heredarse juntos
Los cruzamientos producen nuevas combinaciones de alelos
ligados
10.6 ¿Cómo se determina genéticamente el sexo?
10.7 ¿Cómo se heredan los genes ligados
a los cromosomas sexuales?
De un vistazo
Los genes ligados a los cromosomas sexuales se
encuentran sólo en el cromosoma X o sólo en el
cromosoma Y
10.8 ¿Las leyes de la herencia de Mendel
se aplican en todos los rasgos?
Dominancia incompleta: el fenotipo de los heterocigotos
es intermediario entre los fenotipos de los homocigotos
Un gen único puede tener múltiples alelos
Muchos rasgos están influidos por varios genes
Genes únicos tienen múltiples efectos en un fenotipo
El ambiente influye en la expresión de los genes
Estudio de caso continuación Muerte súbita
en la cancha
10.9 ¿Cómo se investigan los trastornos
genéticos humanos?
10.10 ¿Cómo se heredan los trastornos
genéticos humanos originados por genes únicos?
Algunos trastornos genéticos humanos son causados
por alelos recesivos
Algunos trastornos genéticos humanos son causados
por alelos dominantes
Algunos trastornos genéticos están ligados a los
cromosomas sexuales
10.11 ¿Cómo afectan a los seres humanos
los errores en el número de cromosomas?
Algunos trastornos genéticos son causados por números
anormales de cromosomas sexuales
Guardián de la salud Distrofia muscular
Algunos trastornos genéticos son causados por números
anormales de autosomas
Estudio de caso otro vistazo Muerte súbita
en la cancha
10.1
¿CUÁL ES LA BASE FÍSICA
DE LA HERENCIA?
La herencia es el proceso por el cual se transmiten las característi-
cas de los organismos a su descendencia. Nuestra exploración de la
herencia comenzará con un repaso breve de los genes y cromoso-
mas que conforman la base física de la herencia. En este capítulo
limitaremos nuestra exposición a los organismos diploides (la ma-
yoría de las plantas y los animales) que se reproducen sexualmente
por fusión de gametos haploides.
Los genes son secuencias de nucleótidos en lugares
específicos de los cromosomas
Un cromosoma consta de una única doble hélice de ADN empa-
quetada con diversas proteínas. Los segmentos del ADN, con una
longitud que va de pocos cientos a muchos miles de nucleótidos,
son las unidades de la herencia, los genes, que codifican la in-
formación necesaria para producir proteínas, células y organis-
mos enteros. Por tanto, los genes son parte de los cromosomas.
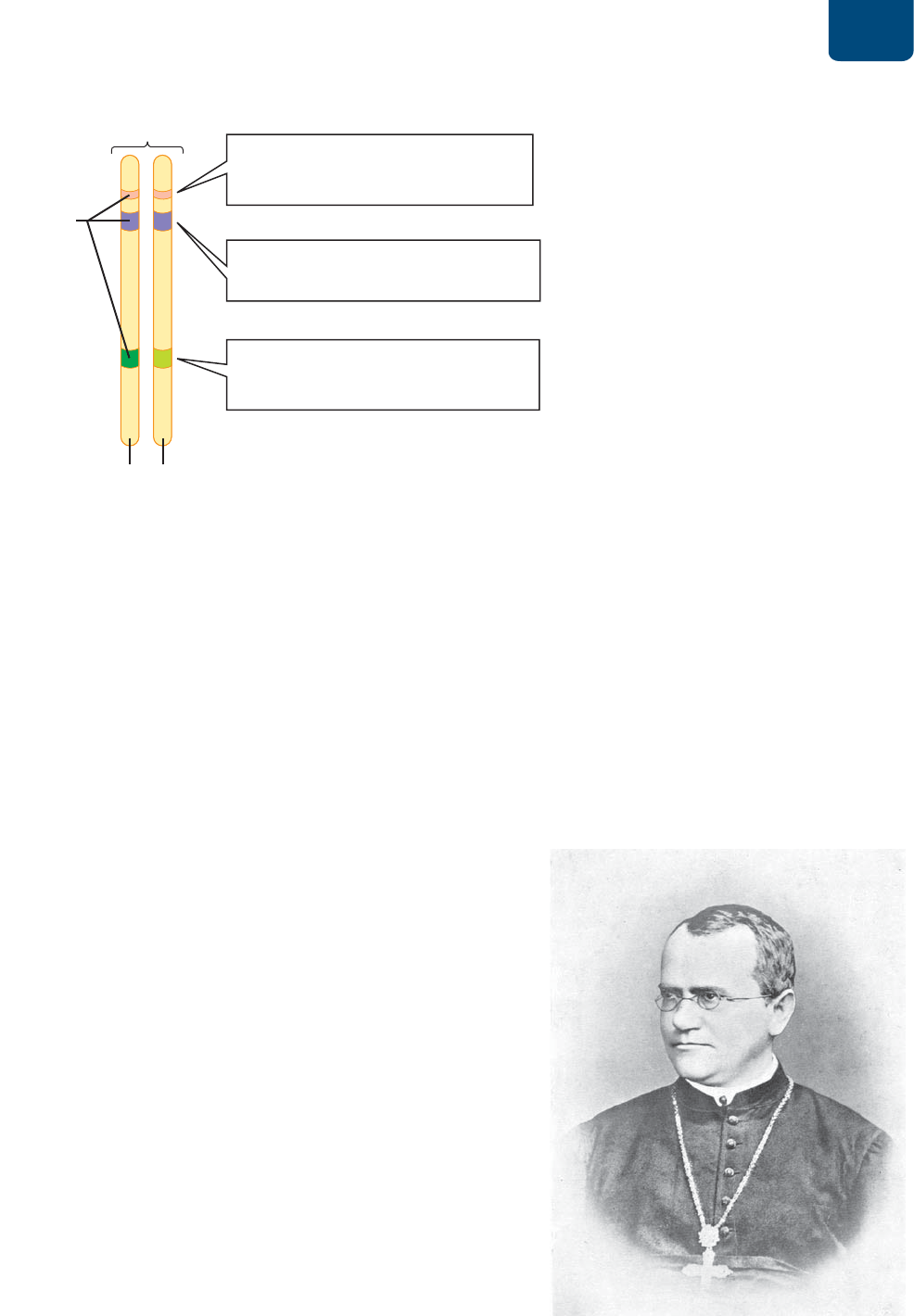
La ubicación física de un gen en un cromosoma se llama locus
(plural loci). Los cromosomas de organismos diploides se pre-
sentan en pares llamados homólogos. Los dos miembros de un par
de homólogos llevan los mismos genes, situados en los mismos
locus (
FIGURA 10-1). Sin embargo, las secuencias de nucleótidos
de un gen dado pueden variar con los miembros de una especie
o incluso en los dos homólogos del mismo organismo. Estas ver-
siones diferentes de un gen en un locus se llaman alelos (véase
la figura 10-1).
Las mutaciones son el origen de los alelos
Piensa en los genes como en frases muy largas, escritas con un al-
fabeto de nucleótidos en lugar de letras. Los alelos surgen como
mutaciones que cambian ligeramente la escritura de estas frases de
nucleótidos. Si ocurre una mutación en las células que se convier-
Patrones de la herencia Capítulo 10 175
ten en óvulos o espermatozoides, pueden pasar del progenitor a su
descendencia.
Casi todos los alelos del ADN de un organismo aparecieron
como mutaciones de las células reproductoras de los antepasados
de dicho organismo, quizá hace cientos o aun millones de años,
y desde entonces se han heredado de una generación a otra. Al-
gunos alelos, que llamaremos “mutaciones nuevas”, pueden ha-
ber surgido en las células reproductoras de los propios padres del
organismo.
Los dos alelos de un organismo
pueden ser iguales o diferentes
Como un organismo diploide tiene pares de cromosomas homó-
logos y los dos miembros del par contienen los mismos locus para
los genes, el organismo tiene dos copias de cada gen. Si los dos
homólogos tienen el mismo alelo en el locus de un gen, se dice que
el organismo es homocigoto para ese locus (“homocigoto” viene
de las palabras griegas que significan “mismo par”). Por ejemplo,
los cromosomas mostrados en la figura 10-1 son homocigotos en
dos locus. Si dos cromosomas homólogos tienen alelos diferentes
en un locus, el organismo es heterocigoto (“diferente par”) en ese
locus. Los cromosomas de la figura 10-1 son heterocigotos en
un locus. Los organismos que son heterocigotos en un locus parti-
cular se llaman híbridos.
10.2
¿CÓMO SE DESCUBRIERON
LOS PRINCIPIOS DE LA HERENCIA?
El esquema común de la herencia y muchos hechos básicos de los
genes y los alelos, así como de la distribución de los alelos en ga-
metos y cigotos durante la reproducción sexual fueron descubiertos
por un monje austriaco llamado Gregorio Mendel (
FIGURA 10-2)
a mediados del siglo XIX, mucho antes de que se descubrieran el
ADN, los cromosomas o la meiosis. Como sus experimentos son
ejemplos concisos y célebres de la práctica de las ciencias, vamos a
seguir la ruta de los descubrimientos de Mendel.
Hacer bien las cosas: los secretos
del éxito de Mendel
Hay tres pasos clave de todo experimento biológico exitoso: esco-
ger el organismo correcto para trabajar, diseñar y ejecutar bien el
experimento, y analizar adecuadamente los datos. Mendel fue
el primer genetista que aplicó los tres pasos.
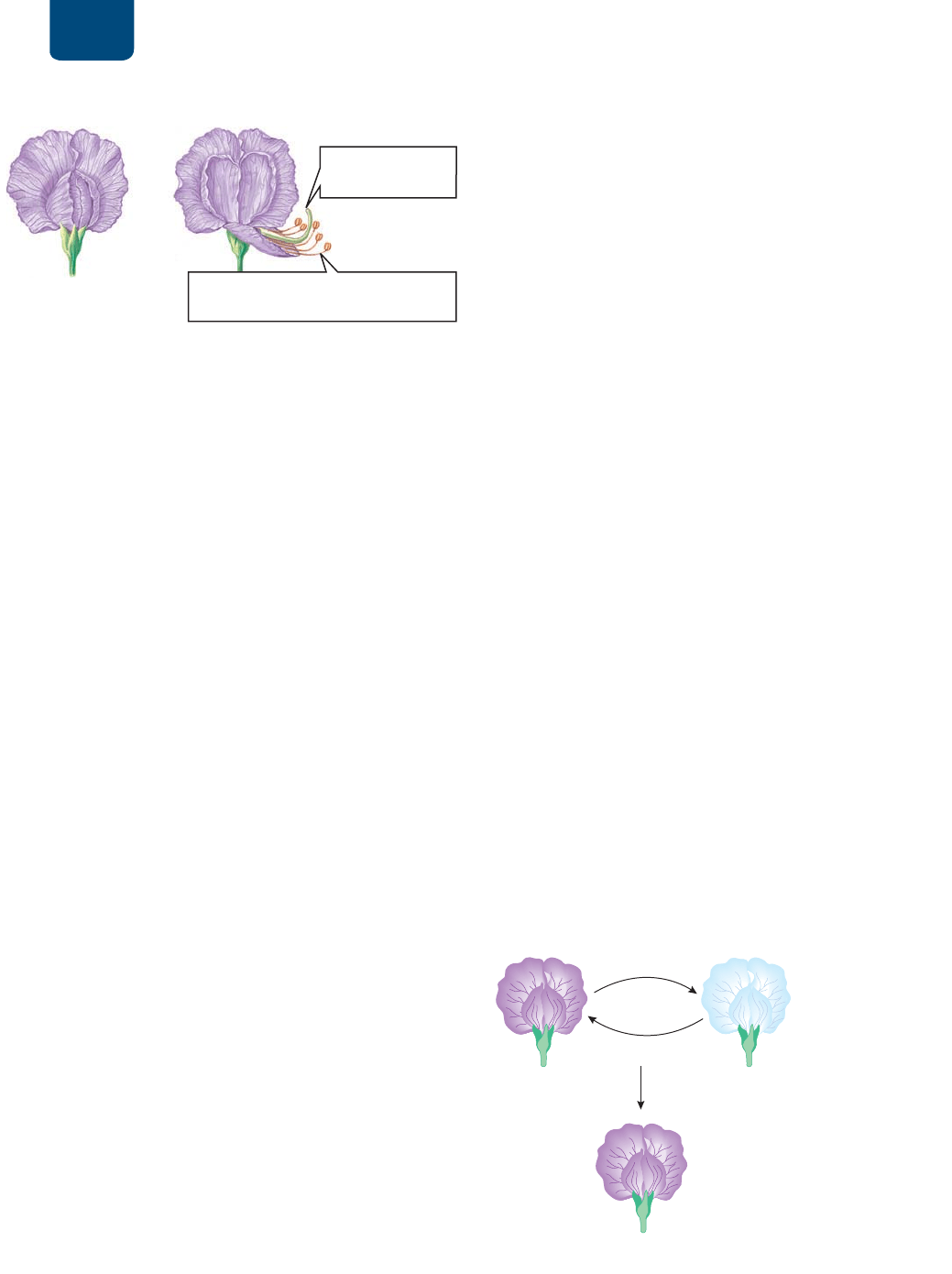
Mendel eligió la planta de chícharo comestible como sujeto
de sus experimentos sobre la herencia (
FIGURA 10-3). El estam-
bre, que es el órgano reproductivo de la flor, produce polen. Cada
grano de polen contiene espermatozoides. Por la polinización, el
espermatozoide fecunda el gameto femenino, el óvulo, situado
dentro del ovario, en la base del órgano reproductivo femenino,
$PERVFURPRVRPDVOOHYDQHOPLVPRDOHOR
GHOJHQHQHVWHORFXV(ORUJDQLVPRHV
KRPRFLJRWRHQHVWHORFXV
(VWHORFXVFRQWLHQHRWURJHQSDUDHOFXDO
HORUJDQLVPRHVKRPRFLJRWR
&DGDFURPRVRPDOOHYDXQDOHORGLIHUHQWH
GHHVWHJHQDVķTXHHORUJDQLVPRHV
KHWHURFLJRWRHQHVWHORFXV
SDUGH
FURPRVRPDV
KRPʼnORJRV
ORFLGHORV
JHQHV
HOFURPRVRPD
GHOSDGUH
HOFURPRVRPD
GHODPDGUH
FIGURA 10-1 Relaciones entre los genes, alelos y
cromosomas Cada cromosoma homólogo lleva el mismo
juego de genes. Cada gen se encuentra situado en la misma
posición, o locus, de su cromosoma. Las diferencias en
las secuencias de nucleótidos en el mismo locus producen
diferentes alelos del gen. Los organismos diploides tienen
dos alelos de cada gen, uno en cada homólogo. Los alelos
de los dos homólogos pueden ser iguales o diferentes.
FIGURA 10-2 Gregorio Mendel
176 UNIDAD 2 Herencia
que se llama carpelo. En las flores de la planta de chícharo, los
pétalos encierran las partes reproductoras, con lo que evitan que
entre el polen de otra flor (véase la figura 10-3). Así, los óvulos de
una flor de chícharo deben ser fecundados por espermatozoides
del polen de la misma flor. Se llama autopolinización al proceso
en el cual el espermatozoide de un organismo fecunda su propio
óvulo.
Ahora bien, Mendel quiso cruzar muchas veces dos plantas
de chícharo para ver qué descendencia producían. Abría una flor
y retiraba los estambres, para impedir la autopolinización. Luego,
espolvoreaba la punta pegajosa del carpelo con polen de la flor de
otra planta. Cuando los espermatozoides de un organismo fecun-
dan los óvulos de otro se llama fecundación cruzada.
El diseño experimental de Mendel era sencillo, pero bri-
llante. Los investigadores anteriores habían tratado de estudiar
la herencia considerando simultáneamente todos los elementos
de los organismos, incluso los que variaban poco entre unos y
otros. No es de sorprender que terminaran más confundidos que
esclarecidos. En cambio, Mendel decidió estudiar características
individuales (llamadas rasgos) que tenían formas diferentes sin
lugar a dudas, como flores blancas o moradas. Se concentró en el
estudio de un rasgo único cada vez.
Mendel siguió la herencia de estos rasgos durante varias
generaciones, contando el número de descendientes con cada
rasgo. Al analizar estas cifras, se le esclareció el esquema gene-
ral de la herencia. En la actualidad, cuantificar los resultados de
los experimentos y aplicar análisis estadísticos son herramientas
esenciales en prácticamente todos los campos de la biología, pero
en la época de Mendel, el análisis numérico era una novedad.
10.3
¿CÓMO SE HEREDAN
LOS RASGOS ÚNICOS?
Para estudiar la herencia, un investigador tiene que comenzar con
organismos que posean rasgos fáciles de identificar y que se trans-
mitan constantemente entre generaciones. Los organismos se lla-
man de raza pura cuando poseen un rasgo específico, como flores
moradas, que siempre heredan sin cambios todos los descendien-
tes producidos por autopolinización. Ya en esa época de mediados
del siglo XIX, los vendedores de semillas comercializaban varios
tipos de chícharos que eran de raza pura con formas distintas de
un solo rasgo. En sus primeros experimentos, Mendel realizó una
fecundación cruzada de plantas que eran de raza pura de diferentes
formas del mismo rasgo, como el color de la flor. Tomó las semillas
producidas y las cultivó el año siguiente para determinar los rasgos
de los descendientes.
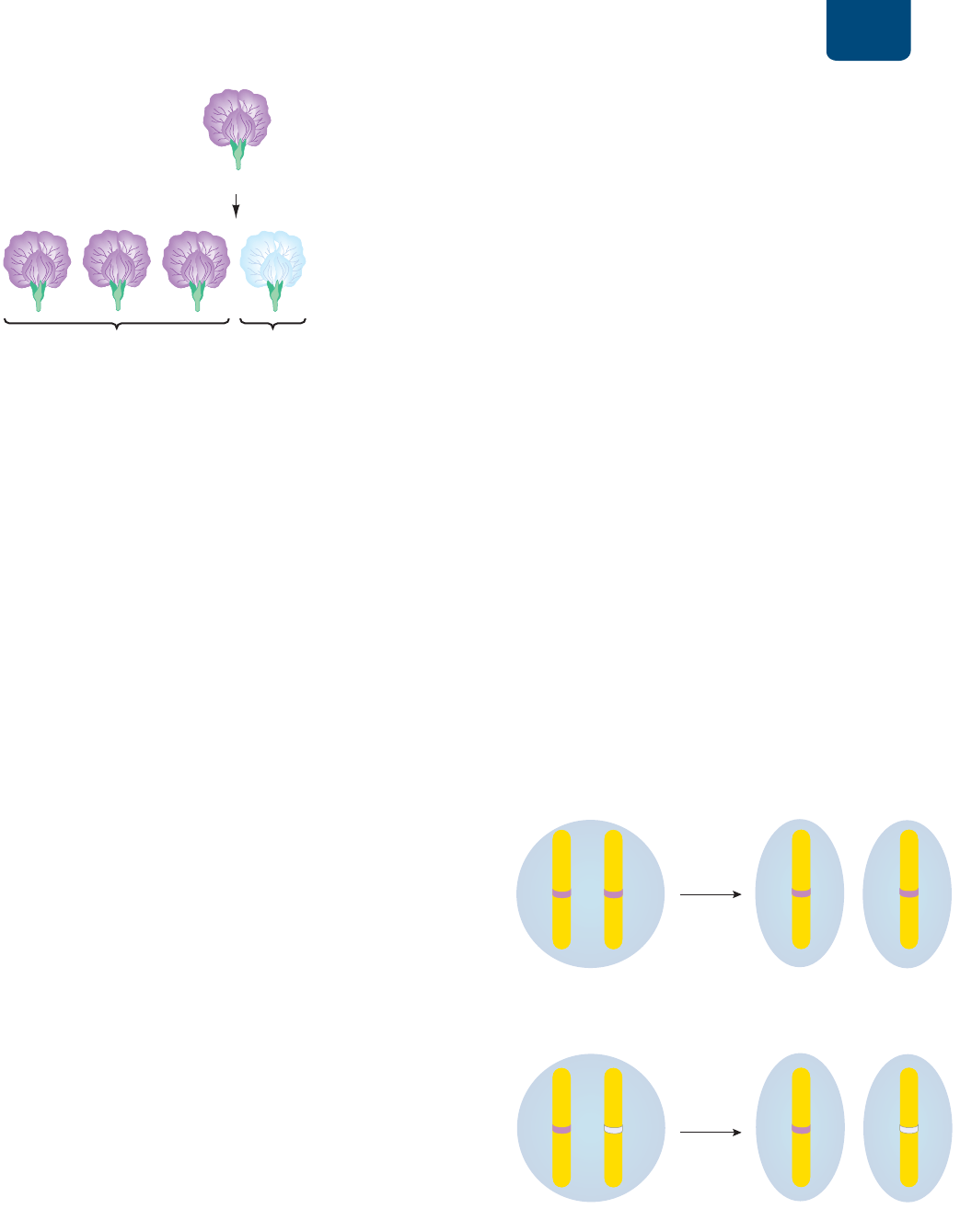
En uno de esos experimentos, Mendel realizó una fecun-
dación cruzada de plantas con flores blancas y plantas con flores
moradas, ambas de raza pura. Ésta fue la generación parental,
denotada con la letra P. Cuando cultivó las semillas producidas,
encontró que todos los descendientes de la primera generación
(la primera generación filial, F
1
) producían flores moradas (FI-
GURA 10-4
). ¿Qué le había pasado al color blanco? Las flores de
los híbridos F
1
eran tan moradas como las de sus padres. El color
blanco había desaparecido de la generación F
1
.
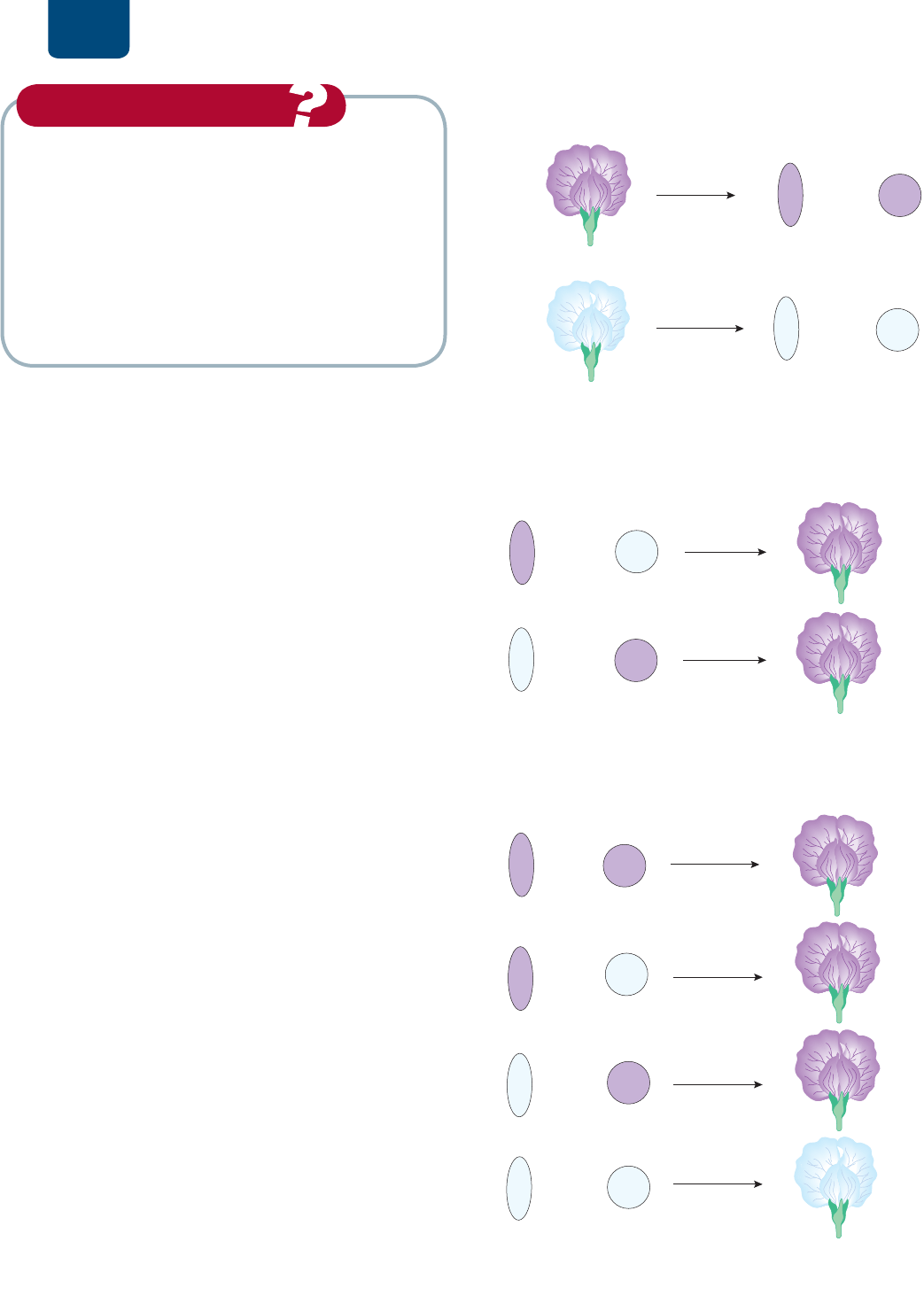
Entonces, Mendel dejó que las flores de las plantas F
1
se
autopolinizaran, recogió las semillas y las plantó la siguiente
primavera. En la segunda generación filial, F
2
, Mendel contó
705 plantas con flores moradas y 224 plantas con flores blancas.
Estas cifras son, aproximadamente, tres cuartas partes de flores
moradas y una cuarta parte de flores blancas, es decir, una pro-
porción de tres moradas por una blanca (
FIGURA 10-5). Este
resultado mostró que la capacidad de producir flores blancas
no desapareció de las plantas F
1
, sino que simplemente se había
“ocultado”.
Mendel dejó que las plantas F
2
se autopolinizaran y produ-
jo una generación más, F
3
. Vio que todas las plantas F
2
de flores
blancas dieron una descendencia de flores blancas, es decir, eran
de raza pura, pues en todas las generaciones que tuvo el tiempo y
la paciencia de cultivar, las plantas de flores blancas siempre die-
ron descendientes de flores blancas. Por el contrario, cuando las
plantas F
2
de flores moradas se autopolinizaron, su descendencia
fue de dos tipos. Alrededor de un tercio fueron plantas de flores
moradas de raza pura, pero los otros dos tercios fueron híbridos
que daban descendencia de flores blancas y moradas, de nueva
cuenta en la proporción de tres moradas por una blanca. Por tan-
to, la generación F
2
comprendía una cuarta parte de plantas de
raza pura para las flores moradas, una mitad de híbridas moradas
y una cuarta parte de raza pura para las flores blancas.
IORUGHFKķFKDURLQWDFWD IORUGLVHFFLRQDGDSDUDPRVWUDU
VXVHVWUXFWXUDVUHSURGXFWRUDV
&DUSHORIHPHQLQR
SURGXFHʼnYXORV
(VWDPEUHPDVFXOLQRSURGXFH
SROHQTXHFRQWLHQHHVSHUPDWR]RLGHV
FIGURA 10-3 Flores del chícharo comestible En la flor
del chícharo intacta (izquierda), los pétalos inferiores forman un
receptáculo que resguarda los órganos reproductores: el estambre
(masculino) y el carpelo (femenino). No puede entrar polen de
fuera en la planta, así que los chícharos se polinizan ellos mismos,
es decir, se autopolinizan. Si la flor se abre (derecha), puede ser
sometida manualmente a polinización cruzada.
SODQWDGHIORUHV
PRUDGDV
GHUD]DSXUD
SODQWDGHIORUHV
EODQFDV
GHUD]DSXUD
IHFXQGDFLʼnQFUX]DGD
SROHQ
SROHQ
WRGDVODVSODQWDVGH
IORUHVPRUDGDV
.LULYHJP}UKL
SVZWHKYLZ7
+LZJLUKPLU[LZKL
SHWYPTLYH
NLULYHJP}UMPSPHS-
FIGURA 10-4 Cruza de plantas de chícharo de raza pura
para flores blancas o moradas Toda la descendencia da flores
moradas.
Patrones de la herencia Capítulo 10 177
La herencia de alelos dominantes y recesivos
en los cromosomas homólogos explica
los resultados de las cruzas de Mendel
Los resultados de Mendel, completados por los conocimientos mo-
dernos sobre los genes y los cromosomas homólogos, nos permi-
ten postular una hipótesis en cinco partes para explicar la herencia
de rasgos únicos.
• Cada rasgo está determinado por pares de unidades físicas
individuales llamadas genes. Cada organismo tiene dos alelos
para cada gen, uno en cada cromosoma homólogo. Las
plantas de chícharos con flores blancas de raza pura tienen
diferentes alelos del gen del color de las flores de las plantas
de chícharos con flores moradas de raza pura.
• Cuando hay dos alelos diferentes en un organismo, uno (el
alelo dominante) puede enmascarar la expresión del otro
(el alelo recesivo); sin embargo, el alelo recesivo sigue presen-
te. En el chícharo comestible, el alelo de las flores moradas es
el dominante y el alelo de las flores blancas, el recesivo.
• Los pares de alelos de los cromosomas homólogos se separan
o se segregan unos de otros en la meiosis. Esta conclusión
se conoce como la ley de la segregación de Mendel. Como
resultado, cada gameto recibe sólo un alelo de cada par [a
diferencia de los animales, en las plantas los gametos no se
forman inmediatamente después de la meiosis (véanse las fi-
guras 9-17c y 9-20); sin embargo, más adelante en el ciclo, la
planta produce gametos que contienen un alelo de cada par].
Cuando un espermatozoide fecunda un óvulo, el descendien-
te recibe un alelo del padre (en el espermatozoide) y uno de
la madre (en el óvulo).
• La casualidad determina qué alelo se encuentra en un ga-
meto. Como los cromosomas homólogos se separan al azar
en la meiosis, la distribución de los alelos a los gametos es
también aleatoria.
• Los organismos de raza pura tienen dos copias del mismo
alelo para un gen dado y, por tanto, son homocigotos para ese
gen. Todos los gametos de un individuo homocigoto reciben
el mismo alelo para ese gen (
FIGURA 10-6a). Los organismos
híbridos tienen dos alelos para un gen y, por consiguiente, son
heterocigotos para ese gen. La mitad de los gametos heteroci-
gotos contiene un alelo para ese gen y la otra mitad contiene el
otro alelo (
FIGURA 10-6b).
Veamos cómo esta hipótesis explica los resultados de los
experimentos de Mendel con los colores de las flores (
FIGURA
10-7
). Si representamos con letras los alelos, asignemos la letra P
mayúscula al alelo dominante para el color morado de las flores
y la p minúscula al alelo recesivo del color blanco. Una planta ho-
mocigota de flores moradas tiene dos alelos para el color morado
(PP), mientras que una planta homocigota de flores blancas tiene
dos alelos para el color blanco (pp). Todos los espermatozoides y
óvulos producidos por una planta PP portan el alelo P, mientras
que todos los espermatozoides y óvulos de una planta pp llevan
el alelo p (
FIGURA 10-7a).
La primera generación filial F
1
fue producida cuando es-
permatozoides P fecundaron óvulos p o cuando espermatozoides
p fecundaron óvulos P. En ambos casos, la generación F
1
era Pp.
Como P domina sobre p, todos los descendientes dieron flores
moradas (
FIGURA 10-7b).
Para la generación F
2
, Mendel permitió que plantas hete-
rocigotas F
1
se autopolinizaran. Cada gameto producido por una
planta heterocigota Pp tenía las mismas probabilidades de recibir
el alelo P o el alelo p. Es decir, una planta heterocigota produce el
mismo número de espermatozoides P y p, y el mismo número
de óvulos P y p. Cuando una planta Pp se autopoliniza, cada tipo de
espermatozoide tiene las mismas probabilidades de fecundar a
cada tipo de óvulo (
FIGURA 10-7c). Por tanto, la generación F
2
comprende tres clases de descendientes: PP, Pp y pp. Los tres tipos
se presentan aproximadamente en una cuarta parte de PP (ho-
7YPTLYHNLULYHJP}U
MPSPHSKL
KLZJLUKPLU[LZ-
PRUDGDV
EODQFDV
:LN\UKHNLULYHJP}U
MPSPHSKL
KLZJLUKPLU[LZ-
VHDXWRSROLQL]D
FIGURA 10-5 Autopolinización de plantas de chícharo F
1
con flores moradas Tres cuartas partes de la descendencia dan
flores moradas y una cuarta parte, flores blancas.
SURJHQLWRUKRPRFLJRWR
H .HTL[VZWYVK\JPKVZWVY\UWYVNLUP[VYOVTVJPNV[V
I .HTL[VZWYVK\JPKVZWVY\UWYVNLUP[VYOL[LYVJPNV[V
((
((
JDPHWRV
SURJHQLWRUKHWHURFLJRWR
( H
H(
JDPHWRV
FIGURA 10-6 Distribución de los alelos en los
gametos (a) Todos los gametos producidos por los organismos
homocigotos contienen el mismo alelo. (b) La mitad de los gametos
producidos por organismos heterocigotos contienen un alelo y la
otra mitad, el otro alelo.
178 UNIDAD 2 Herencia
H .HTL[VZWYVK\JPKVZWVYWHKYLZOVTVJPNV[VZ
I 3HM\ZP}UKLSVZNHTL[VZWYVK\JLSHKLZJLUKLUJPH-
77
7
7
SURJHQLWRUPRUDGR
WRGRVORV
HVSHUPDWR]RLGHV
\ʼnYXORV7
WW
W
SURJHQLWRUEODQFR
WRGRVORV
HVSHUPDWR]RLGHV
\ʼnYXORVW
W
W
W
7W
7W
HVSHUPDWR]RLGHV ʼnYXORV
7
7
RELHQ
WW
W
7W
W
W
JDPHWRVGHSODQWDV7WGH)
ʼnYXORV
GHVFHQGHQFLD)
W
7
7
7
7
77
HVSHUPDWR]RLGHV
7W
J3HM\ZP}UKLNHTL[VZKLSHNLULYHJP}U-
WYVK\JLSH
KLZJLUKLUJPH-
GHVFHQGHQFLD)
FIGURA 10-7 La segregación de los alelos y la fusión de
los gametos pronosticó la distribución de los alelos y los
rasgos en el experimento de Mendel con el color de las
flores del chícharo (a) Generación de los progenitores. Todos los
gametos de los progenitores homocigotos contienen el mismo alelo:
sólo alelos P en los gametos de progenitores PP y sólo alelos pp en
los gametos de los progenitores pp. (b) Generación F
1
: la fusión de
gametos que contienen el alelo P con gametos que contienen el alelo
p produce descendencia exclusivamente Pp. (c) Generación F
2
: la
mitad de los gametos de los progenitores heterocigotos Pp contiene
el alelo P y la otra mitad contiene el alelo p. La fusión de estos
gametos produce descendientes PP, Pp y pp.
mocigoto morado), una mitad Pp (heterocigoto morado) y una
cuarta parte pp (homocigoto blanco).
Dos organismos que se ven idénticos pueden contener
diferentes combinaciones de alelos. La combinación particu-
lar de alelos que lleva un organismo (por ejemplo, PP o Pp)
es su genotipo. Los rasgos de un organismo, como su aspecto,
conducta, enzimas digestivas, tipo de sangre y otros elementos
observables o mensurables, constituyen su fenotipo. Como vi-
mos, las plantas con genotipo PP o Pp tienen el fenotipo de las
flores moradas. Por consiguiente, la generación F
2
de los chícha-
ros de Mendel estaba compuesta por tres genotipos (una cuarta
parte PP, un medio Pp y otra cuarta parte pp), pero sólo dos fe-
notipos (tres cuartas partes de flores moradas y una cuarta parte
de flores blancas).
Un “registro genético” simple puede predecir
genotipos y fenotipos de las células hijas
El método de los cuadrados de Punnett, así nombrado por el
famoso genetista de principios del siglo XX, R.C. Punnett, es una
manera cómoda de pronosticar los genotipos y fenotipos de la des-
cendencia. En la
FIGURA 10-8 se muestra cómo aplicar el cuadra-
do de Punnett para determinar la proporción de células hijas que
nacen de la autopolinización de una planta que es heterocigota
para el color (o las proporciones de descendencia que resulta de
dos organismos que son heterocigotos para un rasgo). Esta cifra
también da las fracciones que permiten calcular los resultados por
medio de las probabilidades de que cada tipo de espermatozoide
fecunde cada tipo de óvulo.
¿Te has preguntado…
por qué los perros tienen tamaños tan variados?
Todos los perros evolucionaron de los lobos. Ahora bien, todos
los lobos son más o menos del mismo tamaño, pero los perros
varían más que cualquier otro mamífero, del gran danés y el
sabueso irlandés al minúsculo chihuahua y poodle miniatura.
En 2007, investigadores descubrieron que las razas de perros
miniatura son homocigotas para el alelo “pequeño” que codifica
el factor de crecimiento parecido a la insulina (FCI), una proteína
que ayuda a regular la talla de muchos mamíferos. Las razas
grandes llevan por lo menos un alelo “grande”. Es probable que
los lobos no tengan el alelo “pequeño”.
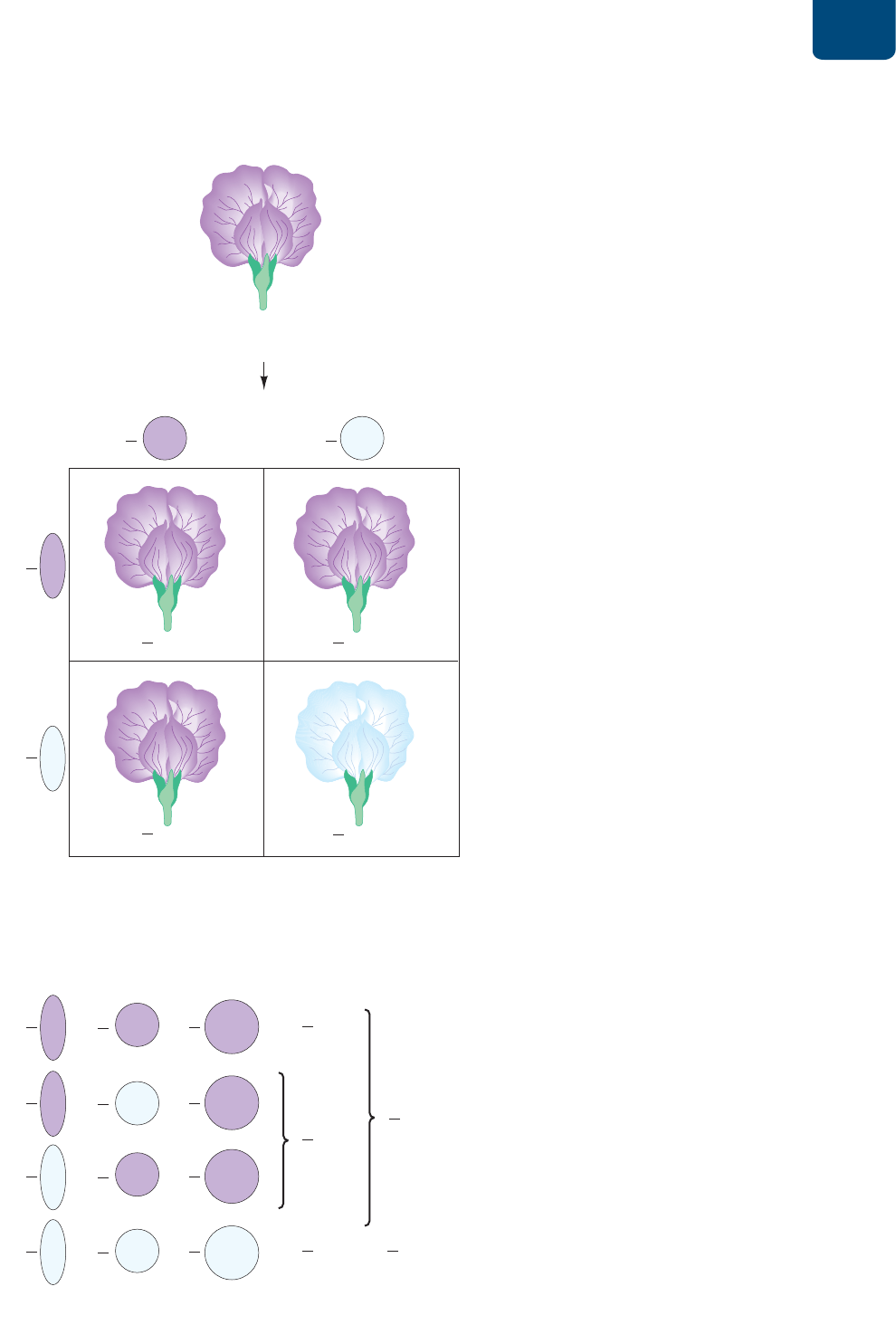
Patrones de la herencia Capítulo 10 179
7
W
ʼnYXORV
VHDXWRSROLQL]D
W7
HVSHUPDWR]RLGHV
7
W
7
7
77
ⴛ
ⴝ
77
7
W
7W
ⴛ
ⴝ
PRUDGDV
W
7
W7
ⴛ
ⴝ
7W
W
W
WW
ⴛ
ⴝ
WW
EODQFDV
HVSHUPDWR]RLGHV
ʼnYXORV
JHQRWLSRVGHOD
GHVFHQGHQFLD
SURSRUFLʼnQ
JHQRWķSLFD
SURSRUFLʼnQ
IHQRWķSLFD
77
WW
7W
W7
H *\HKYHKVKL7\UUL[[KLSJY\JLKL\UYHZNVUPJV
I *mSJ\SVKLWYVIHIPSPKHKLZWHYHKL[LYTPUHYSHKLZJLUKLUJPH
KLSHJY\aHKL\UYHZNVUPJV
FIGURA 10-8 Determinación del resultado de la cruza de
un rasgo único (a) El cuadrado de Punnett permite anticipar los
genotipos y fenotipos de cruzas específicas; aquí, lo usamos para una
cruza entre plantas de chícharo que son heterocigotas para un rasgo
único: el color de las flores.
(1) Se asignan letras a los diferentes alelos: mayúsculas para los
alelos dominantes y minúsculas para los recesivos.
(2) Se determinan todos los tipos de gametos genéticamente
diferentes que pueden producir los progenitores masculino y
femenino.
(3) Se traza el cuadrado de Punnett, con las columnas marcadas con
los posibles genotipos de los óvulos y las hileras con los posibles
genotipos de los espermatozoides (incluimos las fracciones de
esos genotipos en cada designación).
(4)
Se anota el genotipo de la descendencia en cada columna
combinando el genotipo del espermatozoide de su hilera con el
genotipo del óvulo en su columna (se multiplican las fracciones
de los espermatozoides de cada tipo que aparecen en los
encabezados de las columnas por la fracción de los óvulos de
cada tipo en los encabezados de las columnas).
(5) Se cuenta el número de descendientes con cada genotipo.
Observa que Pp es lo mismo que pP.
(6) Se convierte el número de descendientes de cada genotipo a una
fracción del total de descendientes. En este ejemplo, de cuatro
fecundaciones, se prevé que sólo una produzca el genotipo pp,
así que se pronostica que una cuarta parte del número total
de la descendencia producida por la cruza será blanca. Para
determinar las fracciones fenotípicas, se suman las fracciones de
los genotipos que producirían un fenotipo dado. Por ejemplo, las
flores moradas son resultado de
1
–
4
PP +
1
–
4
Pp +
1
–
1
–
4
pP, lo que da un
total de tres cuartas partes de la descendencia.
(b) También pueden calcularse probabilidades para pronosticar el
resultado de la cruza de un solo rasgo. Se determinan las fracciones
de óvulos y espermatozoides de cada genotipo y se multiplican estas
fracciones para calcular la fracción de los descendientes de cada
genotipo. Cuando dos genotipos producen el mismo fenotipo (por
ejemplo, Pp y pP), se suman las fracciones de cada genotipo para
determinar la fracción genotípica.
PREGUNTASi se cruza una planta heterocigota Pp con una planta
homocigota recesiva pp, ¿cuál sería la proporción esperada de la
descendencia? ¿En qué difiere de la descendencia de una cruza
PP pp? Trata de resolver el problema antes de avanzar con tu
lectura del texto.
Mientras aplicas estas técnicas de “registro genético”, re-
cuerda que en un experimento real la descendencia se presentará
aproximadamente en las proporciones pronosticadas, porque los
espermatozoides y los óvulos con alelos diferentes se encuentran
al azar. Veamos un ejemplo, cada vez que se concibe un bebé,
tiene una probabilidad 50:50 de ser niño o niña. Sin embargo,
muchas familias con dos hijos no tienen niña y niño. La propor-
ción 50:50 de niñas y niños aparece únicamente si promediamos
el género de los hijos de muchas familias.
La hipótesis de Mendel puede usarse
para predecir el resultado de nuevos
tipos de cruzas de rasgos únicos
Es probable que te hayas dado cuenta de que Mendel aplicó el mé-
todo científico: hizo una observación y la tomó para formular una
hipótesis. Pero, ¿es atinada la hipótesis de Mendel en predecir los
resultados de otros experimentos? A partir de la hipótesis de que
las plantas heterocigotas F
1
tienen un alelo para las flores moradas
y uno para las blancas (es decir, que tienen el fenotipo Pp), Mendel
predijo el resultado de la fecundación cruzada de plantas Pp con
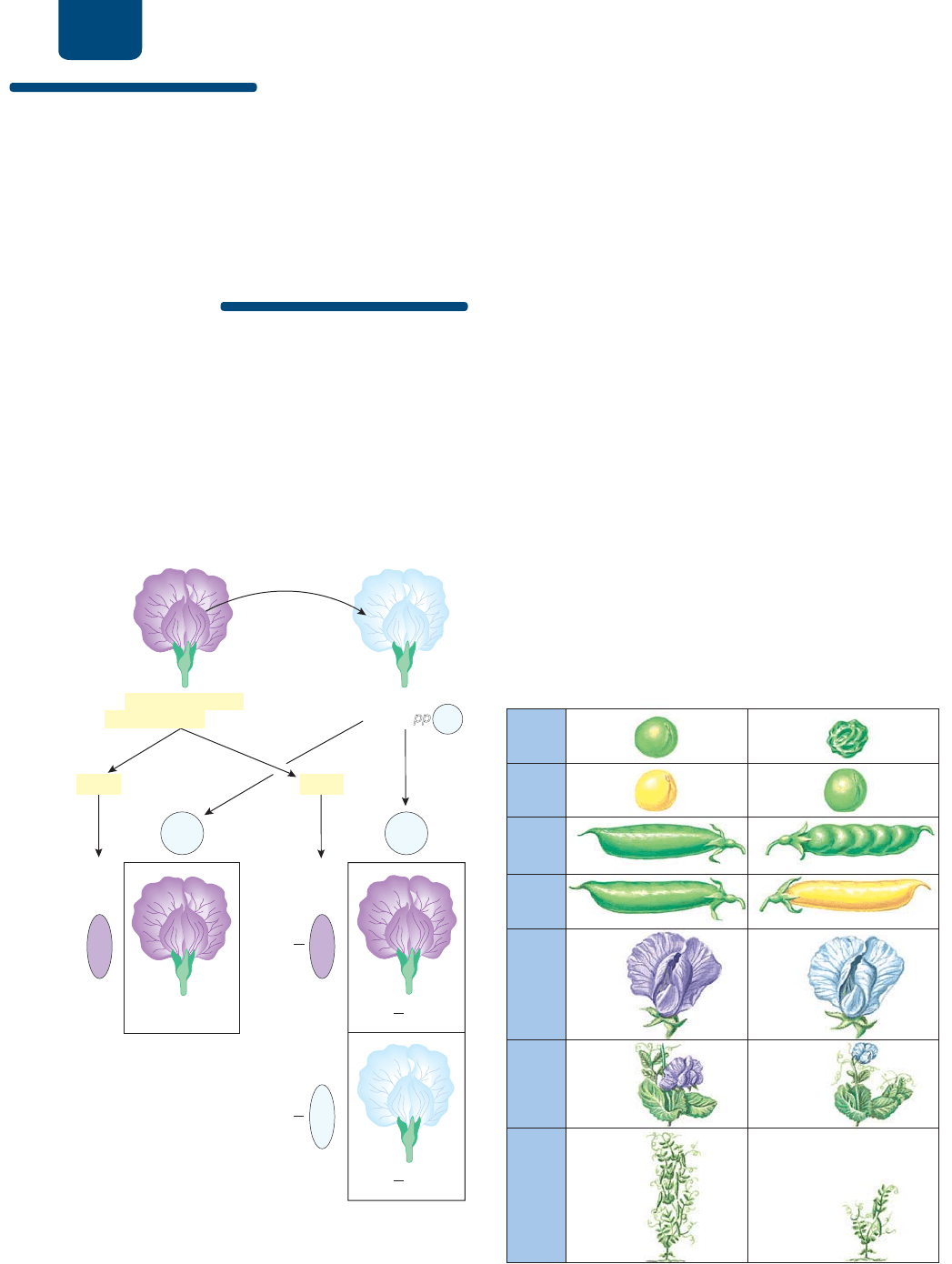
180 UNIDAD 2 Herencia
Cuando se cruza un dominante homocigoto (PP) con un recesivo
homocigoto (pp), produce toda la descendencia fenotípicamente
dominante, mientras que un dominante heterocigoto (Pp) da cé-
lulas hijas con fenotipos dominantes y recesivos en proporción
de 1:1 (uno a uno).
10.4
¿CÓMO SE HEREDAN
LOS RASGOS MÚLTIPLES?
Después de haber determinado las modalidades de la herencia de
rasgos únicos, Mendel pasó a la más compleja cuestión de la he-
rencia de los rasgos múltiples en las plantas de chícharos (
FIGURA
10-10
). Para empezar, cruzó plantas que variaban en dos rasgos;
por ejemplo, el color de la semilla (amarillo o verde) y la forma
(lisa o rugosa). De otras cruzas de plantas con estos rasgos, Mendel
ya sabía que el alelo liso del gen de la forma de la semilla (S) es
dominante sobre el alelo rugoso (s) y que el alelo amarillo del gen
del color de la semilla (Y) es dominante sobre el alelo verde (y).
Cruzó una planta de raza pura con semillas lisas amarillas (SSYY)
con una planta de raza pura de semillas rugosas verdes (ssyy). La
planta SSYY produjo únicamente gametos SY y la planta ssyy pro-
dujo únicamente gametos sy. Por tanto, todos los descendientes F
1
fueron heterocigotos: genotípicamente SsYy, con el fenotipo de las
semillas lisas amarillas.
Al hacer que estas plantas heterocigotas F
1
se autofecunda-
ran, Mendel vio que la generación F
2
constó de 315 plantas con
SROHQ
WRGRV
ORV
HVSHUPDWR]RLGHV
W
W
7
WW
7W
WRGRVORV
ʼnYXORVWW
HVSHUPDWR]RLGHV
GHVFRQRFLGRV77R7W
VL77 VL7W
ʼnYXORV ʼnYXORV
W
HVSHUPDWR]RLGHV
7
W
WRGRV7W
FIGURA 10-9 Cuadrado de Punnett de una cruza de
prueba Un organismo con un fenotipo dominante puede ser
homocigoto o heterocigoto. La cruza de tal organismo con un
organismo recesivo homocigoto puede determinar si el organismo
dominante era homocigoto (izquierda) o heterocigoto (derecha).
Estudio de caso continuación
Muerte súbita en la cancha
Cuando una persona con síndrome de Marfan tiene hijos con
una persona sin el síndrome, los hijos tienen una probabilidad
de 50% de heredar la condición. ¿Crees que el síndrome de
Marfan se hereda como alelo dominante o recesivo? ¿Por qué?
Comprueba tu razonamiento en el apartado “Estudio de caso
otro vistazo”, al final del capítulo.
)RUPD
GHOD
VHPLOOD
&RORU
GHOD
VHPLOOD
&RORU
GHOD
YDLQD
)RUPD
GHOD
YDLQD
&RORU
GHOD
IORU
8ELFDFLʼnQ
GHOD
IORU
HQODV
MXQWXUDV
GHODVKRMDV
HQOD
SXQWDGH
ODVUDPDV
DOWDXQRV
FP
DSUR[
FRUWDGH
DFP
DSUR[
7DPDŅR
GHOD
SODQWD
OLVD
-VYTHKVTPUHU[L9HZNV -VYTHYLJLZP]H
UXJRVD
DPDULOOD
DPDULOOD
LQIODGD
YHUGH
YHUGH
EODQFD
FRQVWUHŅLGD
PRUDGD
FIGURA 10-10 Rasgos de las plantas de chícharo
estudiadas por Gregorio Mendel
plantas homocigotas recesivas blancas (pp): debe haber cifras igua-
les de descendientes Pp (flores moradas) y pp (flores blancas). Eso
fue exactamente lo que encontró.
Este experimento tiene también usos prácticos. La fecunda-
ción cruzada de un organismo con un fenotipo dominante (en
este caso, la flor morada) pero un genotipo desconocido con un
organismo recesivo homocigoto (una flor blanca) prueba si el or-
ganismo con el fenotipo dominante es homocigoto o heterocigo-
to; como es lógico, esto se llama cruza de prueba (
FIGURA 10-9).
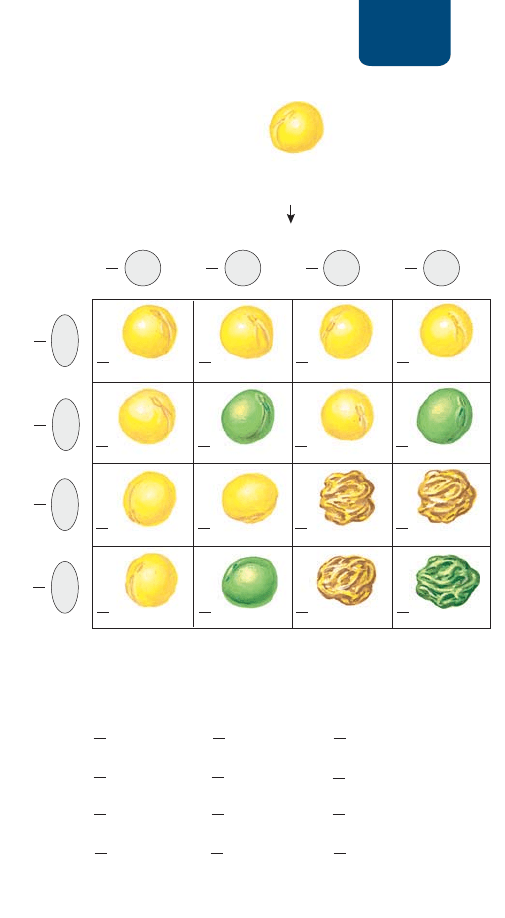
Patrones de la herencia Capítulo 10 181
semillas lisas amarillas, 101 con semillas rugosas amarillas, 108
con semillas lisas verdes y 32 con semillas rugosas verdes: una
proporción de aproximadamente 9:3:3:1. La descendencia pro-
ducida por otras cruzas de plantas heterocigotas para dos rasgos
también daba proporciones fenotípicas de alrededor de 9:3:3:1.
Mendel postuló que los rasgos se heredan
de forma independiente
Mendel se dio cuenta de que estos resultados se explicarían si los
genes del color y de la forma de las semillas se heredaban de forma
independiente y si no se influían unos a otros durante la formación
de los gametos. De ser así, y para cada rasgo, tres cuartas partes de
la descendencia mostrarían el fenotipo dominante y un cuarto
mostraría el fenotipo recesivo. Este resultado fue lo que observó
Mendel. Obtuvo 423 plantas con semillas lisas (de cualquier co-
lor) y 133 con semillas rugosas (una proporción de 3:1); en este
mismo grupo de plantas, 416 produjeron semillas amarillas (de
cualquier forma) y 140 produjeron semillas verdes (también alre-
dedor de 3:1). En la
FIGURA 10-11 se explica cómo trazar un cua-
dro de Punnett o cómo calcular probabilidades para determinar
el resultado de una cruza entre organismos que son heterocigotos
para dos rasgos.
La herencia independiente de dos o más rasgos se llama ley
de la distribución independiente. Los rasgos múltiples se here-
dan de forma independiente si los alelos de un gen están distri-
buidos en los gametos separados de los alelos de otros genes. Se
produce una distribución independiente cuando los rasgos que
se estudian están controlados por genes de diferentes pares de
cromosomas homólogos. ¿Por qué? Recuerda que en el capítulo 9
vimos el movimiento de los cromosomas en la meiosis. Cuando
cromosomas homólogos apareados se alinean en la metafase I,
qué homólogo se dirija a cuál polo de la célula es cuestión del
azar y la orientación de un par de homólogos no influye en los
otros pares. Por tanto, cuando los homólogos se separan en la
anafase I, el alelo de un gen del par homólogo 1 se mueve al “nor-
te” no afectando si también el alelo de un gen del par homólogo
2 se mueve al “norte”; es decir, los alelos de los genes de cromoso-
mas diferentes se distribuyen, o reparten, de forma independiente
(
FIGURA 10-12).
En un mundo que no está preparado para ellos,
los genios pueden pasar inadvertidos
En 1865, Gregorio Mendel presentó los resultados de sus experi-
mentos sobre la herencia a la Sociedad Brünn para el Estudio de las
Ciencias Naturales y fueron publicados al año siguiente. Su trabajo
no marcó el comienzo de la genética, ni hizo mella en el estudio de
la biología durante la vida de Mendel. Sus experimentos, que des-
pués darían lugar a una de las teorías científicas más importantes
de la biología, parecieron desvanecerse. Pocos biólogos leyeron su
artículo y ninguno se dio cuenta de su relevancia.
Fue en 1900 cuando tres biólogos: Carl Correns, Hugo
de Vries y Erich Tschermak, trabajando de forma paralela, inde-
pendientemente unos de otros y sin conocer la obra de Mendel,
redescubrieron el principio de la herencia. Quizá se sintieron
muy decepcionados cuando al buscar en la bibliografía cientí-
fica antes de hacer públicos sus resultados, se dieron cuenta de
que Mendel se les había adelantado más de 30 años. Para méri-
HVSHUPDWR]RLGHV
:@
:@
::@@ :Z@@
ZZ@@
ZZ`@
:Z`@
::@` :Z@`
:Z@`
ZZ@`
ZZ``
:Z``::``
Z:`@ Z:``
Z:@@ Z:@`
::`@
Z@
Z@
Z`
Z`
:`
:`
ʼnYXORV
VHDXWRSROLQL]D
MVYTHKL
SHZLTPSSH
JVSVYKL
SHZLTPSSH
WYVWVYJP}U
MLUV[xWPJH
OLVD
DPDULOOD
OLVDDPDULOOD
OLVDYHUGH
UXJRVDDPDULOOD
UXJRVDYHUGH
DPDULOOD
YHUGH
YHUGH
OLVD
UXJRVD
UXJRVD
ⴛ
ⴛ
ⴛ
ⴛ
ⴝ
ⴝ
ⴝ
ⴝ
H *\HKYHKVKL7\UUL[[KL\UHJY\aHKLKVZYHZNVZ
I *mSJ\SVKLWYVIHIPSPKHKLZWHYHKL[LYTPUHYSHKLZJLUKLUJPH
KL\UHJY\aHKLKVZYHZNVZ
FIGURA 10-11 Predicción de los genotipos y fenotipos
de una cruza entre progenitores heterocigotos para dos
rasgos En las semillas de chícharo, el color amarillo (Y) es dominante
sobre el verde (y) y la forma lisa (S) es dominante sobre la rugosa
(s). (a) Análisis en el cuadrado de Punnett. En esta cruza, un
individuo heterocigoto para ambos rasgos se autopoliniza. En una
cruza de dos genes independientes, los tipos de gametos constan
de todas las posibles combinaciones de alelos de los dos genes: S
con Y, S con y, s con Y y s con y. Se colocan estas combinaciones
de gametos como designaciones de las hileras y las columnas del
cuadrado de Punnett y se calcula la descendencia como se explicó
en la figura 10-8. Observa que el cuadrado de Punnett predice tanto
la frecuencia de las combinaciones de rasgos (
–
9
16
lisa amarilla,
–
3
16
lisa
verde,
–
3
16
rugosa amarilla y
–
1
16
rugosa verde) como la frecuencia de
los rasgos por separado (
3
–
4
amarilla,
1
–
4
verde,
3
–
4
lisa y
1
–
4
rugosa).
(b) La teoría de las probabilidades establece que la probabilidad de
dos eventos independientes es el producto (la multiplicación) de sus
probabilidades por separado. La forma de la semilla es independiente
de su color. Por tanto, multiplicar las probabilidades independientes de
los genotipos o fenotipos de cada rasgo produce las frecuencias
pronosticadas para los genotipos o fenotipos combinados de la
descendencia. Estas proporciones son idénticas a las que se obtienen
con el cuadrado de Punnett.
PREGUNTA ¿El genotipo de una planta con semillas lisas amarillas
puede revelarse mediante una cruza de prueba con una planta que
dé semillas verdes rugosas?
182 UNIDAD 2 Herencia
to suyo, aceptaron donosamente la importancia del trabajo del
monje austriaco, quien murió en 1884.
10.5
¿CÓMO SE HEREDAN LOS GENES
SITUADOS EN EL MISMO CROMOSOMA?
Gregorio Mendel no sabía nada sobre la naturaleza física de los
genes ni de los cromosomas. Mucho después, cuando los cien-
tíficos descubrieron que los cromosomas son los vehículos de la
herencia, se hizo obvio que había muchos más rasgos (y, por tan-
to, muchos más genes) que cromosomas. Ahora sabemos que los
genes son parte de los cromosomas y que cada cromosoma con-
tiene muchos genes (hasta varios miles en los cromosomas muy
grandes). Estos hechos tienen importantes implicaciones para la
herencia.
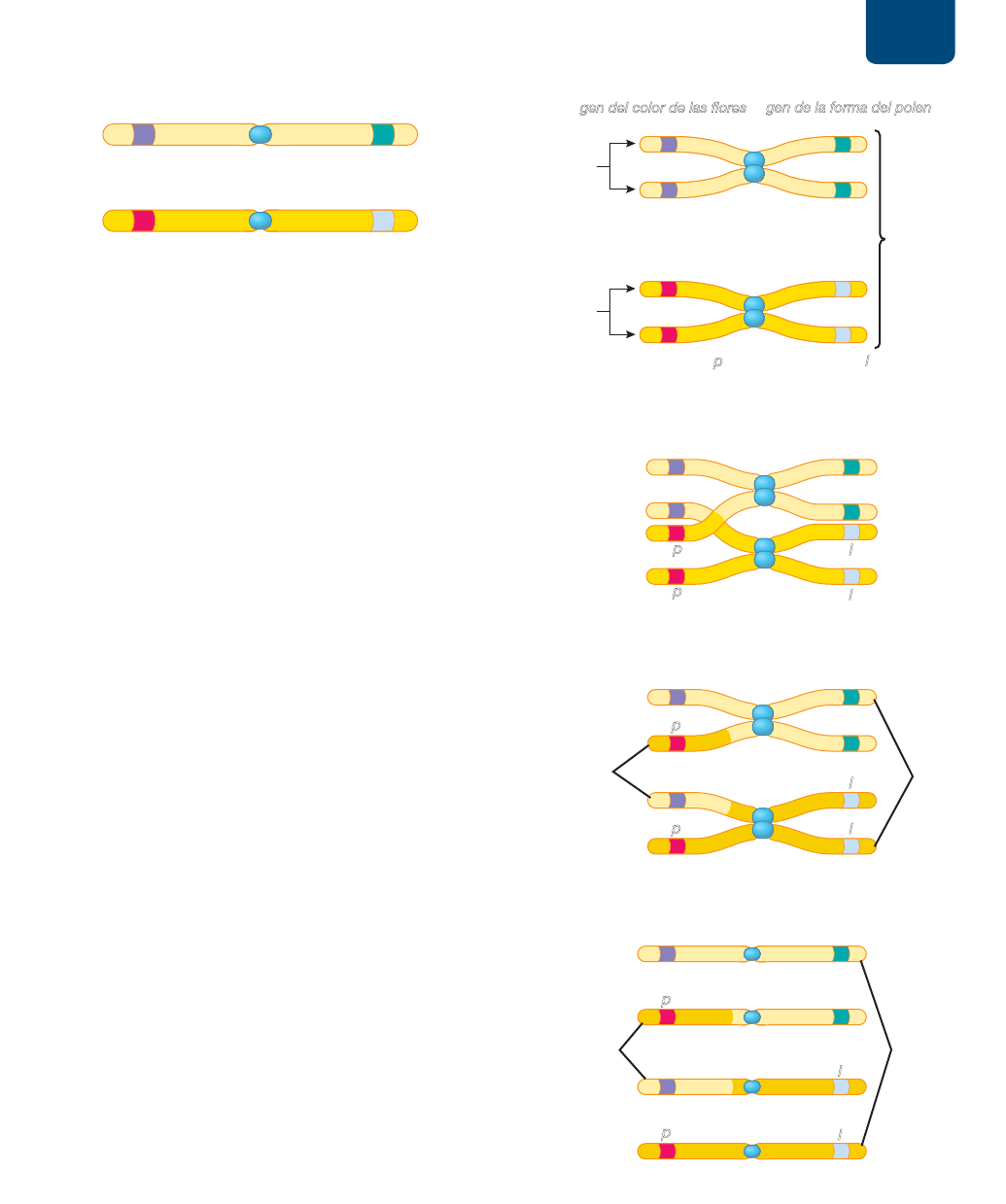
Los genes del mismo cromosoma
tienden a heredarse juntos
Ahora sabemos que los cromosomas, no los genes, se distribuyen
por separado durante la meiosis I. Por tanto, los genes situados en
diferentes cromosomas se distribuyen de forma independiente en ga-
metos. Por el contrario, los genes del mismo cromosoma se heredan
juntos, un fenómeno llamado ligamiento. Uno de los primeros
pares de genes ligados se descubrió en la planta de chícharo de
olor, una especie diferente de la planta de chícharo comestible
de Mendel. En las plantas de chícharos de olor, el gen del color de
la flor (morada o roja) y el gen de la forma del grano de polen
(redondo o alargado) se transmiten en el mismo cromosoma. Así,
los alelos de estos genes se distribuyen juntos en la meiosis y se
heredan juntos.
Pensemos en una planta heterocigota de chícharo de olor
con flores moradas y polen alargado (
FIGURA 10-13). Observa
que el alelo dominante morado del gen del color de la flor y el
alelo dominante alargado del gen de la forma del polen se en-
cuentran en un cromosoma homólogo (véase la figura 10-13,
arriba). El alelo recesivo rojo del gen del color de la flor y el alelo
recesivo redondo del gen de la forma del polen se encuentran
en el otro homólogo (véase la figura 10-13, abajo). Por tanto, es
probable que los gametos producidos por esta planta tengan ale-
los morado y largo o bien alelos rojo y redondo. Este esquema
de herencia no cumple la ley de la distribución independiente
porque los alelos del color de la flor y de la forma del polen no
se distribuyen de forma independiente uno del otro, sino que se
quedan juntos en la meiosis.
ODGLVWULEXFLʼnQLQGHSHQGLHQWHSURGXFHFXDWURFRPELQDFLRQHV
GHDOHORVLJXDOPHQWHSUREDEOHVGXUDQWHODPHLRVLV
SDUHVGHDOHORVHQORVFURPRVRPDV
KRPʼnORJRVGHFÒOXODVGLSORLGHV
SVZJYVTVZVTHZ
ZLK\WSPJHU
ORVKRPʼnORJRVGXSOLFDGRV
VHHPSDUHMDQHQODPHWDIDVH
GHODPHLRVLV,\VH
TLPVZPZ00
TLPVZPZ0
:
:
:
:
:
:
:
:
:
:@ Z@
Z
Z
Z
Z
Z
Z
Z
Z
Z` :`
Z
Z
Z
@
@
@
@
@
:
@
@
@
@
@
@
`
`
`
`
`
`
`
`
`
`
:
`
RULHQWDQDVķ
RDVķ
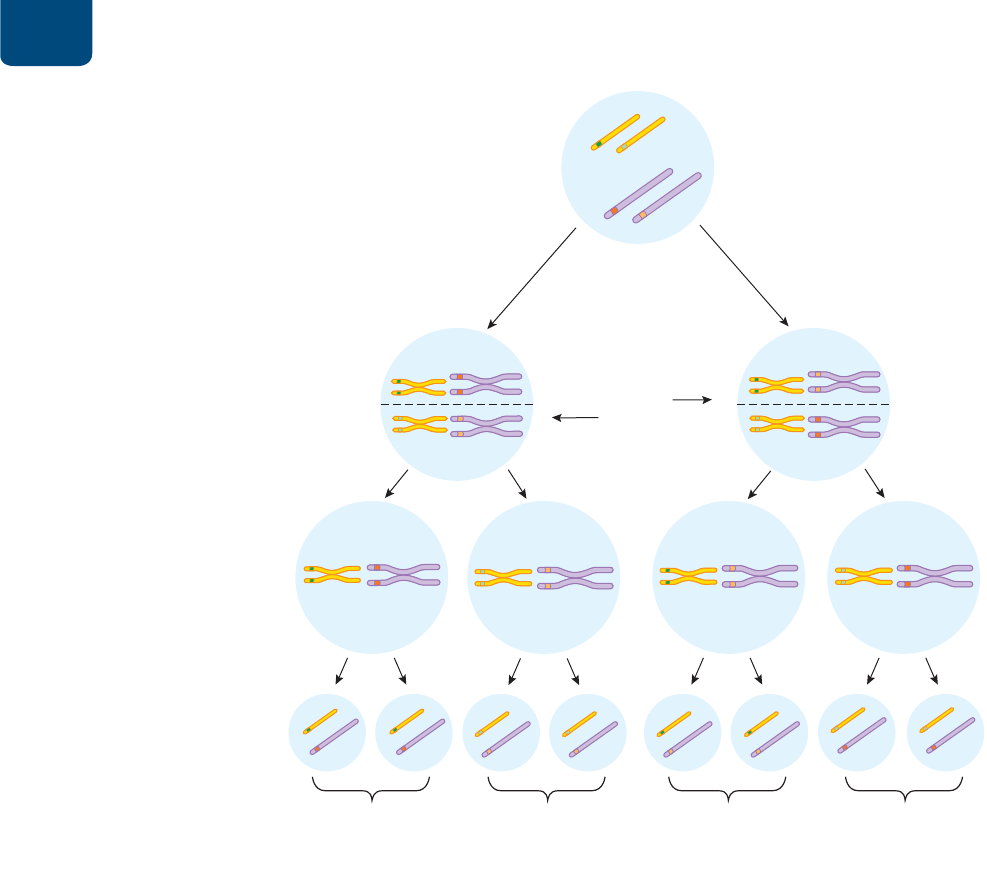
FIGURA 10-12 Distribución independiente de los alelos Los
movimientos de los cromosomas en la meiosis producen distribuciones
independientes de alelos, que aquí se muestran para dos genes. Cada
combinación de alelos tiene la misma probabilidad de ocurrir y producir
gametos en las proporciones predichas de una cuarta parte SY, una cuarta
parte sy, una cuarta parte Sy y una cuarta parte sY.
PREGUNTA Si los genes del color de las semillas y de la forma de éstas
se encontraran en el mismo cromosoma en lugar de en cromosomas
diferentes, ¿sus alelos se distribuirían de forma independiente? ¿Por qué?
Patrones de la herencia Capítulo 10 183
Los cruzamientos producen nuevas
combinaciones de alelos ligados
Aunque tienden a heredarse juntos, los genes del mismo cromoso-
ma no siempre se quedan juntos. Si se hace una fecundación cruza-
da de dos plantas de chícharos de olor con los cromosomas que
se indican en la figura 10-13, sería de esperarse que toda la des-
cendencia tuviera flores moradas con granos de polen alargados,
o bien, flores rojas con granos de polen redondos (trata de resol-
verlo con un cuadrado de Punnett). Sin embargo, en la realidad se
suelen encontrar algunas plantas en las que los genes del color y
la forma del polen se heredaron como si no estuvieran ligados; es
decir, parte de la descendencia tiene flores moradas y polen redon-
do y parte tiene flores rojas y polen alargado. ¿A qué se debe esto?
Como vimos en el capítulo 9, durante la profase I de la
meiosis, los cromosomas homólogos a veces intercambian par-
tes por entrecruzamiento (véase la figura 9-16). En casi todos los
cromosomas ocurre por lo menos un intercambio entre cada par
homólogo durante la división meiótica. El intercambio de seg-
mentos correspondientes de ADN durante el entrecruzamiento
produce nuevas combinaciones de alelos en los dos cromosomas
homólogos. Luego, cuando los homólogos se separan en la ana-
fase I, los cromosomas que reciben las hijas haploides tienen dife-
rentes juegos de alelos que los cromosomas de la célula original.
El entrecruzamiento durante la meiosis explica la recombi-
nación genética, la aparición de nuevas combinaciones de alelos
que estaban previamente enlazados. En la profase I, los cromoso-
mas homólogos duplicados se emparejan (
FIGURA 10-14a). Cada
homólogo tendrá una o más regiones en que haya un entrecruza-
miento. Imagina que en el entrecruzamiento se intercambian los
alelos del color de las flores entre cromátidas no hermanas de
los dos homólogos (
FIGURA 10-14b). En la anafase I, los homólo-
gos separados tendrán una cromátida que lleve parte del ADN de
una cromátida del otro homólogo (
FIGURA 10-14c). En la meio-
sis II se distribuyen cuatro tipos de cromosomas a las cuatro célu-
las hijas haploides: dos cromosomas intactos y dos cromosomas
recombinados (
FIGURA 10-14d).
Por tanto, se producirán algunos gametos con las siguientes
cuatro configuraciones: PL y pl (los tipos de los progenitores) y
Pl y pL (los cromosomas recombinados). Si un espermatozoide
con un cromosoma Pl fecunda un óvulo con un cromosoma pl,
la planta resultante tendrá flores púrpuras (Pp) y polen redondo
(ll). Si un espermatozoide con cromosoma pL fecunda un óvulo
con un cromosoma pl, la descendencia tendrá flores rojas (pp) y
polen alargado (Ll).
NLUKLSJVSVYKLSHZMSVYLZ NLUKLSHMVYTHKLSWVSLU
DOHOR
PRUDGR7
DOHOR
DODUJDGR3
DOHOR
URMR
W
DOHOR
UHGRQGRS
FIGURA 10-13 Genes ligados en cromosomas homólogos
en las plantas del chícharo de olor Los genes del color de las
flores y la forma del polen se encuentran en el mismo cromosoma,
así que tienden a heredarse juntos.
NLUKLSJVSVYKLSHZMSVYLZ
DOHORPRUDGR7 DOHORDODUJDGR3
DOHORURMRW DOHORUHGRQGRS
NLUKLSHMVYTHKLSWVSLU
FURPÀWLGDV
KHUPDQDV
FURPRVRPDV
KRPʼnORJRV
GXSOLFDGRV
HQODPHLRVLV,
FURPÀWLGDV
KHUPDQDV
H*YVTVZVTHZK\WSPJHKVZLUSHWYVMHZLKLSHTLPVZPZ0
I,U[YLJY\aHTPLU[VLUSHWYVMHZL0
7
7
W
W
3
3
S
S
FURPÀWLGDV
UHFRPELQDGRV
73
W
3
7 S
W
S
FURPÀWLGDV
LQWDFWRV
J3VZJYVTVZVTHZOVT}SVNVZZLZLWHYHULUSHHUHMHZL0
73
7
3
W
S
W
S
FURPRVRPDV
UHFRPELQDGRV
K3VZJYVTVZVTHZPU[HJ[VZ`YLJVTIPUHKVZKLZW\tZ
KLSHTLPVZPZ00
FURPRVRPDV
LQWDFWRV
FIGURA 10-14 En el entrecruzamiento se recombinan
los alelos de cromosomas homólogos (a) En la profase de
la meiosis I se emparejan cromosomas homólogos duplicados. (b)
Cromátidas no hermanas de los dos homólogos intercambian partes
por entrecruzamiento. (c) Cuando los cromosomas homólogos se
separan en la anafase de la meiosis I, una cromátida de cada uno de
los homólogos contiene parte del ADN de una cromátida del otro
homólogo. (d) Después de la meiosis II, dos de las células haploides
hijas reciben cromosomas intactos y dos reciben cromosomas
recombinados. Los cromosomas recombinados tienen una disposición
de los alelos que no coincide con los cromosomas originarios.
184 UNIDAD 2 Herencia
mas también se segregan durante la formación de los óvulos,
pero como las hembras tienen dos cromosomas X, cada óvulo
recibe un cromosoma X (junto con un miembro de cada par de
autosomas). Así, se produce un descendiente macho si un óvulo
es fecundado por un espermatozoide que lleve un cromosoma Y
y una hembra si es fecundado por un espermatozoide que lleve
un cromosoma X.
10.7
¿CÓMO SE HEREDAN LOS GENES
LIGADOS A LOS CROMOSOMAS SEXUALES?
Los genes ligados a los cromosomas sexuales
se encuentran sólo en el cromosoma X
o sólo en el cromosoma Y
Se dice que los genes que se encuentran en un cromosoma sexual
y no en el otro están ligados a los cromosomas sexuales. En
muchas especies de animales, el cromosoma Y porta sólo algunos
genes. En los seres humanos, el cromosoma Y tiene algunas doce-
nas de genes (probablemente menos de 80), muchos de los cuales
cumplen una función en la reproducción masculina. El gen ligado
más conocido a un cromosoma Y es SRY, la región que determina
el sexo en el cromosoma Y fue descubierta en 1990. En la vida em-
brionaria, la acción de este gen pone en movimiento toda la ruta
de desarrollo masculina. En condiciones normales, SRY está ligado
100% al sexo masculino en el cromosoma Y.
No es de sorprender que cuanto más lejos estén los genes
en un cromosoma, más probable será que haya un entrecruza-
miento entre ellos. Dos genes muy próximos en un cromosoma
casi nunca se separan por entrecruzamiento, pero si dos genes
están muy apartados, su entrecruzamiento es tan frecuente que
parece distribución independiente, como si estuviesen en cromo-
somas diferentes. Cuando Gregorio Mendel descubrió la distribu-
ción independiente, no sólo fue inteligente y perceptivo, sino que
también tuvo suerte. Los siete rasgos que estudió eran controla-
dos por genes que estaban en cuatro cromosomas diferentes. Ob-
servó la distribución independiente porque los genes que estaban
en el mismo cromosoma se hallaban muy separados.
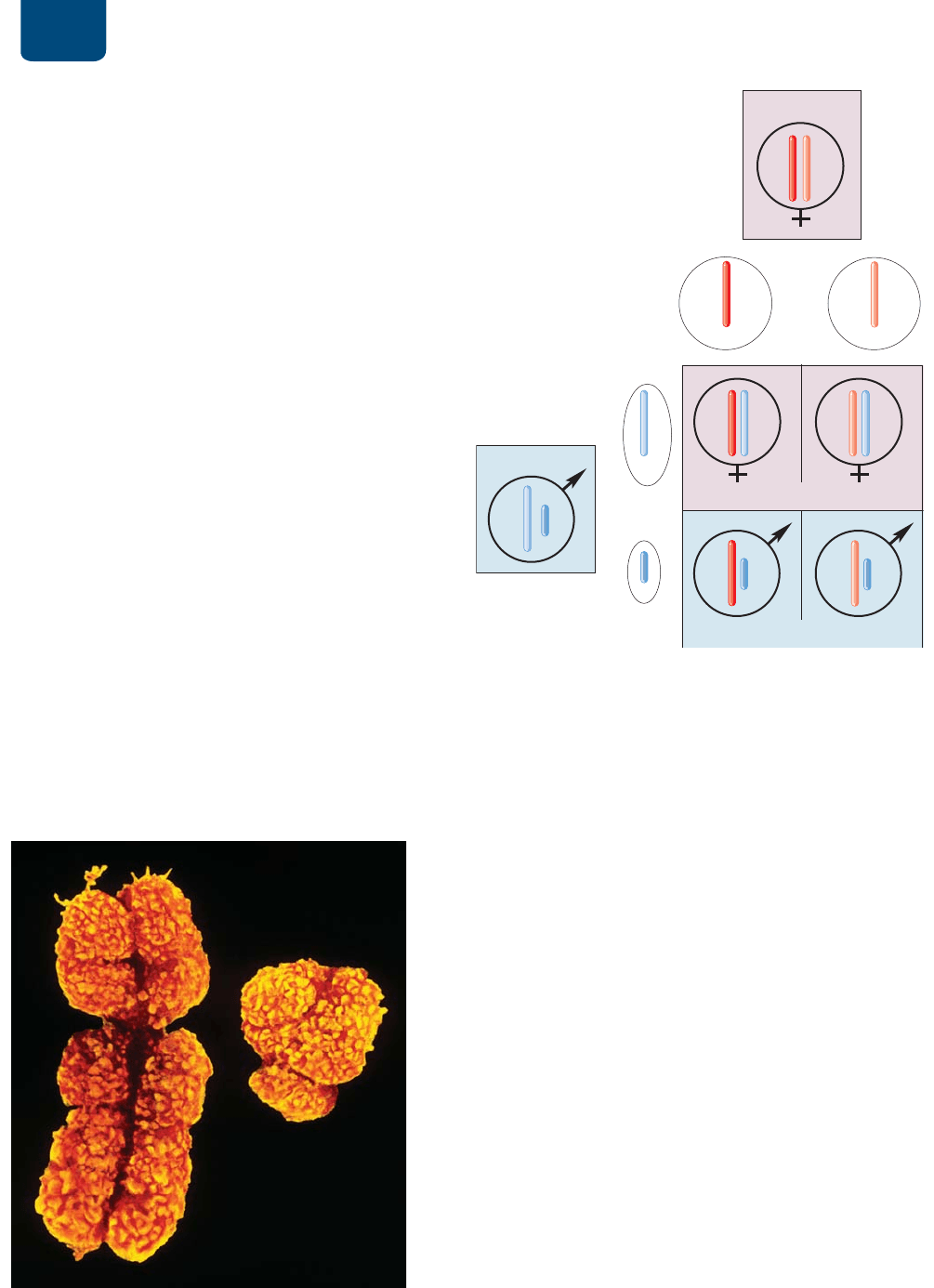
10.6
¿CÓMO SE DETERMINA
GENÉTICAMENTE EL SEXO?
En los mamíferos, el sexo de un individuo está determinado por
un par especial de cromosomas, los cromosomas sexuales. Las
hembras tienen dos cromosomas sexuales idénticos, llamados
cromosomas X, mientras que los machos tienen un cromosoma
X y un cromosoma Y (
FIGURA 10-15). Aunque el cromosoma Y
es mucho más pequeño que el cromosoma X, una pequeña parte
de ambos cromosomas sexuales es homólogo. Como resultado, los
cromosomas X y Y se emparejan durante la profase de la meiosis y
se separan en la anafase I. Los demás cromosomas, que se presen-
tan en pares con la misma apariencia en hembras que en machos,
se llaman autosomas.
En los organismos en que los machos son XY y las hem-
bras XX, el cromosoma sexual que porta el espermatozoide de-
termina el sexo del producto (
FIGURA 10-16). En la formación
de los espermatozoides, los cromosomas sexuales se separan y
cada espermatozoide recibe el cromosoma X o el cromosoma
Y (más un miembro de cada par de autosomas). Los cromoso-
FIGURA 10-15 Micrografía de los cromosomas sexuales
humanos El cromosoma Y (derecha), que tiene pocos genes, es
mucho más pequeño que el cromosoma X (izquierda).
ʼnYXORV
THKYL
KLZJLUKLUJPHMLTLUPUH
KLZJLUKLUJPHTHZJ\SPUH
WHKYL
HVSHUPDWR]RLGHV
;
;
;
;
;
P
;
P
<
<
;
P
;
P
;
;
<
<
;
;
FIGURA 10-16 Determinación del sexo en los mamíferos
La descendencia masculina recibe su cromosoma Y de su padre; la
descendencia femenina recibe el cromosoma X del padre (marcado
X
m
). Tanto los machos como las hembras reciben un cromosoma X
(X
1
o X
2
) de su madre.
FURPRVRPD<
FURPRVRPD;
Patrones de la herencia Capítulo 10 185
En contraste con el pequeño cromosoma Y, el cromosoma
X contiene más de 1,000 genes, pocos de los cuales tienen una
función concreta en la reproducción femenina. Casi ninguno de
los genes del cromosoma X tiene un equivalente en el cromo-
soma Y, incluso en cuanto a rasgos que son tan importantes en
ambos sexos como la visión cromática, coagulación de la sangre
y ciertas proteínas estructurales de los músculos.
¿Qué efecto tiene en la herencia los genes ligados al cromo-
soma X? Como las hembras tienen dos cromosomas X, pueden
ser homocigotas o heterocigotas para los genes del cromosoma
X y se expresarán las relaciones de dominantes y recesivos entre
los alelos. Por el contrario, los machos expresan completamente
todos los alelos que tienen en su único cromosoma X, indepen-
dientemente de que sean dominantes o recesivos.
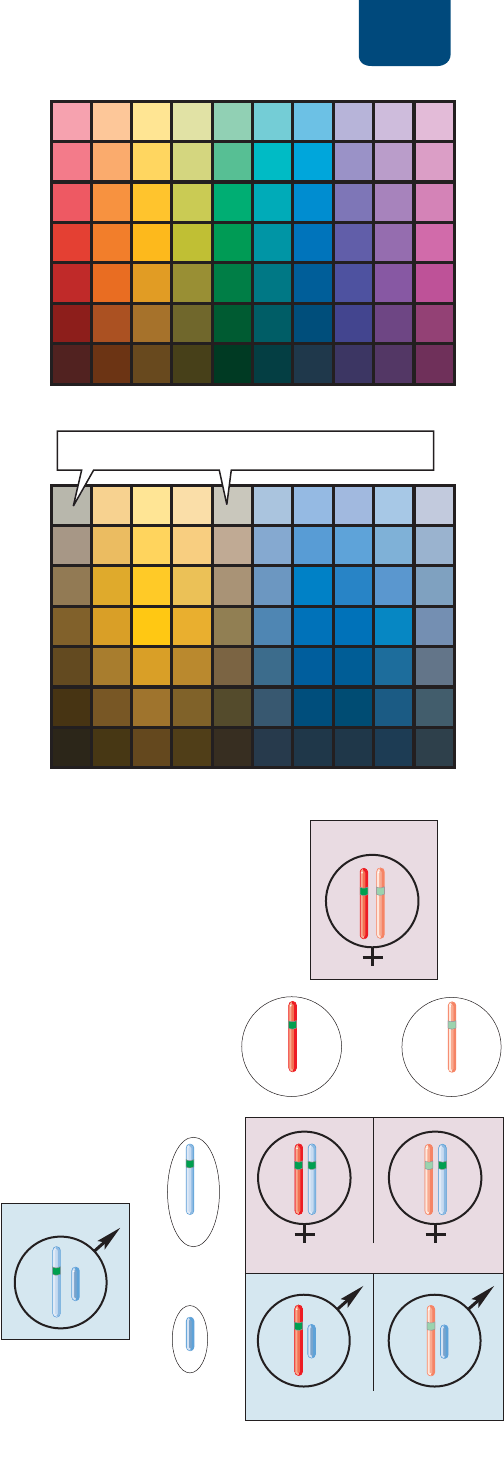
Tomemos un ejemplo conocido: la ceguera a los colores
rojo y verde (
FIGURA 10-17). La ceguera al color está causada por
alelos recesivos en cualquiera de dos genes situados en el cromo-
soma X. Los alelos dominantes normales de estos genes (vamos a
llamarlos C) codifican proteínas, gracias a las cuales un conjunto
de células oculares para la visión cromática, llamadas conos, son
más sensibles a la luz roja y los de otro grupo son más sensibles
a la luz verde (
FIGURA 10-17a). Hay varios alelos recesivos de-
fectuosos de estos genes (vamos a llamarlos c). En los casos más
extremos, uno de los genes falta en un cromosoma X, o bien, ale-
los defectuosos codifican proteínas que hacen que ambos grupos
de conos sean igualmente sensibles a la luz roja y a la verde. Por
consiguiente, la persona afectada no puede distinguir un color
del otro (
FIGURA 10-17b).
¿Cómo se hereda la ceguera al color? Un hombre puede te -
ner un genotipo CY o cY, lo que significa que tiene un alelo de
visión cromática C o c en su cromosoma X y ningún gen corres-
pondiente en su cromosoma Y. Por tanto, tendrá visión cromática
normal si su cromosoma X lleva el alelo C o será ciego al color si
lleva el alelo c. Una mujer puede ser CC, Cc o cc. Las mujeres con
los genotipos CC o Cc tendrán una visión cromática normal; sólo
las mujeres con genotipo cc serán ciegas al color. Aproximada-
mente 7% de los hombres tienen defectos de la visión cromática.
Entre las mujeres, alrededor de 93% son homocigotas normales
CC, 7% son heterocigotas normales Cc y menos de 0.5% son cie-
gas al color cc.
Un hombre ciego al color (cY) transmitirá su alelo defec-
tuoso sólo a sus hijas, porque únicamente ellas heredan su cro-
mosoma X. Ahora bien, las hijas tendrán visión cromática nor-
mal, porque también heredan un alelo C normal de su madre,
que muy probablemente es homocigota normal CC.
Una mujer heterocigota (Cc), aunque tenga visión cromáti-
ca normal, transmitirá su alelo defectuoso a la mitad de sus hijos
(
FIGURA 10-17c). Estos hijos serán ciegos al color (cY). La otra
mitad de los hijos heredarán su alelo funcional y tendrán visión
cromática normal (CY).
madre
descendencia femenina
descendencia masculina
espermatozoide
X
C
X
C
X
c
X
c
X
C
X
C
Y
Y
X
C
X
C
X
C
X
c
Y
Y
X
C
X
c
(a) Visión a color normal
(b) Ceguera a los colores rojo y verde
(c) Hijos esperados de un hombre con visión cromática normal
(
CY
)
y
una mu
j
er heteroci
g
ota
(
Cc
)
El individuo no diferencia el rojo del verde
óvulos
padre
FIGURA 10-17 Herencia ligada a los cromosomas
sexuales de la ceguera al color Las rejillas cromáticas sirven
para comparar (a) la visión normal al color y (b) la deficiencia de
la visión de los colores rojo y verde. Por lo regular, una persona
afectada no es realmente “ciega” al color, pues ve la mayor parte
de los colores que ven las personas normales, pero no tan bien.
(c) Cuadro de Punnett que muestra la herencia de la ceguera al
color de una mujer heterocigota (Cc) a sus hijos.

186 UNIDAD 2 Herencia
el cabello rizado; dos copias del alelo C
2
producen cabello lacio.
Los heterocigotos, con el genotipo C
1
C
2
, tienen cabello ondulado.
Si dos personas de cabello ondulado se casan, pueden tener hijos
con cualquiera de los tres tipos, con probabilidades de una cuarta
parte rizado (C
1
C
1
), un medio ondulado (C
1
C
2
) y una cuarta parte
lacio (C
2
C
2
).
Un gen único puede tener múltiples alelos
Los alelos surgen por mutación y el mismo gen en diferentes indi-
viduos puede tener mutaciones distintas que produzcan, cada una,
un nuevo alelo. Por consiguiente, aunque un individuo puede tener
cuando más dos alelos diferentes, una especie puede tener alelos
múltiples de muchos de sus genes. Hay alelos múltiples para mu-
chos trastornos genéticos humanos, como el síndrome de Marfan,
la distrofia muscular de Duchenne (véase el apartado “Guardián
de la salud: Distrofia muscular” en las páginas 194-195) y la fibro-
sis quística (véase el estudio de caso del capítulo 12).
Los tipos de sangre humana son un ejemplo de alelos múl-
tiples de un único gen, con un añadido al esquema de la herencia.
Los tipos sanguíneos A, B, AB u O son resultado de tres alelos de
un solo gen en el cromosoma 9 (por simplicidad, los llamare-
mos alelos A, B y o). Este gen codifica la enzima que agrega mo-
léculas de carbohidratos a los extremos de glucoproteínas que se
proyectan de la superficie de los glóbulos rojos. Los alelos A y B
codifican las enzimas que agregan diferentes carbohidratos a las
glucoproteínas (llamaremos a las glucoproteínas resultantes A y
B). El alelo o codifica una enzima no funcional que no agrega
ninguna molécula de carbohidrato.
Una persona puede tener uno de seis genotipos: AA, BB,
AB, Ao, Bo u oo (
Tabla 10-1). Los alelos A y B son dominantes
sobre o; por tanto, las personas con genotipos AA o Ao tienen
únicamente glucoproteínas de tipo A y su tipo de sangre es A.
Quienes tienen los genotipos BB o Bo sintetizan únicamente las
glucoproteínas de tipo B y tienen tipo de sangre B. Los individuos
homocigotos recesivos oo carecen de ambos tipos de glucoproteí-
nas y tienen sangre tipo O. En las personas con tipo de sangre AB
están presentes las dos enzimas, así que la membrana plasmática
de sus glóbulos rojos tiene glucoproteínas A y B. Cuando los he-
terocigotos expresan fenotipos de los dos homocigotos (en este
caso, glucoproteínas A y B) el esquema de la herencia se llama
codominancia y se dice que los alelos son codominantes uno
del otro.
Las personas forman anticuerpos del tipo de las gluco-
proteínas que no tienen. Estos anticuerpos son proteínas en el
plasma sanguíneo que se unen a glucoproteínas extrañas cuando
reconocen diversas moléculas de carbohidratos. Los anticuerpos
hacen que los glóbulos que llevan las glucoproteínas extrañas se
agrupen y se rompan. Los grupos resultantes y fragmentos pue-
den obstruir vasos sanguíneos pequeños y dañar órganos vitales,
como el cerebro, corazón, pulmones o riñones. Esto significa que
el tipo de sangre debe ser identificado y concordado con preci-
sión antes de hacer una transfusión sanguínea.
El tipo de sangre O, que no tiene carbohidratos, no es
atacado por anticuerpos en sangre A, B ni AB, así que puede ser
transfundido con seguridad a los otros tipos (los anticuerpos pre-
sentes en la sangre transfundida quedan demasiado diluidos para
causar problemas). A las personas con tipo de sangre O se les
llama “donadores universales”. Pero la sangre tipo O lleva anti-
cuerpos de las glucoproteínas A o B, así que los individuos de tipo
10.8
¿LAS LEYES DE LA HERENCIA
DE MENDEL SE APLICAN EN
TODOS LOS RASGOS?
En nuestra exposición de la herencia, hasta este momento, hemos
hecho algunas suposiciones simplificadoras: que cada rasgo está
controlado completamente por un gen único, que sólo hay dos
posibles alelos en cada gen y que un alelo es completamente domi-
nante respecto de otro, que es recesivo. Sin embargo, casi todos los
rasgos tienen influencias más variadas y sutiles.
Dominancia incompleta: el fenotipo
de los heterocigotos es intermediario
entre los fenotipos de los homocigotos
Cuando uno de los alelos es completamente dominante respecto
a otro, los heterocigotos con un alelo dominante tienen el mismo
fenotipo que los homocigotos con dos alelos dominantes (véan-
se las figuras 10-8 y 10-9). Sin embargo, las relaciones entre ale-
los no siempre son simples. Cuando el fenotipo heterocigoto es
intermedio de dos fenotipos homocigotos, el esquema de la he-
rencia se llama dominancia incompleta. En los seres humanos,
la textura del cabello está influida por un gen con dos alelos que
no son completamente dominantes, a los que llamaremos C
1
y C
2
(
FIGURA 10-18). Una persona con dos copias del alelo C
1
tiene
*
*
ʼnYXORV
*
SDGUH
*
*
*
*
*
*
*
*
*
*
*
*
PDGUH
*
HVSHUPDWR]RLGHV
FIGURA 10-18 Dominancia incompleta La herencia de la
textura del pelo en los seres humanos es un ejemplo de dominancia
incompleta. En tales casos, denotamos con letras mayúsculas ambos
alelos, aquí C
1
y C
2
. Los homocigotos pueden tener cabello rizado
(C
1
C
1
) o lacio (C
2
C
2
). Los heterocigotos (C
1
C
2
) tienen el pelo ondulado.
Los hijos de un hombre y una mujer de pelo rizado pueden tener el
pelo rizado, lacio u ondulado, en la proporción aproximada de una
cuarta parte rizado, una mitad ondulado y una cuarta parte lacio.
Patrones de la herencia Capítulo 10 187
O sólo pueden recibir transfusiones de sangre del mismo tipo. En
la tabla 10-1 se resumen los tipos de sangre y las características de
las transfusiones.
Muchos rasgos están influidos por varios genes
Si miras a tus compañeros, verás que tienen distinta estatura, color
de piel y complexión. Estos rasgos no están gobernados por genes
únicos, sino por la interacción de dos o más genes, así como por
las interacciones con el entorno. Muchos rasgos, como la estatura,
peso, color de ojos o de piel, pueden tener varios fenotipos, o bien,
mostrar variaciones continuas que no pueden separarse en catego-
rías distintas y bien definidas. Lo anterior es un ejemplo de heren-
cia poligénica, una forma de herencia en la que la interacción de
dos o más genes contribuye a un fenotipo único.
Como te imaginarás, cuantos más genes contribuyan a un
rasgo único, mayor es el número de fenotipos y más estrechas las
distinciones entre ellos. Cuando tres o más pares de genes contri-
buyen a un rasgo, las diferencias entre fenotipos son muy peque-
ñas. Por ejemplo, aunque nadie entiende completamente cómo
se hereda el color de la piel en los seres humanos, probablemente
este factor está controlado por al menos tres genes, cada uno con
pares de alelos de dominancia incompleta (
FIGURA 10-19a). Si
el entorno también contribuye significativamente al rasgo (por
ejemplo, la exposición a la luz solar), hay prácticamente una va-
riación continua en el fenotipo (
FIGURA 10-19b).
Genes únicos tienen múltiples
efectos en un fenotipo
Acabamos de ver que un fenotipo único puede ser el resultado de
la interacción de varios genes. Lo contrario también ocurre: es co-
mún que genes únicos tengan múltiples efectos fenotípicos, un fe-
nómeno llamado pleiotropía. Un buen ejemplo es el gen SRY del
cromosoma Y. El gen SRY codifica una proteína que activa otros
genes, los cuales codifican proteínas que estimulan el desarrollo
masculino de un embrión. Por la influencia de los genes que activa
la proteína SRY, los órganos sexuales se convierten en testículos.
A continuación, los testículos producen hormonas sexuales que
estimulan el desarrollo de los órganos reproductores masculinos
internos y externos, como la próstata y el pene.
El ambiente influye en la expresión de los genes
Un organismo no es únicamente la suma de sus genes. Además
de su genotipo, el ambiente en el que vive afecta profundamen-
te su genotipo. Un sorprendente ejemplo del efecto del ambiente
en la acción de los genes se encuentra en los gatos siameses.
Todos los gatos siameses nacen con el pelaje claro, pero en las
primeras semanas, sus orejas, nariz, garras y cola se oscurecen
(
FIGURA 10-20). De hecho, un gato siamés puede tener el ge-
notipo del pelaje oscuro en todo el cuerpo, pero la enzima que
produce el pigmento oscuro se inactiva a temperaturas de más
de 34°C. Antes de nacer, las crías están tibias dentro del útero
materno, así que los gatitos recién nacidos tienen todo el cuerpo
cubierto de pelaje claro. Cuando nacen, las orejas, nariz, garras
y cola se enfrían más que el resto del cuerpo y ahí se produce el
pigmento.
Casi todas las influencias ambientales son más complica-
das y sutiles que esto. Las influencias ambientales complejas son
muy comunes en las características humanas. Los efectos ambien-
tales de la exposición al Sol modifican el rasgo poligénico del
color de la piel (véase la figura 10-19b). La nutrición influye en
otro rasgo poligénico, el peso.
Las interacciones ambientales entre sistemas genéticos
com plejos y condiciones ambientales variadas pueden crear un
continuo de fenotipos que dificultan la separación exacta entre
los componentes genéticos y ambientales. El tiempo de la ges-
tación humana es prolongado y el número de hijos por pareja
es pequeño. Agréguense a estos factores las incontables y sutiles
formas en que responden las personas a su entorno y puede verse
por qué es tan difícil determinar la base genética precisa de rasgos
humanos complejos como la inteligencia o las dotes para la mú-
sica, la pintura o los deportes.
Tabla 10-1 Características de los grupos sanguíneos
Tipo de sangre Genotipo Glóbulos rojos
Tiene anticuerpos
en plasma para:
Puede recibir
sangre de:
Puede donar
sangre a:
Frecuencia
en Estados
Unidos
A AA o Ao Glucoproteína B A u O (sangre sin
glucoproteína B)
A o AB 40%
B BB o Bo
Glucoproteína A B u O (sangre sin
glucoproteína A)
B o AB
10%
AB AB
Ninguna
glucoproteína
A ni B
AB, A, B, O (receptor
universal)
AB
4%
O oo Glucoproteínas
A y B
O (sangre sin
glucoproteínas A ni B)
O, AB, A, B
(donador
universal)
46%
Glucoproteína A
Glucoproteína B
Glucoproteínas A y B
Sin glucoproteínas A ni B
Este documento contiene más páginas...
Descargar Completo
Transmision sináptica215.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.