Oxidación de los ácidos grasos
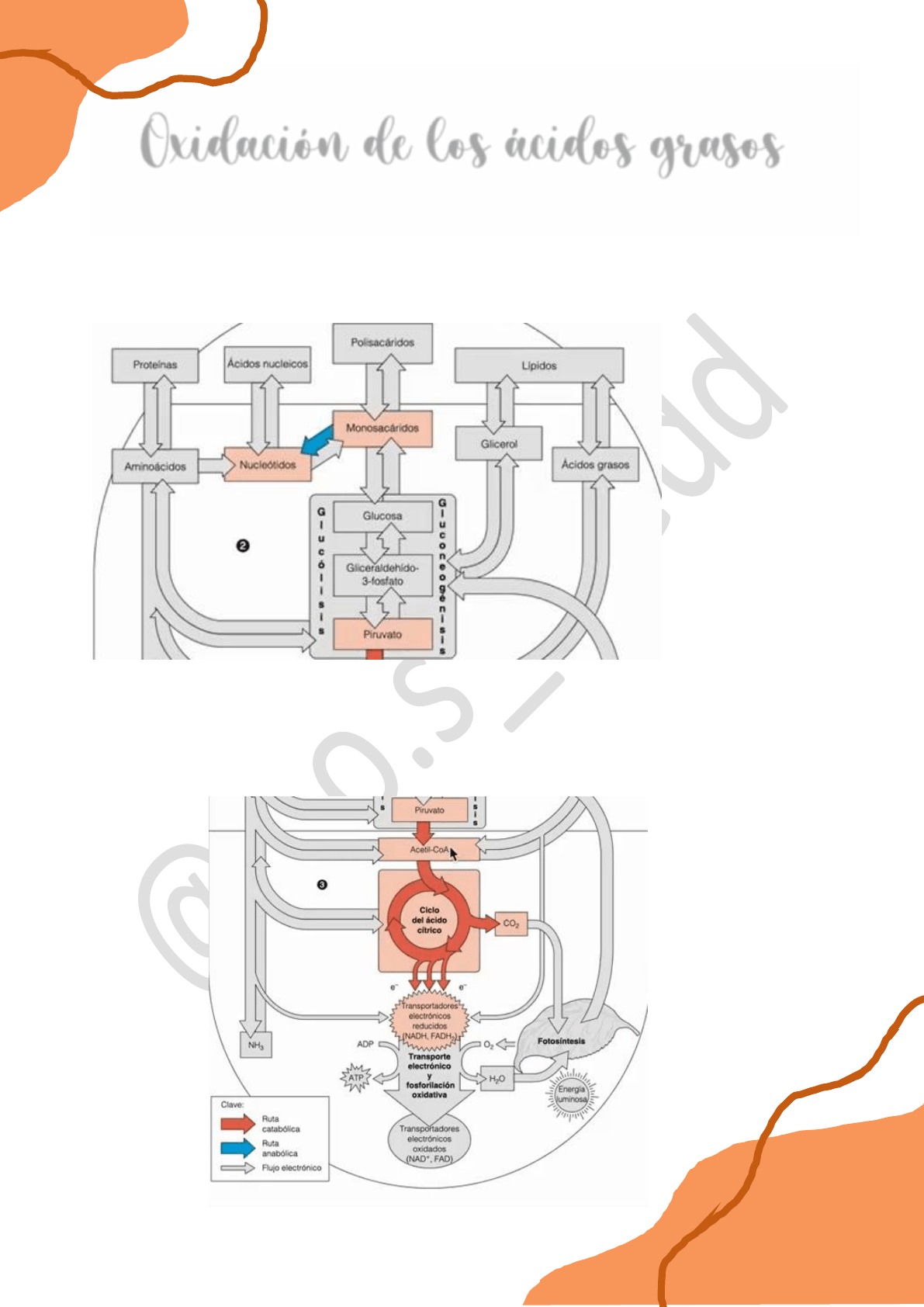
Habíamos visto en el módulo anterior que los polisacáridos pueden oxidarse por vía catabólica.
En principio se hidrolizan y dan monosacáridos y luego, estos monosacáridos ingresan al
interior celular y por un proceso denominado glucólisis, en 10 reacciones, la glucosa se termina
de oxidar a nivel de piruvato.
Luego este piruvato ingresa a la piruvato deshidrogenasa en el interior de la matriz
mitocondrial, se descarboxila formándose acetil coA, y ésta última termina de oxidarse
completamente en el ciclo de Krebs. Los dos carbonos que venían del piruvato y en definitiva
los carbonos que venían de la glucosa u otro monosacárido terminan totalmente oxidados a
CO2.
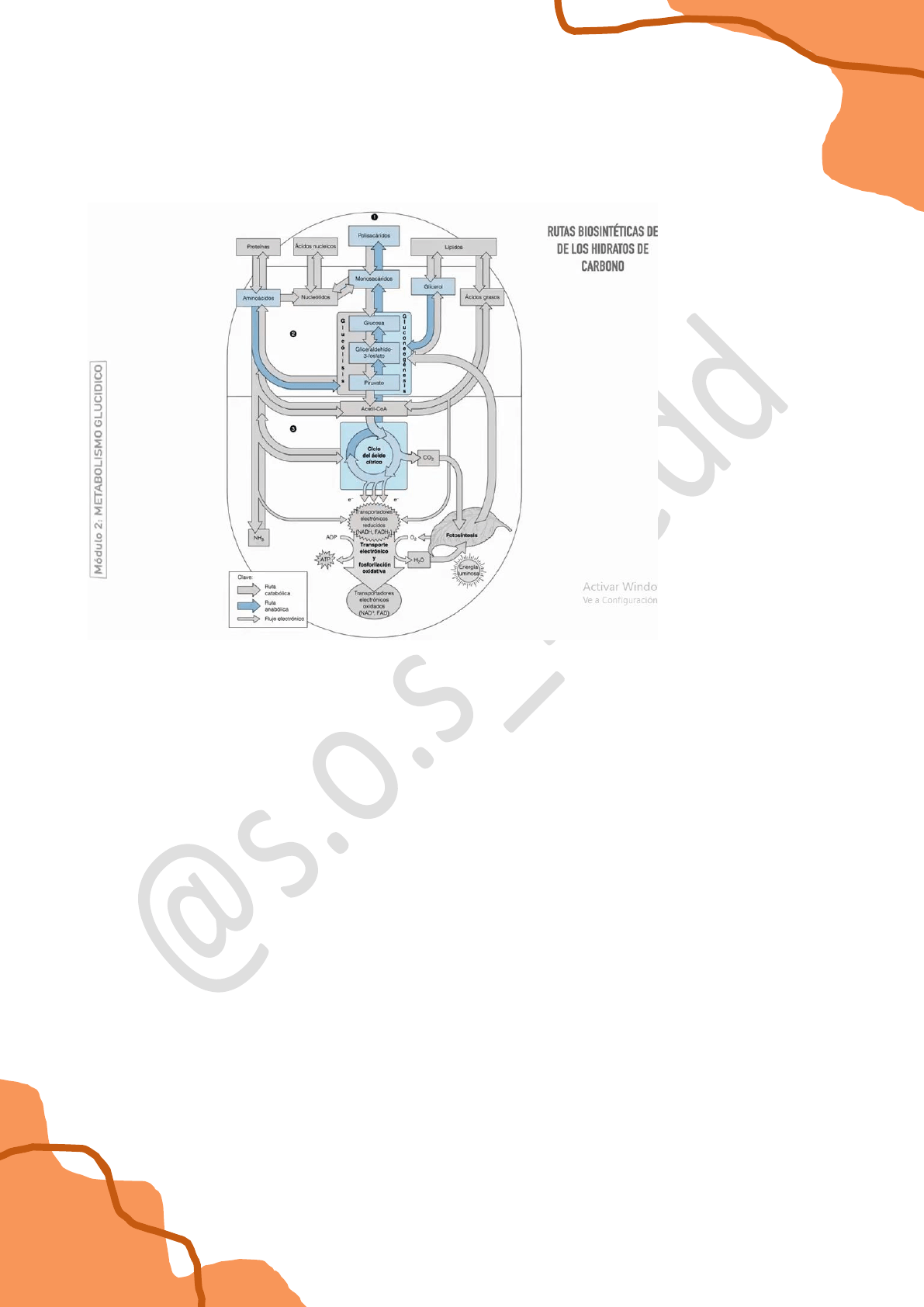
También habíamos visto que no sólo existían vías catabólicas, sino que también teníamos vías
anabólicas, en donde el proceso era diferente, había una construcción a partir de una
reducción biológica del carbono, se sintetizaba piruvato y luego este piruvato podía
anabolizarse nuevamente a glucosa.
En el caso de los polisacáridos la oxidación o la reducción de los mismos para catabolizarse o
par anabolizarse respectivamente, es simple por el hecho de que son moléculas que están
bastante oxidadas. Si uno recuerda la fórmula de la glucosa: C6H12O6 puede ver que hay
tantos oxígenos como carbonos en la molécula.
Sin embargo para el caso de los lípidos, estos compuestos están muy reducidos. Hay una
enorme cantidad de carbonos e hidrógenos y entonces cuando estos compuestos y
particularmente los ácidos grasos, se oxidan en el interior mitocondrial para dar acetil coA,
esta energía de oxidación es muy grande, se almacena en transportadores electrónicos
reducidos y luego cuando estos transportadores electrónicos se reoxidan en la cadena de
electrones generan una gran cantidad de protones y esos protones cuando re-ingresan a la
matriz mitocondrial en el proceso conocido como fosforilación oxidativa se genera una enorme
cantidad de ATP.
¿De dónde provienen estos ácidos grasos?
Principalmente provienen de la dieta, constituida básicamente por tag en sus formas más
comunes: aceites y grasas, y también fosfolípidos.
Los aceites y las grasas al ser tag tienen 3 colas de ácidos grasos esterificadas al glicerol, y
los fosfolípidos tienen 2 moléculas de ácidos grasos. Cuando se produce el proceso de
digestión de todos los componentes grasos que ingerimos, los tag aportan por cada molécula
de aceites o grasas, 3 moléculas de ácidos grasos y los fosfolípidos 2 moléculas de ácidos
grasos.
También los ácidos grasos pueden provenir de las reservas internas que tengamos en el tejido
adiposo. O bien pueden provenir de la síntesis celular endógena.
Digestión de lípidos
Muy resumidamente en qué consiste el proceso de
digestión: los lípidos empiezan a digerirse muy
precariamente en la boca a través de la acción de una
lipasa lingual o salival pero la mayor parte de la digestión
de los lípidos ocurre en el duodeno. Si recuerdan de
anatomía, entre la segunda, tercera y cuarta porción del
duodeno está la cabeza del páncreas y en la segunda
porción del duodeno desemboca el conducto biliar común
que trae la secreción exocrina del hígado que es la bilis, y
las secreciones pancreáticas que vienen del páncreas.
Entonces, los lípidos tienen un gran problemas al ser lípidos, por definición son insolubles en
agua, en cambio las proteínas que van a degradar a esos lípidos, naturalmente al ser proteínas
son solubles en medio acuoso, entonces se plantea un problema de heterogeneidad porque los
lípidos están en una fase y las proteínas que digieran a esos lípidos están en otra, y allí es
donde entra la acción biológica de la bilis. La bilis lo que hace es actuar como un detergente,
esta tiene iones, agua y básicamente las sales biliares que son las que actúan como
detergentes. Estas moléculas que están presentes en la bilis tienen una cara hidrofílica y una
cara hidrofóbica, entonces varias de estas moléculas se posicionan alrededor de las moléculas
lipídicas exponiendo su cara hidrófila hacia afuera y de esa manera logran emulsionar a las
moléculas lipídicas y en los intersticios que quedan por la yuxtaposición de estas moléculas de
ácidos biliares es por donde actúan las lipasas pancreáticas. Estas lipasas pancreáticas para
poder actuar necesitan de un cofactor que son las colipasas, y de esa manera en esta
interfase de emulsión entre la gota lipídica y la fase acuosa en donde están las enzimas se
produce la hidrólisis o degradación de los ácidos grasos.
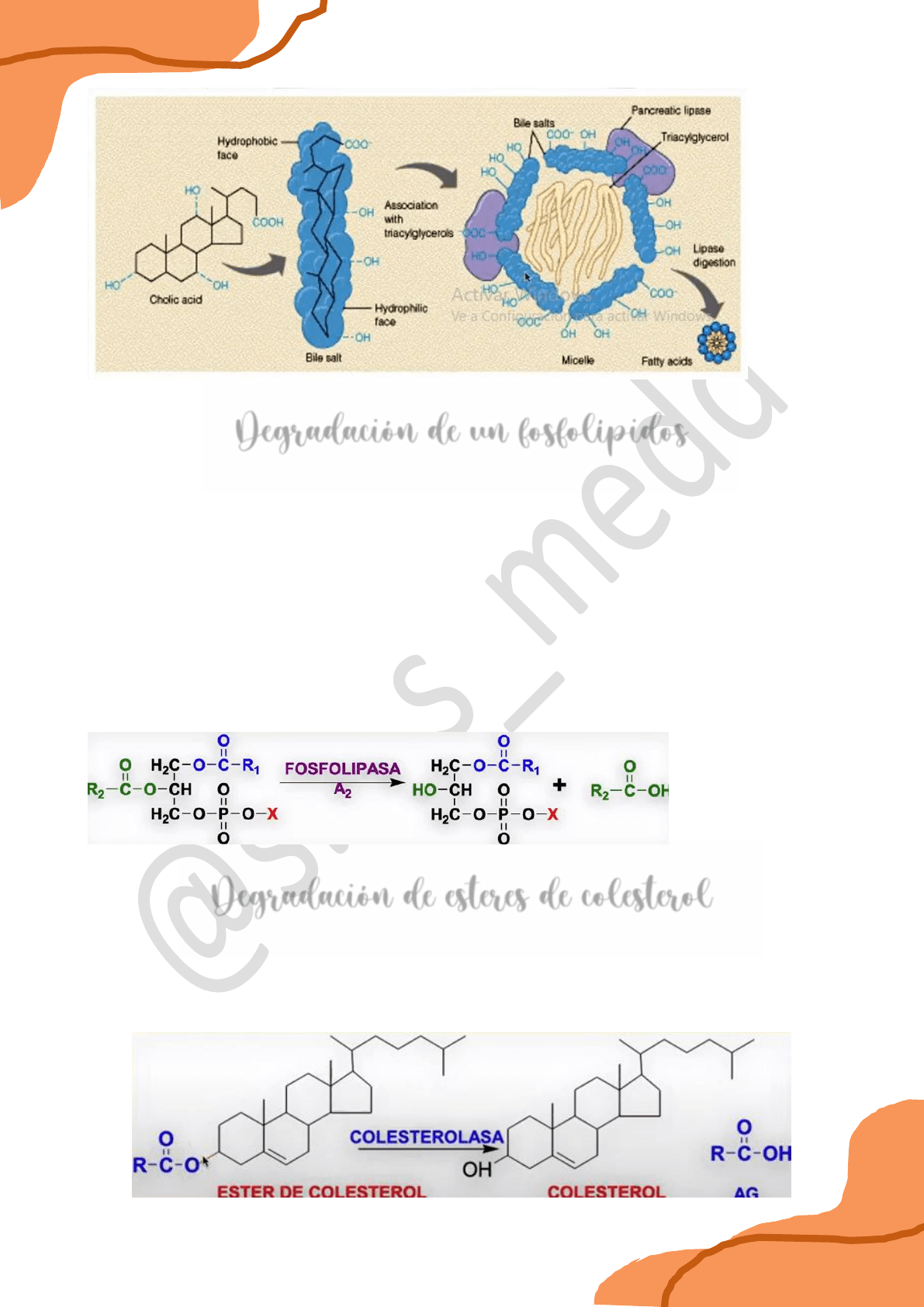
Degradación de un fosfolípidos
Son otra fuente importante de ácidos grasos y no actúa una lipasa sino una fosfolipasa.
Fosfolipasa A1: rompe la unión éster 1 del ácido graso al oxhidrilo 1 del glicerol.
Fosfolipasa A2: rompe la unión éster 2 del ácido graso al oxhidrilo 2 del glicerol.
Fosfolipasa C: hidroliza la unión del glicerol al grupo fosfato.
Fosfolipasa D.
De acuerdo a qué fosfolipasa esté actuando sobre el fosfolípido se obtienen distintos
productos de hidrólisis.
Degradación de esteres de colesterol
Donde actúa una acilcolesterolasa degradará la unión del ácido graso al oxhidrilo en posición 3
del colesterol. Al romperse la unión se libera colesterol por un lado y una molécula de ácido
graso por otro.

Degradación del glicerol
Una gliceroquinasa, es una enzima que fosforila al glicerol en posición 3 obteniéndose glicerol –
3 – fosfato y luego este glicerol-3-P en el carbono 2 donde tiene un grupo alcohol, es oxidado
a un grupo cetona por la acción de la glicerolfosfato deshidrogenasa, pasando a ser
dihidroxiacetona fosfato. Luego la dihidroxiacetona fosfato se oxida, otra vez, en el carbono
1 y el grupo alcohol se transforma en un grupo aldehído, obteniendo así el gliceraldehído
fosfato.
Si recuerdan del módulo anterior, el G3-P es el reactivo clave que constituye la oxidación de
la glucólisis, ya que puede oxidarse a uno 1,3 bifosfoglicerato, la cual es la reacción principal
de la glucólisis. En este caso se reduce el NAD+ a NADH,H+ y este NADH,H+ puede ser
captado por la lanzadera mitocondrial de glicerol-3-fosfato, y de esta manera trasladar este
equivalente de reducción del citosol hacia la matriz mitocondrial en forma de FADH2.
Lipolisis
Una vez que los ácidos grasos se liberan del glicerol o del colesterol para poder β oxidarse
previamente se deben activar, formando acil coA del ácido graso y en eucariotas pasar del
citosol a la mitocondria.
Tiene dos etapas:
Activación y transporte (con un aminoácido que se denomina carnitina).
B oxidación

Activación de un ácido graso
1. el oxígeno del grupo carboxilo de la molécula de ácido graso libre ataca a una molécula de
ATP y este se rompe liberando un pirofosfato y entonces el adenilato del ATP queda unido al
grupo acilo.
2. Posteriormente una molécula de coenzima A produce un mismo ataque, ya no con oxígeno
sino con azufre, sobre el carbono 1 del grupo acilo, se rompe el adenilato del ácido graso y se
libera AMP y Se forma el acil coA correspondiente.
3. Una vez que se forma acil coA este va a ser transportado al interior de la mitocondria,
porque son impermeables a la membrana mitocondrial interna, a través de un sistema que se
denomina lanzadera de carnitina o sistema transportador de carnitina.
Transportador de carnitina
El grupo acilo unido a la coenzima A se etioliza, es decir se rompe el grupo tiol de la coenzima
y el grupo acilo queda unido a la carnitina.
La carnitina es un aminoácido pero no es un α aminoácido como los que están presentes en las
proteínas sino que es un γ aminoácido, ya que el grupo amino está unido al carbono γ de la
carnitina, entonces esta molécula es un aminoácido NO proteico.
El grupo acilo queda unido a la carnitina y de esta manera es transportado al interior de la
mitocondria a través de una enzima llamada carnitina acil transferasa I y en la cara interna de
la membrana mitocondrial interna una acil carnitina II vuelve a romper esta unión del grupo
acilo con la carnitina y se vuelve a unir el grupo acilo a una coenzima A, de esta manera
tenemos nuevamente acil coA del ácido graso idéntico al que estaba afuera.
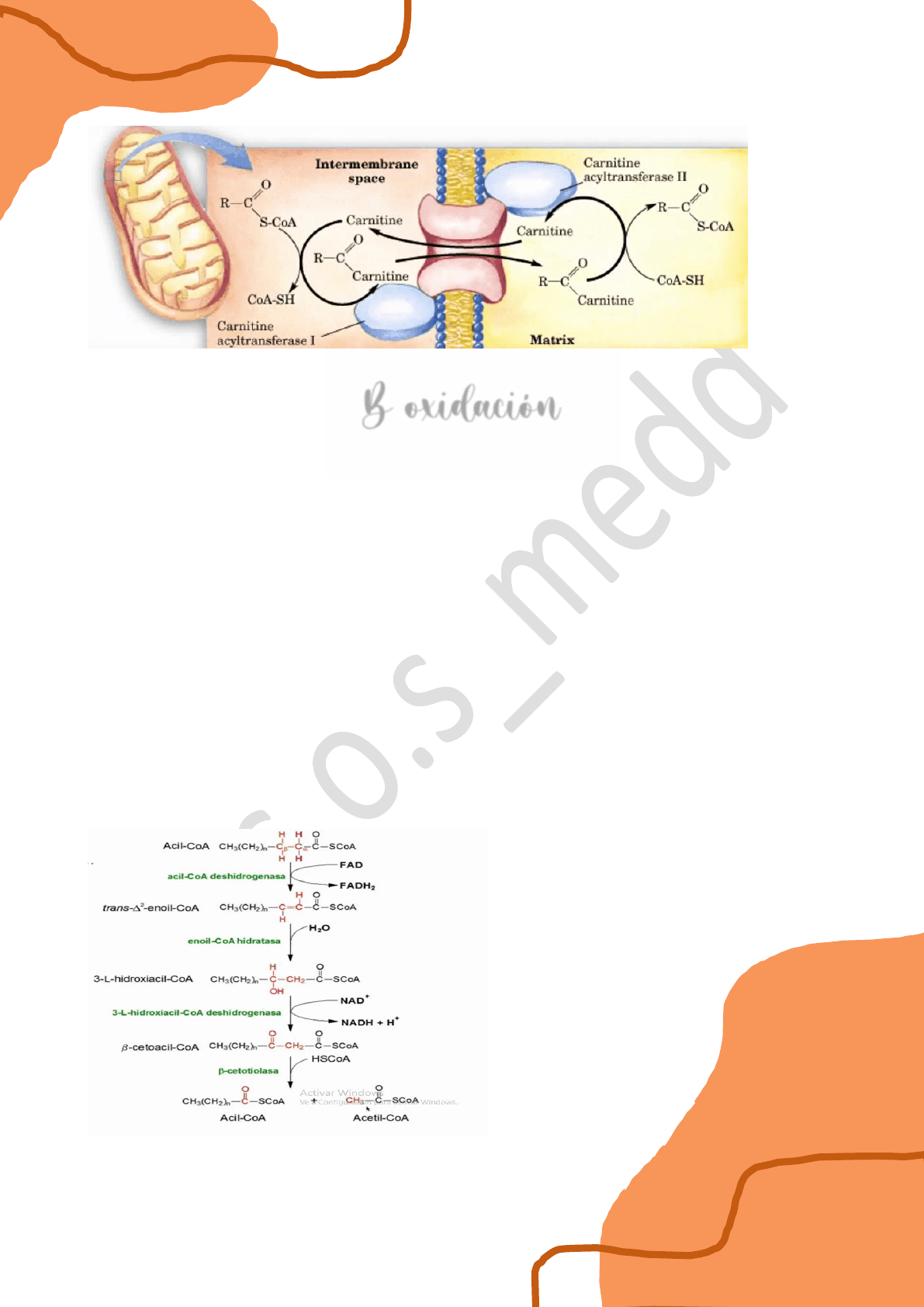
B oxidación
Consta de 4 etapas y es un proceso cíclico.
1. el acil coA correspondiente se oxida entre el carbono α y β. Entonces ambos que tienen un
enlace simple C-C se oxidan a un alqueno (se produce un doble enlace), reduciendo al FAD+ a
FADH2.
2. Luego el doble enlace se hidrate. Un oxhidrilo de la molécula de agua se adiciona sobre el
carbono β y el protón sobre el carbono α (CH2).
3. Luego el grupo alcohol secundario que se encuentra en el carbono 3 o β se oxida a una
cetona, reduciendo el NAD+ a NADH, H+.
4. Finalmente el carbono 2 o α va a romperse a través de un proceso que se llama
fragmentación tiólica, y entonces una molécula de coenzima A ataca la unión obteniéndose un
acil coA con dos carbonos menos y un acetil coA. El ciclo vuelve a iniciarse y en cada proceso
se obtiene un acetil coA y un acil coA con dos carbonos menos.

¿Cuántas vueltas van a dar la β oxidación para oxidar totalmente a un ácido graso y
convertirlo totalmente en acetil coA? Va a depender del número de átomos de carbono que
tenga ese ácido graso.
¿Qué ocurre cuando tenemos que oxidar ácidos grasos que
tienen un número impar de átomos de carbonos?
Una enzima que se llama propionilcoA carboxilasa introduce un carbono más a través de un
bicarbonato con gasto de una molécula de ATP. Entonces el propionil coA se elonga con un
carbono más, formándose metilmalonilcoA y en la siguiente reacción hay una epimerización, en
el cual el grupo carboxilo del carbono 3 pasa a ser el carbono 4 y se obtiene succinil-coA,
sigue la β oxidación normal como si fuera un acil coA de 4 átomos de carbono, es decir en el
siguiente paso se convierte en dos unidades de acetil coA.
Otra complicación adicional que podemos tener Es que tengamos ácidos grasos con una
insaturación o con muchas insaturaciones.
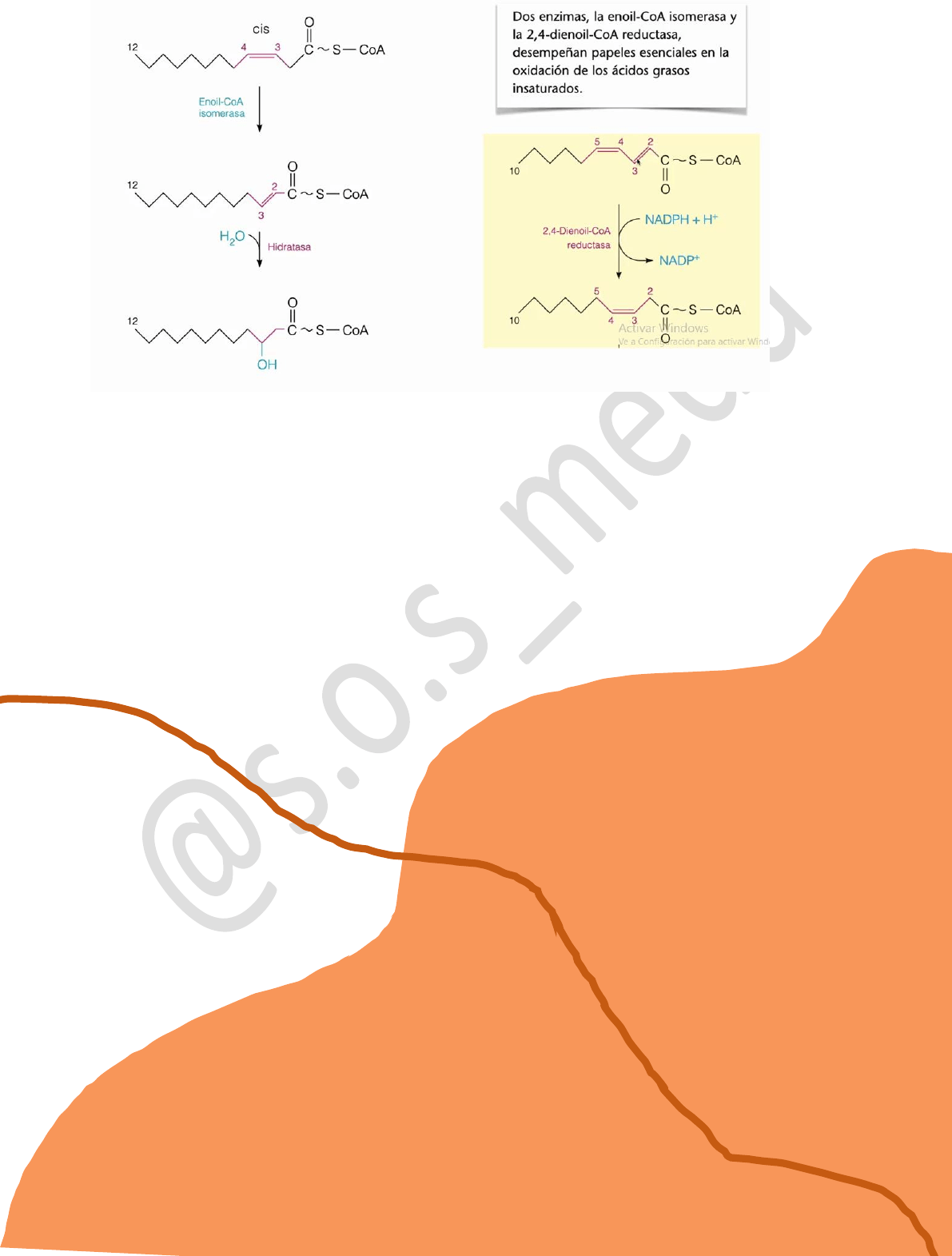
Si tenemos un doble enlace cis, la enzima que produce la hidratación siempre produce la
introducción del oxhidrilo en el carbono más alejado del grupo carbonilo entonces Lo que hace
la célula es mover el doble enlace más próximo al grupo carbonilo por medio de la enzima enoil-
coA isomerasa, por lo que ahora sí se obtiene el doble enlace entre el carbono α y β, Entonces
en la siguiente reacción, una hidratasa introduce un oxhidrilo en el carbono β y se obtiene el
βhidroxiacilcoA por lo que la β oxidación transcurre normalmente.
Si tenemos varios dobles enlaces Allí van a tomar preponderancia dos enzimas: enoil-coA
isomerasa y 2,4 dienoil-coA reductasa. Lo que va a ocurrir es que El primer doble enlace más
cercano al carbonilo se reduce, oxidando el NADPH+ a NADP+ , desapareciendo el doble
enlace, y el doble enlace que le seguía se mueve. Ahora estamos en la situación que nos
encontrábamos anteriormente, la enoil-coA isomerasa mueve el doble enlace y obtenemos el
mismo resultado: β-hidroxiacilcoA.

oxidacion de acidos grasos desgrabado .pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.