
CAPÍTULO
Médula espinal: orga interna nización
18
La médula espinal proporciona inervación para el tronco y las
extremidades a través de los nervios espinales y sus ramificaciones
periféricas. Recibe fibras aferentes primarias de receptores periféricos
ubicados en estructuras somáticas y viscerales generalizadas y envía
axones motores al músculo esquelético. También contiene los cuerpos
celulares de todas las neuronas preganglionares responsables de la
inervación simpática del músculo cardíaco y liso y de las glándulas
secretoras, y de la inervación parasimpática del músculo liso en la parte
distal del intestino posterior, las vísceras pélvicas y los tejidos eréctiles de
los genitales externos. Muchas funciones corporales están reguladas en
un nivel inconsciente por conexiones reflejas intraespinales entre
neuronas aferentes y eferentes. Las vías ascendentes y descendentes
profusas unen la médula espinal con el cerebro,
consta de cuatro masas celulares enlazadas, los cuernos dorsal y ventral
derecho e izquierdo, que se proyectan dorsolateralmente y ventrolateralmente
hacia la superficie respectivamente. La sustancia gris que rodea
inmediatamente el canal central y une los dos lados constituye las comisuras
grises dorsal y ventral. El asta dorsal es el sitio de terminación de las fibras
aferentes primarias que entran en la médula a través de las raíces dorsales de
los nervios espinales. La punta del asta dorsal está separada de la superficie
dorsolateral del cordón por un fascículo o trayecto delgado (de Lissauer) en el
que las fibras aferentes primarias ascienden y descienden una corta distancia
antes de terminar en la sustancia gris subyacente. El cuerno ventral contiene
neuronas eferentes cuyos axones salen de la médula espinal en las raíces
nerviosas ventrales. Un pequeño intermedio, o lateral, el cuerno está presente
en los niveles torácico y lumbar superior; contiene los cuerpos celulares de las
neuronas simpáticas preganglionares.
La sustancia gris espinal (fig. 18.2) es una mezcla compleja de cuerpos de células
neuronales, sus procesos y conexiones sinápticas, neuroglia y vasos sanguíneos. Las
neuronas de la materia gris son multipolares. Varían en tamaño y características
como la longitud y la disposición de sus axones y dendritas. Las neuronas pueden ser
intrasegmentarias, es decir, estar contenidas dentro de un solo segmento, o
intersegmentarias, es decir, sus ramificaciones se extienden a través de varios
segmentos.
CARACTERÍSTICAS Y RELACIONES EXTERNAS
La anatomía topográfica de la médula espinal, sus características
externas y sus relaciones se describen con más detalle en el capítulo 43.
En resumen, la médula se encuentra dentro del canal vertebral. Continúa
rostralmente con la médula oblonga, justo debajo del nivel del foramen
magnum (fig. 28.11) y termina caudalmente como el cono medular, que
es continuo con el fi lum terminale y está anclado al dorso del cóccix. El
cordón está envuelto por meninges espinales que se continúan con las
meninges craneales a través del foramen magnum. Aunque tiene una
sección transversal aproximadamente circular, el diámetro de la médula
espinal varía según el nivel; lleva dos ampliaciones, cervical y lumbar.
La médula espinal es esencialmente una estructura segmentaria, dando lugar a 31 nervios
espinales emparejados bilateralmente. Estos se adhieren al cordón como series lineales de raicillas
nerviosas ventrales y dorsales más pequeñas. Las raicillas dorsales contienen fibras nerviosas
aferentes y las raicillas ventrales contienen fibras eferentes (ver Fig.
15,4). Los grupos de raicillas adyacentes se fusionan para formar raíces nerviosas dorsales o
ventrales que cruzan el espacio subaracnoideo y se unen para formar nervios espinales
funcionalmente mixtos a medida que atraviesan los agujeros intervertebrales. Las raíces
dorsales tienen ganglios de la raíz dorsal que contienen los cuerpos celulares de las
neuronas aferentes primarias.
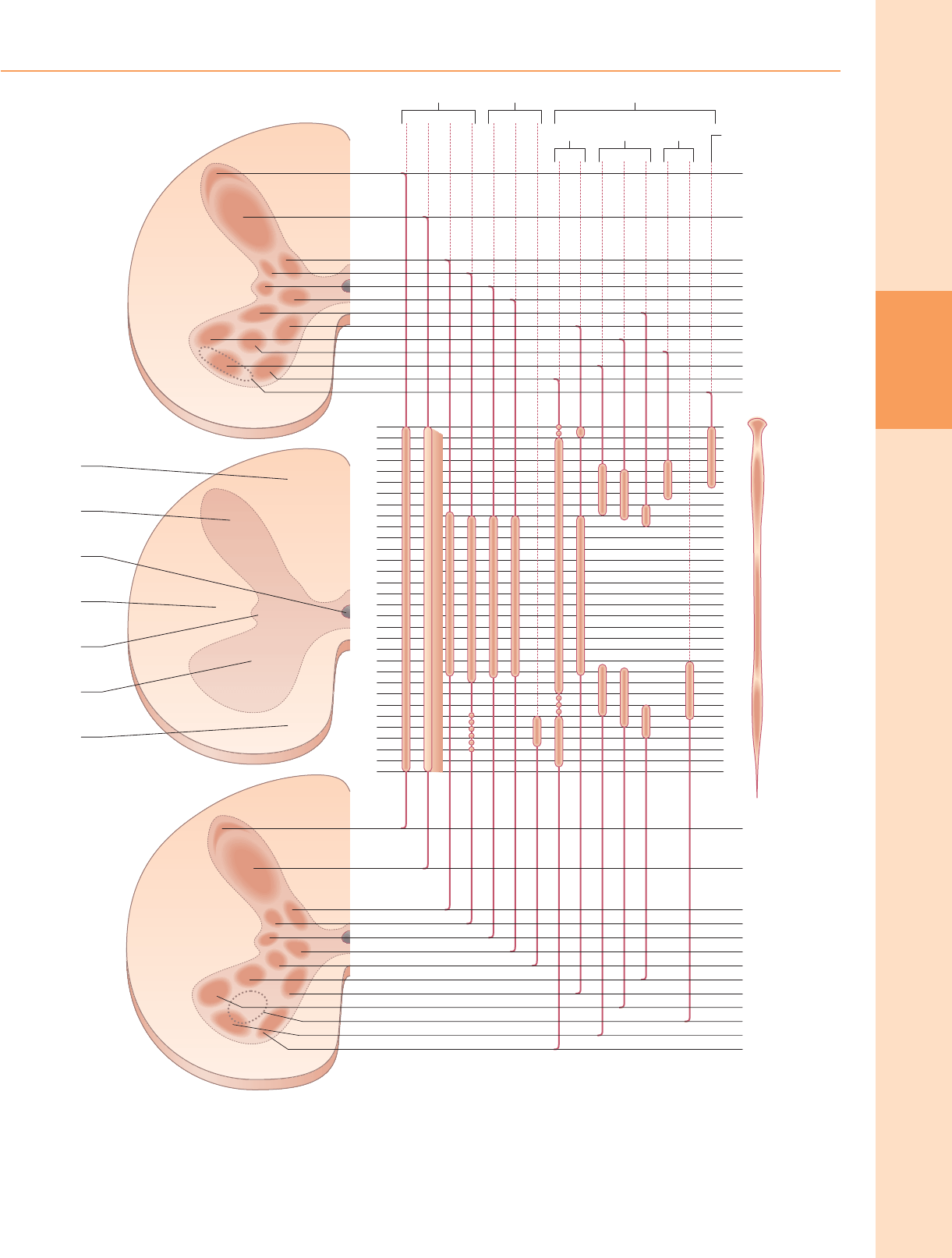
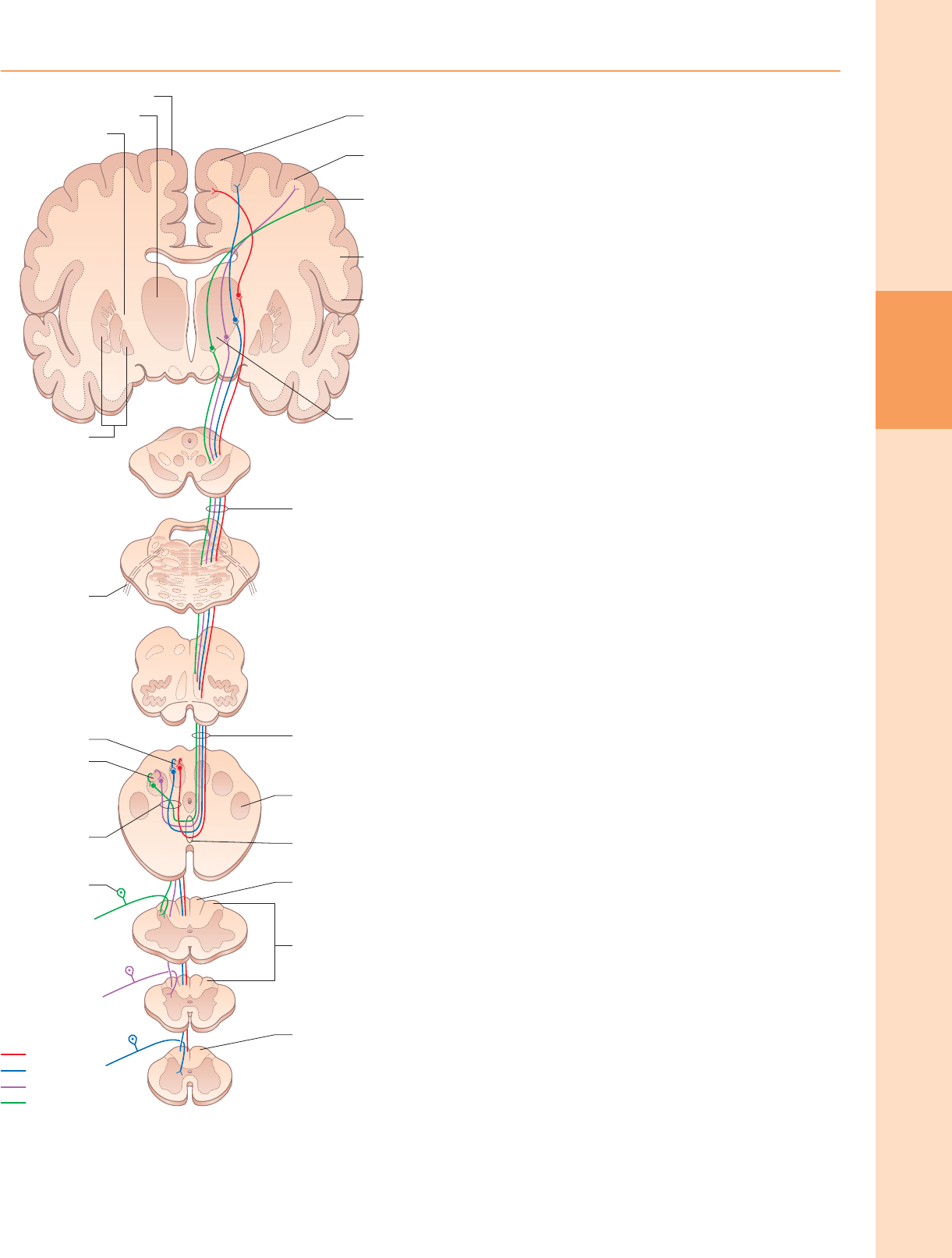
Grupos de células neuronales de la médula espinal.
Vista desde la perspectiva de su organización longitudinal columnar, la materia gris
de la médula espinal consiste en una serie de agrupaciones de células discontinuas
asociadas con sus correspondientes nervios espinales dispuestos segmentados. En
cualquier nivel de sección transversal particular, estos grupos de células a menudo
se consideran que se corresponden aproximadamente con una o más de diez capas
de células, conocidas como láminas de Rexed. Estas láminas se definen sobre la base
del tamaño neuronal, la forma, las características citológicas y la densidad y están
numeradas en una secuencia dorsoventral.
Las láminas I-IV corresponden a la parte dorsal del asta dorsal y son el sitio
principal de terminación de las terminales aferentes primarias cutáneas y sus
colaterales. Muchas rutas complejas de reflejos polisinápticos (ipsilateral,
contralateral, intrasegmental e intersegmental) parten de esta región, al igual que
muchas fibras largas del tracto ascendente que pasan a niveles superiores. Lamina I
(lámina marginalis) es una capa muy delgada con un límite mal definido en la punta
dorsolateral del asta dorsal. Tiene una apariencia reticular, lo que refleja su
contenido de haces entremezclados de fibras nerviosas finas y gruesas. Contiene
somas neuronales pequeños, intermedios y grandes, muchos de los cuales tienen
forma fusiforme. La lámina II, mucho más grande, consta de pequeñas neuronas
densamente agrupadas, responsables de su apariencia oscura en las secciones
teñidas con Nissl. Con manchas de mielina la lámina II se distingue de forma
característica de las láminas adyacentes por la falta casi total de fibras mielinizadas.
La lámina II corresponde a la sustancia gelatinosa. La lámina III consta de somas que
son en su mayoría más grandes, más variables y menos empaquetados que los de la
lámina II. También contiene muchas fibras mielinizadas. Algunos trabajadores
consideran que la sustancia gelatinosa contiene parte o la totalidad de la lámina III
así como de la lámina.
II. El núcleo propio mal definido del asta dorsal corresponde a algunos de los
constituyentes celulares de las láminas III y IV. La lámina IV es una zona
heterogénea, gruesa, poco compacta, impregnada de fibras. Sus somas neuronales
varían considerablemente en tamaño y forma, desde pequeños y redondos, pasando
por intermedios y triangulares, hasta muy grandes y estrellados.
Las láminas V y VI se encuentran en la base del asta dorsal. Reciben la mayoría de
las terminales de las aferentes primarias propioceptivas, proyecciones
corticoespinales profusas de la corteza motora y sensorial e información de los
niveles subcorticales, lo que sugiere su participación en la regulación del
movimiento. La lámina V es una capa gruesa, divisible en un tercio lateral y dos
tercios medial. Ambos tienen una población celular mixta, pero el primero contiene
muchos somas prominentes bien teñidos entrelazados por numerosos haces de
fibras transversales, dorsoventrales y longitudinales. La lámina VI es más prominente
en las ampliaciones de las extremidades. Tiene una densa
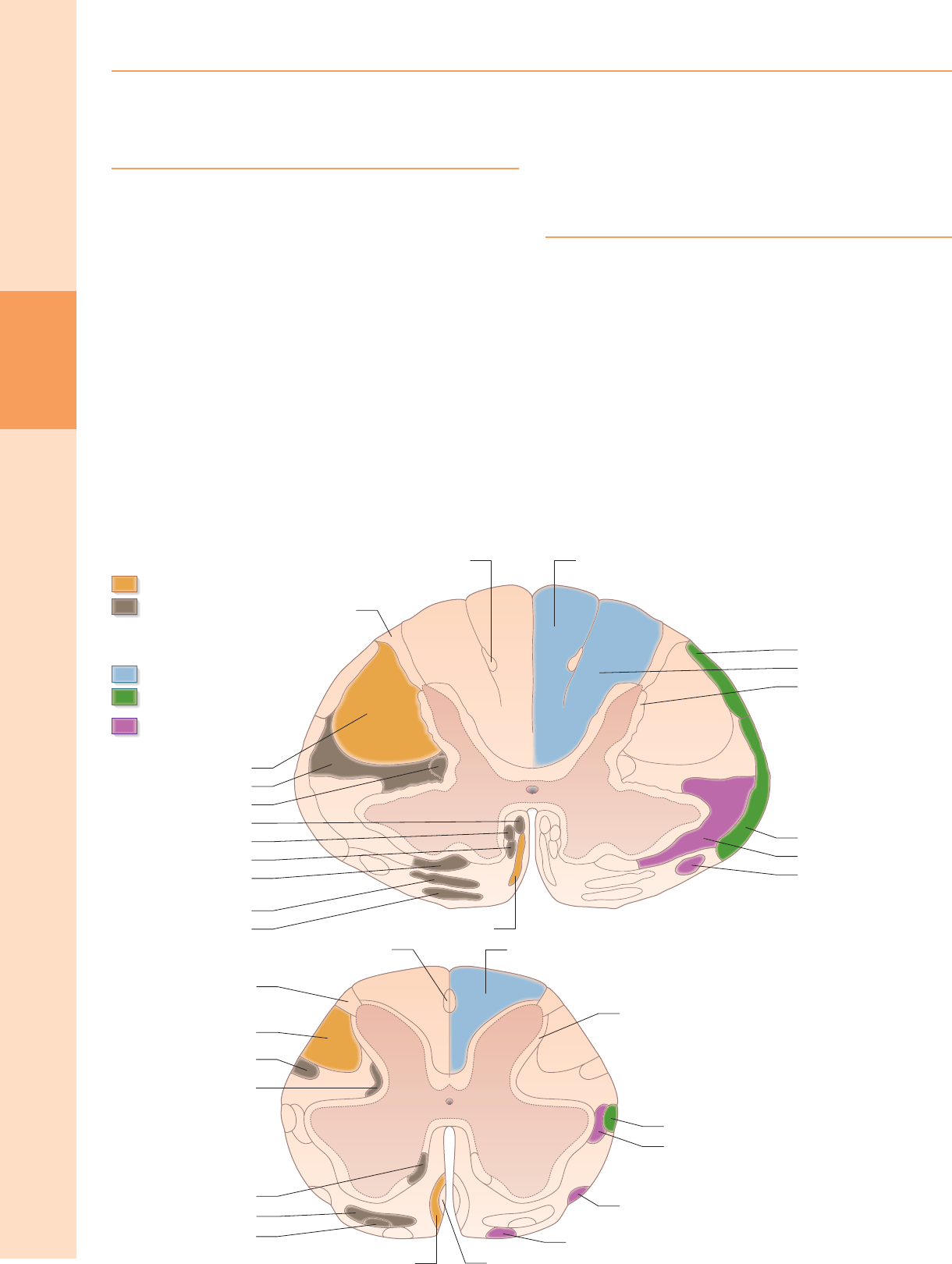
ORGANIZACIÓN INTERNA
En la sección transversal, la médula espinal está dividida de forma incompleta
en mitades simétricas por un tabique mediano dorsal (posterior) y un surco
mediano ventral (anterior) (fig. 18.1). Consiste en una capa externa de materia
blanca y un núcleo interno de materia gris; sus tamaños relativos y su
configuración varían según el nivel. La cantidad de materia gris refleja el
número de neuronas presentes; es proporcionalmente mayor en las
ampliaciones cervicales y lumbares, que contienen las neuronas que inervan
las extremidades. La cantidad absoluta de sustancia blanca es mayor en los
niveles cervicales y disminuye progresivamente en los niveles inferiores,
porque los tractos descendentes desprenden fibras a medida que descienden
y los tractos ascendentes acumulan fibras a medida que ascienden.
Un canal central diminuto, revestido por epitelio cilíndrico cilíndrico
(ependyma) y que contiene líquido cefalorraquídeo (LCR), se extiende por toda
la longitud de la médula espinal que se encuentra en el centro de la sustancia
gris espinal. Rostralmente, el canal central se extiende hacia la mitad caudal
del bulbo raquídeo y luego se abre hacia el cuarto ventrículo.
MATERIA GRIS ESPINAL
En tres dimensiones, la sustancia gris espinal tiene la forma de una columna
acanalada (fig. 43.1F). En la sección transversal, la columna se describe a menudo
como "en forma de mariposa" o se asemeja a la letra "H" (Fig. 18.1). Eso
257
Traducido del inglés al español - www.onlinedoctranslator.com
MÉDULA ESPINAL: ORGANIZACIÓN INTERNA
Surco mediano dorsal
Cervical
A
Fisura ventral mediana
Funículo dorsal
Torácico
Funículo lateral
B
Funículo ventral
Lumbar
Cuerno dorsal
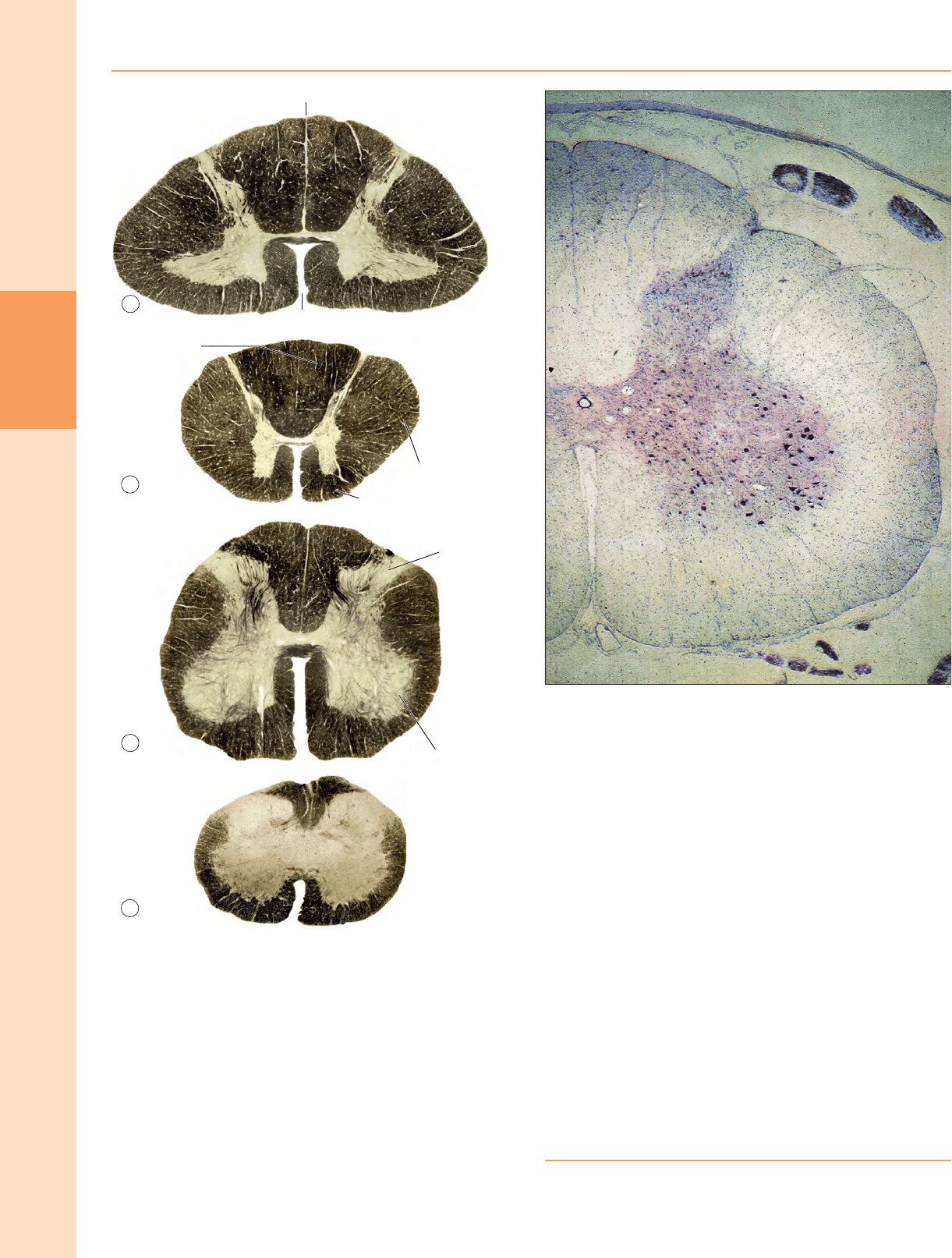
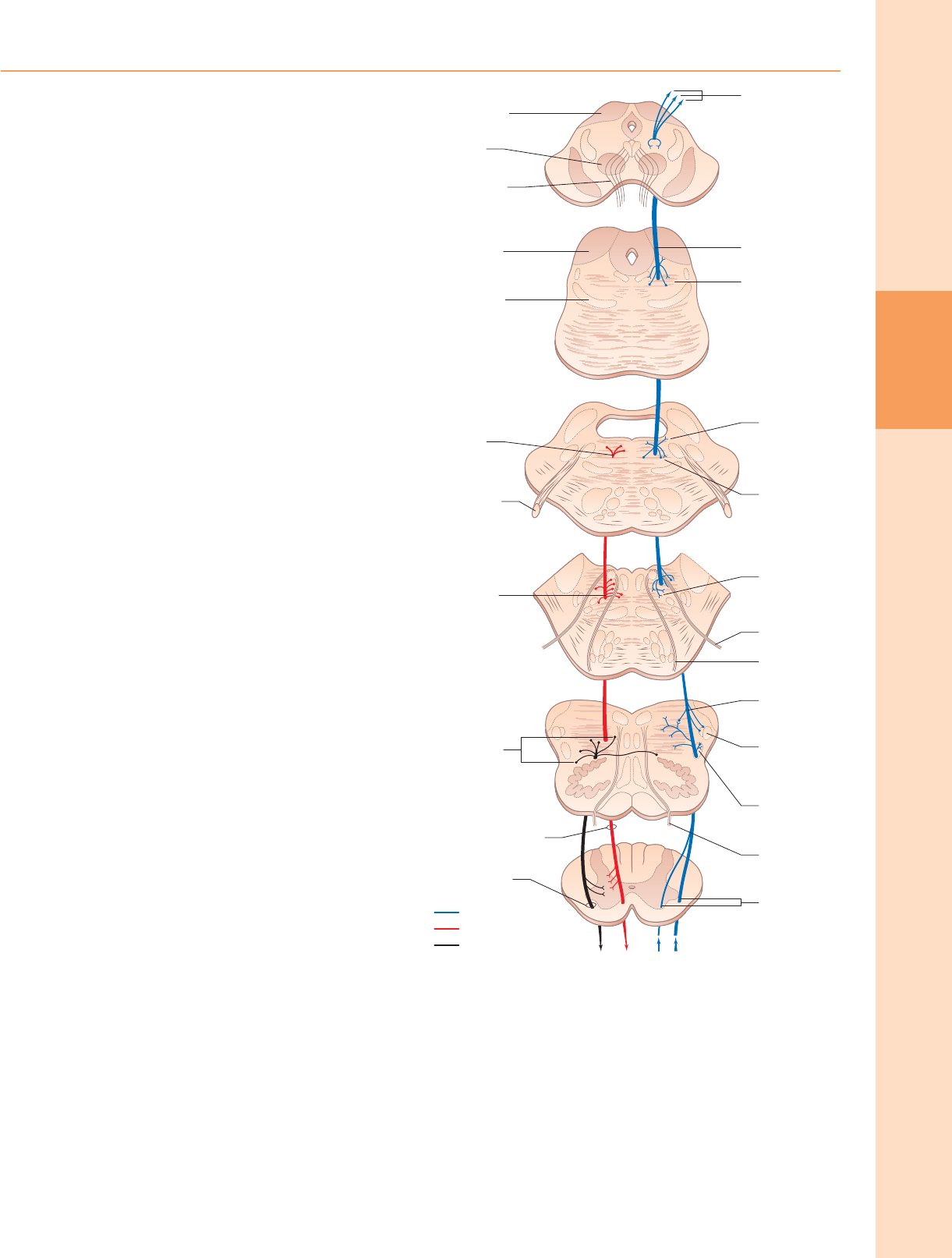
Figura 18.2 Sección transversal de la médula espinal a nivel lumbar medio. Las
neuronas motoras más grandes de la columna gris ventral están agrupadas
visiblemente. Teñido con violeta rápido de cresilo.
C
Cuerno ventral
Sacro
las láminas adyacentes, muchas fibras comisurales de la lámina VIII
contralateral y conexiones descendentes de los tractos intersticioespinal,
reticuloespinal y vestibuloespinal y el fascículo longitudinal medial. Los axones
de estas interneuronas influyen α actividad de la neurona motora
bilateralmente, tal vez directamente, pero más probablemente por excitación
de pequeñas γ neuronas motoras que suministran fibras eferentes a los husos
musculares. Lamina IX es una matriz compleja de células que consta de α y γ
neuronas motoras y muchas interneuronas. El grande α las neuronas motoras
inervan las placas terminales motoras de las fibras musculares extrafusales en el
músculo estriado. Las técnicas de grabación han demostrado ser tónicas y fásicas. α
neuronas motoras. Los primeros tienen una menor tasa de disparo y menor
velocidad de conducción y tienden a inervar las unidades musculares de tipo S. Estos
últimos tienen una mayor velocidad de conducción y tienden a suministrar unidades
musculares de contracción rápida (tipo FR, FF). El pequeño γ las neuronas motoras
dan lugar a axones eferentes de pequeño diámetro (fibras fusimotoras), que inervan
las fibras musculares intrafusales en los husos musculares. Hay varios tipos
funcionalmente distintos de γ neurona motora. Las respuestas "estáticas" y
"dinámicas" de los husos musculares tienen controles separados mediados por fibras
fusimotoras estáticas y dinámicas, que se distribuyen de diversas formas a las fibras
de la cadena nuclear y de la bolsa nuclear.
La lámina X rodea el canal central y está formada por las comisuras
grises dorsal y ventral.
D
Figura 18.1 Secciones transversales a través de la médula espinal a niveles
representativos. Aproximadamente × 5. (Figura mejorada por B Crossman.)
tinción del tercio medial de neuronas pequeñas y densamente empaquetadas y dos
tercios laterales que contienen somas triangulares o estrellados más grandes, más
sueltos.
Las láminas VII-IX muestran una variedad de formas en el agrandamiento de las
extremidades. La lámina VII incluye gran parte del cuerno intermedio (lateral).
Contiene neuronas prominentes de la columna de Clarke (núcleo dorsal, núcleo
torácico, núcleo torácico) y agrupaciones de células intermediomediales e
intermediolaterales (fig. 18.3). La parte lateral de la lámina VII tiene amplias
conexiones ascendentes y descendentes con el mesencéfalo y el cerebelo (a través
de los tractos espinocerebeloso, espinotectal, espinorreticular, tectoespinal,
reticuloespinal y rubroespinal) y, por lo tanto, participa en la regulación de la postura
y el movimiento. Su parte medial tiene numerosas conexiones de reflejo
proprioespinal con la sustancia gris adyacente y los segmentos relacionados tanto
con el movimiento como con las funciones autónomas. La lámina VIII se extiende por
la base del asta ventral torácica, pero está restringida a su cara medial en los
agrandamientos de las extremidades. Sus neuronas muestran una mezcla
heterogénea de tamaños y formas, desde pequeñas hasta moderadamente grandes.
Lamina VIII es una masa de interneuronas proprioespinales. Recibe terminales de
Cuerno dorsal
El asta dorsal es una zona importante de terminación de las fibras aferentes
primarias, que entran en la médula espinal a través de las raíces dorsales de los
nervios espinales. Las fibras de la raíz dorsal contienen numerosas moléculas, que se
sabe o se sospecha que cumplen una función neurotransmisora o
neuromoduladora. Estos incluyen ácido glutámico, sustancia P, péptido relacionado
con el gen de la calcitonina (CGRP), bombesina, polipéptido intestinal vasoactivo
(VIP), colecistoquinina (CCK), somatostatina, dinorfina y angiotensina II.
258
Organización interna
Columna dorsal Columna lateral Columna ventral
Medio Lateral Central Accesorio
Sustancia gelatinosa
Núcleo propio
Columna de Clarke
(núcleo torácico)
Gris visceral
Intermediolateral
Intermediomedial
Retrodorsolateral
Dorsomedial
Dorsolateral
Núcleo frénico
Ventrolateral
Ventromedial
Accesorio
C 1
2
3
4
5
6
7
8
1
2
3
4
5
6
7
8
9
10
11
12
1
2
3
4
5
1
2
3
4
5
C 1
Funículo dorsal
Columna gris dorsal
T
Canal central
Funículo lateral
Columna lateral gris
L
Columna ventral gris
S
Funículo ventral
Sustancia gelatinosa
Núcleo propio
Columna de Clarke
(núcleo torácico)
Gris visceral
Intermediolateral
Intermediomedial
Parasimpático sacro
Retrodorsolateral
Dorsomedial
Dorsolateral
Lumbosacro
Ventrolateral
Ventromedial
Figura 18.3 Los grupos de células nerviosas en las columnas grises de la médula espinal. Se indican las posiciones relativas de estos grupos columnares y su extensión a
través de los segmentos espinales. (Modificado con permiso de Simon & Schuster de Correlative Anatomy of the Nervous System por E. Crosby,
T. Humphrey, E. Lauer. Copyright © 1962 Macmillan Publishing Company.)
Las aferencias de la raíz dorsal llevan información exteroceptiva, propioceptiva
e interoceptiva. Las láminas I-IV son las principales áreas receptivas cutáneas;
la lámina V recibe aferencias finas de la piel, los músculos y las vísceras; La
lámina VI recibe aferencias propioceptivas y algunas cutáneas. La mayoría, si
no todas, las fibras aferentes primarias se dividen en ramas ascendentes y
descendentes al entrar en el cordón. Estos luego viajan a distancias variables
en el tracto de Lissauer, cerca de la superficie del cordón, y envían colaterales.
en la materia gris subyacente. La formación, topografía y división de las raíces
espinales dorsales se han confirmado en el hombre.
La lámina marginal es una lámina delgada de neuronas en la punta dorsolateral
del asta dorsal, profunda al tracto de Lissauer. Debajo se encuentra la sustancia
gelatinosa (láminas II y III), que está presente en todos los niveles y está formada
principalmente por neuronas pequeñas, junto con algunas neuronas más grandes.
La sustancia gelatinosa recibe aferentes a través de la dorsal.
259
CAPÍTULO 18

MÉDULA ESPINAL: ORGANIZACIÓN INTERNA
raíces, y sus neuronas dan lugar a fibras que forman el tracto espinotalámico
contralateral. Las grandes neuronas proprioespinales del núcleo propio se
encuentran ventrales a la sustancia gelatinosa; enlazan segmentos para la
mediación de la coordinación intraespinal (fig. 18.3).
La columna de Clarke se encuentra en la base del asta dorsal. En la mayoría de
los niveles, está cerca del funículo blanco dorsal y puede proyectarse en él. En la
médula espinal humana, por lo general se puede identificar desde el octavo
segmento cervical hasta el tercero o cuarto lumbar. Las neuronas de la columna de
Clarke varían en tamaño, pero la mayoría son grandes, especialmente en los
segmentos lumbares y torácicos inferiores. Algunos envían axones a los tractos
espinocerebelosos dorsales y otros son interneuronas.
C8
Músculos de la mano
Músculos intrínsecos de las extremidades
Músculos de las extremidades
con origen en el maletero
Cuerno lateral
Músculos axiales
El cuerno lateral es una pequeña proyección lateral de materia gris ubicada
entre los cuernos dorsal y ventral. Está presente desde el octavo segmento
cervical o primer segmento torácico hasta el segundo o tercer segmento
lumbar. Contiene los cuerpos celulares de las neuronas simpáticas
preganglionares. Estos se desarrollan en el cordón embrionario dorsolateral al
canal central y migran lateralmente, formando columnas de células
intermediomediales e intermediolaterales. Sus axones viajan a través de raíces
espinales ventrales y ramas comunicantes blancas hasta el tronco simpático.
Un grupo de células similar se encuentra en el segundo al cuarto segmento
sacro, pero a diferencia de la columna de células lateral toracolumbar, no
forma una proyección lateral visible. Es la fuente del flujo sacro de las fibras
nerviosas preganglionares parasimpáticas.
Figura 18.4 La ubicación aproximada de los grupos de células motoras en el nivel segmentario C8
de la médula espinal.
L1 L2 L3 L4 L5 S1 S2 S3
Iliopsoas
Sartorio
Pectíneo
Gracilis
Aductor largo
Aductor corto
Aductor mayor
Cuádriceps femoral
Obt. ext.
Tib. hormiga.
Cuerno ventral
Las neuronas del cuerno ventral varían en tamaño. Los cuerpos celulares más
grandes, que pueden superar los 25 μ m de diámetro, son las de α neuronas
motoras, cuyos axones emergen en las raíces ventrales para inervar las fibras
extrafusales en los músculos esqueléticos estriados. Gran cantidad de neuronas más
pequeñas, 15-25 μ m de diámetro, también están presentes. Algunos de estos son γ
las neuronas motoras, que inervan las fibras intrafusales de los husos musculares, y
el resto son interneuronas. Las neuronas motoras utilizan acetilcolina como su
neurotransmisor.
Consideradas longitudinalmente, las neuronas del asta ventral están
dispuestas en grupos alargados y forman varias columnas separadas, que se
extienden a través de varios segmentos. Estos se ven más fácilmente en
secciones transversales. El asta ventral se puede dividir en columnas de células
medial, central y lateral, todas las cuales exhiben subdivisión en ciertos niveles,
generalmente en partes dorsal y ventral (Fig. 18.3). El grupo medial se extiende
por todo el cordón, pero puede estar ausente en el quinto segmento lumbar y
el primer sacro. En el torácico y los cuatro segmentos lumbares superiores, se
subdivide en grupos ventromedial y dorsomedial. En los segmentos craneal y
caudal a esta región, el grupo medial tiene sólo una fracción ventromedial,
excepto en el primer segmento cervical, donde sólo existe el grupo
dorsomedial.
El grupo central de células es el menos extenso y se encuentra solo en
algunos segmentos cervicales y lumbosacros. El núcleo frénico situado en el
centro, que contiene las neuronas motoras que inervan el diafragma, se
encuentra en los segmentos cervicales tercero a séptimo. Un grupo accesorio
irregular de neuronas en los cinco o seis segmentos cervicales superiores en el
borde ventral del cuerno ventral da lugar a axones que se cree que entran en
el nervio espinal accesorio (fig. 18.3).
El grupo lateral de células en el asta ventral se subdivide en grupos
ventral, dorsal y retrodorsal, en gran parte confinados a los segmentos
espinales que inervan las extremidades. El núcleo de Onuf, que se cree
que inerva los músculos estriados perineales, es un grupo ventrolateral
de células en el primer y segundo segmento sacro.
Las neuronas motoras del asta ventral están organizadas somatotópicamente. La
disposición básica es que los grupos de células mediales inervan la musculatura axial
y los grupos de células laterales inervan las extremidades. El bloque de construcción
básico de las poblaciones de neuronas motoras somáticas está representado por un
grupo de neuronas dispuestas longitudinalmente, que inervan un músculo
determinado, y en el que el α y γ las neuronas motoras están entremezcladas. Los
diversos grupos que inervan diferentes músculos se agregan en dos columnas
longitudinales principales, medial y lateral. En la sección transversal, estos forman
los grupos de células medial y lateral en el asta ventral (Fig. 18.4).
La columna motora longitudinal medial se extiende a lo largo de la médula
espinal. Sus neuronas inervan grupos de músculos epaxiales e hipaxiales.
Básicamente, los músculos epaxiales incluyen el grupo erector de la columna (que
extiende la cabeza y la columna vertebral), mientras que los músculos hipaxiales
incluyen los músculos prevertebrales del cuello, los músculos intercostales y de la
pared abdominal anterior (que flexionan el cuello y el tronco). los
Tib. correo.
Diez. fas. lata
Exceso. medicina y
min. Semimembranoso
Semitendinoso
Ext. hall.l.
Ext. cavar. l.
Mentira. tert.
Mentira. brevis
Mentira. longus
Lat. cadera. putrefacción
Gastrocn.
Soleus y planta.
Bíceps femoral
Glúteo máx.
Flexionar. pasillo l. y B.
Flexionar. cavar. l. y B.
Intrínsecos del pie
Figura 18.5 Disposición segmentaria de las neuronas motoras que inervan los
músculos del miembro inferior.
Los músculos epaxiales están inervados por ramas de las ramas primarias dorsales
de los nervios espinales y los músculos hipaxiales por ramas de las ramas primarias
ventrales. En la columna medial, las neuronas motoras que inervan los músculos
epaxiales están ubicadas ventrales a las que inervan los músculos hipaxiales.
La columna motora longitudinal lateral se encuentra solo en las
ampliaciones de la médula espinal. Las neuronas motoras de esta
columna en las ampliaciones cervical y lumbar inervan los músculos de
las extremidades superiores e inferiores, respectivamente. En el
agrandamiento cervical, las neuronas motoras que inervan los músculos
intrínsecos del miembro superior están situadas dorsalmente en la
columna gris ventral, y las que inervan los músculos más distales (de la
mano) están situadas más dorsalmente. Las neuronas motoras de los
músculos de la cintura se encuentran en la parte ventrolateral del asta
ventral. Existe una organización somatotópica adicional en el sentido de
que los músculos proximales de la extremidad son inervados por grupos
de células motoras ubicadas más rostralmente en el agrandamiento que
los que inervan los músculos distales. Por ejemplo, las neuronas motoras
que inervan los músculos intrínsecos de la mano están ubicadas en los
segmentos C8 y T1,
Las principales conexiones aferentes a las neuronas motoras son:
conexiones monosinápticas directas de aferentes de la raíz dorsal
propioceptiva en el mismo segmento o en segmentos cercanos; conexiones de
colaterales axonales del asta dorsal y otras interneuronas; conexiones
monosinápticas directas de los tractos vestibuloespinal y corticoespinal.
260
SECCIÓN 3
Organización interna
Reflejos espinales
Reflejo flexor
La estimulación dolorosa de las extremidades conduce a la retirada de la fl exión,
que está mediada por un reflejo polisináptico (fig. 18.8) en el que las interneuronas
unen las neuronas aferentes y eferentes. Por tanto, la activación de aferentes
primarios nociceptivos provoca indirectamente la activación de las neuronas motoras
fl exoras de las extremidades. La colateralización de las fibras a los segmentos
espinales cercanos media la fl exión de una extremidad en varias articulaciones,
según la intensidad del estímulo. Se activan las conexiones decusado al lado
contralateral del cordón. α neuronas motoras que inervan los músculos extensores
correspondientes, lo que produce el llamado reflejo extensor cruzado. En principio,
prácticamente cualquier estímulo cutáneo tiene el potencial de inducir un reflejo fl
exor, pero, salvo en el caso de los estímulos nocivos, esta respuesta normalmente se
inhibe por vías descendentes. Cuando se pierden las influencias descendentes,
incluso la estimulación cutánea inofensiva puede provocar la flexión de las
extremidades. El re fl ejo de Babinski (extensor plantar), que generalmente se
considera patognomónico del daño del tracto corticoespinal, es parte de un
Las conexiones intrínsecas de la médula espinal y el tronco encefálico sirven a una
serie de reflejos mediante los cuales las funciones de las estructuras periféricas se
modulan en respuesta a la información aferente de una manera relativamente
automática o autónoma. Los componentes fundamentales de tales «arcos» reflejos
son, por tanto, una neurona aferente y una eferente. Sin embargo, en todos los
reflejos, excepto en el más simple, las interneuronas intervienen entre los
componentes aferente y eferente, lo que confiere una mayor versatilidad y
complejidad a las respuestas del reflejo. Los reflejos, por su propia naturaleza, tienen
una forma relativamente fija y estereotipada. Sin embargo, están fuertemente
influenciados y modulados por conexiones descendentes. En el caso de los reflejos
espinales, estos controles descendentes provienen tanto del tronco encefálico como
de la corteza cerebral. La patología de las vías supraespinales descendentes suele
causar anomalías en la actividad del reflejo espinal, que se evalúan de forma
rutinaria en el examen neurológico. Durante el desarrollo, los mecanismos de control
descendente suprimen lo que puede considerarse como respuestas del reflejo
espinal "primitivo", como el reflejo plantar extensor y el reflejo de prensión. Cuando
los mecanismos de control superiores se dañan, estos reflejos se liberan y
reaparecen como un signo de patología del SNC (p. Ej., El reflejo de Babinski).
1a neurona aferente Extrafusal
fibras musculares
Estirar re fl exión
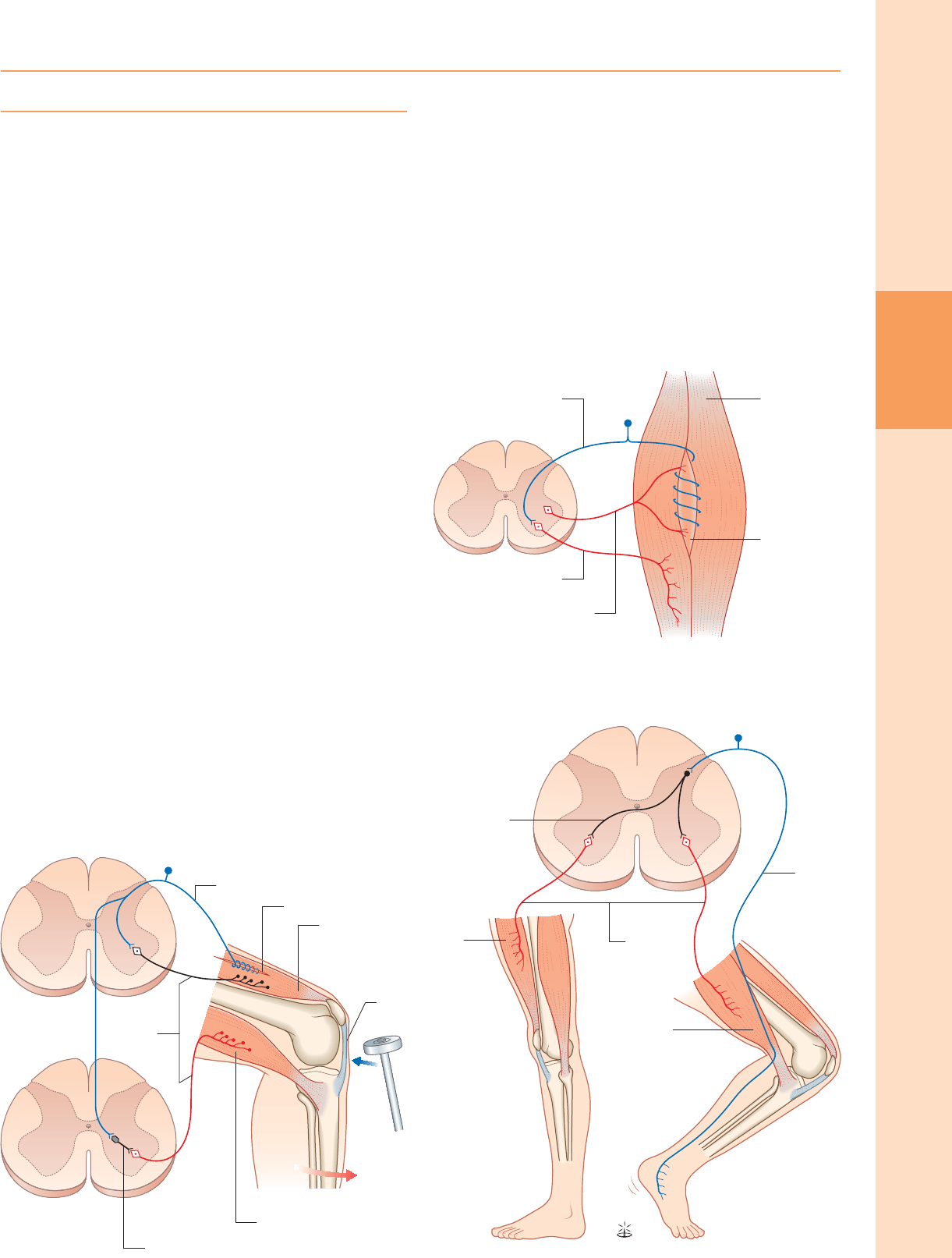
El reflejo del estiramiento es el mecanismo por el cual el estiramiento aplicado a un
músculo provoca su contracción del reflejo. Es fundamental para el mantenimiento
tanto del tono muscular como de la postura erguida (a través de la inervación de los
músculos posturales del cuello, la espalda y las extremidades inferiores).
Anatómicamente es el más simple de los reflejos, ya que está mediado únicamente
por una neurona aferente y una eferente. El componente aferente surge de los
receptores de estiramiento asociados con las fibras musculares intrafusales ubicadas
dentro de los husos musculares. Las terminaciones primarias o anuloespirales de
estas células receptivas dan lugar a fibras aferentes primarias que entran en la
médula espinal, donde hacen contacto sináptico excitador directamente sobre α
neuronas motoras que inervan el mismo músculo (fig. 18.6). los α Las neuronas
motoras de los músculos antagonistas se inhiben simultáneamente a través de
conexiones colaterales con las interneuronas inhibidoras.
Intrafusal
fibras musculares
Neurona motora alfa
Neurona motora gamma
Reflejo gamma
Figura 18.7 El reflejo gamma. (Con permiso de Crossman AR, Neary D 2000
Neuroanatomy, 2nd edn. Edimburgo: Churchill Livingstone.)
Al igual que α neuronas motoras que inervan las fibras musculares extrafusales, los
músculos también reciben γ neuronas motoras, que inervan las fibras musculares
intrafusales. Activación de γ las neuronas motoras aumentan la sensibilidad de las
fibras intrafusales al estiramiento (fig. 18.7). Por lo tanto, los cambios en γ
La actividad tiene un efecto profundo sobre el reflejo del estiramiento y sobre el tono
muscular. Igual que α neuronas motoras, γ las neuronas motoras están bajo la
influencia de vías descendentes desde el tronco encefálico y la corteza cerebral. Los
cambios en la actividad del reflejo de estiramiento y del tono muscular se encuentran
comúnmente en los trastornos del SNC y del SNC.
Cordón lumbar
Interneurona
A
Cutáneo
aferente
neurona
1a aferente
Fibra muscular intrafusal
Músculo cuádriceps
Cuadríceps
músculo
Motor alfa
neuronas
Rotuliano
tendón
Flexor de rodilla
músculos
Motor alfa
neurona
B
Músculos flexores de la rodilla
Interneurona inhibidora
Figura 18.8 El fl exor re fl exor y el re fl ejo extensor cruzado. (Con permiso de
Crossman AR, Neary D 2000 Neuroanatomy, 2nd edn. Edimburgo: Churchill
Livingstone.)
Figura 18.6 El re fl ejo de estiramiento. (Con permiso de Crossman AR, Neary D
2000 Neuroanatomy, 2nd edn. Edimburgo: Churchill Livingstone.)
261
CAPÍTULO 18
MÉDULA ESPINAL: ORGANIZACIÓN INTERNA
fl exión retracción del miembro inferior en respuesta a la estimulación de la
planta del pie.
contienen sólo pequeñas fibras, por ejemplo, el tracto dorsolateral, el fascículo
gracilis y la parte central del funículo lateral. El fascículo cuneatus, el funículo
anterior y la zona periférica del funículo lateral contienen muchas fibras de
gran diámetro.
Si bien los tramos ascendentes y descendentes son en gran medida discretos y
están ubicados regularmente, se produce una superposición significativa entre
tramos adyacentes. Su disposición general se muestra en la figura 18.9.
MATERIA BLANCA ESPINAL
La sustancia blanca espinal rodea el núcleo central de la materia gris. La sustancia
blanca se compone principalmente de fibras nerviosas que discurren
longitudinalmente. Las fibras que sirven a funciones relacionadas, o aquellas con
orígenes o destinos comunes, generalmente se agrupan anatómicamente para
formar tractos (fascículos), que pueden ser ascendentes, descendentes y
propioespinales. Los tractos ascendentes consisten en fibras aferentes primarias,
que ingresan a la médula a través de las raíces dorsales de los nervios espinales, o
fibras derivadas de neuronas espinales intrínsecas, que transportan impulsos
aferentes a niveles supraespinales. Los tractos descendentes contienen fibras que
descienden de la corteza cerebral o de los núcleos del tronco encefálico para
controlar la actividad de las neuronas espinales. Los tractos propioespinales
contienen los axones de neuronas que se localizan completamente en la médula
espinal: contienen componentes ascendentes y descendentes,
La materia blanca espinal se describe convencionalmente como dispuesta en tres
grandes masas emparejadas bilateralmente, los funículos dorsal, lateral y ventral,
cada uno de los cuales contiene varios tractos predominantemente específicos
(véanse la figura 18.1 y la figura 18.9). Las comisuras blancas dorsal y ventral
estrechas corren entre las dos mitades del cordón.
Las fibras de la sustancia blanca espinal varían en calibre. Muchos son pequeños y
ligeramente mielinizados o no. La mayoría de las regiones contienen un amplio espectro
10 μ metro. Algunos tratados típicamente
Caminos ascendentes
Columnas dorsales
El funículo dorsal consta de dos grandes extensiones ascendentes, el fasciculus
gracilis y el fasciculus cuneatus (fig. 18.10), que también se conocen como
columnas dorsales. Están separados por un tabique posterointermedio. Las
columnas dorsales contienen una alta proporción de fibras mielinizadas que
llevan la propioceptiva (sentido de posición y cinestesia), exteroceptiva (tacto-
presión) y sensación vibratoria a niveles más altos. Estas fibras proceden de
varias fuentes: fibras aferentes primarias largas que entran en la médula por
las raíces dorsales de los nervios espinales y ascienden a los núcleos de la
columna dorsal en el bulbo raquídeo; fibras aferentes primarias más cortas
que se proyectan a las neuronas de la columna de Clarke y otras neuronas
espinales; axones de neuronas secundarias de la médula espinal que
ascienden a los núcleos de la columna dorsal. Las columnas dorsales también
contienen axones de neuronas propioespinales.
El fasciculus gracilis comienza en el extremo caudal de la médula
espinal. Contiene largas ramas ascendentes de aferentes primarios, que
de diámetros de fi bra, desde 1 μ mo menos a
A
Vías descendentes
Fascículo interfascicularis
Fascículo gracilis
De la corteza cerebral
Del tronco encefálico
Fascículo dorsolateral
(de Lissauer)
Caminos ascendentes
Fasciculus cuneatus tracto
espinocerebeloso dorsal
Fasciculus proprius
Columnas dorsales
Tractos espinocerebelosos
Espinotalámico y
Tractos espinorreticulares
Tracto corticoespinal lateral
Tracto rubroespinal
Tracto reticuloespinal lateral
Fascículo longitudinal medial
Tracto reticuloespinal medial
Tracto tectoespinal
Tracto reticuloespinal ventrolateral
Tracto espinocerebeloso ventral
Tracto espino-olivar Tracto espino-
olivar
Tracto vestibuloespinal
lateral (ventrolateral)
Tracto reticuloespinal ventral Tracto corticoespinal ventral
B Fascículo septomarginalis Fascículo gracilis
Fascículo dorsolateral (de Lissauer)
Fasciculus proprius
Tracto corticoespinal lateral
Tracto tegmentoespinal
Tracto reticuloespinal lateral
Tracto espinocerebeloso ventral
Tracto espinotalámico y espinotectal lateral
Tracto reticuloespinal medial
Tracto vestibuloespinal ventrolateral
Tracto reticuloespinal ventral
Tracto espino-olivar
Tracto espinotalámico ventral
Tracto corticoespinal ventral Fascículo sulcomarginal
262
Figura 18.9 Las posiciones aproximadas de los tractos de fibras nerviosas en la médula espinal en la mitad de las cervicales ( A) y lumbar B) niveles.
SECCIÓN 3
Organización interna
Corteza cerebral (circunvolución poscentral)
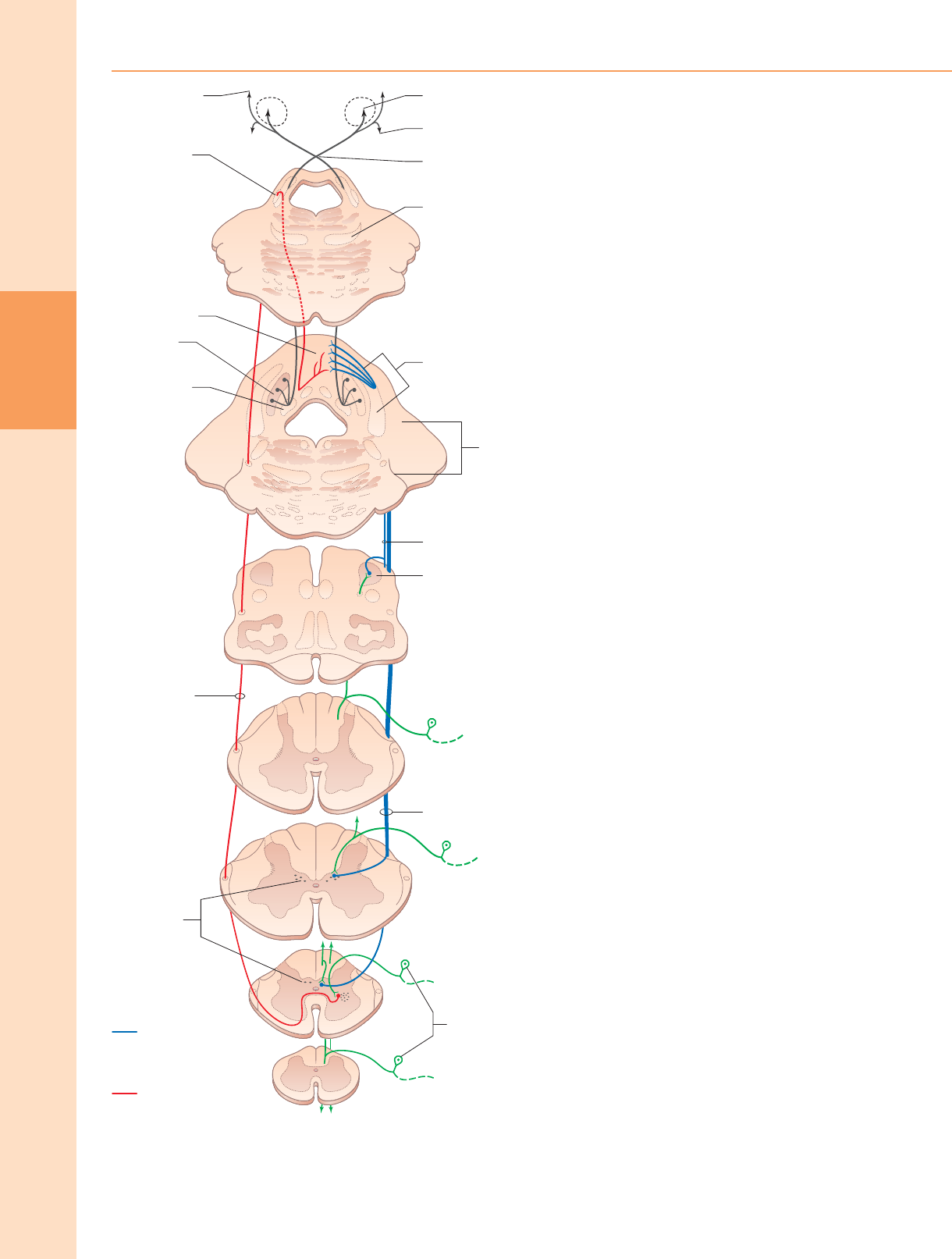
El fascículo gracilis se encuentra medial al fascículo cuneatus en la médula
espinal superior (fig. 18.9). En los niveles cervicales superiores, el fasciculus gracilis
contiene una mayor proporción de aferentes de los receptores cutáneos que de los
propioceptores profundos porque muchos de estos últimos dejan el fascículo en los
segmentos inferiores para hacer sinapsis en la columna de Clarke. De hecho, la
propiocepción de la extremidad inferior llega en su mayor parte al tálamo por
retransmisión en la columna de Clarke y luego nuevamente en el núcleo Z (pág. 280).
Los axones del fasciculus gracilis, tanto de las neuronas primarias como secundarias,
terminan en el núcleo gracilis de la médula dorsal.
El fasciculus cuneatus (fig. 18.9) comienza a nivel torácico medio y se
encuentra lateral al fasciculus gracilis. Se compone principalmente de fibras
aferentes primarias de las raíces dorsales torácicas superiores y cervicales. En
los niveles cervicales superiores contiene una gran población de aferentes de
receptores tanto profundos como cutáneos del miembro superior. Además,
algunos de sus axones surgen de neuronas secundarias en las láminas IV-VI
del asta dorsal ipsilateral. Muchos axones (tanto primarios como secundarios)
que ascienden en el fascículo cuneatus terminan en el núcleo cuneatus de la
médula dorsal. Algunas también terminan en el núcleo cuneiforme lateral
(externo o accesorio); las neuronas de este núcleo se proyectan hacia el
cerebelo a través de la vía cuneocerebelosa.
Muchas fibras ascendentes del fasciculus gracilis y fasciculus cuneatus
terminan en sinapsis con neuronas de los núcleos de la columna dorsal
(núcleo gracilis y núcleo cuneatus, respectivamente) en el bulbo raquídeo.
(Las conexiones de los núcleos de la columna dorsal se describen con
más detalle con el bulbo raquídeo, pág. 280.) Los axones que surgen de
las neuronas en el arco del núcleo de la columna dorsal
ventromedialmente alrededor de la sustancia gris central de la médula
como fibras arqueadas internas (Fig. 19.5) y decusarse en la gran
decusación sensorial (decusación del lemnisco medial) para formar el
lemnisco medial mismo. Ascienden al núcleo posterolateral ventral del
tálamo, desde donde se proyectan las neuronas hacia la corteza
somatosensorial en la circunvolución poscentral del lóbulo parietal (áreas
3, 1 y 2).
El alto grado de organización somatotópica que está presente en las columnas
dorsales se conserva a medida que las vías ascienden a través de los núcleos de la
columna dorsal y el tálamo para llegar a la corteza somatosensorial primaria. En los
núcleos de la columna dorsal, el miembro inferior está representado en el núcleo
gracilis, el miembro superior en el núcleo cuneatus y el tronco está representado en
una posición intermedia entre ellos. Las fibras también se segregan por modalidad
en las columnas dorsales: las fibras de los receptores capilares son más superficiales,
mientras que las de los receptores táctiles y vibratorios se encuentran en capas más
profundas.
Tálamo
Capsula interna
Área de la pierna
Área del maletero
Área del brazo
Área de la cabeza
Área de la cara
Ventral
posterolateral
núcleo de
tálamo
Ganglios basales
Mesencéfalo
Lemnisco medial
Puente de Varolio
Nervio trigémino
Núcleo gracilis
Núcleo cuneatus
Lemnisco medial
Tractos espinocerebelosos
Hay dos tractos espinocerebelosos principales: dorsal (posterior) y ventral
(anterior). Ocupan la periferia de la cara lateral de la sustancia blanca
espinal (fig. 18.9, fig. 18.11) y transportan información propioceptiva y
cutánea al cerebelo para la coordinación del movimiento. Ambos tractos
contienen fibras mielinizadas de gran diámetro, pero hay más en el tracto
dorsal. Las fibras de calibre más fino están asociadas con el tracto ventral.
El haz espinocerebeloso dorsal se encuentra lateral al haz corticoespinal lateral.
Comienza aproximadamente al nivel del segundo o tercer segmento lumbar y se
agranda a medida que asciende. Los axones del haz se originan ipsilateralmente a
partir de las neuronas más grandes de la columna de Clarke, en la lámina VII a lo
largo de los segmentos espinales T1-L2. La columna de Clarke recibe información de
colaterales de aferentes primarios ascendentes largos de las columnas dorsales y
terminales de aferentes primarios ascendentes más cortos de las columnas dorsales.
Muchas de estas fibras aferentes ascienden desde los segmentos caudales hasta L2.
En la médula, el haz espinocerebeloso dorsal pasa a través del pedúnculo cerebeloso
inferior para terminar ipsilateralmente en las partes rostral y caudal del vermis
cerebeloso.
El haz espinocerebeloso ventral se encuentra inmediatamente ventral al haz
dorsal. Las células de origen se encuentran en las láminas V-VII del cordón
lumbosacro y el tracto transporta información de la extremidad inferior. La mayoría
de los axones que forman el haz se decusan, pero algunos permanecen ipsolaterales.
El trayecto comienza en la región lumbar superior y asciende a través del bulbo
raquídeo para alcanzar el nivel pontino superior, desde donde desciende en la parte
dorsal del pedúnculo cerebeloso superior para terminar, principalmente
contralateralmente, en el vermis cerebeloso anterior.
Los tractos espinocerebelosos están organizados de manera que las fibras de los
segmentos espinales inferiores son más superficiales. Ambos tractos transmiten
información propioceptiva y exteroceptiva, pero son funcionalmente diferentes. Las
neuronas de la columna de Clarke son excitadas monosinápticamente por las fibras
aferentes primarias Ia y Ib (de los husos musculares y los órganos tendinosos,
respectivamente) y también por las aferencias musculares del grupo II y las aferentes
cutáneas del tacto y de la presión. Los impulsos propioceptivos a menudo surgen de
un solo músculo o de músculos sinérgicos que actúan en una articulación común.
Núcleo espinal del trigémino
Médula
Fibras arqueadas internas
Discusión de
lemnisco medial
Espina dorsal
neurona ganglionar
Fascículo gracilis
C7
Fasciculus cuneatus
T4
Fascículo gracilis
Sacro
Lumbar
Torácico
Cervical
L4
Figura 18.10 Las columnas dorsales. Las fibras aferentes primarias de diferentes
niveles y sus neuronas de segundo y tercer orden asociadas se representan en
diferentes colores.
entran en la médula a través de las raíces espinales dorsales ipsolaterales y los
axones ascendentes de las neuronas secundarias en las láminas IV a VI del asta
dorsal ipsolateral. A medida que ascienden las fibras, se les unen axones de raíces
dorsales sucesivas. Las fibras que entran en las regiones coccígea y sacra inferior se
desplazan medialmente por adiciones sucesivas de fibras que entran en niveles
superiores.
263
CAPÍTULO 18
MÉDULA ESPINAL: ORGANIZACIÓN INTERNA
Fibras dentatotalámicas Fibras cerebelorubrales
Mits información para el movimiento coordinado y la postura de toda la
extremidad inferior.
Dado que la columna de Clarke disminuye rostralmente (fig. 18.3) y no
se extiende por encima del segmento cervical más bajo, se deduce que el
haz espinocerebeloso dorsal transporta información desde el tronco y la
extremidad inferior. La información propioceptiva y exteroceptiva del
miembro superior viaja en las fibras aferentes primarias del fascículo
cuneatus. Estas fibras terminan somatotópicamente en el núcleo
cuneiforme accesorio (externo o lateral) y en la parte adyacente del
núcleo cuneiforme situado en el bulbo raquídeo. Las células de estos
núcleos dan lugar a las fibras arqueadas externas posteriores que forman
el haz cuneocerebeloso (fig. 18.11), que entra en el cerebelo a través del
pedúnculo cerebeloso inferior ipsolateral. El núcleo cuneiforme accesorio
y la parte lateral del núcleo cuneiforme se consideran homólogos a las
células de la columna de Clarke.
Los axones de todos los tractos espinocerebelosos y cuneocerebelosos
forman parte del "sistema de fibras musgosas". Terminan en la corteza
cerebelosa en un patrón funcional, somatotópico y altamente organizado (pág.
302).
Tracto espinocerebeloso
anterior en la superficie del
pedúnculo cerebeloso superior
Fibras dentatoreticulares
Decusación del pedúnculo
cerebeloso superior
Lemnisco medial
Puente superior
Vermis del cerebelo
Núcleo dentado
Posterior
fibras espinocerebelosas
en cerebelo inferior
pedúnculo
Superior
pedúnculo cerebeloso
Tractos espinotalámicos
Los tractos espinotalámicos (fig. 18.9) constan de neuronas de segundo orden que
transmiten información sobre el dolor, la temperatura, el tacto grosero (no
discriminativo) y la presión a la región somatosensorial del tálamo. Las células de
origen se encuentran en varias láminas de todos los segmentos de la médula espinal.
Las fibras se decusan en la comisura blanca ventral para alcanzar los tractos
espinotalámicos contralaterales: las fibras de dolor y temperatura lo hacen
rápidamente, dentro de aproximadamente un segmento de su origen, mientras que
las fibras que llevan otras modalidades pueden ascender varios segmentos antes de
cruzar. Las fibras espinotalámicas ascienden principalmente en la sustancia blanca
ventrolateral al asta ventral, en parte entremezcladas con fibras espinorreticulares
ascendentes y fibras reticuloespinales descendentes. Algunas autoridades describen
dos tractos espinotalámicos (lateral y ventral) con ubicaciones y funciones
anatómicas más o menos distintas. Sin embargo, los estudios fisiológicos en
animales apoyan la idea de que estos tractos pueden considerarse mejor como un
continuo estructural y funcional.
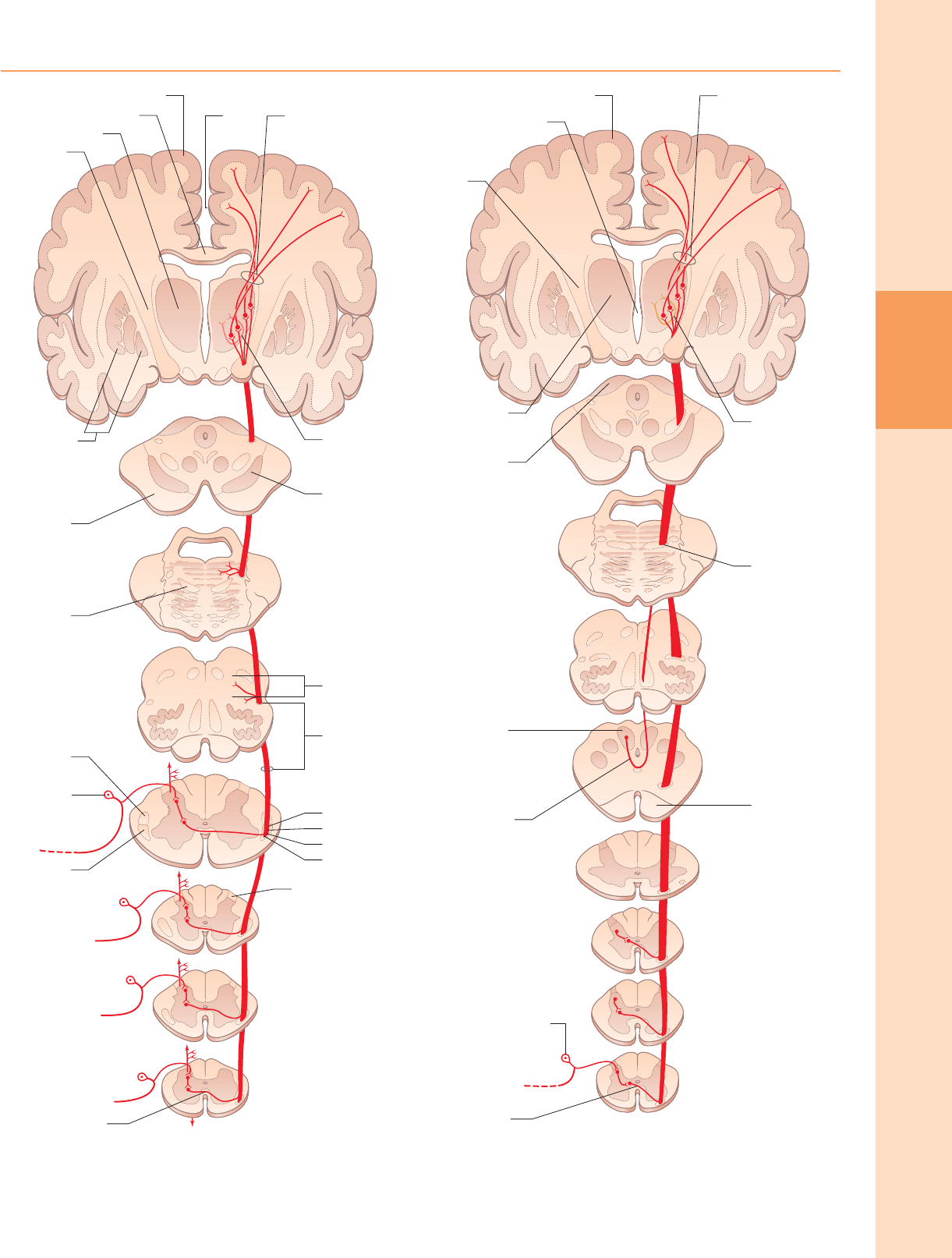
El haz espinotalámico lateral (fig. 18.12) está ubicado en el funículo
lateral, medial al haz espinocerebeloso ventral. La evidencia clínica indica
que favorece las sensaciones de dolor y temperatura. El haz
espinotalámico ventral (fig. 18.13) se encuentra en el funículo anterior
medial al punto de salida de las raíces nerviosas espinales ventrales y
dorsal al haz vestibuloespinal, que se superpone. Sobre la base de la
evidencia clínica, favorece las modalidades táctiles y de presión toscas.
En animales se ha descrito un tracto espinotalámico dorsolateral. Los
axones surgen principalmente de neuronas en la lámina I y cruzan para
ascender en el funículo dorsolateral contralateral. Estas neuronas responden al
máximo a estímulos cutáneos nocivos, mecánicos y térmicos. Los ejemplos de
alivio clínico del dolor después de la cordotomía dorsolateral sugieren que
existe una proyección similar en el hombre.
Al llegar al tronco encefálico inferior, los axones del tracto espinotalámico se
separan. Los axones del tracto ventral se unen al lemnisco medial. Los axones en el
tracto lateral continúan como el lemnisco espinal.
Existe una clara organización somatotópica de las fibras en los tractos
espinotalámicos en toda su extensión. Las fibras que se cruzan a cualquier nivel del
cordón se unen a la cara profunda de las que ya se han cruzado, lo que significa que
ambas extensiones están laminadas segmentariamente (Fig. 18.14). La somatotopía
se mantiene en todo el bulbo raquídeo y la protuberancia. En el mesencéfalo, las
fibras del lemnisco espinal que transmiten la sensación de dolor y temperatura del
miembro inferior se extienden dorsalmente, mientras que las del tronco y el
miembro superior se colocan más ventralmente. Ambos lemnisci ascienden para
terminar en el tálamo. Las principales proyecciones espinotalámicas en el hombre
son hacia el núcleo posterolateral ventral y también hacia el núcleo intralaminar
centrolateral.
Medio
cerebeloso
pedúnculo
Puente de Varolio
Tracto cuneocerebeloso
Accesorio
núcleo cuneado
Médula
Anterior
tracto espinocerebeloso
C4
Posterior
tracto espinocerebeloso
C8
Núcleo dorsal
(Columna de Clarke)
Órgano del tendón de Golgi
(Cuádriceps femoral)
L3
Espina dorsal
ganglio
neuronas
Células de la parte posterior (dorsal)
tracto espinocerebeloso
y cuneocerebeloso
tracto
S2
Órgano del tendón de Golgi
(Músculos isquiotibiales)
Células del tracto espinocerebeloso
anterior (ventral)
Neuronas de los tractos espinotalámicos
La localización específica de los cuerpos celulares del tracto espinotalámico está poco
documentada en el hombre. En los animales, aproximadamente un tercio se localiza en los
tres segmentos cervicales superiores. Aproximadamente el 20% se localiza en los
segmentos cervicales inferiores, el 20% en la región torácica (principalmente en los
segmentos T1–3), el 20% en la región lumbar y el 10% en el cordón sacrococcígeo. Las
células se encuentran en las láminas I y IV-VIII, la mayor concentración se encuentra en las
láminas VI y VII. Los cuerpos celulares que dan lugar a los axones del tracto espinotalámico
son predominantemente contralaterales, aunque un número relativamente pequeño (10%),
principalmente en los tres segmentos cervicales superiores, son ipsolaterales.
Las neuronas de los tractos espinotalámicos tienen campos receptivos muy diferentes.
La especificidad de canales separados, tal como existe en los núcleos de la columna dorsal,
está ausente en las láminas del cordón. Convergencia de diferentes
Figura 18.11 Los tractos espinocerebelosos. (Redibujado de Carpenter MB 1991
Core Text of Neuroanatomy, 4th edn. Baltimore: Williams and Wilkins, con permiso
del autor y editor).
Por tanto, el tracto espinocerebeloso dorsal transmite información específica de la
modalidad y del espacio que se utiliza en la coordinación fina de los músculos
individuales de las extremidades. Por otro lado, las células del tracto ventral son
activadas monosinápticamente por aferentes Ib y transmiten información desde
grandes campos receptivos que incluyen diferentes segmentos de una extremidad.
El tracto ventral carece de subdivisiones para diferentes modalidades y trans-
264
SECCIÓN 3
Organización interna
Corteza cerebral (circunvolución poscentral)
Corteza cerebral (circunvolución poscentral)
Axones de neuronas en la extremidad
posterior de la cápsula interna
Cuerpo calloso
Tálamo
Capsula interna
Pie
Pierna
Axones de neuronas en la extremidad
posterior de la cápsula interna
III ventrículo
Pierna
Interno
cápsula
Tálamo
Mesencéfalo
Posterolateral ventral
núcleo (VPL)
Ganglios basales
Mesencéfalo
Posterolateral ventral
núcleo del tálamo
Colículo superior
Sustancia negra
Crus cerebri
Puente de Varolio
Espinotalámico anterior
tracto y medial
lemniscus
Puente de Varolio
Lemnisco medial
Médula
Formación reticular
Médula
Lateral
tracto espinotalámico
Núcleo gracilis
Temperatura
Médula
Espina dorsal
neurona ganglionar
Axón de la neurona
decidirse a ascender
en lemnisco medial
Pirámide
Fibras sacras
Fibras lumbares
Fibras torácicas
Fibras cervicales
C8
Dolor
C8
Fascículo dorsolateral
(de Lissauer)
T4
T12
L2
Neurona del ganglio de la raíz dorsal
Axón de la neurona
cruzando en anterior
comisura blanca
ascender en el tracto
espinotalámico anterior
S2
L3
Cruce de axones
al lado opuesto en la
comisura blanca anterior
Figura 18.12 El tracto espinotalámico lateral. (Redibujado de Carpenter MB 1991
Core Text of Neuroanatomy, 4th edn. Baltimore: Williams and Wilkins, con permiso
del autor y editor).
Figura 18.13 El tracto espinotalámico ventral (anterior). (Redibujado de
Carpenter MB 1991 Core Text of Neuroanatomy, 4th edn. Baltimore: Williams
and Wilkins, con permiso del autor y editor).
los tipos funcionales de fibras aferentes en una célula del tracto individual es una
característica común en el cordón. Sobre la base del sitio laminar, las propiedades
funcionales y la terminación talámica específica de sus axones, las neuronas del
tracto espinotalámico pueden dividirse en tres grupos separados. Estas son las
células apicales de la columna gris dorsal (lámina I), las células de la columna dorsal
profunda (láminas IV-VI) y las células de la columna gris ventral.
(láminas VII, VIII). Existen diferencias de especies y la descripción a
continuación se deriva de estudios en primates no humanos.
Las células de la lámina I que se proyectan al tálamo muestran las siguientes
características. En esencia, responden al máximo a la estimulación cutánea térmica o
nociva, y consisten principalmente en unidades de umbral alto, pero también de
rango dinámico amplio. Sus campos receptivos suelen ser
265
CAPÍTULO 18
D
D
norte
norte
a
a
H
H
metro
r
metro
A
r
A
MÉDULA ESPINAL: ORGANIZACIÓN INTERNA
ic
cil
Más alto
centros
C4 C8 T2 T6 T7 T12 L3 S1S5
Presión
Vibración
Movimiento
Posición
Tocar
Tractos descendentes
Diametro largo
fibras aferentes
Sustantia
gelatinosa
neurona
Lámina IV
neurona
Respuesta refleja
C1
S5
C6 C5
Pequeño diámetro
fibras aferentes
S1
7
L1 T12 T2 T1
C8 C
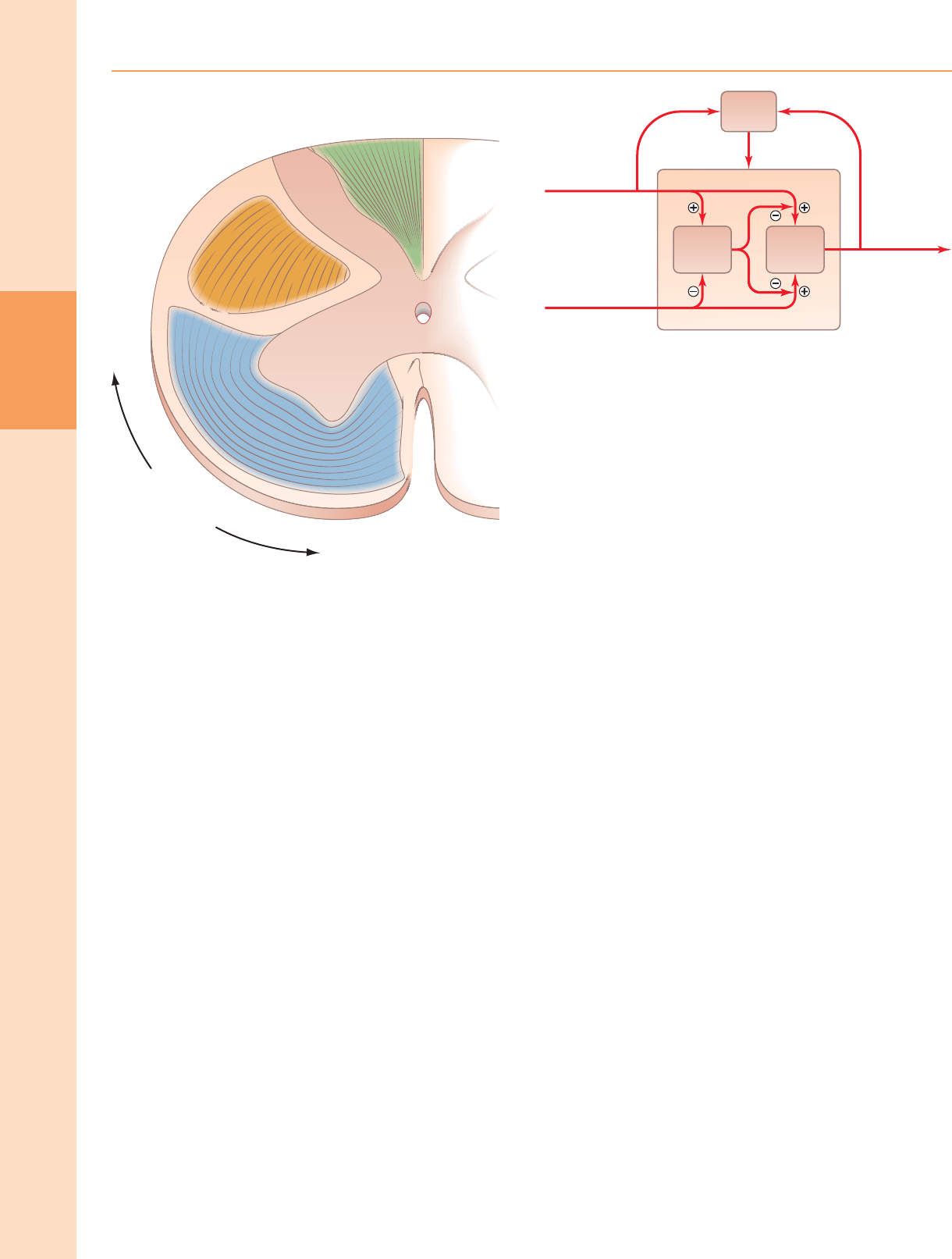
Figura 18.15 La disposición básica del mecanismo sensorial de "puerta" en las
láminas dorsales de la materia gris de la médula espinal. (Redibujado con permiso
de Melzack R, Wall PD. Mecanismos del dolor: una nueva teoría. Science 150: 971–
979. Copyright 1965 American Association for the Advancement of Science.)
C4
C8
T2
T6
T7
T12
L3
Mecanismos del dolor
S1
S5
Las conexiones ascendentes a través de las cuales la información sensorial
llega a los centros superiores no son simples relés, ya que se sabe que están
sujetas a modulación por conexiones intraespinales y por vías descendentes
desde el tronco encefálico y la corteza cerebral. Esto es particularmente
importante en relación con las vías espinotalámicas y espinorreticulares y la
percepción del dolor.
La inhibición presináptica influye en muchas, posiblemente todas, las
terminales aferentes primarias. Un sitio muy investigado de efectos
presinápticos es la sustancia gelatinosa. Se ha propuesto que los impulsos de
aferentes cutáneos (y otros) están aquí sujetos a control tónico por modulación
presináptica de terminales aferentes primarios, mediada por pequeñas
neuronas de la sustancia gelatinosa.
La "teoría del control de la puerta" (Melzack y Wall 1965) propuso un mecanismo
para la modulación del flujo de información a lo largo de las vías nociceptivas y otras
vías aferentes (fig. 18.15). La hipótesis era que las aferencias de gran diámetro (p. Ej.,
De los pelos y los corpúsculos del tacto) excitan las neuronas grandes de la lámina IV,
de las que surgen las fibras espinotalámicas, y las interneuronas de la sustancia
gelatinosa. Por el contrario, los aferentes finos no mielinizados excitan las células del
tracto pero inhiben las interneuronas. Se presume que los axones de las
interneuronas de la sustancia gelatinosa inhiben presinápticamente las terminales
de todas las aferencias que hacen sinapsis con las células del tracto. En tal sistema, la
baja actividad de las aferentes finas inhibe las interneuronas y, por lo tanto, evita que
inhiban las células del tracto. Por lo tanto, la "puerta" a las células en la lámina IV se
abre para transmitir pequeñas descargas intermitentes de impulsos de las fibras
grandes. Una descarga prolongada de impulsos de alta frecuencia en las aferentes
de gran diámetro se transmitiría inicialmente a las células del tracto de la lámina IV,
pero esto cesaría pronto cuando la actividad en las interneuronas cerrara la puerta.
Por el contrario, una alta actividad persistente en las aferentes finas abriría la puerta
dando como resultado un bombardeo masivo de neuronas de la lámina IV (que
incluyen algunas neuronas de umbral alto que sólo son activadas por tal
bombardeo). Se asumió que la transmisión hacia adelante en el tracto
espinotalámico lateral evocaría la percepción de dolor a niveles supraespinales.
Dolor, por tanto, resultaría de un desequilibrio entre las variedades de impulsos
aferentes cuando hubiera un tráfico desproporcionadamente grande a lo largo de
las aferentes finas. De hecho, la inhibición de las neuronas del tracto espinotalámico
puede producirse mediante la estimulación eléctrica de los nervios periféricos,
siendo las más efectivas las descargas que activan A δ fibras.
La actividad de las neuronas del tracto espinotalámico también puede modularse
selectivamente por vías que descienden del cerebro a la médula espinal. Muchos
estudios han demostrado que la respuesta de las células del tracto espinotalámico a
los estímulos nocivos se inhibe por la estimulación de ciertas regiones del cerebro.
Obviamente, esto tiene un interés clínico considerable en el tratamiento del dolor
crónico e intratable.
En el tronco encefálico, las regiones que inducen tales efectos
corresponden a una serie de núcleos mesencéfalo y rombencefálico que, con
sus conexiones, constituyen un sistema analgésico endógeno. En el
mesencéfalo, estas regiones son la sustancia gris periacueductal, el núcleo del
rafe dorsal y parte del núcleo cuneiforme. Las neuronas en estos sitios
contienen serotonina (5-HT), γ- ácido aminobutírico (GABA), sustancia P, CCK,
neurotensina, encefalina y dinorfina. La sustancia gris periacueductal recibe
aferencias de la neocorteza frontal somatosensorial y cingulada, la amígdala,
numerosos núcleos reticulares locales y el hipotálamo. Los aferentes de este
último son haces separados, que transportan histamina, hormona liberadora
de hormona luteinizante (LHRH), vaso-
Figura 18.14 Plano general de la organización segmentaria de las fibras en el funículo
dorsal, el tracto corticoespinal lateral y los tractos espinotalámicos. Las áreas de
sección transversal probables de estos tractos se amplían esquemáticamente. Este
plan general se aplica a todos los tractos organizados por segmentos, ya sean
ascendentes, descendentes, ipsilaterales o contralaterales.
pequeño, que representa una parte de un dedo o un área pequeña de piel que involucra varios dedos. Las
neuronas del tracto espinotalámico de la lámina I reciben información de A δ y fibras C, y algunas responden a
impulsos convergentes de receptores somáticos y viscerales profundos. Las células del tracto espinotalámico
en el cordón torácico muestran una marcada convergencia viscerosomática. Las neuronas del tracto
espinotalámico de la lámina I se proyectan preferentemente hacia el núcleo posterolateral ventral del tálamo,
con proyecciones limitadas hacia los núcleos talámico centrolateral y mediodorsal. La población de neuronas
espinotalámicas de la columna dorsal profunda (láminas IV-VI) del cordón lumbar contiene unidades de tipo de
rango dinámico amplio (60%), umbral alto (30%) y umbral bajo (10%). Pueden codificar con precisión estímulos
cutáneos tanto inocuos como nocivos. Algunas células también responden a la información de receptores
somáticos y viscerales profundos. En el cordón lumbar sus campos receptivos son de tamaño pequeño o
mediano; son más grandes que el área del pie, pero más pequeños que toda la pierna. En el cordón torácico,
los campos de estas células laminares son más grandes ya menudo incluyen todo el miembro superior más
parte del tórax. Muchas de las neuronas del tracto espinotalámico de la columna gris dorsal profunda en los
segmentos torácicos reciben impulsos convergentes de las fibras aferentes simpáticas. Las unidades del tracto
espinotalámico de las láminas IV-VI se proyectan al núcleo posterolateral ventral o al núcleo centrolateral del
tálamo y, a veces, a ambos. Las unidades que se proyectan hacia el núcleo posterolateral ventral reciben
información de todas las clases (A Las unidades del tracto espinotalámico de las láminas IV-VI se proyectan al
núcleo posterolateral ventral o al núcleo centrolateral del tálamo y, a veces, a ambos. Las unidades que se
proyectan hacia el núcleo posterolateral ventral reciben información de todas las clases (A Las unidades del
tracto espinotalámico de las láminas IV-VI se proyectan al núcleo posterolateral ventral o al núcleo
centrolateral del tálamo y, a veces, a ambos. Las unidades que se proyectan hacia el núcleo posterolateral
ventral reciben información de todas las clases (A β, A δ y C) de fibras cutáneas.
Las células del tracto espinotalámico de la columna gris ventral (láminas VII y VIII)
responden principalmente a estímulos somáticos profundos (músculos y
articulaciones), pero también a estímulos cutáneos inocuos y / o nocivos. En las
regiones torácicas de la médula espinal también reciben información convergente de
fuentes viscerales. La mayoría de las neuronas del tracto espinotalámico de las
láminas VII y VIII tienen campos receptivos grandes y complejos (a menudo
bilaterales), que abarcan áreas extensas del cuerpo. Las células de este grupo, que se
proyectan exclusivamente al tálamo medial, reciben información de A β, A δ
y clases C de fibras aferentes, y muchas responden a entradas convergentes
de receptores de estructuras profundas. Esta población de neuronas contiene
unidades de rango dinámico amplio (25%), de umbral alto (63%) y de umbral
bajo (12%). La mayoría de las células del tracto espinotalámico de la columna
gris ventral se proyectan hacia los núcleos intralaminares del tálamo. Las
neuronas de amplio rango dinámico son particularmente efectivas para
discriminar entre diferentes intensidades de estimulación dolorosa.
266
C
real academia de bellas artes
PAG
como
gramo
r
F
mi
nosotros
s
cul
atu
s
ci
mi
s
s
norte
a
tu
tu
F
C
r
mi
t
C
a
r
t
gramo
l
a
mi
L
en
pag
k
s
norte
o
C
tu
I
r
rt
T
o
C
metro
l
a
r
r
A
mi
t
a
L
SECCIÓN 3
T
mi
metro
pag
mi
r
a
t
tu
r
mi
PAG
a
S
I
pag
norte
I
norte
o
t
h
a
l
a
metro
I
C
t
r
a
C
t
s
T
o
tu
C
h
s
ulu
es
Organización interna
presina, oxitocina, hormona adrenocorticotrófica (ACTH), hormona estimulante
de la melanocitos ( γ- MSH), endorfina y angiotensina II. Algunas fibras
descienden de la sustancia gris periacueductal a los centros rombencefálicos,
otras pasan directamente a la médula espinal.
En el rombencéfalo, el núcleo rafe grande y la columna reticular
medial constituyen un importante centro multineuromediador. Las
neuronas en estos sitios contienen serotonina, sustancia P, CCK, hormona
liberadora de tirotropina (TRH), encefalina y dinorfina: algunas neuronas
contienen dos o incluso tres neuromediadores. Las fibras bulboespinales
descendentes pasan al núcleo del tracto espinal del nervio trigémino y su
continuación, la sustancia gelatinosa. Este último se extiende a lo largo
del cordón y contiene poblaciones de neuronas que expresan muchos
neuromediadores diferentes, por ejemplo, GABA, sustancia P,
neurotensina, encefalina y dinorfina. Existe abundante evidencia
fisiológica y farmacológica de que todas estas regiones están
íntimamente relacionadas con el control de las entradas nociceptivas (y
probablemente de otras modalidades).
La estimulación de los sitios del prosencéfalo, incluida la sustancia gris
periventricular, el núcleo posterolateral ventral del tálamo y las cortezas
sensitiva primaria (SI) y parietal posterior, inhibe las células del tracto
espinotalámico. Por el contrario, algunas células del tracto espinotalámico se
excitan mediante la estimulación de la formación reticular medular y la corteza
motora primaria (este último efecto probablemente esté mediado por el tracto
corticoespinal).
La percepción del dolor se describe con más detalle en el capítulo 23.
Reticular ascendente
proyecciones a
talámico intralaminar
núcleos e hipotálamo
Colículo superior
Núcleo rojo
Nervio oculomotor
Mesencéfalo
Colículo inferior
Reticular ascendente
sistema de fibra
Tegmentum del mesencéfalo
Lemnisco medial
Formación reticular pontino
(núcleo reticularis
Pontis oralis)
Terminaciones colaterales en
formación reticular
Vía espinorreticular
Ascendente
fibras colaterales
Las fibras espinorreticulares se entremezclan con las de los tractos espinotalámicos y
ascienden en el cuadrante ventrolateral de la médula espinal (fig.
18,16). La evidencia de estudios en animales sugiere que las células de origen se encuentran en todos los niveles de la médula espinal,
particularmente en los segmentos cervicales superiores. La mayoría de las neuronas se encuentran en la lámina VII, algunas en la lámina
VIII y otras en el asta dorsal, especialmente en la lámina V. La mayoría de los axones de las ampliaciones lumbares y cervicales cruzan la
línea media, pero hay un gran componente no cruzado en las regiones cervicales. La mayoría de los axones están mielinizados. El patrón
de degeneración anterógrada, tanto en estudios post mortem humanos como en animales de experimentación después de cordotomía
anterolateral, indica la existencia de proyecciones espinorreticulares a muchos núcleos de la formación reticular pontomedular medial.
También hay una proyección hacia el núcleo reticular lateral (un núcleo de relevo precerebeloso). Estas proyecciones no parecen estar
organizadas somatotópicamente. Las neuronas espinorreticulares responden a las entradas de la piel o los tejidos profundos. Los
estímulos cutáneos inocuos pueden inhibir o excitar una célula en particular, mientras que los estímulos nocivos suelen ser excitadores. Se
ha propuesto una vía espino-retículo-tálamo-cortical como una vía importante al servicio de la percepción del dolor. Al igual que otras vías
ascendentes, las células del tracto se ven influidas por el control descendente. Por ejemplo, la estimulación eléctrica de la materia gris
periacueductal inhibe las respuestas de ciertas células espinorreticulares a la entrada de aferentes cardiopulmonares. La estimulación de
la formación reticular también altera la actividad de las neuronas espinorreticulares. Se ha propuesto una vía espino-retículo-tálamo-
cortical como una vía importante al servicio de la percepción del dolor. Al igual que otras vías ascendentes, las células del tracto se ven
influidas por el control descendente. Por ejemplo, la estimulación eléctrica de la materia gris periacueductal inhibe las respuestas de
ciertas células espinorreticulares a la entrada de aferentes cardiopulmonares. La estimulación de la formación reticular también altera la
actividad de las neuronas espinorreticulares. Se ha propuesto una vía espino-retículo-tálamo-cortical como una vía importante al servicio
de la percepción del dolor. Al igual que otras vías ascendentes, las células del tracto se ven influidas por el control descendente. Por
ejemplo, la estimulación eléctrica de la materia gris periacueductal inhibe las respuestas de ciertas células espinorreticulares a la entrada
de aferentes cardiopulmonares. La estimulación de la formación reticular también altera la actividad de las neuronas espinorreticulares.
Nervio trigémino
Puente de Varolio
Formación reticular pontino
(núcleo reticularis
Pontis caudalis)
Terminaciones colaterales en
formación reticular
Nervio facial
Nervio Abducens
Ascendente
fibras colaterales
Reticular medular
formación
(núcleo reticularis
gigantocellularis)
Núcleo de espinal
tracto del trigémino
nervio
Médula
Reticular lateral
núcleo
Fibras reticuloespinales pontinas
Nervio hipogloso
Vía espinocervicotalámica
Medular
fibras reticuloespinales
El núcleo cervical lateral es pequeño en el hombre. Se encuentra en el funículo
lateral, ventrolateral al asta dorsal en los dos segmentos cervicales superiores. En
algunas muestras de cordón humano, el núcleo no está claramente definido y
posiblemente esté incorporado al asta dorsal. Recibe axones del tracto
espinocervical, que asciende en el funículo dorsolateral. Las células del tracto se
encuentran en las láminas III-V en todos los niveles de la médula espinal, ipsilateral
al núcleo. La mayoría de las neuronas del núcleo se proyectan hacia el tálamo
contralateral a través del lemnisco medial y algunas se proyectan hacia el
mesencéfalo contralateral. Las dianas talámicas específicas incluyen el núcleo
posterolateral ventral y parte del complejo posterior. Las neuronas del tracto
espinocervical responden al movimiento del cabello, la presión, el pellizco y los
estímulos térmicos y al estímulo muscular de alto umbral; muchos también
responden a estímulos nocivos. Al igual que las células del tracto de otras vías
ascendentes, están bajo control inhibitorio tónico descendente.
Ascendente
fibras espinorreticulares
Tracto ascendente
Tracto descendente
Tracto medular
Figura 18.16 Tractos reticulares. (Redibujado de Carpenter MB 1991 Core Text of
Neuroanatomy, 4th edn. Baltimore: Williams and Wilkins, con permiso del autor y
editor).
en su mayoría mielinizados y ascienden en la sustancia blanca del cuadrante
ventrolateral de la médula espinal, en asociación con los tractos
espinotalámico y espinorreticular.
Las neuronas espinomesencefálicas pertenecen a clases de umbral bajo,
rango dinámico amplio o umbral alto. Sus campos receptivos pueden ser
pequeños o muy complejos y abarcan grandes áreas de superficie del cuerpo.
Muchas células espinomesencefálicas son nociceptivas y es probable que
participen en el componente motivacional-afectivo del dolor. La estimulación
eléctrica de su sitio de terminación en la sustancia gris periacueductal produce
un dolor severo en el hombre. Además, las células de las capas más profundas
del colículo superior, donde hacen sinapsis las fibras espinotectales, son
activadas por estímulos nocivos.
Vía espinomesencefálica
La vía espinomesencefálica consta de varios tractos que ascienden desde la médula
espinal a varias regiones del mesencéfalo. Incluye el tracto espinotectal que se
proyecta hacia el colículo superior, neuronas que hacen sinapsis en la sustancia gris
periacueductal y otras proyecciones de la médula espinal que terminan en el núcleo
parabraquial, los núcleos pretectales y el núcleo de Darkschewitsch. Las células de
origen se localizan a lo largo de la médula espinal, particularmente en los segmentos
cervicales y el agrandamiento lumbosacro, principalmente en la lámina I, pero
también están presentes en las láminas IV-VIII, donde se concentran en la lámina V.
La mayoría son contralaterales , pero también se encuentra un grupo ipsilateral
prominente en los niveles cervicales superiores. Las fibras espinomesencefálicas son
Tracto espino-olivar
El tracto espino-olivar se describe en animales como surgido de neuronas en las láminas
más profundas de la materia gris. Los axones que forman el tracto se cruzan y
267
CAPÍTULO 18

MÉDULA ESPINAL: ORGANIZACIÓN INTERNA
luego asciende super fi cialmente en la unión de los funículos blancos anterior
y lateral, para terminar en los núcleos olivar accesorios dorsal y medial. El
tracto transporta información de los propioceptores de músculos y tendones, y
también de receptores cutáneos. Una ruta funcionalmente similar, el tracto
espino-olivar dorsal, asciende en los funículos blancos dorsales y se traslada en
los núcleos de la columna dorsal al núcleo olivar inferior contralateral. La
información sobre estos tractos en primates es escasa, pero la evidencia post
mórtem después de cordotomías en el hombre ha revelado terminales
axonales degenerados en el núcleo olivar inferior.
Corteza motora (circunvolución precentral)
Tálamo
Extremidad posterior
capsula interna
Grande
piramidal
células de Betz
Fibras para
más bajo
extremidad
Tractos descendentes
Lentiforme
complejo
Las vías descendentes a la médula espinal se originan principalmente en la
corteza cerebral y en numerosos sitios dentro del tronco encefálico (Fig.
18.16, figura 18.17, figura 18.18). Se preocupan por el control del movimiento,
el tono muscular y la postura, la modulación de los mecanismos del reflejo
espinal y la transmisión de información aferente a niveles superiores. También
median en el control de las neuronas autónomas espinales.
Cápsula externa
Claustrum
Fibras
para maletero
Extremo
cápsula
Fibras para
superior
extremidad
Corteza de ínsula
Genu de
interno
cápsula
Tractos corticoespinal y corticobulbar
Extremidad anterior de
capsula interna
Las fibras corticoespinales y corticobulbares surgen de neuronas en la corteza
cerebral. Se proyectan, de una manera organizada somatotópicamente, a neuronas
que se encuentran principalmente en el lado contralateral de la médula espinal o del
tronco encefálico, respectivamente (fig. 18.17). La mayoría de las fibras
corticoespinales y corticobulbares proceden de células situadas en la corteza motora
primaria (área 4) y la corteza premotora (área 6). Una pequeña contribución proviene
de las células de la circunvolución poscentral (corteza somatosensorial; áreas 3, 1 y 2)
y la corteza parietal adyacente (área 5). En el mono, el 30% de las fibras
corticoespinales surgen del área 4, el 30% del área
6 y 40% de las regiones parietales. Las células de origen de las fibras corticoespinales
y corticobulbares varían de tamaño en las diferentes áreas corticales y se agrupan en
grupos o tiras. Las células más grandes (neuronas piramidales gigantes o células de
Betz) se encuentran en la corteza motora primaria de la circunvolución precentral.
Las fibras corticoespinales y corticobulbares descienden a través de la
sustancia blanca subcortical para entrar en la rodilla y la rama posterior de la
cápsula interna. Luego pasan a través de la parte ventral del mesencéfalo en el
crus cerebri. A medida que continúan caudalmente a través de la
protuberancia, se separan de su superficie ventral y se fragmentan en
fascículos mediante fibras pontocerebelosas que discurren transversalmente.
Las fibras corticobulbares dejan de terminar en asociación con los núcleos
motores de los nervios craneales del mesencéfalo, la protuberancia y la
médula. En el bulbo raquídeo, las fibras corticoespinales residuales forman un
haz discreto, la pirámide (fig. 19.2) que forma una columna longitudinal
prominente en la superficie ventral del bulbo raquídeo. Por lo tanto, el tracto
corticoespinal también se conoce como tracto piramidal. Cada pirámide
contiene alrededor de un millón de axones de diámetro variable. μ metro; El
9% tiene diámetros de 5 a 10 μ metro; y menos del 2% tienen diámetros de 11
a 22 μ metro. Los axones de mayor diámetro surgen de las células de Betz
gigantes.
Justo en posición rostral al nivel de la unión espinomedular,
aproximadamente el 75-90% de las fibras corticoespinales de la pirámide
cruzan el plano mediano en la decusación piramidal (decusación de las
pirámides) y continúan caudalmente como el haz corticoespinal lateral. El
resto de las fibras continúan sin cruzar como el haz corticoespinal ventral.
El tracto lateral también contiene algunas fibras corticoespinales no
cruzadas. El haz corticoespinal lateral (fig. 18.17) desciende en el funículo
lateral a lo largo de la mayor parte de la médula espinal. Ocupa un área
ovalada, ventrolateral al asta dorsal y medial al haz espinocerebeloso
dorsal (fig. 18.9). En las regiones lumbar y sacra, donde el haz
espinocerebeloso dorsal está ausente, el haz corticoespinal lateral
alcanza la superficie dorsolateral de la médula. Mientras desciende
El haz corticoespinal ventral más pequeño (fig. 18.17) desciende en el
funículo ventral. Se encuentra cerca de la fisura media ventral y está separada
de ella por el fascículo sulcomarginal (fig. 18.9). El tracto corticoespinal ventral
disminuye a medida que desciende y, por lo general, desaparece por completo
en los niveles de la médula torácica media. Puede estar ausente o, muy
raramente, contener casi todas las fibras corticoespinales. Cerca de su
terminación, la mayoría de las fibras del haz cruzan el plano medio en la
comisura blanca ventral para hacer sinapsis con las neuronas contralaterales.
Por tanto, la gran mayoría de las fibras corticoespinales, independientemente
del trayecto por el que descienden, terminan en la médula espinal en el lado
contralateral a su origen cortical.
El conocimiento de la terminación detallada de las fibras corticoespinales se basa
en gran medida en estudios en animales, pero se complementa con datos de
Núcleo caudado (cabeza)
Tracto corticoespinal
Mesencéfalo
Tracto temporopontino
Crus cerebri
Tracto frontopontino
Nervio oculomotor
Puente de Varolio
Fibras longitudinales
en porción basilar
de pons
Nervio Abducens
Médula
Nervio hipogloso
Piramidal
Médula
Decusación piramidal
Tracto corticoespinal lateral
Tracto corticoespinal anterior
C8
A las terminaciones motoras
en los músculos de
antebrazo y mano
A las terminaciones motoras
en intercostal y
segmentario
músculos de la espalda
T4
Fibra de raíz ventral
A las terminaciones motoras
en glúteo medio
y tibial anterior
L4
A los segmentos sacros del cordón
Figura 18.17 Los tractos corticoespinales. (Redibujado de Carpenter MB 1991 Core
Text of Neuroanatomy, 4th edn. Baltimore: Williams and Wilkins, con permiso del
autor y editor).
268
gramo
mi
L
SECCIÓN 3
metro
r
A
k
norte
tu
r
T
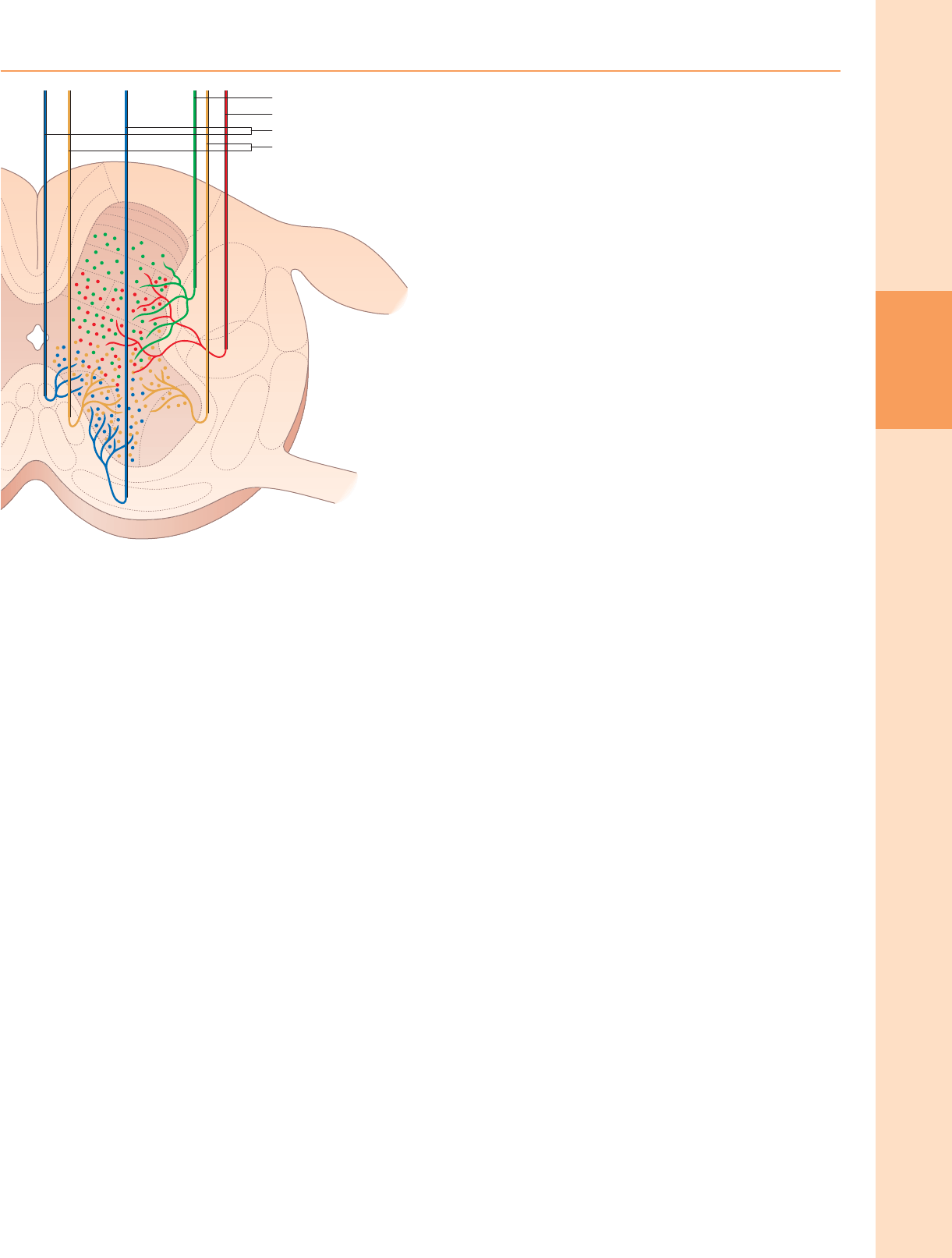
Organización interna
Tracto corticoespinal lateral
Tracto rubroespinal
Tractos vestibuloespinales
Tractos reticuloespinales
accidente vascular cerebral o "ictus", resulta en una hemiplejía
contralateral. La parálisis es inicialmente flácida, pero luego se vuelve
espástica y es más marcada en los músculos distales de las extremidades,
especialmente en aquellos relacionados con los movimientos individuales
de los dedos y la mano. Los signos asociados en el lado paralizado son:
reflejos tendinosos profundos hiperactivos; hipertonicidad; la pérdida de
reflejos super fi ciales abdominales y cremastéricos; y la aparición de
flexión dorsal de los dedos de los pies (signo de Babinski) en respuesta a
las caricias en la planta del pie. Este último suele interpretarse como
patognomónico de daño corticoespinal, pero no siempre está presente
en pacientes con lesiones corticoespinales confirmadas. Es más,
Algunas de las secuelas del daño por accidente cerebrovascular en la
cápsula interna, en particular la hiperreflexia y la hipertonía, se deben a la
participación de otras vías además del tracto corticoespinal. Estos incluyen
fibras corticales descendentes a los núcleos del tronco encefálico, como los
núcleos vestibular y reticular, que a su vez dan lugar a proyecciones
descendentes que in fl uyen en la actividad de las neuronas motoras.
Tracto rubroespinal
El haz rubroespinal surge de neuronas en la parte magnocelular caudal del
núcleo rojo (una masa ovoide de células situada en el centro del tegmento del
mesencéfalo (pág. 290)). Esta parte del núcleo contiene entre 150 y 200
neuronas grandes, intercaladas con neuronas más pequeñas.
El origen, la localización, la terminación y las funciones de las conexiones
rubroespinales están mal definidas en el hombre y el tracto parece ser
rudimentario. Las fibras rubroespinales se cruzan en la decusación tegmental
ventral y descienden en el funículo lateral de la médula, donde se encuentran
ventrales y se entremezclan con las fibras del haz corticoespinal lateral (fig.
18.9). En los animales, el tracto desciende hasta los niveles lumbosacros,
mientras que en el hombre parece proyectarse sólo a los tres segmentos
superiores del cordón cervical. Las fibras rubroespinales se distribuyen a las
partes laterales de las láminas V-VI y la parte dorsal de la lámina VII de la
sustancia gris espinal. Las zonas terminales del tracto corresponden a las de
las fibras corticoespinales de la corteza motora. Los estudios en animales
demuestran que los efectos de las fibras rubroespinales en α y γ las neuronas
motoras son similares a las de las fibras corticoespinales.
Figura 18.18 Un esquema simplificado de algunas de las principales vías
descendentes de la médula espinal, incluidas sus zonas superpuestas de
terminación en la sustancia gris. Dentro de la materia gris las líneas punteadas
muestran el patrón laminar, mientras que dentro de la materia blanca son una
guía aproximada de la topografía de los tractos.
estudios post mortem en cerebros humanos utilizando métodos de degeneración
anterógrada. Se cree que la mayoría de las fibras corticoespinales terminan
contralateralmente en las interneuronas de las partes laterales de las láminas IV-VI y
de las partes lateral y medial de la lámina VII. Algunos también se distribuyen a la
lámina VIII de forma bilateral. Los terminales también se asocian con grupos de
células de neuronas motoras contralaterales en la lámina IX, en el grupo dorsolateral
y en las partes laterales de los grupos central y ventrolateral (fig. 18.18).
Las fibras corticoespinales de la corteza frontal, incluidas las áreas motoras y
premotoras 4 y 6, terminan principalmente en las interneuronas de las láminas V-
VIII, y la concentración más densa termina en la parte lateral de la lámina VI. Influyen
α y γ neuronas motoras de la lámina IX, a través de estas interneuronas. Debido a
que las dendritas generalizadas de las neuronas multipolares en la lámina IX
penetran en la lámina VII, los contactos axodendríticos monosinápticos directos
también ocurren en grandes α neuronas motoras. La terminación directa en las
neuronas motoras es más abundante en los agrandamientos de la columna.
La evidencia experimental muestra que los axones corticoespinales precentrales
influyen en las actividades de ambos α y γ neuronas motoras, que facilitan los músculos fl
exores e inhiben los extensores, que son los efectos opuestos a los mediados por las fibras
vestibuloespinales laterales. La evidencia de estudios en animales muestra que las
proyecciones directas desde las áreas corticales precentrales hasta las neuronas motoras
espinales se relacionan con movimientos de precisión y muy fraccionados de las
extremidades. En consecuencia, en los primates, las fibras corticoespinales precentrales se
distribuyen principalmente a las neuronas motoras que inervan los músculos distales de las
extremidades. Las proyecciones corticoespinales pueden utilizar glutamato o aspartato, a
menudo co-localizados, como neurotransmisores excitadores.
Las fibras corticoespinales de origen parietal terminan principalmente en el asta
dorsal contralateral, en las partes laterales de las láminas IV-VI y lámina VII.
Filogenéticamente, estas fibras representan la parte más antigua del sistema
corticoespinal. Los axones de la corteza sensorial terminan principalmente en las
láminas IV y V. Están relacionados con la modulación supraespinal de la transmisión
de impulsos aferentes a los centros superiores, incluida la corteza motora.
Los estudios experimentales en primates indican que la sección aislada de
fibras corticoespinales a nivel de la pirámide (piramidotomía) produce parálisis
o paresia flácida de las extremidades contralaterales y pérdida de los
movimientos independientes de la mano y los dedos. Destrucción de las fibras
corticoespinales a nivel de la cápsula interna, comúnmente causada por una
Tracto tectoespinal
El tracto tectoespinal surge de neuronas en las capas intermedia y
profunda del colículo superior del mesencéfalo. Cruza ventral a la
sustancia gris periacueductal en la decusación tegmental dorsal y
desciende en la parte medial del funículo ventral de la médula espinal
(fig. 18.9). Las fibras del tracto se proyectan solo a los segmentos
superiores del cordón cervical y terminan en las láminas VI-VIII.
Establecen conexiones polisinápticas con las neuronas motoras que
sirven a los músculos del cuello, facilitando las que inervan los músculos
contralaterales e inhibiendo las que inervan los ipsilaterales. En los
animales, la estimulación eléctrica unilateral del colículo superior provoca
el giro de la cabeza hacia el lado contralateral, un efecto mediado
principalmente a través del tracto tectoespinal.
Tractos vestibuloespinales
El complejo nuclear vestibular se encuentra en la parte lateral del piso del
cuarto ventrículo, al nivel de la unión pontomedular. Da lugar a los
tractos vestibuloespinal lateral y medial, que son funcional y
topográficamente distintos (fig. 18.19).
El haz vestibuloespinal lateral surge de neuronas pequeñas y grandes del
núcleo vestibular lateral (núcleo de Deiters). Desciende ipsolateralmente,
inicialmente en la periferia de la sustancia blanca espinal ventrolateral, pero
luego migra hacia la parte medial del funículo ventral a niveles espinales
inferiores. Las fibras de este tracto están organizadas somatotópicamente. Así,
las fibras que se proyectan a los segmentos cervical, torácico y lumbosacro del
cordón surgen de neuronas en las partes rostroventral, central y dorsocaudal,
respectivamente, del núcleo vestibular lateral. Las fibras vestibuloespinales
laterales terminan ipsolateralmente, principalmente en la parte medial del asta
ventral en la lámina VIII y en la parte medial de la lámina VII. Los axones del
tracto vestibuloespinal lateral excitan las neuronas motoras de los músculos
extensores del cuello, la espalda y las extremidades. γ Es probable que
también se faciliten las neuronas motoras. Los axones del tracto
vestibuloespinal lateral también inhiben las neuronas motoras de los músculos
fl exores de las extremidades, a través de interneuronas inhibidoras.
El haz vestibuloespinal medial (fig. 18.19) surge principalmente de
neuronas en el núcleo vestibular medial, pero algunas también se localizan en
los núcleos vestibulares inferior y lateral. El tracto vestibuloespinal medial
desciende en el fascículo longitudinal medial hacia el ventral.
269
CAPÍTULO 18
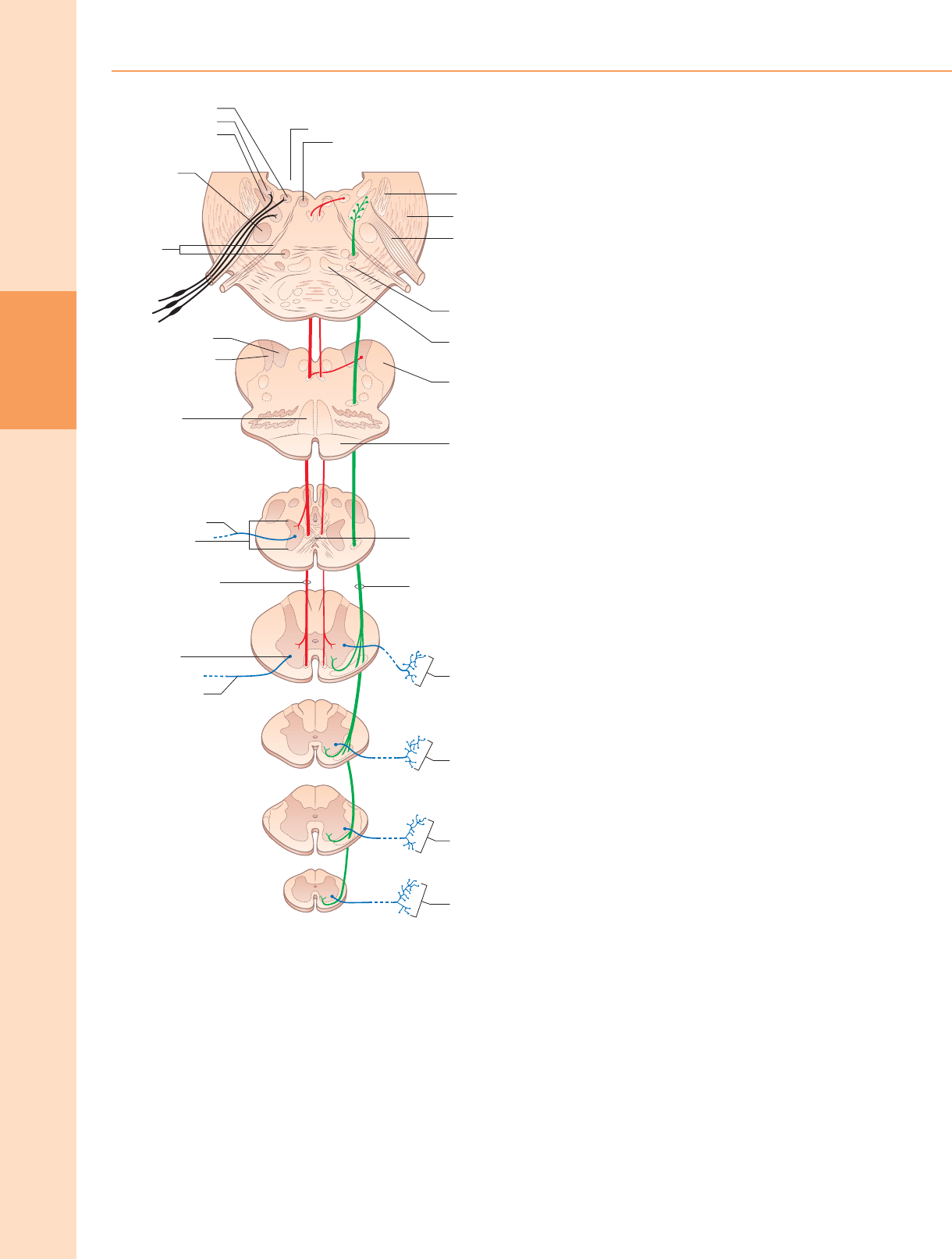
MÉDULA ESPINAL: ORGANIZACIÓN INTERNA
Núcleos vestibulares:
Núcleo medial
Núcleo superior
Núcleo lateral
los núcleos reticulares pontinos oral y caudal y el núcleo reticular
gigantocelular en la médula. Las fibras pontinas descienden,
principalmente ipsolateralmente, en el funículo ventral del cordón. Las
fibras medulares descienden, tanto ipsilateral como contralateralmente,
en el funículo ventral y la parte ventral del funículo lateral. Estas fibras
tienen muchas colaterales y dos tercios de las neuronas reticuloespinales
que llegan a la médula cervical también descienden a niveles
lumbosacros. Los terminales de las fibras reticuloespinales se distribuyen
hasta la lámina VIII y las partes central y medial de la lámina VII. Las
terminales reticuloespinales medulares están más ampliamente
distribuidas, terminando adicionalmente en las partes laterales de las
láminas VI y VII y también directamente en las neuronas motoras. Las
terminaciones de las fibras reticuloespinales que se originan en la médula
son, en general,
Ambos α y γ las neuronas motoras están influenciadas por fibras reticuloespinales, a
través de conexiones polisinápticas y monosinápticas. La evidencia fisiológica muestra que
las fibras reticuloespinales de fuentes pontinas excitan las neuronas motoras de los
músculos axiales y de las extremidades, mientras que las fibras medulares excitan o inhiben
las neuronas motoras de los músculos cervicales y excitan las neuronas motoras de los
músculos axiales. Funcionalmente, el tracto reticuloespinal medial se ocupa de la postura, la
dirección de los movimientos de la cabeza y el tronco en respuesta a estímulos externos y
los movimientos crudos y estereotipados de las extremidades.
El haz reticuloespinal lateral se encuentra en el funículo lateral de la médula
espinal, estrechamente asociado con los haces rubroespinal y corticoespinal
lateral (fig. 18.9). Sus fibras proceden de neuronas del campo tegmental
ventrolateral de la protuberancia. Las fibras se cruzan en el bulbo raquídeo
rostral y se proyectan, con un alto grado de colateralización, a lo largo de la
médula espinal. Los axones de este tracto terminan en las láminas I, V y VI, y
también de forma bilateral en el núcleo cervical lateral. La evidencia sugiere
que esta vía está involucrada en el control de la percepción del dolor y en las
funciones motoras.
Ventrículo IV
Núcleo abducens
Núcleo espinal de
nervio trigémino
Pontomedular
unión
Cuerpo Restiforme
Brachium pontis
Nervio vestibular
Nervio facial
y núcleo
Núcleo de
aceituna superior
Núcleo vestibular medial
Núcleo vestibular inferior
Lemnisco medial
Cuerpo Restiforme
Médula
Lemnisco medial
Pirámide
Médula
Nervio espinal accesorio
Columna gris anterior
Decusación piramidal
Tracto vestibuloespinal medial
Tracto vestibuloespinal lateral
Tracto intersticioespinal
El tracto intersticioespinal surge de las neuronas del núcleo intersticial (de Cajal) y el
área circundante inmediata, y desciende a través del fascículo longitudinal medial
hacia el funículo ventral de la médula espinal. Sus fibras se proyectan,
principalmente ipsilateralmente, hasta los niveles lumbosacros, y se distribuyen
mayoritariamente hacia la parte dorsal de la lámina VIII y la parte dorsal contigua de
la lámina VII. Establecen algunas conexiones monosinápticas con las neuronas
motoras que inervan los músculos del cuello, pero sus conexiones principales son
disinápticas con las neuronas motoras que inervan los músculos de las extremidades.
C3
Célula del asta anterior
Placas de extremo de motor
en trapecio y
músculos escalenos
Fibra nerviosa de la raíz ventral
T4
Placas de extremo de motor
en intercostal
y segmentario
músculos de la espalda
Tracto solitarioespinal
El tracto solitarioespinal es un pequeño grupo de fibras en su mayoría
cruzadas que surge de neuronas en la parte ventrolateral del núcleo solitario
de la médula. Descendiendo en el funículo ventral y la parte ventral del
funículo lateral del cordón, estos axones terminan en neuronas motoras
frénicas que inervan el diafragma y las neuronas motoras torácicas que
inervan los músculos intercostales. Una vía con un curso y terminaciones algo
similares a la del tracto solitarioespinal se origina en el núcleo retroambiguus.
Ambas vías sirven a las actividades respiratorias impulsando los músculos
inspiratorios, y algunos axones descendentes del núcleo retroambiguo facilitan
las motoneuronas espiratorias. Existe evidencia clínica de que la cordotomía
ventrolateral bilateral a niveles cervicales altos suprime los movimientos
ventilatorios rítmicos.
L3
Placas de extremo de motor
en cuadriceps
femoral
S2
Placas de extremo de motor
en gastrocnemio
Figura 18.19 Los tractos vestibuloespinales. (Redibujado de Carpenter MB 1991
Core Text of Neuroanatomy, 4th edn. Baltimore: Williams and Wilkins, con permiso
del autor y editor).
Fibras hipotálamoespinales
Existen fibras hipotálamospinales en animales. Surgen del núcleo
paraventricular y otras áreas del hipotálamo y descienden
ipsilateralmente, principalmente en la región dorsolateral del cordón,
para distribuirse a neuronas preganglionares simpáticas y parasimpáticas
en la columna intermediolateral. Las fibras del núcleo paraventricular
muestran inmunorreactividad a oxitocina y vasopresina. También se
distribuyen a las láminas I y X. Las fibras descendentes del grupo de
células dopaminérgicas (A11) situadas en el hipotálamo caudal inervan
neuronas preganglionares simpáticas y neuronas del asta dorsal. La
existencia de vías similares en el hombre puede inferirse de los déficits
simpáticos ipsolaterales (p. Ej., Síndrome de Horner), que siguen a las
lesiones del hipotálamo, el tronco encefálico tegmental lateral o el
funículo lateral del cordón.
funículo de la médula espinal donde se encuentra cerca de la línea media en el
llamado fascículo sulcomarginal (Fig. 18.9). A diferencia del tracto lateral, contiene
fibras cruzadas y no cruzadas, y no se extiende más allá del nivel medular torácico.
Las fibras del tracto medial se proyectan principalmente a los segmentos de la
médula cervical, terminando en la lámina VIII y la parte dorsal adyacente de la
lámina VII. Los datos de la estimulación de los núcleos vestibulares en animales
indican que los axones del tracto medial inhiben monosinápticamente las neuronas
motoras que inervan los músculos axiales del cuello y la parte superior de la espalda.
Tractos reticuloespinales
Los tractos reticuloespinales pasan de la formación reticular del tronco encefálico a
la médula espinal. El conocimiento detallado de sus orígenes y conexiones se ha
obtenido principalmente de estudios en animales.
El haz reticuloespinal medial (fig. 18.16) se origina en los campos tegmentales
medial de la protuberancia y la médula. Las principales fuentes son
Vías espinales monoaminérgicas
Los grupos de células monoaminérgicas utilizan dopamina, adrenalina
(epinefrina), noradrenalina (norepinefrina) y 5-HT (5-hidroxitriptamina,
serotonina) como neurotransmisores. Ocurren ampliamente en todo el
270
SECCIÓN 3
Lesiones de la médula espinal
tronco encefálico y en el hipotálamo. Se proyectan rostralmente a muchas
áreas del prosencéfalo y caudalmente a la médula espinal y parecen
preocuparse por la modulación de la transmisión sensorial y el control de las
actividades neuronales motoras somáticas y autónomas.
Las proyecciones a la médula espinal surgen de varias fuentes. Las
proyecciones coeruleospinales se originan a partir de los grupos de células
noradrenérgicas A4 y A6 en el complejo locus coeruleus en la protuberancia y
descienden a través de la sustancia blanca ventrolateral para inervar todos los
segmentos del cordón bilateralmente. Terminan en la sustancia gris dorsal
(láminas IV-VI) y los cuernos intermedios y ventrales. También se proyectan
ampliamente a las neuronas parasimpáticas preganglionares del cordón sacro.
Las fibras noradrenérgicas descendentes, que surgen de los grupos de células
tegmentales laterales A5 y A7 de la protuberancia, viajan en la sustancia blanca
dorsolateral. Se distribuyen a las láminas I-III, y particularmente al cuerno gris
intermedio. Las fibras descendentes de los grupos de células adrenérgicas C1 y
C3 del bulbo raquídeo se han rastreado hasta el funículo anterior del cordón y
se distribuyen ampliamente a la columna intermediolateral. Las fibras
dopaminérgicas que se proyectan a la médula espinal viajan por la vía
hipotalámica-espinal.
Los núcleos del rafe pallidus (B1), obscurus (B2) y magnus (B3) en el
tallo cerebral dan lugar a dos haces descendentes serotoninérgicos. El
haz espinal del rafe lateral, de las neuronas B3, se ocupa del control de la
nocicepción. Desciende cerca del haz corticoespinal lateral y termina en el
asta dorsal (láminas I, II y V). El haz ventral, compuesto principalmente
por axones de neuronas B1, viaja en la parte medial de la columna blanca
ventral y termina en el asta ventral (láminas VIII y IX). Facilita las
motoneuronas extensoras y fl exoras. Algunas fibras serotoninérgicas
descendentes se proyectan a las neuronas preganglionares simpáticas y
están relacionadas con el control central de la función cardiovascular.
El tracto de Lissauer
El haz de Lissauer, o haz dorsolateral, se encuentra entre el vértice del
asta dorsal y la superficie de la médula espinal, donde rodea las fibras de
la raíz dorsal entrantes. Está presente en toda la médula espinal y está
más desarrollada en las regiones cervicales superiores.
El tracto consta de axones finos mielinizados y no mielinizados. Muchas son
las ramas de los axones en los haces laterales de las raíces dorsales. Estos
axones se bifurcan en ramas ascendentes y descendentes a medida que
ingresan al cordón. Las ramas viajan en el tracto de Lissauer por uno o dos
segmentos y emiten colaterales, que terminan en y alrededor de las neuronas
en el asta dorsal. El tracto también contiene fibras proprioespinales, algunas
de las cuales son axones cortos de neuronas de sustancia gelatinosa pequeña,
que vuelven a entrar en el asta dorsal.
LESIONES DE LA MÉDULA ESPINAL
La compresión mecánica y el daño isquémico secundario al tejido nervioso
subyacente provocan una enfermedad de la médula espinal quirúrgicamente
relevante (mielopatía). El sitio y el nivel de daño de la médula determinan el
síndrome clínico particular, por ejemplo, si la lesión afecta a la médula espinal
cervical superior o inferior, torácica o lumbosacra. En cada uno de estos niveles, los
síntomas y signos están determinados por la destrucción directa del tejido
segmentario, es decir, el daño distribuido transversalmente, y la desconexión de los
tractos suprasegmentarios ascendentes y descendentes por encima y por debajo del
nivel de una lesión, es decir, daño distribuido longitudinalmente (Fig.
18,20). Por ejemplo, una lesión de la médula espinal cervical inferior daña las
contribuciones sensoriales y motoras segmentarias a las raíces nerviosas y el plexo
braquial, causando pérdida sensorial, debilidad y atrofia de los músculos y pérdida
de reflejos tendinosos en las extremidades superiores. La interrupción de las vías
sensoriales ascendentes en las columnas lateral y dorsal de la médula espinal
cervical conduce a la pérdida de la sensibilidad al dolor y la temperatura (tractos
espinotalámicos laterales) y al tacto y la propiocepción (fascículos dorsales) por
debajo del 'nivel sensorial' correspondiente al segmento de la medula espinal.
La lesión de los tractos corticoespinales descendentes en las columnas laterales
de la médula espinal produce una paraparesia espástica, es decir, aumento del tono
de los músculos, debilidad de los movimientos de fl exión, reflejos tendinosos
exagerados y reflejos superficiales anormales, por ejemplo, respuestas plantares
extensores y ausencia de reflejos abdominales. Las vías descendentes a la vejiga se
interrumpen y esto produce una "vejiga neurogénica".
Los mismos principios se aplican a las lesiones en otros niveles de la
médula espinal y se ilustran en forma de diagrama en la figura 18.20.
El síndrome clínico preciso está determinado solo por el sitio anatómico y
no por la patología. Sin embargo, es de utilidad práctica clasificar las lesiones
en función de su relación anatómica con la médula espinal y las meninges, es
decir, si son extradurales, intradurales o intramedulares (ver tabla). Esta
clasificación anatómica proporciona una guía para las probabilidades de
diagnóstico, así como una ayuda para la interpretación neurorradiológica
antes de la intervención neuroquirúrgica. Por ejemplo, los neurofibromas son
frecuentes en el canal espinal cervical, los meningiomas en el canal espinal
torácico y los ependimomas en el canal espinal lumbosacro. La enfermedad
degenerativa de la columna vertebral es común en las vértebras cervicales y
lumbosacras, pero rara en las vértebras torácicas. Lesiones intramedulares
anteriores y centrales discretas, p. Ej. debido a siringomielia y angiomas
respectivamente, destruyen preferentemente las vías espinotalámicas en las
columnas anterolaterales y áreas centrales de la médula espinal. Esto conduce
a una pérdida sensorial 'disociada' característica,
es decir, pérdida de la sensación de dolor y temperatura, pero con conservación de la
sensación táctil y la propiocepción en el nivel de la lesión y por debajo del mismo.
Vías propioespinales
Las vías propioespinales (fasciculi proprii) consisten en las fibras ascendentes y
descendentes de las neuronas espinales intrínsecas. Se ponen en contacto con otras
neuronas dentro del mismo segmento y / o en segmentos más distantes de la médula
espinal y, por lo tanto, sirven a la integración y coordinación intrasegmentarias e
intersegmentarias. La mayoría de las neuronas espinales son neuronas proprioespinales, la
mayoría de las cuales se encuentran en las láminas V-VIII. Las fibras propioespinales se
concentran principalmente alrededor de los márgenes de la sustancia gris (Fig. 18.9), pero
también se dispersan de manera difusa en los funículos blancos.
El sistema proprioespinal juega un papel importante en las funciones de la
columna. Las vías descendentes terminan en subgrupos específicos de neuronas
proprioespinales y estas, a su vez, se transmiten a las neuronas motoras y otras
neuronas espinales. El sistema media todas aquellas funciones automáticas que
continúan después de la sección de la médula espinal, por ejemplo, las actividades
sudomotoras y vasomotoras, las funciones del intestino y la vejiga.
Algunos axones proprioespinales son muy cortos y abarcan solo un segmento,
mientras que otros recorren toda la longitud del cordón. Los axones más cortos se
encuentran inmediatamente adyacentes a la sustancia gris y los más largos están
situados más periféricamente. Las neuronas propioespinales se pueden clasificar
según la longitud de sus axones como neuronas largas, intermedias o cortas. Las
neuronas proprioespinales largas distribuyen sus axones a lo largo de la médula,
principalmente a través de los funículos ventrales y laterales; sus cuerpos celulares
están en la lámina VIII y la parte dorsalmente contigua de la lámina VII. Los axones
de las neuronas proprioespinales largas de la médula cervical descienden
bilateralmente, mientras que los de las neuronas lumbosacras correspondientes
ascienden principalmente contralateralmente. La mayoría de las fibras están bien
(menos de 3 μ m de diámetro). Algunos son los primeros axones del tracto espinal
que se mielinizan. Las neuronas proprioespinales intermedias ocupan las partes
central y medial de la lámina VII y se proyectan principalmente ipsilateralmente. Las
neuronas proprioespinales cortas se encuentran en las partes laterales de las
láminas V-VIII y sus axones corren ipsilateralmente en el funículo lateral.
Las fibras propioespinales en las diferentes partes de los funículos blancos
se distribuyen preferentemente a regiones específicas de la sustancia gris
espinal. En los aumentos de tamaño de la columna, las fibras proprioespinales
del funículo dorsolateral se proyectan hacia las partes dorsal y lateral de la
zona intermedia, y también hacia las neuronas motoras espinales que inervan
los músculos distales de las extremidades, especialmente los de la mano y el
pie. Las fibras proprioespinales en la parte ventral del funículo ventrolateral se
distribuyen a las partes central y medial de la lámina VII ya las neuronas
motoras de los músculos proximales de la cintura y las extremidades. Otras
fibras proprioespinales corren en la parte medial del funículo ventral y viajan
principalmente a la parte ventromedial de la zona intermedia, que de manera
característica contiene neuronas proprioespinales largas,
Clasificación de las lesiones de la médula espinal
Lesiones extradurales A. Trastorno de la osteoartritis degenerativa (osteofitos y
discos intervertebrales prolapsados vertebrales)
columna Tumor (carcinoma metastásico, mieloma, linfoma,
cordoma, sarcoma)
B. Absceso
C.Hematoma (complicación de sangrado, diátesis o
anticoagulación)
Lesiones intradurales Meningioma, neurofibroma, lipoma, angioma
Intramedular
lesiones
Siringomielia (malformación de Arnold-Chiari),
angioma, glioma, ependimoma, tumor
epidermoide
271
CAPÍTULO 18

Organización de la médula espinal y vías nerviosas.en.es.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.