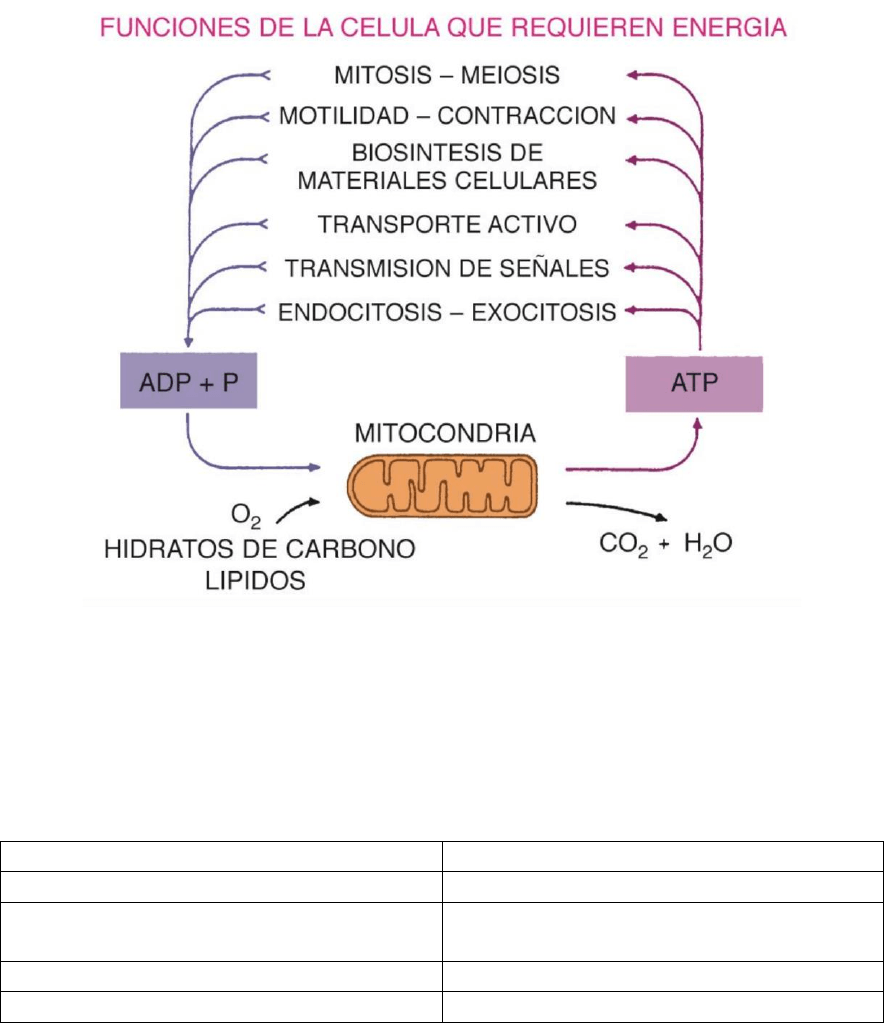
MITOCONDRIAS
Origen de las mitocondrias.
La mayor parte de la energía que usa la célula es provista por el ATP.
Las células necesitan energía para realizar los siguientes trabajos:
Fig. 9-1. Esquema que muestra a la mitocondria como la planta de producción energética de
la célula. El ATP producido se emplea en las funciones indicadas.
La energía es tomada de los alimentos.
Diferencias entre los procesos energéticos de las plantas (fotosíntesis) y de los animales
(fosforilación oxidativa).
FOTOSÍNTESIS
FOSFORILACIÓN OXIDATIVA
En cloroplastos
En mitocondrias
Reacción endergónica:
Energía + CO2 + H2O → Alimentos + H2O
Reacción exergónica:
Alimentos + O2 → Energía + CO2 + H2O
Hidroliza el H2O
Forma H2O
Libera O2
Libera CO2

Sólo en presencia de luz
Independiente de la luz
Periódica
Continua
La energía de las moléculas alimenticias es extraída mediante oxidaciones.
La mayor parte de la energía contenida en las moléculas de los alimentos es extraída
mediante una sucesión de numerosísimas oxidaciones, al cabo de las cuales el oxígeno
atmosférico se une a los átomos de hidrógeno de las citadas moléculas para formar H2O.
Vinculando este último con lo anterior, son tales oxidaciones las que se dan gradualmente,
liberándose en algunas de ellas pequeñas porciones de energía que han de ser captadas por
el ATP. Si las oxidaciones no fueran graduales, la energía química se liberaría súbitamente y
se disiparía como calor.
Deberá recordarse que una molécula se oxida no solamente cuando gana Oxígeno (O) sino
también cuando pierde Hidrógeno (H), debido a que este puede disociarse en un electrón
(e-) y un protón (H+), en un sentido general
toda remoción de e- de cualquier átomo o
molécula constituye una reacción de oxidación. Si el e- removido es aportado por un átomo
de H, el H+ puede permanecer en la molécula (que entonces queda con una carga positiva)
o puede ser removido y pasar al medio acuoso. Ulteriormente los e- y los H+ pueden volverse
a unirse.
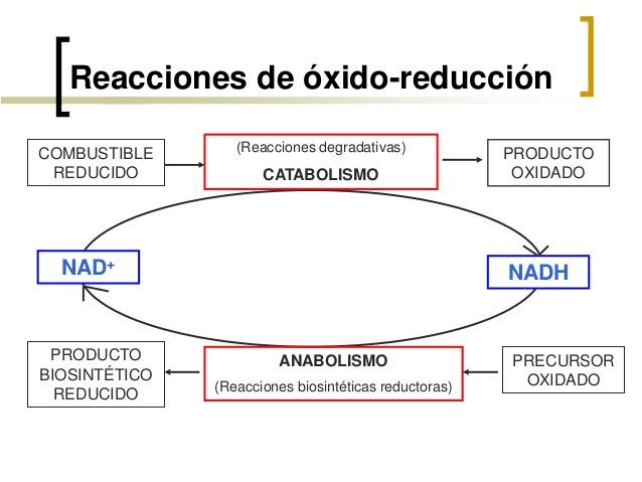
Toda oxidación de un átomo o de una molécula está atada a la reducción de otro átomo u
otra molécula, que entonces ganan Hidrógeno o e-, o pierden Oxígeno.
En el epicentro de algunas reacciones de oxidación y reducción durante el procesamiento de
los alimentos intervienen dos moléculas intermediarias cardinales: el NAD y el FAD. En su
forma oxidativa se representa con las siglas NAD+, y en su forma reducida con las siglas
NADH. La segunda, con las siglas FAD y FADH2, respectivamente.
NAD+ →
Oxidado
Puede aceptar electrones
NADH + H+
Reducido
Aceptó electrones y 1 protón
NADH + H+ →
Reducido
Puede perder electrones y 1 protón
NAD+
Oxidado
Perdió electrones
La degradación enzimática de los alimentos comienza en el aparato digestivo.
La primera etapa en la escisión enzimática de los alimentos tiene lugar en la luz del tubo
digestivo y es entonces extracelular. Así, mediante enzimas producidas por algunas de las
células de dicho tubo, los hidratos de carbono se degradan a monosacáridos -especialmente
glucosa-, los lípidos (en su mayoría triglicéridos) se convierten en ácidos grasos y glicerol, y
las proteínas se degradan a aminoácidos. Luego de ser absorbido por el epitelio intestinal,
estas moléculas ingresan en la sangre, desde donde pasan al interior de las células.
Para asegurarse un abastecimiento continuo de energía, las células guardan en el citosol
parte de la glucosa y de los ácidos grasos bajo la forma de glucógeno y de triglicéridos,
respectivamente. Más aún, existen células especializadas en las que se depositan grandes
reservas de estas moléculas -por ejemplo-, glucógeno en los hepatocitos y triglicéridos en las
células adiposas-, desde las cuales se movilizan cuando es necesario.
La glucólisis tiene lugar en el citosol.
Mediante una serie de reacciones químicas agrupadas bajo el nombre de glucólisis -en las
que intervienen 10 enzimas localizadas en el citosol, cada molécula de glucosa, que posee 6
átomos de carbono, da lugar a 2 moléculas de piruvato, cada una compuesta por 3 carbonos.
Durante este proceso -que constituye la segunda etapa de degradación de los glúcidos- se
invierte la energía de 2 ATP, pero se generan otros 4, de modo que se produce una ganancia
de 2 ATP, uno por cada piruvato.
Más aún, una parte de la energía liberada durante la glucólisis no es transferida al ATP
directamente, sino que promueve la reducción de 2 NAD+, 1 por cada piruvato. Más adelante
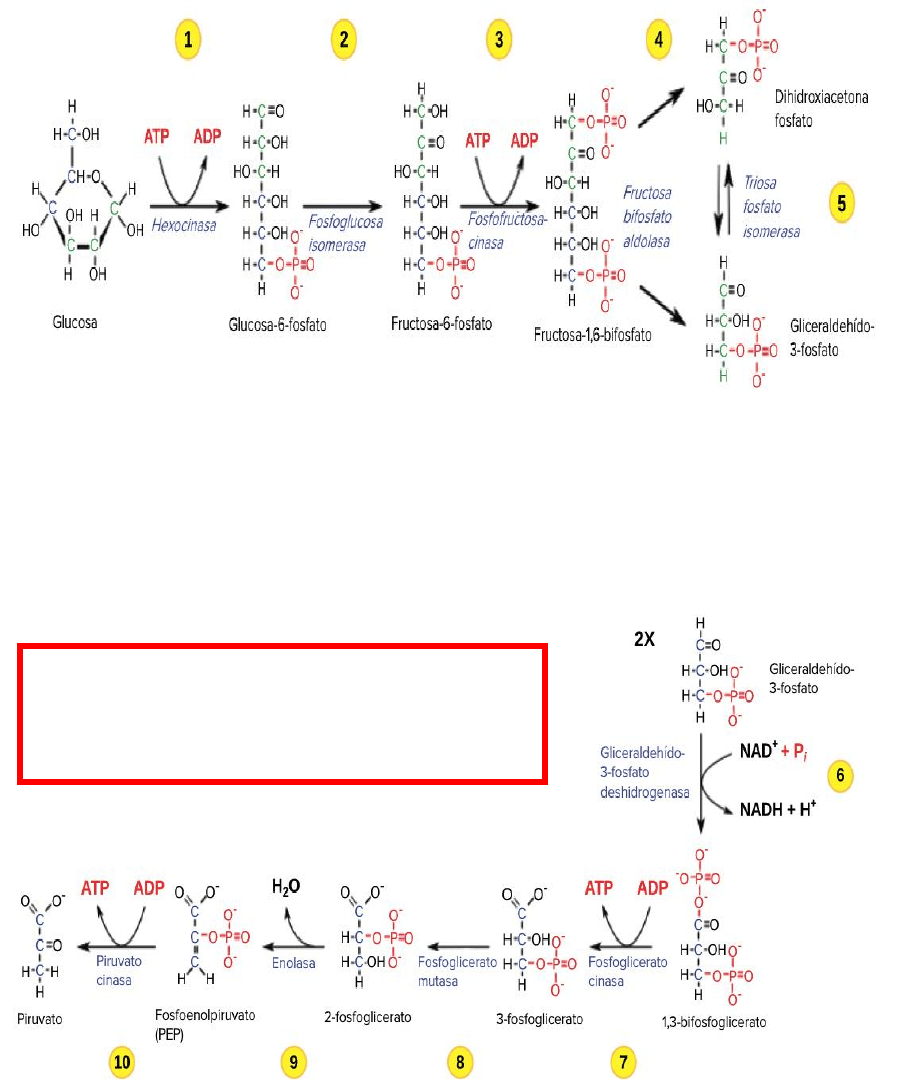
veremos como, en las mitocondrias, la energía contenida en los 2 NADH surgidos de la
glucólisis es transferida al ATP.
Volviendo a los piruvatos, digamos que dejan el citosol e ingresan en las mitocondrias.
Energía: gasto de 2 ATP / molécula de Glucosa.
GLUCÓLISIS: 2ª FASE DE RENDIMIENTO ENERGÉTICO
¿QUÉ OCURRE?
Transformación de Gliceraldehído 3 P en Piruvato. Acidos Carboxilicos (Piruvato o Ácido
Piruvico).
Preguntas:
No se supone que siempre para que ocurra una reacción la energía potencial inicial debe
ser > que la energía potencial final?
Energía: ganancia de 4 ATP y 2 de
NADH + H
+
/ molécula de Glucosa
En las mitocondrias se producen la descarboxilación oxidativa, el ciclo de Krebs y la
fosforilación oxidativa.
Descarboxilación oxidativa
Por acción de un complejo multienzimático llamado piruvato deshidrogenasa, presente en
las mitocondrias, cada piruvato (3 C) se convierte en una molécula de 2 carbonos (2 C). Esta
molécula es un grupo acetilo que se liga a una coenzima -la coenzima A (CoA)-, con la que
compone la acetilcoenzima A (acetil CoA). El carbono es removido del piruvato junto a 2
oxígenos, lo que produce un CO2. El piruvato cede también un ión hidruro (1 H+ y 2 e-). Estas
reacciones reciben el nombre de descarboxilación oxidativa, la tercera etapa en la
degradación de los glúcidos.
Durante la descarboxilación oxidativa se genera energía suficiente para reducir un NAD+, lo
cual se traduce en la formación de una molécula de NADH por cada grupo acetilo producido.
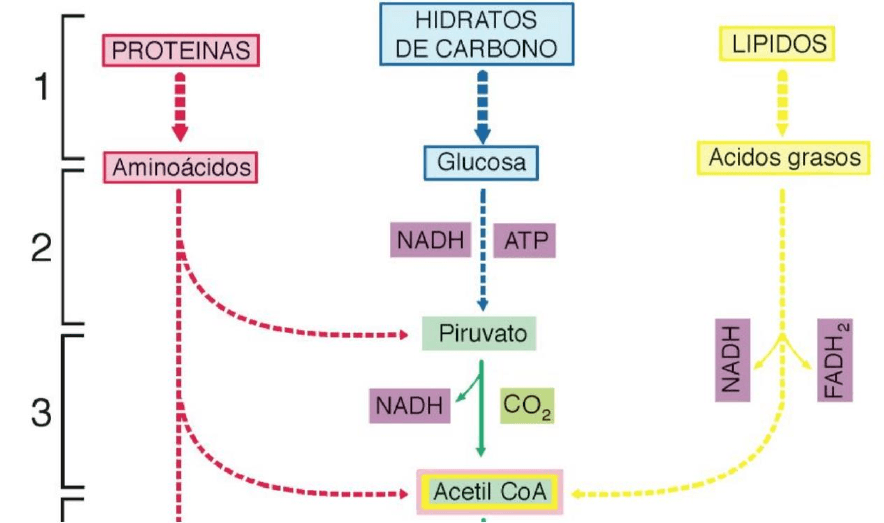
Veamos entonces el proceso completo:
1. Digestión. Las macromoléculas de los alimentos son degradadas en el proceso
digestivo a sus componentes básicos.
2. Glucólisis (citosol). Éstos ingresan a la célula. Si bien la glucosa es el principal
combustible energético, también es posible obtener energía a partir de los ácidos
grasos y, en última instancia, de los aminoácidos. En el citosol, ocurre el proceso de
glucólisis, en el cual la glucosa es transformada a piruvato y esta glucólisis arroja
además como resultado la obtención de 2 ATP y 2 NADH.
3. Descarboxilación oxidativa (matriz mitocondrial). El piruvato ingresa a la mitocondria
y ocurre la descarboxilación oxidativa del mismo que se transforma en acetil CoA,
obteniéndose además, 2 NADH y 2 CO2 (desecho).
Ciclo de Krebs (Oxidación progresiva del PIRUVATO para obtener CO2 y H2O).
A continuación -siempre en las mitocondrias- los átomos de C e H del grupo acetilo
(recordemos que se halla unido a la CoA) son oxidados, de modo que se generan CO2 y H2O.
Las oxidaciones son graduales, y en su transcurso se libera la energía depositada en las
uniones covalentes entre átomos, que pasa al ATP. Ambos procesos -las oxidaciones y la
formación de ATP- ocurren en 2 tiempos; el CO2 se genera en el primero y el H2O en el
segundo.
En el primero de esos tiempos -que representa la cuarta etapa de la degradación de los
glúcidos- abarca una sucesión de oxidaciones en el llamado ciclo de Krebs
De la energía liberada en esta etapa, una pequeña fracción se utilizará generar 1 ATP en
forma directa, pero la mayor parte es aprovechada para reducir 3 NAD+ -que entonces se
convierte en 3 NADH- y 1 FAD, que de estar oxidado pasa a su estado reducido o FADH2.
En el segundo tiempo, contemporáneo al del ciclo de Krebs, los NADH y los FADH2 son
oxidados de manera gradual por una serie de moléculas que se agrupan bajo el nombre de
cadena transportadora de electrones –o cadena respiratoria-, se modo que vuelven a
convertirse en NAD+ y FAD, respectivamente. Cuando ambos dinucleótidos son oxidados se
libera energía depositada en sus moléculas y ésta es transferida al ADP presente en las
mitocondrias, el cual, al fosforilarse, se convierte en ATP. Esta etapa -que es la quinta y última
de la degradación de los glúcidos-, a causa de que da lugar a oxidaciones acopladas a
fosforilaciones, recibe el nombre de fosforilación oxidativa.
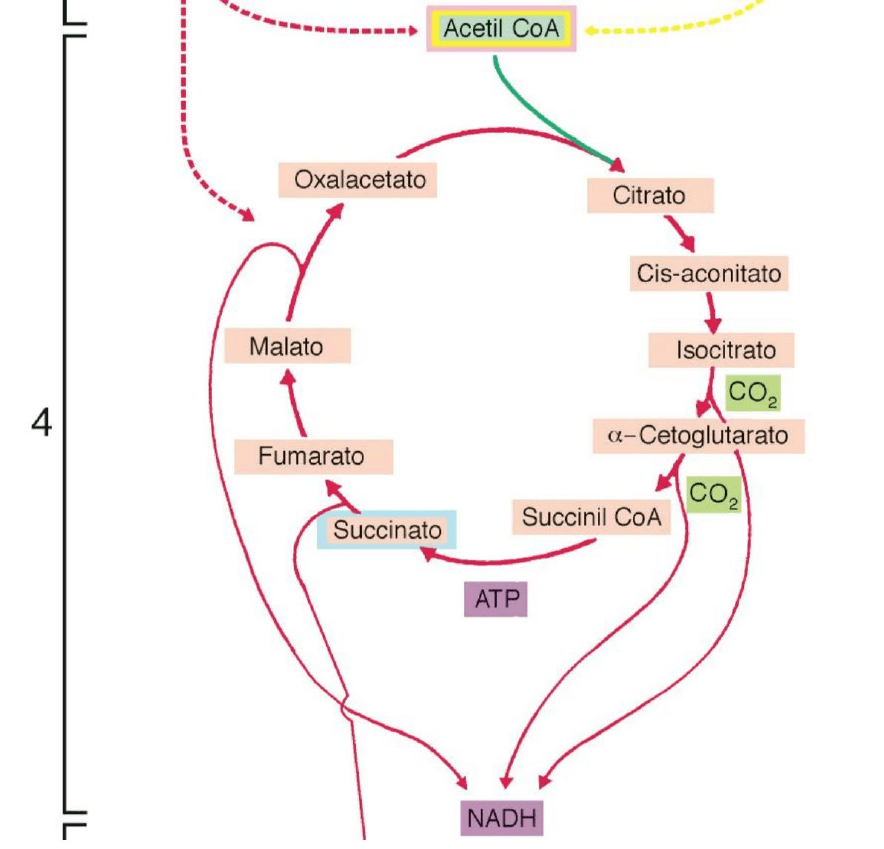
4. Ciclo de Krebs (matriz mitocondrial). Este acetil CoA, siempre dentro de la
mitocondria, ingresa en una cadena cíclica de reacciones químicas llamada ciclo de
Krebs, en el cual se obtiene 2 ATP, 6 NADH, 2 FADH y 4 CO2.
5. Fosforilación oxidativa (membrana mitocondrial interna). Los NADH y los FADH son
oxidados en las enzimas de la cadena respiratoria que al sacarles los H+ les quita
también la energía. Estos H+ son tomados por la ATP sintasa, enzima que les saca la
energía y la utiliza para la síntesis de ATP. Por último los H+ , con electrones de la
cadena respiratoria y O2 , que respiramos, forma H2O.
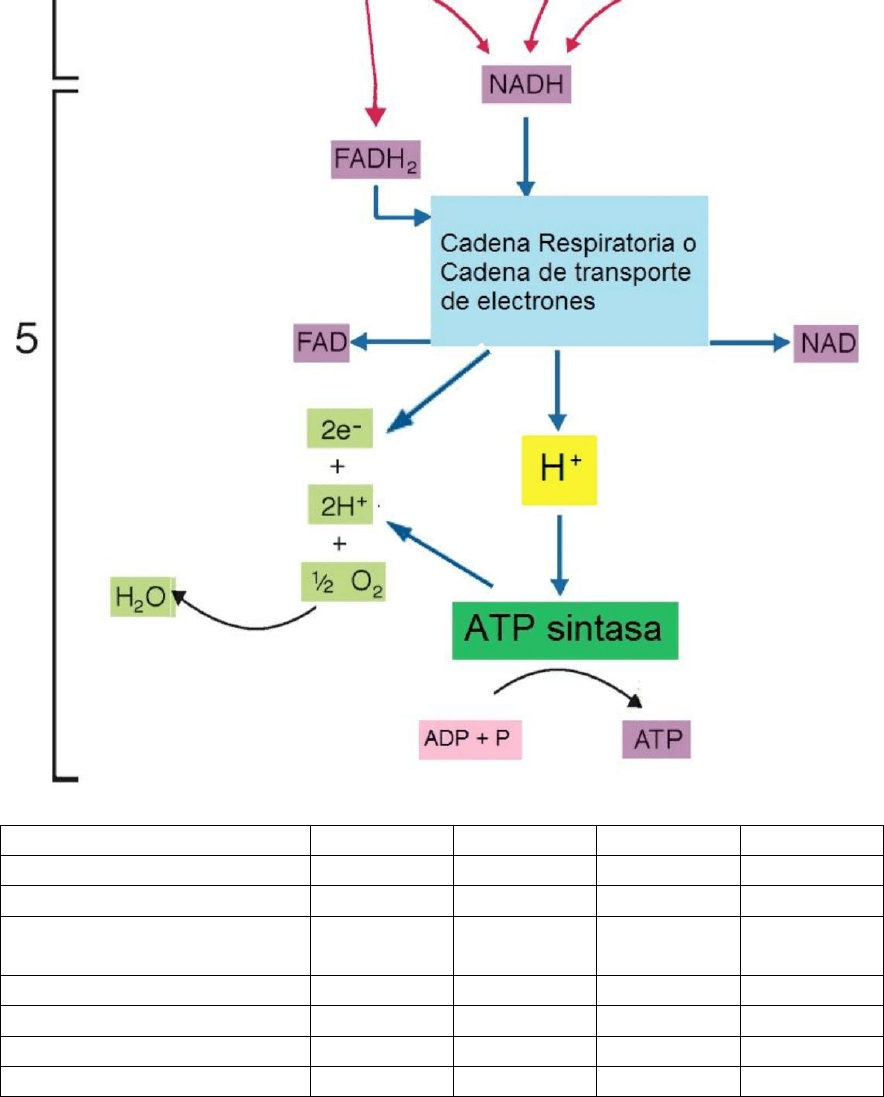
La cadena transportadora de electrones está constituida por un conjunto de
proteínas localizadas en la membrana interna mitocondrial y se encarga de
transportar los electrones que provienen del ciclo de Krebs hacia el oxígeno.
PROCESO
NADH
FADH
CO2
ATP
GLUCÓLISIS (Glucosa)
2
2
DESCARBOXILACIÓN
OXIDATIVA (2 piruvatos)
2
2
CICLO DE KREBS (2 Acetil-CoA)
6
2
4
2
SUB-TOTALES
10
2
4
RENDIMIENTO EN ATPs
X 3 ATP c/u
X 2 ATP c/u
TOTAL DE ATPs (38)
30 ATP
4 ATP
4 ATP

Las mitocondrias se encuentran en todos los tipos celulares.
Se encuentran ubicadas en las regiones de las células donde la demanda de energía es mayor,
por lo que se desplazan de un lado a otro del citoplasma hacia las zonas necesitadas de
energía. Los microtúbulos y sus proteínas asociadas intervienen en tales desplazamientos.
No obstante, en algunos tipos celulares, como los espermatozoides, las células musculares y
las células grasa, las mitocondrias permanecen en lugares fijos.
FUNCIONES DE LAS MITOCONDRIAS
La función principal de las mitocondrias es generar ATP.
La descarboxilación oxidativa se produce en la matriz mitocondrial.
Proveniente del citosol, el piruvato (3 C) ingresa en la matriz mitocondrial, donde por acción
de la piruvato deshidrogenasa pierde un C y se convierte en el grupo acetilo (2 C) de la Acetil
CoA.
Las reacciones del ciclo de Krebs ocurren en la matriz mitocondrial.
Las oxidaciones de la fosforilación oxidativa tienen lugar en la membrana interna de la
mitocondria.
La energía contenida en los NADH y FADH2 -provenientes de la descarboxilación oxidativa y
del ciclo de Krebs- se transfiere al ATP luego de una serie de reacciones químicas que
comienzan con la ionización de ambos dinucleótidos. Luego los átomos de H liberados como
consecuencia de las ionizaciones se disocian en H+ y e-. Ambas reacciones pueden
expresarse mediante las siguientes ecuaciones:
NADH → NAD+ + 2 e- + H+
FADH2
→ FAD + 2 e- + 2 H+
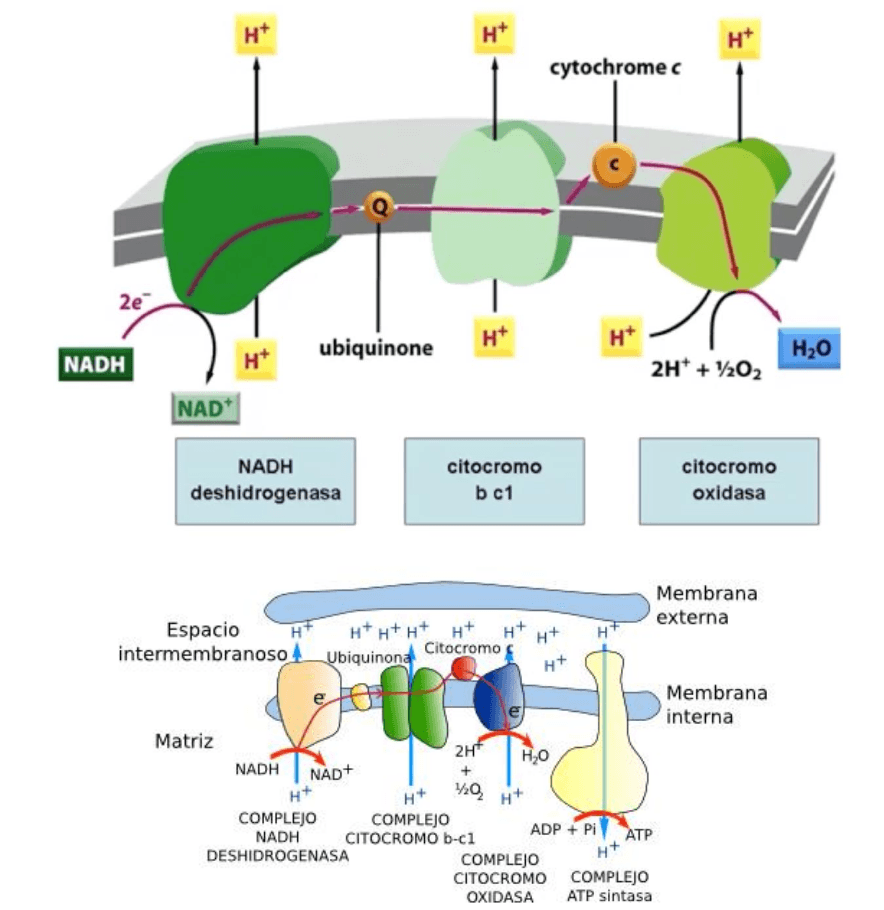
Es importante señalar que los e- surgidos de estos procesos poseen un elevado potencial de
transferencia, es decir, una elevada carga de energía. En esas condiciones ingresan en la
cadena transportadora de electrones.
Dado que cada componente de la cadena posee por los e- una afinidad mayor que su
predecesor, los e- -ricos en energía al ingresar en la cadena- fluyen por ella en el siguiente
orden:
Para los e- cedidos por los átomos de H del NADH, el punto de entrada es la NADH
deshidrogenasa. Desde ésta pasan a la ubiquinona, que los transfiere al complejo b-c1. Los
e- dejan este complejo y pasan al citocromo C, desde el cual pasan al quinto y último eslabón
de la cadena, la citocromo oxidasa. Finalmente, luego de haber gastado la mayor parte de su
energía, los e- pasan a la matriz mitocondrial (membrana externa).
Por su lado, los e- cedidos por los H del FADH2 tienen como punto de entrada la ubiquinona,
a partir de la cuál fluyen por los restantes eslabones de la cadena en el mismo orden en que
lo hacen los e- cedidos por los H del NADH.
Como vemos, en el curso de las reacciones de oxidorreducción que tienen lugar a lo largo de
la cadena respiratoria, el potencial de transferencia de los electrones va disminuyendo, de
modo que en cada etapa pasan a un estado de menor energía, que termina
considerablemente reducida cuando abandonan el último eslabón de la cadena. A
continuación, analizaremos en que se gasta esa energía.
Es utilizada para transferir los H+ disueltos en la matriz mitocondrial al espacio
intermembranoso. Ello requiere energía, ya que los H+ deben ser transportados desde un
medio en el que se hallan menos concentrados a otro en el que su concentración es mayor
(transporte activo). El mecanismo que hace posible el bombeo de los H+ no ha podido ser
determinado. No obstante, se sabe que los H+ salen al espacio intermembranoso a través de
algunas proteínas de los complejos enzimáticos que integran la cadena respiratoria, las
cuales actuarían como verdaderas bombas de H+.
Al existir un gradiente de concentración de H+ entre ambos lados de la membrana
mitocondrial interna, se establece un gradiente de pH (más alto en la matriz mitocondrial) y
se afirma el potencial de membrana, que se hace más positivo en el espacio
intermembranoso. Ambos -el gradiente de pH y el potencial de membrana- componen un
gradiente electroquímico.
La fosforilación del ADP es mediada por la ATP sintetasa.
Los H+ y los e- se combinan con el oxígeno atmosférico para formar agua.
Cabe ahora indagar sobre el destino de los e-, los cuales, luego de perder una parte sustancial
de su energía, han abandonado la cadena respiratoria y regresado a la matriz mitocondrial.
Se combinan tanto con el O2, que procede de la atmósfera como con los H+ provenientes
del espacio intermembranoso, lo que da lugar a la formación de H2O. la atracción de los e-
por el O2, se debe a que poseen una gran afinidad por éste, mayor de la que tienen por la
citocromo oxidasa, lugar por donde salen de la cadena transportadora.
Con la formación de H2O culmina la fosforilación oxidativa; se necesitan 4 e- y 4 H+ por O2
para que se produzcan 2 moléculas acuosas. El H2O es uno de los productos finales del
metabolismo (el otro es el CO2); pasa al citosol, desde dónde sale al espacio extracelular para
ingresar en la sangre y eliminarse con la orina.
Los NADH generados durante la glucólisis no ingresan en las mitocondrias.
En presencia de oxígeno, por cada molécula de glucosa se generan 36 ATP.

PROCESO
NADH
FADH
CO2
ATP
GLUCÓLISIS (Glucosa)
2
2
DESCARBOXILACIÓN
OXIDATIVA (2 piruvatos)
2
2
CICLO DE KREBS (2 Acetil-CoA)
6
2
4
2
SUB-TOTALES
10
2
4
RENDIMIENTO EN ATPs
X 3 ATP c/u
X 2 ATP c/u
TOTAL DE ATPs (38)
30 ATP
4 ATP
4 ATP
En las células musculares el piruvato puede convertirse en lactato.
En las mitocondrias de la grasa parda la energía generada por las oxidaciones se disipa en
forma de calor.
Remoción de Ca+2 del citosol.
Síntesis de aminoácidos.
Síntesis de esteroides.
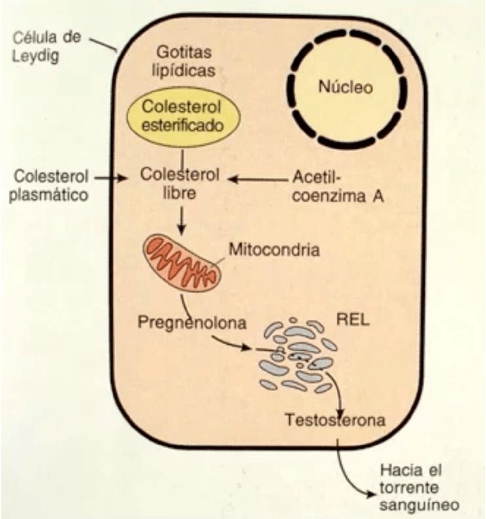
En algunas células de la
corteza suprarrenal, los ovarios y los testículos, las mitocondrias
participan en la síntesis de diversos esteroides (
función esteroidogénica). Para ello el
colesterol captado por las células debe ser transportado hacia las mitocondrias, donde, por
acción de una enzima localizada en la membrana mitocondrial interna, se convierte en
pregnenolona. La pregnenolona sale de la mitocondria e ingresa en el retículo
endoplasmático liso. Allí continua su metabolismo mediante diversas enzimas que actúan
secuencialmente.
REPRODUCCIÓN DE LAS MITOCONDRIAS
Las mitocondrias se reproducen para reemplazar a las que se desaparecen y su número se
duplica antes de cada división celular.
Para que las células se puedan dividir todos sus componentes -incluídas las mitocondrias-
deben previamente duplicarse. En las células que no se multiplican, o que poseen interfaces
prolongadas, las mitocondrias envejecen y son degradadas por los autofagosomas; no
obstante, su número permanece relativamente estable, debido a la formación de nuevas
mitocondrias. Así, sea para su renovación o por necesidades derivadas de la proliferación
celular, las mitocondrias deben ser dotadas de un mecanismo que les permita multiplicarse
en cantidades adecuadas y en los momentos oportunos.
La reproducción de las mitocondrias no se produce como consecuencia de un ensamblaje
espontáneo de los componentes que las integran sino por la división (fisión) de mitocondrias
preexistentes, las cuales deben previamente duplicar su tamaño.
La división de las mitocondrias se produce durante todo ciclo celular, tanto en la interfase
como en la mitosis. No obstante, no todas las mitocondrias se multiplican, por ende, algunas
deben dividirse repetidas veces durante el curso de un mismo ciclo celular a fin de
compensar a ñas que omiten hacerlo.
El ADN mitocondrial es diferente del ADN nuclear.
1. Es circular, carece de histonas y se replica a partir de un solo punto de origen.
2. Es muy pequeño.
3. Posee muy pocas y a la vez muy cortas secuencias no esenciales, es decir, que no
codifican ninguna clase de ARN.
4. Las mitocondrias poseen varias copias de un mismo ADN y no dos como el ADN
nuclear. Pero lo más singular no es la diferencia en el número de copias, sino que son
heredadas enteramente de la madre, ya que, como se sabe, en el momento de la
fecundación virtualmente todas las mitocondrias (y por ende las moléculas de ADN
mitocondrial) son aportadas por el ovocito.
La síntesis de las proteínas mitocondriales requiere una adecuada coordinación.
Aunque la mitocondria posee ADN, ARNm, ARNt y ribosomas propios, las proteínas que
fabrica son muy pocas -13 en total-; por consiguiente, la mayor parte de las que necesita
para su crecimiento y reproducción debe importarlas desde el citosol.

MITOCONDRIAS.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.