
1
UNION NEUROMUSCULAR
las neuronas que estimulan la contracción de las fibras musculares esqueléticas se denominan neuronas
motoras somáticas. Cada neurona motora somática tiene un axón filiforme que se extiende desde el encéfalo
o la médula espinal hasta un grupo de fibras musculares esqueléticas. Una fibra muscular se contrae en
respuesta a uno o más potenciales de acción
que se propagan a lo largo de su sarcolema y a través de su sistema de túbulos T. Los potenciales de acción
musculares se originan en la unión neuromuscular (UNM), la sinapsis entre una neurona motora somática y
una fibra muscular esquelética (Figura 10.9a). Una sinapsis es una región donde se produce la comunicación
entre dos neuronas o entre una neurona y una célula diana; en este caso, entre una neurona motora somática
y una fibra muscular. En la mayoría de las sinapsis, las dos células están separadas por un pequeño espacio,
denominado hendidura sináptica. Como las células no están en contacto físico, el potencial de acción no
puede “saltar la hendidura” de una célula a otra, por lo que la primera célula se comunica con la segunda y
libera una sustancia química conocida como neurotransmisor.
En la UNM, el extremo de la neurona motora, denominado axón terminal, se divide en un racimo de bulbos
sinápticos terminales (Figura 10.9a, b), la zona nerviosa de la UNM. Suspendidas en el cito-sol dentro de cada
bulbo sináptico terminal, hay cientos de sacos delimitados por una membrana llamados vesículas sinápticas.
Dentro de cada vesícula sináptica, se encuentran miles de moléculas de acetilcolina, que se abrevia ACh, el
neurotransmisor liberado en la UNM.
La región del sarcolema frente a los bulbos sinápticos terminales, denominada placa motora (Figura 10.9b,
c), es la fibra muscular parte de la UNM. Dentro de cada placa motora, hay de 30 a 40 millones de receptores
colinérgicos
P R E G U N T A S D E R E V I S I Ó N
5. Describa las partes de una neurona y la función de cada una.
6. Proporcione varios ejemplos de las clasificaciones estructurales y funcionales de las neuronas.
7. ¿Qué es el neurolema? ¿Cuál es su importancia?
8. En referencia al sistema nervioso, ¿qué es un núcleo?
SEÑALES ELÉCTRICAS EN LAS NEURONAS
1) Los potenciales graduados (descritos más adelante) se utilizan sólo para las comunicaciones a corta
distancia.
(2) Los potenciales de acción (también descritos más adelante) permiten la comunicación con lugares
cercanos y lejanos, dentro del cuerpo. Recuerde que un potencial de acción en una fibra muscular se
denomina potencial de acción muscular. Cuando se produce un potencial de acción en una neurona (célula
nerviosa), recibe el nombre de potencial de acción nervioso (impulso nervioso)
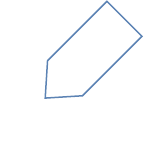
Canales iónicos
Cuando los canales iónicos están abiertos permiten el paso de iones específicos a través de la membrana
plasmática, los cuales siguen su gradiente electroquímico: una diferencia en la concentración de sustancias
químicas sumada a una diferencia de cargas eléctricas.
Recuerde que los iones se mueven desde áreas en las que se encuentran en mayor concentración hacia
regiones en las que se hallan en menor concentración (el componente químico del gradiente). Además, los
cationes con cargas positivas se desplazan hacia zonas con cargas negativas, y los aniones con cargas
negativas se dirigen hacia áreas con cargas positivas (el componente eléctrico del gradiente). A medida que
los iones se mueven, crean una corriente eléctrica que puede cambiar el potencial de membrana.
Las señales eléctricas que producen las neuronas y las fibras musculares dependen de cuatro tipos de canales:
canales pasivos, canales dependientes de ligandos, canales accionados mecánicamente y canales
dependientes del voltaje:
•
canales pasivos alternan al azar entre las posiciones abierta y cerrada, la permeabilidad de la
membrana para el K+ es mucho más alta que para el Na+.
•
canales dependientes de ligandos se abren y se cierran en res-puesta a estímulos ligandos (químicos)
específicos. Los canales dependientes de ligandos se localizan en las dendritas de algunas neuronas
sensitivas, como los receptores del dolor, y en las dendritas y en los cuerpos celulares de
interneuronas y neuronas motoras. La generación de potenciales graduados y de potenciales de
acción depende de dos características fundamentales de la membrana plasmática de las células
2
excitables: la existencia de un potencial de membrana de reposo y la presencia de tipos específicos
de canales iónicos.
•
Los canales accionados mecánicamente se abren o se cierran luego de una estimulación mecánica,
que puede tomar la forma de una vibración (como las ondas sonoras), el tacto, la presión o el
estiramiento de un tejido. Los receptores auditivos del oído, en los receptores que monitorizan el
estiramiento de los órganos internos y en los receptores para el tacto y la presión de la piel.
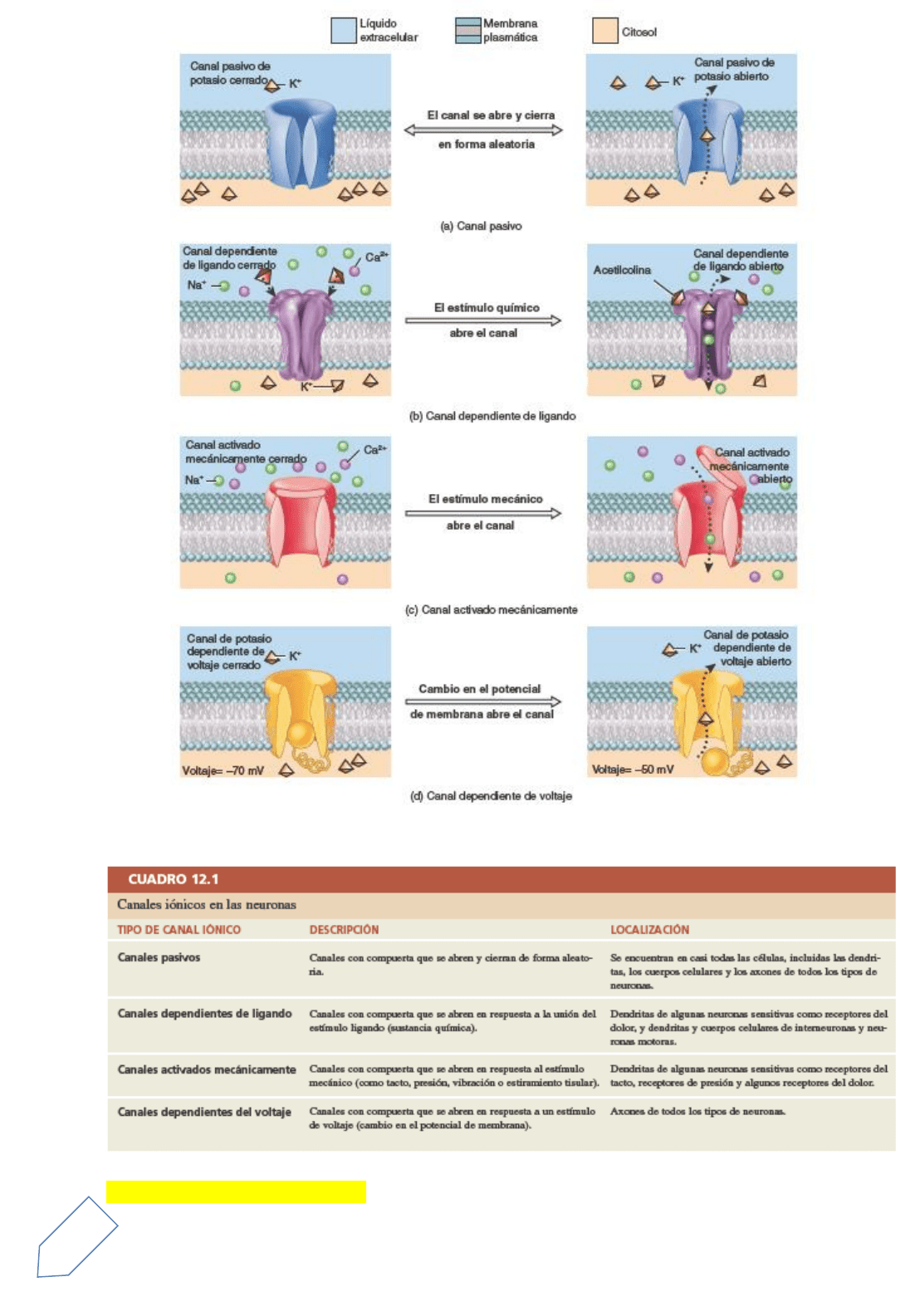
En las células excitables, este voltaje se denomina potencial de membrana de reposo. El potencial de
membrana es similar al volta-je que se almacena en un acumulador. Si se conectan los terminales
positivo y negativo de un acumulador con un cable, los electrones fluirán a lo largo de éste. Ese flujo
de partículas cargadas se denomina corriente. En las células vivas, el flujo de iones (más que de
electrones) constituye las corrientes eléctricas.
•
Los canales dependientes del voltaje se abren en respuesta a un cambio en su potencial de
membrana (voltaje
Los potenciales graduados y los potenciales de acción se producen porque las membranas de las neuronas
contienen muchos tipos diferentes de canales iónicos que se abren o se cierran, en respuesta a estímulos
específicos. Como la bicapa lipídica que constituye la membrana plasmática es un buen aislante eléctrico, las
vías principales que utiliza el flujo iónico para pasar a través de la membrana son los canales iónicos.
3
Potencial de membrana en reposo
4
El potencial de membrana en reposo es la consecuencia de la pequeña acumulación de iones negativos en el
citosol a lo largo de la superficie interna de la membrana y de la acumulación semejante de iones positivos
en el líquido extracelular, a lo largo de la superficie externa de la membrana. Esta separación de cargas
eléctricas positivas y negativas es una forma de energía potencial, que se mide en voltios o en milivoltios
El potencial de membrana de reposo se origina a partir de tres factores principales
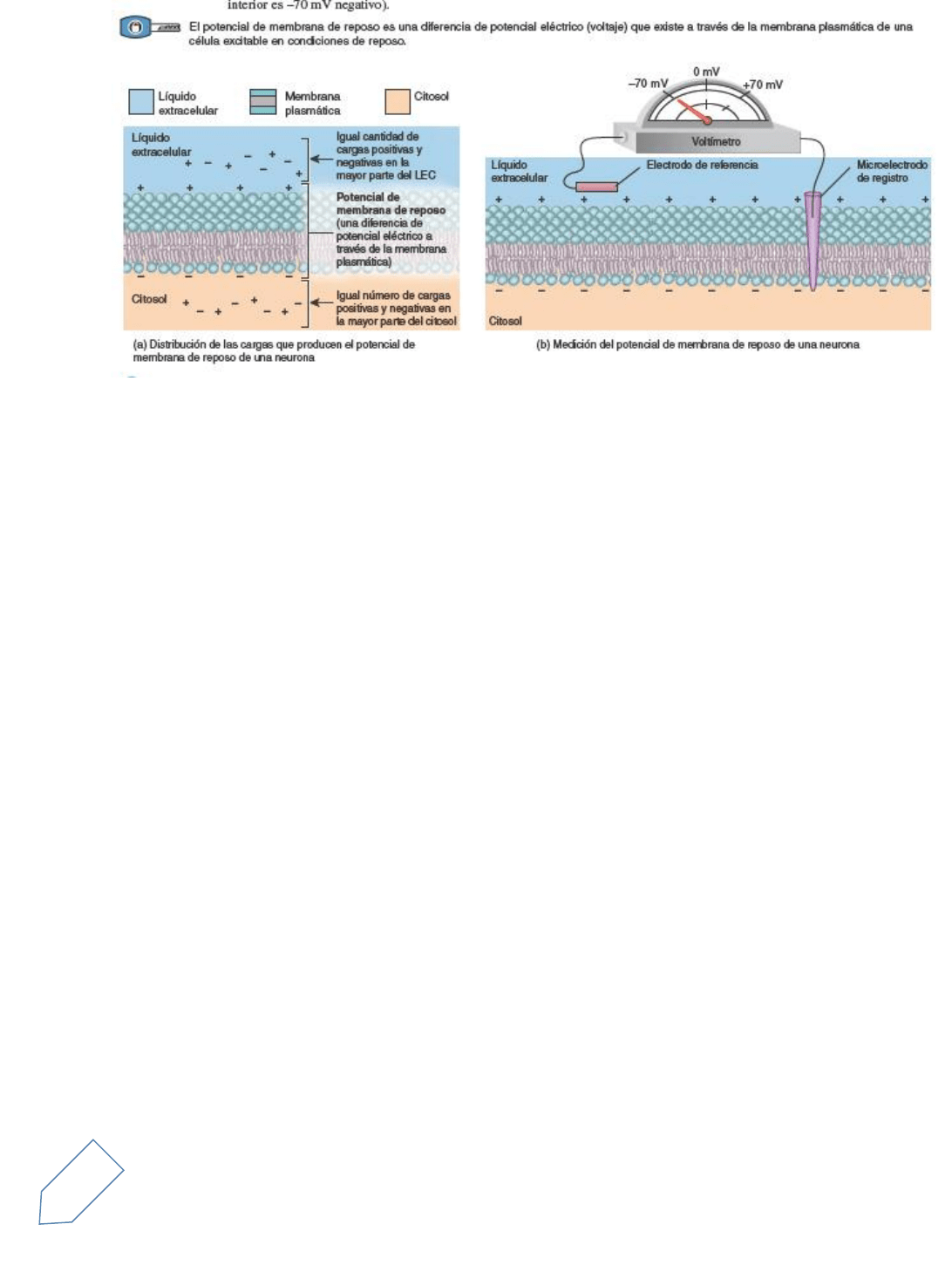
1. La distribución desigual de diversos iones en el líquido extracelular y en el citosol.
El líquido extracelular es rico en iones de Na+ y de cloruro (Cl–). En el citosol, sin embargo, el catión
principal es el K+, y los dos aniones predominantes son los fosfatos que se unen a distintas moléculas,
como los tres fosfatos en el ATP y los aminoácidos en las proteínas. Dado que la membrana
plasmática típicamente tiene más canales pasivos a los iones K+ que canales pasivos a los iones Na+,
la cantidad de iones K+ que difunde a favor de su gradiente de concentración hacia afuera de la célula
y adentro del líquido extracelular es mayor que la cantidad de iones Na+ que difunden a favor de su
gradiente de concentración desde el líquido extracelular hacia la célula. En la medida en que cada
vez más iones positivos de potasio abandonan la célula, el interior de la membrana se vuelve más
negativo y el exterior, más positivo
2. Incapacidad de la mayoría de los aniones para abandonar la célula. Otro factor contribuye a la
negativización del interior: la mayoría de los iones con carga negativa que se encuentra dentro de la
célula no puede abandonarla (Figura 12.13). No pueden seguir al K+ hacia el exterior de la célula, ya
que están adheridos a moléculas no difusibles, como ATP y proteínas grandes.
5
3. La naturaleza electrogénica de las ATPasas Na+/K+
La pequeña cantidad de iones Na+ que ingresan y de iones K+ que salen en forma pasiva de la célula
es compensada por las ATPasas Na+/K+ (bombas de sodio-potasio)
Estas bombas ayudan a mantener el potencial de membrana en reposo bombeando el Na+ hacia el
exterior de la célula a medida que éste ingresa pasivamente
Recuerde que las ATPasas Na+/K+ expulsan tres iones Na+ por cada dos iones K+ importados (véase
la Figura 3.10). Dado que estas bombas extraen más cargas positivas de la célula de las que llevan
hacia el interior, se dice que son electrogénicas, lo que significa que contribuyen a la negativización
del potencial de membrana en reposo
Potenciales graduados
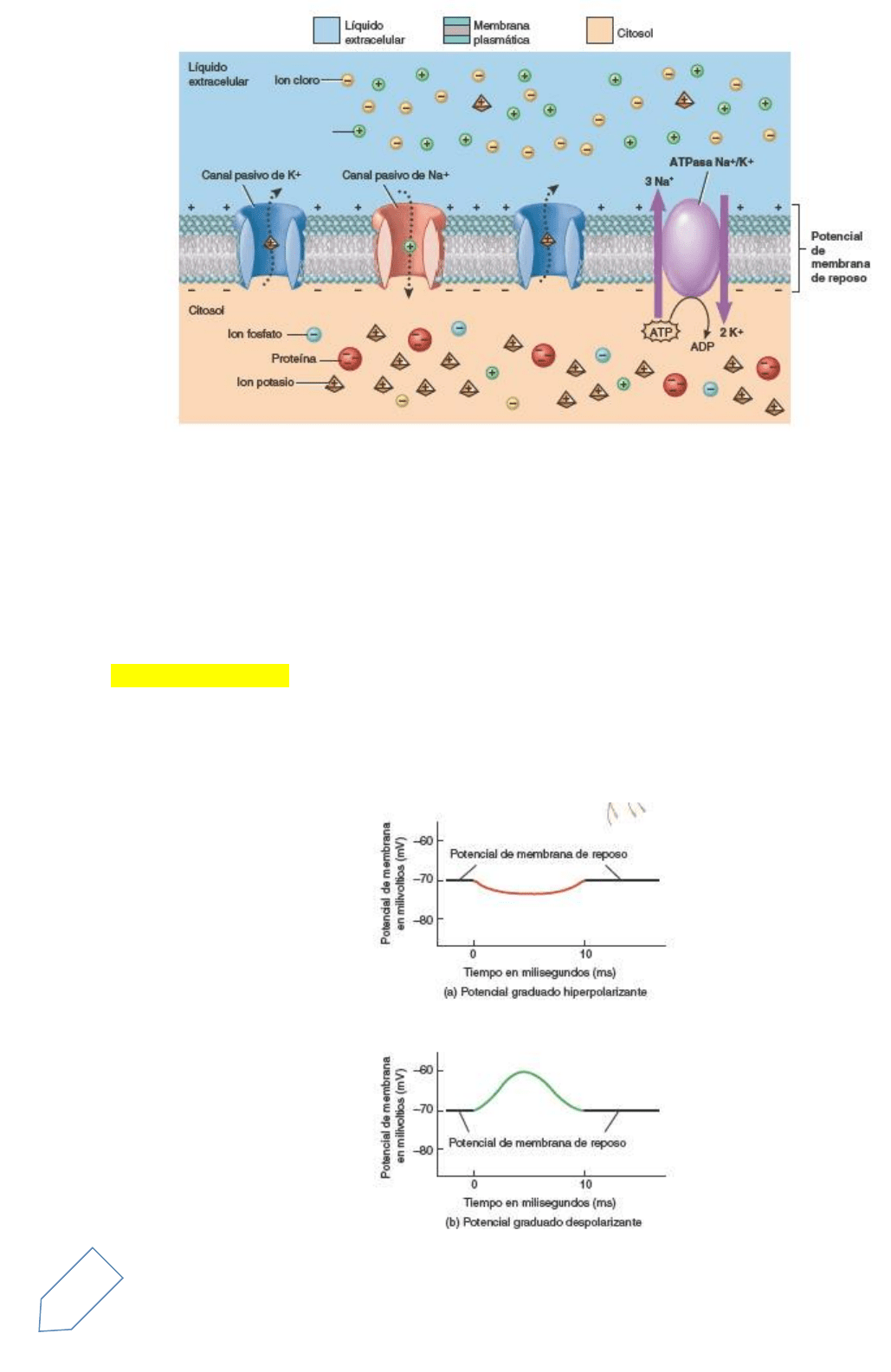
Un potencial graduado es una pequeña desviación del potencial de membrana que hace que ésta se halle
más polarizada (con el interior más negativo) o bien menos polarizada (con el interior menos negativo).
Cuando la respuesta polariza aún más la membrana (y vuelve el interior más negativo), se denomina potencial
graduado hiperpolarizante. Cuando la respuesta torna a la membrana menos polarizada (hace el interior
menos negativo), se denomina potencial graduado despolarizante

6
Un potencial graduado se produce cuando un estímulo hace que los canales activados mecánicamente o los
canales dependientes de ligan-do se abran o cierren en la membrana plasmática de una célula excitable
Los potenciales graduados se producen fundamentalmente en las dendritas y en el cuerpo celular de una
neurona. Decir que estas señales eléctricas están graduadas significa que varían en amplitud (tamaño) de
acuerdo con la intensidad del estímulo.
La apertura o cierre de los canales iónicos altera el flujo de iones específicos a través de la membrana y
produce un flujo de corriente localizado; esto significa que se propaga hacia regiones adyacentes a lo largo
de la membrana plasmática en cualquier dirección desde el origen del estímulo, por una distancia corta en la
membrana plasmática y luego desaparece gradualmente a medida que se pierden las cargas a través de la
membrana, mediante los canales pasivos. Este modo de viajar, por medio del cual el potencial graduado
desaparece a medida que se propaga a lo largo de la membrana, se conoce como conducción decremental.
Como desaparecen a los pocos milímetros de su punto de origen, los potenciales graduados son útiles
solamente para la comunicación a corta distancia.
Aunque un potencial graduado individual sufre conducción decremental, puede tornarse más fuerte y durar
más por la sumación con otros potenciales graduados. Sumación es el proceso por medio del cual se suman
los potenciales graduados. Si dos potenciales graduados despolarizantes se suman, el resultado neto es un
potencial graduado despolarizante más grande el tipo de estímulo que los origina y el lugar donde se
producen. Por ejemplo, cuando un potencial graduado se genera en las dendritas o el cuerpo celular de una
neurona en respuesta a un neurotransmisor, se denomina potencial postsináptico (tratado más adelante).
Por su parte, los potenciales graduados que se originan en las neuronas y en los receptores sensitivos se
denominan potenciales generadores y potenciales receptores.
Generación de los potenciales de acción
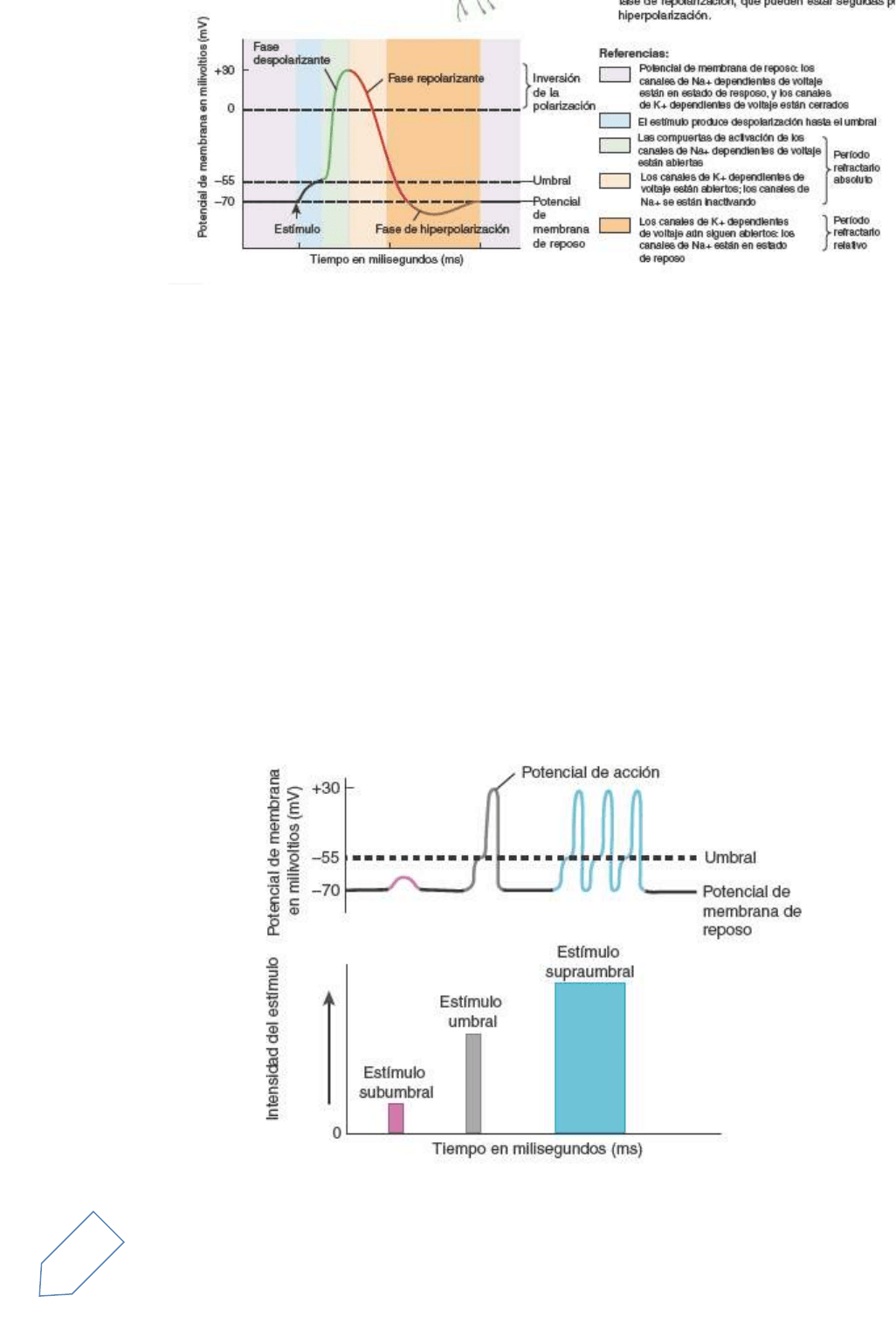
Un potencial de acción (PA) o impulso nervioso consiste en una secuencia de procesos que se suceden con
rapidez y disminuyen o revierten el potencial de membrana y que, finalmente, lo restablecen al estado de
7
reposo. Un potencial de acción tiene dos fases principales: una fase despolarizante y una fase de
repolarización
Durante la fase de despolarización, el potencial de membrana negativo se vuelve menos negativo, llega a
cero y luego se vuelve positivo. Durante la fase de repolarización, el potencial de membrana retorna a su
estado de reposo de –70 mV. Después de la fase de repolarización puede haber una fase de
poshiperpolarización, durante la cual el potencial de membrana se torna transitoriamente más negativo que
el nivel de reposo. Durante un potencial de acción, se abren y luego se cierran dos tipos de canales
dependientes del voltaje.
Estos canales están presentes, fundamentalmente, en la membrana plasmática del axón y en los axones
terminales. El primer canal que se abre, el canal de Na+, permite el ingreso rápido del Na+ hacia el interior
de la célula, lo que produce la fase de despolarización. Luego se abren los canales de K+ y permiten el flujo
hacia afuera del K+, que genera la fase de repolarización. La fase de poshiperpolarización se produce cuando
los canales de K+ dependiente del voltaje se mantienen abiertos, una vez terminada la fase de repolarización.
Un potencial de acción se produce en la membrana del axón de una neurona cuando la despolarización
alcanza cierto nivel denominado umbral (aproximadamente –55 mV en muchas neuronas). Diferentes
neuronas pueden tener distintos umbrales para la generación de un potencial de acción, pero el umbral de
una neurona en particular habitualmente es constante. La generación de un potencial de acción depende de
que un estímulo particular sea capaz de llevar el potencial de membrana hasta el umbral (Figura 12.19). Un
potencial de acción no ocurre en respuesta a un estímulo subumbral, un estímulo de despolarización débil,
que no puede llevar el potencial de membrana hasta el umbral. Sin embargo, un potencial de acción se
produce en respuesta a un estímulo umbral, el estímulo que es lo suficientemente intenso como para
despolarizar la membrana hasta el umbral

8
Una vez generado un potencial de acción, su amplitud es siempre la misma y no depende de la intensidad
del estímulo. Un potencial de acción ocurre totalmente o no ocurre en absoluto. Esta característica del
potencial de acción se conoce como principio del todo o nada. Este principio del potencial de acción es similar
a empujar la primera ficha del dominó en una larga hilera de fichas paradas.
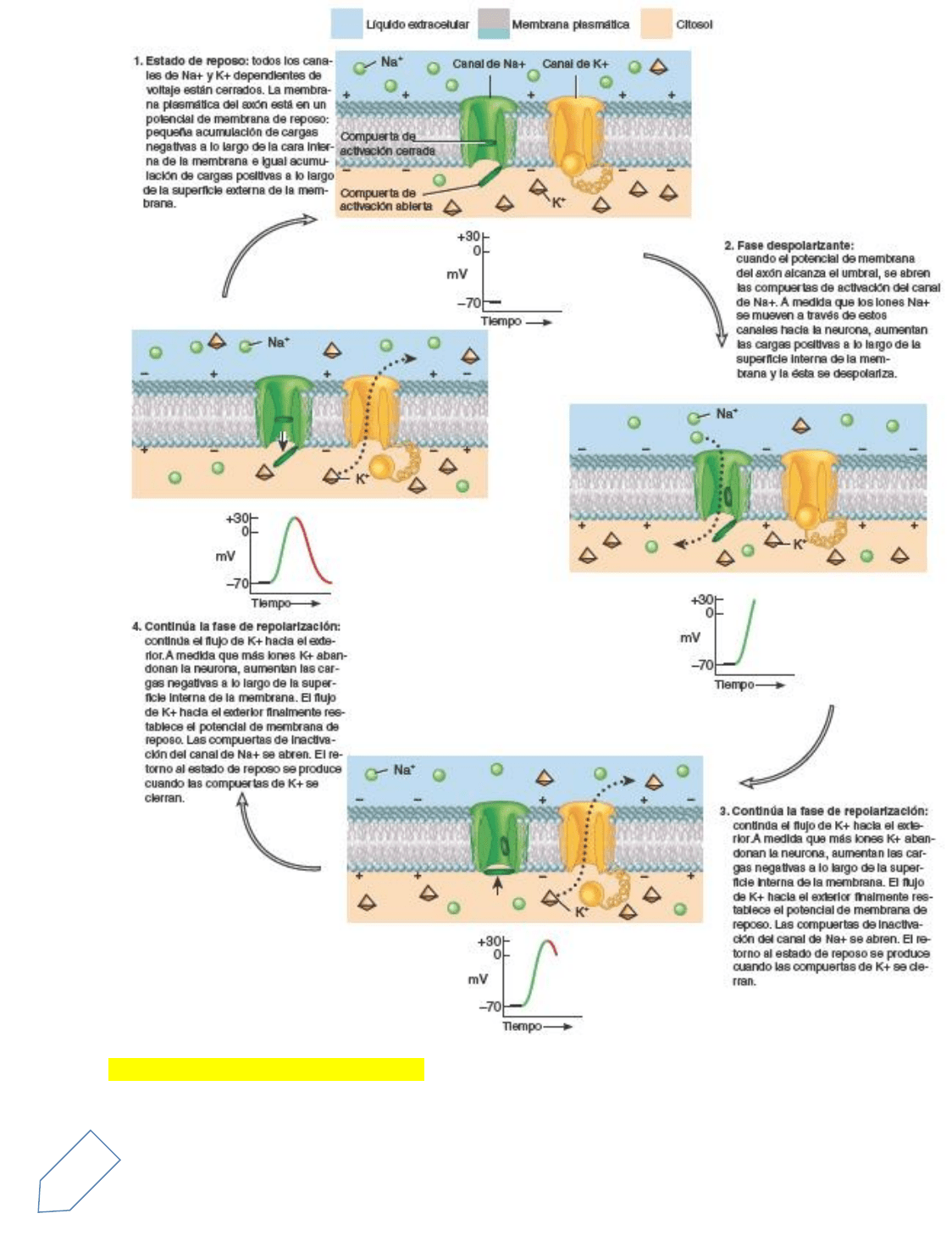
Fase de despolarización
Cuando un potencial graduado despolarizante o algún otro tipo de estímulo produce la despolarización de la
membrana hasta el valor umbral, los canales de Na+ dependientes de voltaje se abren rápidamente. Los
gradientes, tanto eléctrico como químico, actúan a favor del ingreso de Na+, y la entrada consecuente de
Na+ genera la fase despolarizante del potencial de acción. El flujo de entrada del Na+ modifica el potencial
de membrana desde un valor de –55 mV hasta un valor de +30 mV.
En el punto más alto del potencial de acción, la superficie interna de la membrana es 30 mV más positiva que
la superficie externa. Cada canal de Na+ dependiente del voltaje tiene dos compuertas separadas: una
compuerta de activación y una compuerta de inactivación. En el estado de reposo de los canales de Na+
dependientes del voltaje, la compuerta de inactivación está abierta pero la compuerta de activación está
cerrada. Como resultado, el Na+ no puede ingresar a la célula a través de estos canales. Al llegar al umbral,
los canales son activados. En el estado activado de un canal de Na+ dependiente del voltaje, tanto la
compuerta de activación como la de inactivación se hallan abiertas y comienza la corriente de ingreso del
Na+. A medida que se abren más canales, el flujo del ingreso de Na+ aumenta, la membrana se despolariza
aún más, y todavía más canales de Na+ se abren. Éste es un ejemplo de un mecanismo de retroalimentación
positiva. A pesar de ello, la concentración de Na+ se modifica muy poco porque en el líquido extracelular hay
millones de iones de Na+. La bomba de sodio-potasio expele fácilmente los 20 000 iones Na+ que habían
entrado en la célula durante ese único potencial de acción y mantiene así la baja concentración de Na+ en el
interior celular.
Fase de repolarización
Poco tiempo después de que se abren las compuertas de activación de los canales de Na+ dependientes del
voltaje, se cierran las compuertas de inactivación (paso 3 en la Figura 12.20). Ahora el canal se encuentra en
el estado inactivado. Además de la apertura de los canales de Na+ dependientes del voltaje, una
despolarización que alcance el nivel umbral también producirá la apertura de canales de K+ (pasos 3 y 4 en
la Figura 12.20). Dado que los canales de K+ se abren más lentamente, su apertura se produce casi en el
mismo momento en el que los canales de Na+ se están cerrando. La apertura más lenta de los canales de K+
y el cierre de los canales de Na+ previamente abiertos produce la fase de repolarización del potencial de
acción. A medida que los canales de Na+ se inactivan, el ingreso de Na+ disminuye. Al mismo tiempo, los
canales de K+ se están abriendo y se acelera de este modo la salida del K+. El ingreso más lento del Na+ y la
aceleración del flujo de egreso de K+ causan una variación en el potencial de membrana que pasa de un valor
de +30 mV a –70 mV. La repolarización también permite que los canales de Na+ inactivos puedan volver al
estado de reposo.
Fase de poshiperpolarización
Mientras que los canales de K+ permanecen abiertos, el egreso del K+ puede ser lo suficientemente
importante como para causar una fase de poshiperpolarizacion del potencial de acción
Durante esta fase, los canales de K+ dependientes del voltaje se mantienen abiertos y el potencial de
membrana se torna incluso más negativo (unos –90 mV). A medida que los canales de K+ dependientes del
voltaje se cierran, el potencial de membrana retorna a los valores de reposo de –70 mV. A diferencia de los
canales de Na+ dependientes del voltaje, la mayoría de los canales de K+ no presentan un estado inactivo.
En lugar de ello, alternan entre los estados cerrado (en reposo) y abierto (activo).
Período refractario
El tiempo luego del inicio de un potencial de acción durante el cual una célula excitable no puede generar
otro potencial de acción se denomina período refractario (véase referencias de Figura 12.18).
En el período refractario absoluto ni siquiera un estímulo muy intenso podrá iniciar un segundo potencial de
acción. Este lapso coincide con el período de activación e inactivación de los canales de Na+
El período refractario relativo es el intervalo de tiempo durante el cual un segundo potencial de acción puede
ser iniciado, pero sólo por un estímulo más potente que lo normal. Coincide con el período en el cual los
9
canales de K+ están todavía abiertos, después de que los canales de Na+ inactivos ya han vuelto a su estado
de reposo.
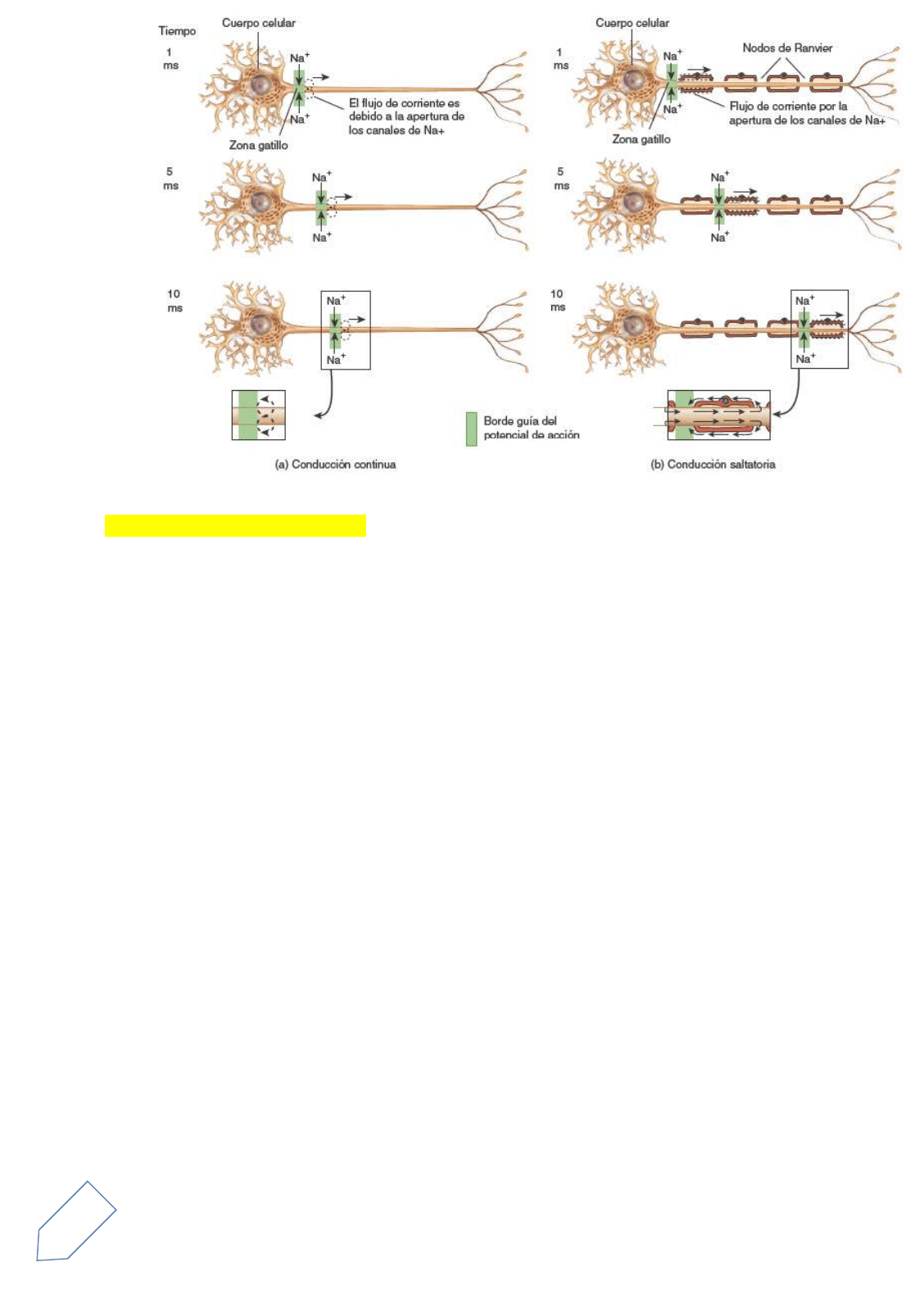
Propagación de los potenciales de acción
Para transmitir información entre distintos sectores del cuerpo, los potenciales de acción de una neurona
deben trasladarse desde la zona gatillo, donde se originan, hasta los axones terminales. Al contrario del
potencial graduado, un potencial de acción no es decremental, no desaparece.

10
En cambio, mantiene su intensidad a medida que se propaga a lo largo de la membrana. Esta forma de
conducción se denomina propagación y depende de la retroalimentación positiva.
En realidad, no es el mismo potencial de acción que se propaga a lo largo de todo el axón. En cambio, se
regenera una y otra vez en regiones adyacentes de la membrana, a partir de la zona gatillo hasta los axones
terminales. Como los potenciales de acción pueden viajar a lo largo de una membrana sin desaparecer,
funcionan en comunicación sobre largas distancias.
Conducción continua y conducción saltatoria
El tipo de propagación de los impulsos que se ha descrito se denomina conducción continua e involucra
despolarización y repolarización paso por paso de cada segmento adyacente de la membrana plasmática
La conducción saltatoria, un tipo especial de propagación de los impulsos que tiene lugar en los axones
mielínicos, se produce por la distribución desigual de canales dependientes del voltaje.
Algunos pocos de estos canales se localizan en la región del axolema cubierta por la vaina de mielina. En
contraste, en los nodos de Ranvier (donde no hay vaina de mielina), el axolema tiene muchos canales
dependientes del voltaje. Por lo tanto, la corriente producida por el Na+ y el K+ fluye a través de la membrana
principalmente en esos nodos.
El potencial de acción del primer nodo origina corrientes iónicas en el citosol y en el líquido extracelular;
dichas corrientes despolarizan la membrana hasta el umbral y provocan la apertura de los canales de Na+
dependientes del voltaje del segundo nodo. La corriente iónica que se establece a través de estos canales
abiertos produce un impulso nervioso en el segundo nodo. Luego, el impulso nervioso genera en el nodo una
corriente iónica que da lugar a la apertura de los canales de Na+ dependientes del voltaje del tercer nodo, y
así sucesivamente. Cada nodo se repolariza después de despolarizarse.
El flujo de corriente a través de la membrana que se produce sólo en los nodos de Ranvier tiene dos
consecuencias:
1. Como un potencial de acción salta a lo largo de los extensos segmentos de axolema rodeados de
mielina, a medida que la corriente fluye de un nodo al siguiente, se desplaza a mayor velocidad que
en un axón amielínico del mismo diámetro.
2. La apertura de un número menor de canales y sólo a nivel de los nodos, en vez de producirse la
apertura de muchos canales en cada segmento adyacente de la membrana, representa un modo de
conducción más eficiente en cuanto al gasto de energía
Como sólo se despolarizan y repolarizan pequeñas regiones de la membrana cada vez que un impulso
nervioso pasa por esa región, hay un ingreso mínimo de Na+ y una salida también mínima de K+. De
esa forma, se consume menos ATP en las bombas de sodio y potasio para mantener la baja
concentración intracelular de Na+ y la baja concentración extracelular de K+.
Factores que afectan la velocidad de propagación
1. Grado de desmielinización: se propagan más rápidamente a lo largo de los axones mielínicos que de
los amielínicos.
2. Diámetro del axón: Los axones de mayor diámetro propagan los potenciales de acción más
rápidamente que los más pequeños, debido a sus áreas de superficie mayores.
3. Temperatura: Los axones propagan los potenciales de acción a menor velocidad cuando se enfrían.
11
Clasificación de las fibras nerviosas
Los axones pueden ser clasificados en 3 grupos principales sobre la base del grado de mielinización, sus
diámetros y sus velocidades de propagación:
• Las fibras A son los axones de mayor diámetro (5-20 μm), que son mielínicos. Las fibras tipo A tienen un
corto período refractario absoluto y conducen los impulsos nerviosos (potenciales de acción) a velocidades
de 12 a 130 m/s. Los axones de las neuronas sensitivas que propagan los impulsos relacionados con el tacto,
la presión, la posición de las articulaciones y algunas sensaciones térmicas, como también los axones de las
neuronas motoras que conducen impulsos hacia los músculos esqueléticos son fibras de tipo A.
• Las fibras B son axones cuyos diámetros oscilan entre 2 y 3 μm.
Como las fibras A, las fibras B son mielínicas y presentan conducción saltatoria con velocidades hasta de 15
m/s. Las fibras B tienen un período refractario absoluto algo más largo que las A. Conducen impulsos
nerviosos sensitivos desde las vísceras hasta el encéfalo y la médula espinal. Además, constituyen todos los
axones de las neuronas motoras autonómicas que se extienden desde el encéfalo y la médula hasta las
estaciones de relevo del SNA llamadas ganglios autónomos.
• Las fibras C son las de diámetro más pequeño (0,5 a 1,5 μm) y todas carecen de vaina de mielina. La
velocidad de propagación de los impulsos nerviosos a lo largo de una fibra C es de 0,5 a 2 m/s.
Las fibras C presentan los períodos refractarios absolutos más largos. Estos axones amielínicos conducen
algunos impulsos sensitivos relacionados con el dolor, tacto, presión, calor y frío desde la piel, e impulsos
dolorosos desde las vísceras. Las fibras motoras autónomas que se extienden desde los ganglios autónomos
para estimular el corazón, el músculo liso y las glándulas son también fibras C. Entre los ejemplos de funciones
motoras de las fibras B y C, encontramos la miosis y la midriasis pupilares, el aumento o disminución de la
frecuencia cardíaca y la contracción o relajación de la vejiga.

12
TRANSMISIÓN DE SEÑALES EN LAS SINAPSIS
una sinapsis es una región en la que se produce la comunicación entre dos neuronas o entre una neurona y
una célula efectora (célula muscular o célula glandular). El término neurona presináptica (pre-, antes) se
refiere a una célula nerviosa que transporta el impulso nervioso hacia la sinapsis. Una célula postsináptica es
la célula que recibe una señal. Puede ser una célula nerviosa denominada neurona postsináptica (post-,
después) que transmite un impulso nervioso lejos de la sinapsis o una célula efectora que responde al impulso
en la sinapsis.
La mayoría de las sinapsis pueden clasificarse en axodendríticas (entre un axón y una dendrita), mientras que
otras son axosomáticas (entre un axón y el cuerpo celular o soma) o axoaxónicas (entre axones). Además, las
sinapsis pueden ser eléctricas o químicas y difieren tanto estructural como funcionalmente.
Las sinapsis son esenciales para la homeostasis, ya que permiten que la información pueda ser filtrada e
integrada.
Sinapsis eléctricas
En una sinapsis eléctrica, los potenciales de acción (impulsos) se transmiten directamente entre las
membranas plasmáticas de células adyacentes, a través de estructuras llamadas uniones comunicantes o en
hendidura. Cada unión en hendidura contiene alrededor de 100 conexones tubulares, que actúan como
conductos para conectar directamente el citosol de las dos células
A medida que los iones fluyen de una célula a la siguiente a través de los conexones, el potencial de acción
se propaga de célula en célula. Las uniones en hendidura son frecuentes en el músculo liso visceral, el
músculo cardíaco y el embrión en desarrollo. También se encuentran presentes en el encéfalo.
Las sinapsis eléctricas tienen dos ventajas principales:
1. Comunicación más rápida: Como los potenciales de acción se transmiten directamente a través de las
uniones en hendidura, las sinapsis eléctricas son más rápidas que las sinapsis químicas. En una sinapsis
eléctrica, el potencial de acción pasa directamente de la célula presináptica a la célula postsináptica.
Cuando un potencial postsináptico despolarizante alcanza el umbral, desencadenará un potencial de acción
en el axón de la neurona postsináptica.
2. Sincronización. Las sinapsis eléctricas pueden sincronizar (coordinar) la actividad de un grupo de neuronas
o de fibras musculares y así pueden producir potenciales de acción al unísono (sincitio).
El valor que tienen estos potenciales de acción sincronizados en el corazón o en el músculo liso visceral se ve
reflejado en la contracción coordinada de estas fibras para producir un latido cardíaco o para facilitar la
progresión del alimento a lo largo del tubo digestivo.
13
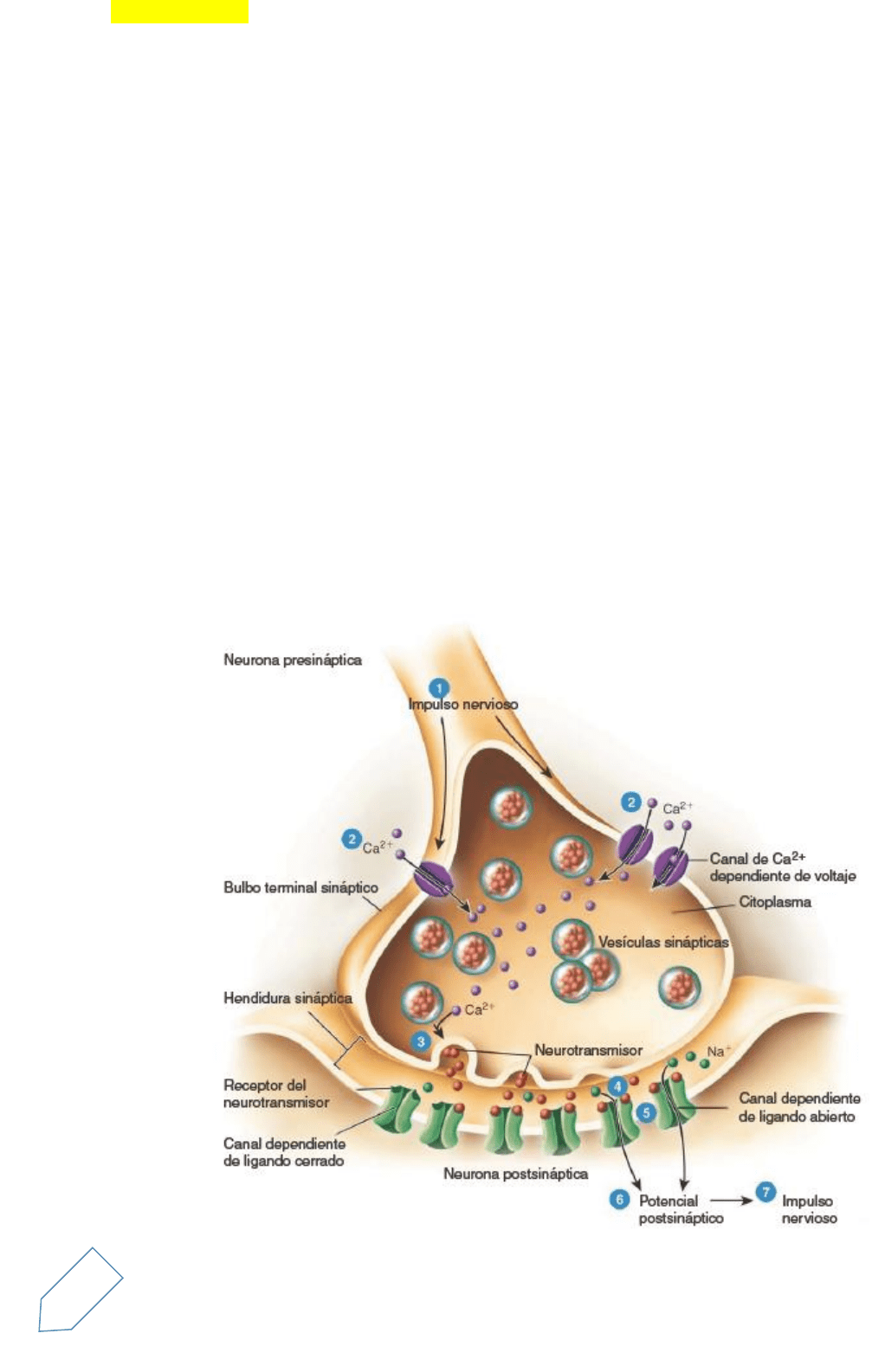
Sinapsis químicas
A pesar de la cercanía entre las membranas plasmáticas de las neuronas presinápticas y postsinápticas en
una sinapsis química, ambas no se tocan. Están separadas por la hendidura sináptica, un espacio de 20 a 50
nm* lleno de líquido intersticial.
En respuesta a un impulso nervioso, la neurona presináptica libera un neurotransmisor que se difunde a
través del líquido de la hendidura sináptica y se une a receptores específicos en la membrana plasmática de
la neurona postsináptica. La neurona postsináptica recibe la señal química y, como resultado, produce un
potencial postsináptico, un tipo de potencial graduado. De esta forma, la neurona presináptica convierte una
señal eléctrica (el impulso nervioso) en una señal química (el neurotransmisor liberado). La neurona
postsináptica recibe esta señal química y, en respuesta, genera una señal eléctrica (el potencial
postsináptico). El tiempo que se requiere para llevar a cabo estos procesos en una sinapsis química, el retardo
sináptico de alrededor de 0,5 ms
Una sinapsis química típica genera la transmisión de una señal de la siguiente manera (Figura 12.22):
1. Un impulso nervioso arriba al bulbo terminal sináptico (o a una varicosidad) de un axón presináptico.
2. La fase de despolarización del impulso nervioso abre los canales de Ca2+dependientes del voltaje
que están en la membrana plasmática de los bulbos sinápticos. Dado que la concentración de iones
de calcio es mayor en el líquido extracelular, el Ca2+fluye hacia el interior de la célula a través de los
canales abiertos.
3. El aumento en la concentración de Ca2+dentro de la neurona presináptica actúa como una señal que
desencadena la exocitosis de las vesículas sinápticas. A medida que la membrana de las vesículas se
fusiona con la membrana plasmática, las moléculas de neurotransmisores contenidos dentro de
estas vesículas se liberan hacia la hendidura sináptica. Cada vesícula sináptica contiene varios miles
de moléculas de neurotransmisor.
4. Las moléculas del neurotransmisor difunden a través de la hendidura sináptica y se unen a los
receptores de los neurotransmisores localizados en la membrana plasmática de la neurona
postsináptica.

14
5. La unión de las moléculas de neurotransmisor con sus receptores en los canales dependientes del
ligando provoca la apertura de éstos y permite el flujo de determinados iones a través de la
membrana.
6. A medida que los iones fluyen a través de los canales abiertos, se producen cambios en el voltaje de
la membrana. Este cambio en el voltaje constituye un potencial postsináptico. Según el tipo de iones
que permita pasar el canal, el potencial postsináptico puede ser despolarizante (excitación) o
hiperpolarizante (inhibición). Por ejemplo, la apertura de los canales de Na+ hace posible el ingreso
de este ion, que produce la despolarización. Sin embargo, la apertura de los canales de Cl– o de K+
genera hiperpolarización. La apertura de los canales de Cl– permite el ingreso de este ion en la célula,
mientras que la apertura de los canales de K+ da lugar a su salida; en cualquiera de los dos casos, el
interior de la célula se vuelve más negativo.
7. Cuando un potencial postsináptico despolarizante alcanza el umbral, desencadenará un potencial de
acción en el axón de la neurona postsináptica.
En la mayoría de las sinapsis químicas, la transferencia de información
es unidireccional, desde una neurona presináptica hacia una neurona postsináptica o hacia un efector, como
una fibra muscular o una célula glandular. Por ejemplo, la transmisión sináptica en una sinapsis o unión
neuromuscular se dirige de una neurona motora somática a una fibra muscular esquelética (pero nunca en
la dirección opuesta). Solamente los bulbos terminales sinápticos de las neuronas presinápticas pueden
liberar neurotransmisores, y sólo en la membrana de la neurona postsináptica se hallan las proteínas
receptoras que pueden reconocer el neurotransmisor y unirse a éste. En consecuencia, los potenciales de
acción se propagan sólo en una dirección.
Potenciales postsinápticos excitatorios e inhibitorios
Un neurotransmisor puede producir un potencial graduado excitatorio o inhibitorio. Un neurotransmisor que
despolarice la membrana postsináptica es excitatorio porque el valor del potencial de membrana se acerca
al valor umbral. Un potencial postsináptico despolarizante se denomina, en consecuencia, potencial
postsináptico excitatorio (PPSE). A pesar de que un único PPSE normalmente no inicia un impulso nervioso,
la célula postsináptica se vuelve más excitable. Como está parcialmente despolarizada, es más probable que
pueda alcanzar el umbral cuando se produzca el siguiente PPSE.
El neurotransmisor que produce hiperpolarización de la membrana postsináptica es inhibitorio. Durante la
hiperpolarización, la generación de un impulso nervioso se vuelve más difícil que lo normal, debido a que el
potencial de membrana se torna más negativo y, de esta forma, se aleja aún más del umbral que en el estado
de reposo. El potencial postsináptico hiperpolarizante se denomina potencial postsináptico inhibitorio (PPSI).
REVISIÓN DEL CAPÍTULO 12.1
Generalidades del sistema nervioso
1. El sistema nervioso central (SNC) está constituido por el encéfalo y la médula espinal.
2. El sistema nervioso periférico (SNP) está constituido por todo el tejido nervioso que se halla por fuera del
SNC.
Los componentes del SNP incluyen nervios, ganglios, plexos entéricos y receptores sensoriales.
3. Los componentes del SNP son el sistema nervioso somático (SNS), el sistema nervioso autónomo (SNA) y
el sistema nervioso entérico (SNE).
4. El SNS está constituido por neuronas que conducen impulsos desde receptores somáticos y receptores
para los sentidos especiales hacia el SNC y por neuronas motoras que conducen impulsos desde el SNC hacia
los músculos esqueléticos.
5. El SNA contiene neuronas sensoriales localizadas en los órganos viscerales y neuronas motoras que
transportan impulsos desde el SNC hacia el tejido muscular liso, el tejido muscular cardíaco y las glándulas.
6. El SNE está formado por neuronas de los plexos entéricos del tubo digestivo que funcionan, de alguna
manera, en forma independiente del SNA y del SNC. El SNE monitoriza los cambios químicos en el tubo
digestivo y el estiramiento de sus paredes; el SNE también controla la contracción del músculo liso del tubo
digestivo.
7. El sistema nervioso contribuye a mantener la homeostasis e integra todas las actividades del cuerpo por
medio del control de los cambios (función sensitiva), la interpretación de éstos (función integradora) y la
reacción a ellos (función motora).
15
Histología del tejido nervioso
1. El tejido nervioso está compuesto por neuronas (células nerviosas) y la neuroglia. Las neuronas tienen la
propiedad de ser eléctricamente excitables y son responsables de varias funciones exclusivas del sistema
nervioso:
las sensaciones, el pensamiento, los recuerdos, el control de la actividad muscular y la regulación de la
secreción glandular.
2. La mayoría de las neuronas tienen tres componentes. Las dendritas constituyen la principal región de
recepción o de entrada. La integración se produce en el cuerpo celular, donde se alojan los orgánulos típicos.
La región eferente es generalmente un único axón, que propaga los impulsos nerviosos hacia otra neurona,
una fibra muscular o una célula glandular.
3. Las sinapsis son los sitios de contacto funcional entre dos células excitables. Los axones terminales
contienen vesículas sinápticas que están ocupadas por moléculas de neurotransmisores.
4. Los transportes axónicos rápido y lento son sistemas que se encargan del transporte de sustancias entre
el cuerpo celular y los axones terminales.
5. De acuerdo con su estructura, las neuronas se clasifican en multipolares, bipolares y unipolares.
6. Las neuronas se clasifican funcionalmente como neuronas sensitivas (aferentes), neuronas motoras
(eferentes) e interneuronas. Las neuronas sensitivas transportan la información sensitiva hacia el SNC. Las
neuronas motoras transportan información hacia afuera del SNC hacia los efectores (músculos y glándulas).
Las interneuronas se localizan dentro del SNC, entre las neuronas sensitivas y motoras.
7. La neuroglia proporciona sostén, nutrición y protección a las neuronas y mantiene el líquido intersticial
que baña las células nerviosas. La neuroglia del SNC está constituida por los astrocitos, los oligodendrocitos,
la microglia y las células ependimarias. La neuroglia del SNP está formada por células de Schwann y células
satélite.
8. Dos tipos de células gliales producen las vainas de mielina: los oligodendrocitos mielinizan los axones del
SNC, y las células de Schwann mielinizan los axones del SNP.
9. La sustancia blanca está constituida por agregados de axones mielínicos; la sustancia gris contiene cuerpos
celulares, dendritas y axones terminales provenientes de neuronas, axones amielínicos y células gliales.
10. En la médula espinal, la sustancia gris forma el núcleo interno en forma de H, que está rodeado de
sustancia blanca. En el encéfalo, una delgada capa superficial de sustancia gris cubre los hemisferios
cerebrales y cerebelosos.
Señales eléctricas en las neuronas
1. Las neuronas se comunican entre sí por medio de potenciales graduados, que se utilizan sólo para la
comunicación a corta distancia, y por medio de potenciales de acción, que permiten la comunicación a través
de distancias tanto cortas como largas dentro del cuerpo.
2. Las señales eléctricas producidas por las neuronas y por las fibras musculares dependen de cuatro tipos de
cana-les iónicos: los pasivos, los dependientes de ligando, los accionados mecánicamente y los dependientes
del volta-je.
3. Hay un potencial de reposo en la membrana plasmática de las células excitables que no son estimuladas
(en repo-so). Este potencial existe por la pequeña acumulación de iones negativos en el citosol, a lo largo de
la superficie interna de la membrana, y una acumulación igual de iones positivos en el líquido extracelular
(LEC) a lo largo de la superficie externa de la membrana. Un valor típico para el potencial en reposo de la
membrana es –70 mV. Se dice que cualquier célula que presenta un potencial de membrana está polarizada.
El potencial de reposo de la membrana es determinado por tres factores:
1) la distribución desigual de los iones entre el LEC y el citosol,
2) la incapacidad de la mayoría de los aniones citoplasmáticos para abandonar la célula y
3) la naturaleza electrógena de las ATPasas Na+/K+.
4. Un potencial graduado es una pequeña desviación del potencial de membrana de reposo que se produce
a partir de la apertura o cierre de canales dependientes de ligandos o de canales operados mecánicamente.
En la hiperpolarización, un potencial graduado torna más negativo el potencial de membrana (más
polarizado); en la despolarización, un potencial graduado hace menos negativo el potencial de membrana
(menos polarizado). La amplitud de un potencial graduado presenta variaciones que dependen de la
intensidad del estímulo.
Este documento contiene más páginas...
Descargar Completo
Neurobiologia resumen Tortora.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.