
1
I.S.U Instituto Superior Universitario
METABOLISMO DE HIDRATO DE CARBONO
En el proceso de digestión de los hidratos de carbono se degrada a los glúcidos provenientes
de los alimentos hasta el estado de monosacáridos, ya que sólo este tipo de compuestos
pueden absorberse en la mucosa intestinal y metabolizarse luego en las células. Después de la
absorción, estos monosacáridos son transportados hacia el hígado por la circulación portal. En
hígado, tanto la galactosa como la fructosa pueden ser transformadas en glucosa.
La principal función de la glucosa en el organismo es de servir como combustible de las células;
su oxidación produce energía utilizable para la realización de trabajo. También es utilizada
como materia prima para algunas síntesis.
El hígado es capaz de captar buena parte de la glucosa que le llega para formar una
macromolécula polimérica, el glucógeno, que constituye un verdadero material de reserva.
Ésta síntesis de glucógeno (glucógeno-génesis), es un proceso que requiere energía, es decir es
anabólico.
DURANTE EL PERÍODO POST PANDRIAL (PERÍODO DE ABSORCIÓN DESPUÉS DE UNA
COMIDA), Y SI LA COMIDA HA SIDO RICA EN GLÚCIDOS, EL HÍGADO NO ALCANZA A
CAPTURAR TODA LA GLUCOSA QUE LE LLEGA Y TRANSFORMARLA EN GLUCÓGENO, POR LO
CUAL PARTE DE ESTA GLUCOSA PASA A LA CIRCULACIÓN GENERAL (EL NIVEL DE GLUCOSA
NORMAL EN SANGRE ES DE 80 A 120 MG).
Todos los tejidos reciben un aporte de glucosa. El tejido muscular puede almacenar esa
glucosa en forma de glucógeno, es decir que existen otros tejidos además del hígado donde se
puede producir glucógeno.
Según necesidades metabólicas, el glucógeno del hígado puede ser desdoblado en glucosa,
liberándola a la circulación en general (glucogenólisis). La glucogenólisis hepática es
importante para mantener el nivel de glucosa en sangre (glucemia).
A diferencia del hígado, el tejido muscular no puede liberar glucosa libre a partir de su
glucógeno, ya que la degradación del glucógeno en el músculo lleva a la producción de
piruvato y lactato como productos finales.
Como ya dijimos la glucosa sirve de combustible a las células; también el glucógeno del
músculo y de otros tejidos es degradado para proveer energía. Éste proceso catabólico tanto
del glucógeno como de la glucosa se puede dividir en dos partes:
Una vía metabólica que se cumple aún en ausencia de oxígeno (en anaerobiosis),
denominada GLUCÓLISIS O VÍA DE EMBDEN – MEYERHOF. La etapa final de la
glucólisis comprende la formación del piruvato, que se reduce a lactato cuando la
provisión de oxígeno es insuficiente o nula. El aumento de lactato en sangre y en orina
después de un ejercicio intenso es expresión de la actividad glucolítica.
En presencia de oxígeno, el piruvato formado durante la glucólisis sufre un proceso de
oxidación total hasta CO
2
y H
2
O. el piruvato experimenta una descarboxilación que lo
convierte en acetato. Luego éste acetato es oxidado totalmente en el ciclo de krebs
para la producción de energía.

2
I.S.U Instituto Superior Universitario
El lactato producido por la degradación del glucógeno o de la glucosa producido por la
contracción muscular, es convertido de nuevo a glucosa en el hígado y devuelto a los músculos
a través de la circulación. Éste mecanismo fisiológico se denomina CICLO DE CORI.
Gracias a la gran difusibilidad del lactato, buena parte del mismo pasa a la sangre desde donde
es captado por el hígado, el cual tiene la capacidad de convertirlo en glucosa y glucógeno. La
transformación de aquellos metabolitos no glucídicos en glucosa y glucógeno se denomina
gluco-neo-génesis.
Ingreso de glucosa en las células
Para que la glucosa se absorba en el intestino existen las vellosidades intestinales, las cuales
poseen un sistema de co-transporte glucosa-Na
+
, que introduce glucosa en las células
intestinales aprovechando el gradiente de sodio creado por el funcionamiento de la bomba de
sodio. Éste mecanismo permite acumular glucosa en el citosol y de aquí pasa a la circulación
portal por difusión facilitada. Una vez en sangre, la glucosa llega a las células de diversos
tejidos y penetra en ellas por difusión facilitada.
Fosforilación de la glucosa: la fosforilación de la glucosa es el paso inicial
de todas las vías de utilización de los monosacáridos. Cualquiera sea el destino de la
glucosa, la primera transformación que experimenta es su esterificación con
ortofosfato para formar glucosa-6-fosfato. (G-6-P). la reacción es catalizada por la
enzima hexoquinasa, la cual se encuentra presente en todas las células. Ésta enzima
posee cuatro isozimas de hexoquinasa I, II, III, que se encuentran en distintos tejidos y
la IV, denominada glucoquinasa que se encuentra exclusivamente en el hígado y es
altamente específica, ya que sólo actúa sobre la D-glucosa. Las hexoquinasas I, II, III

3
I.S.U Instituto Superior Universitario
son inhibidas por la G-6-P, producto de la reacción, que actúa como efector alostérico
negativo. Éstas aseguran a continua utilización de la glucosa por las células y la
provisión de energía, aunque los niveles de glucemia varíen. La glucoquinasa sólo
permite la captación de glucosa por el hepatocito cuando los niveles de glucosa en
sangre aumentan (ejemplo después de una comida). Tanto las hexoquinasas como la
glucoquinasa requieren de la presencia de ATP y Mg
++
.
La reacción catalizada por las hexoquinasas comprenden dos reacciones acopladas, una
de síntesis del éster glucosa 6 fosfato, endergónica e irreversible, donde las membranas
celulares son impermeables a éste tipo de éster, lo que impide difundirse hacia fuera de la
célula, y poder seguir las distintas vías metabólicas. Por otra parte, la rápida conversión de
glucosa en G-6-P mantiene baja la concentración intracelular de glucosa y favorece el
ingreso de más glucosa.
VÍAS METABÓLICA DE LA GLUCOSA
La glucosa 6 fosfato pude seguir diferentes vías:
GLUCÓGENO-GÉNESIS:
la formación de glucógeno a partir de glucosa puede realizarse
en muchos tejidos del organismo, pero adquiere real importancia en el hígado y en
músculo, el primero puede llegar a contener 5% de su peso en glucógeno si se lo
analiza después de una comida rica en carbohidratos, y el músculo posee 1%.. Es un
proceso anabólico, el cual se lleva a cabo mediante una serie de reacciones:
1) Fosforilación de la glucosa (G-6-P).
2) Formación de glucosa-1-fosfato: una transferasa llamada fosfogluco mutasa
cataliza la transferencia intramolecular del grupo fosfato del carbono 6 al
carbono 1. Entonces se convierte en glucosa 1 fosfato. Ésta enzima requiere la
presencia de Mg
++
y utiliza como cofactor a la glucosa 1.6-bifosfato. Es una
reacción reversible.
Hexoquinasa
GLUCOSA
GLUCOSA-6-fosfato.
Fosfo-gluco-mutasa
Glucosa1.6bifosfato

4
I.S.U Instituto Superior Universitario
3) Activación de la glucosa: la glucosa 1 fosfato reacciona con un nucleótido de
alta energía (uridín trifosfato UTP) y forman el uridín difosfato glucosa (UDPG)
y pirofosfato (PPi). La reacción es catalizada por la uridín difosfato
pirofosforilasa y es irreversible. Se dice que la glucosa se ha activado por su
unión al UDP.
GLUCOSA-1-P + UTP UDPG + PPi
4) Adición de la glucosa la estructura polimérica: la glucosa activada es
transferida a uno de los restos de glucógeno pre-existentes en el tejido. Se
produce la unión glucosídica entre el carbono 1 y el carbono 6 de una glucosa
terminal en las cadenas del glucógeno del tejido. Ésta reacción es catalizada
por la enzima glucógeno sintetasa. La reacción es prácticamente irreversible.
5) Formación de las ramificaciones: cuando se ha formado una cadena de hasta 8
o más residuos de glucosa, interviene otra enzima transfiriendo un segmento
de no menos de 6 glucosas para insertarlo, mediante una unión glucosídica 1-
6, sobre otra cadena vecina.
COMO DIJIMOS ÉSTE ES UN PROCESO ENDERGÓNICO, POR LO QUE SE REQUIERE
SUMINISTRO DE ENERGÍA. LA PRIMERA REACCIÓN DE FOSFORILACIÓN, COMÚN DE
TODAS LAS VÍAS CONSUME UNA MOLÉCULA DE ATP. EN A REACCIÓN DE ACTIVACIÓN
INTERVIENE UN UTP. EN LA REACCIÓN SIGUIENTE SE LIBERA UDP EL CUAL PUEDE
REGENERAR UTP. POR LO TANTO LA INCORPORACIÓN DE UNA MOLÉCULA DE GLUCOSA A
GLUCÓGENO REPRESENTA UN GASTO TOTAL DE 2 ATP.
uridín difosfato pirofosforilasa

5
I.S.U Instituto Superior Universitario
GLUCOGENÓLISIS:
no es simplemente el proceso inverso de la glucogenogénesis.
Como en esta última vía existen etapas irreversibles, la degradación del glucógeno
debe realizarse utilizando, en esos pasos, enzimas distintas a las de la vía anabólica.
Las etapas son las siguientes:
1) Fosforólisis del glucógeno: la degradación del glucógeno es iniciada por la
acción de fosforilasa, que cataliza la ruptura de uniones glucosídicas
1,4 por
inserción de fosfato en el carbono 1. El ortofosfato utilizado en esta reacción
proviene del medio (Pi); no es necesario gasto de energía (ATP). La fosforilasa
actúa a partir del extremo no reductor de las ramificaciones y libera glucosa-1-
fosfato. La acción enzimática se detiene cuatro restos de glucosa antes de la
próxima unión 1,6. Aquí interviene otra enzima, oligo- (1,4)
glucotransferasa, que desprende el trisacárido terminal de la ramificación y lo
transfiere al extremo de una rama vecina, al cual lo une por enlace
1,4. La
ramificación queda reducida a una sola glucosa con unión
1,6.

6
I.S.U Instituto Superior Universitario
2) Hidrólisis de uniones glucosídicas
1,6: la ruptura de este enlace se realiza
por hidrólisis, catalizada por
1,6 glucosidasa o enzima desramificante, que
deja glucosa en libertad. Después de esta intervención por la enzima
desramificante, la cadena es nuevamente atacada por la fosforilasa, que
continúa liberando glcucosa-1-fpsfato hasta que la próxima unión
1,6 se
encuentre a una distancia de cuatro restos glucosa; entonces se repite la
participación de las otras enzimas. La acción concertada de fosforilasa, oligo-
alfa1,4glucotransferasa y alfa 1,6 glucosidasa libera glucosa 1 fosfato y algunas
glucosas (la enzima desramificante actúa por hidrólisis y no por Fosforólisis).
En promedio se produce una glucosa libre por cada nueve glucosas 1fosfato, lo
cual da una idea del grado de ramificación en la molécula de glucógeno. Sólo
unidades de glucosa en la posición de ramificación son liberadas como glucosa
libre. Todas las otras aparecen como G-1-P.
3) Formación de glucosa-6-fosfato: la glucosa-1-fosfato es convertida en glucosa-
6-fosfato por la fosfoglucomutasa. Es la misma reacción de la
glucogenogénesis, en sentido inverso.
4) Formación de glucosa libre: la última etapa es la hidrólisis de glucosa-6-fosfato
a glucosa y fosfato inorgánico, catalizada por glucosa-6-fosfatasa.
GLUCOSA-6-P GLUCOSA + Pi.
H
2
O
La glucosa-6-fosfatasa se encuentra en membrana del retículo endoplásmico (RE) del hígado,
riñón e intestino, pero no en músculo. Esto explica porqué el hígado, riñón e intestino pueden
ceder glucosa a la circulación y el músculo no. En el músculo, el glucógeno inicia su
GLUCOSA-6-FOSFATASA

7
I.S.U Instituto Superior Universitario
degradación con etapas similares a estas. La glucosa-6-fosfato formada no puede hidrolizarse
por falta de la enzima glucosa-6-fosfatasa y sigue su camino catabólico en el propio músculo,
principalmente por vía de la glucólisis.
PAPEL FUNCIONAL DEL GLUCOGENO
En la mayoría de los tejidos el glucógeno representa una reserva a la cual se recurre para
obtener glucosa durante períodos de hipoglucemia o hipoxia.
Sin embargo, el papel del glucógeno no es el mismo en todos los órganos. Especialmente
notable es la diferencia entre el hígado y músculo, ambos muy ricos en glucógeno.
El hígado cumple un rol muy importante como regulador de la glucemia, asegurando la
provisión constante de glucosa a todos los tejidos.
Inmediatamente después de una comida aumenta transitoriamente la glucemia.
En estos períodos de exceso, el hígado sustrae glucosa de la circulación y la almacena como
glucógeno. En los intervalos entre comidas, el hígado degrada su glucógeno y libera glucosa a
la sangre.
En el músculo el glucógeno actúa como reserva rápidamente movilizable que provee
combustible para la contracción. El músculo no puede liberar glucosa, sus depósitos de
glucógeno son utilizados exclusivamente por el propio tejido.

8
I.S.U Instituto Superior Universitario
GLUCOLISIS: la principal vía inicial del catabolismo de glucosa es la serie de reacciones
llamada glucólisis o vía de Ember.Meyerhof. en el curso de esta vía, una molécula de
glucosa es desdoblada en dos moléculas de piruvato y se produce energía utilizable. El
proceso puede cumplirse en ausencia de oxígeno. Este mecanismo metabólico
proveedor de energía es evolutivamente el más antiguo. La glucólisis es un notable
ejemplo de la unidad del mundo biológico; funciona en todos los organismos vivientes,
aún filogenéticamente muy distantes, siguiendo exactamente las mimas etapas. En
distintas especies puede variar el destino final del piruvato formado. Muchos
microorganismos realizan por esta vía la degradación anaeróbica de glucosa y otros
monosacáridos, el proceso es denominado fermentación. Los productos terminales
difieren en distintos microorganismos. Algunos forman lactato (fermentación láctica),
otros producen etanol y dióxido de carbono (fermentación alcohólica), otros ácidos
acético. En seres aerobios, la glucólisis constituye la primera parte del catabolismo de
glucosa. en ellos el piruvato continúa su degradación por vía oxidativa hasta CO
2
y H
2
O.
sin embargo, en organismo, en organismos aerobios cuando un tejido funciona con
insuficiente provisión de oxígeno, por ejemplo en el músculo esquelético durante un
ejercicio brusco e intenso, el piruvato es convertido en lactato como fermentación
láctica. Las transformaciones químicas de la glucólisis comprenden cambios en la
molécula del sustrato original (glucosa) con producción de metabolitos ricos en
energía, que pueden transferir restos fosforilo a ADP.
La s series de reacciones de la glucólisis puede dividirse en dos fases:
1) En la primera, la hexosa sufre dos fosforilaciones y termina dividida en dos
triosas-fosfato. Es ésta una fase preparatoria durante la cual se invierte
energía para formar compuestos incapaces de escapar de la célula y más
reactivos que la glucosa, es decir, más aptos para sufrir nuevas
transformaciones. El resultado del primer grupo de reacciones es la ruptura de
la molécula inicial de 6 carbonos en dos de tres carbonos, gliceraldehído-3-
fosfato (G3P) y dihidroxiacetonafosfato (DHAP). Esta última es transformada
en G3P, razón por la cual puede considerarse que cada molécula de glucosa
ingresada en la vía se convierte en dos G3P.
2) En la segunda etapa, el gliceraldehído-3-fosfato sufre oxidación y
redistribución de sus átomos con formación de intermediarios de alta energía
que participan en la síntesis de ATP por fosforilación a nivel de sustrato. En
esta fase se obtiene el rédito energético de la vía.
TODAS LAS ENZIMAS INVOLUCRADAS SE ENCUNETRAN EN EL CITOSOL, RAZÓN POR LA
CUAL LA GLUCÓLISIS SE CUMPLE ÍNTEGRAMENTE EN EL CITOPLASMA DE LAS CELULAS.
PRIMERA FASE DE LA GLUCÓLISIS:
A. Formación de la glucosa-6-fosfato: la utilización de glucosa exige, como etapa
inicial obligatoria, su fosforilación en el carbono 6. Las reacciones necesarias
para obtener G-6P son distintas si la materia prima utilizada por el tejido es
glucosa o glucógeno. A partir de la glucosa la fosforilación es catalizada por
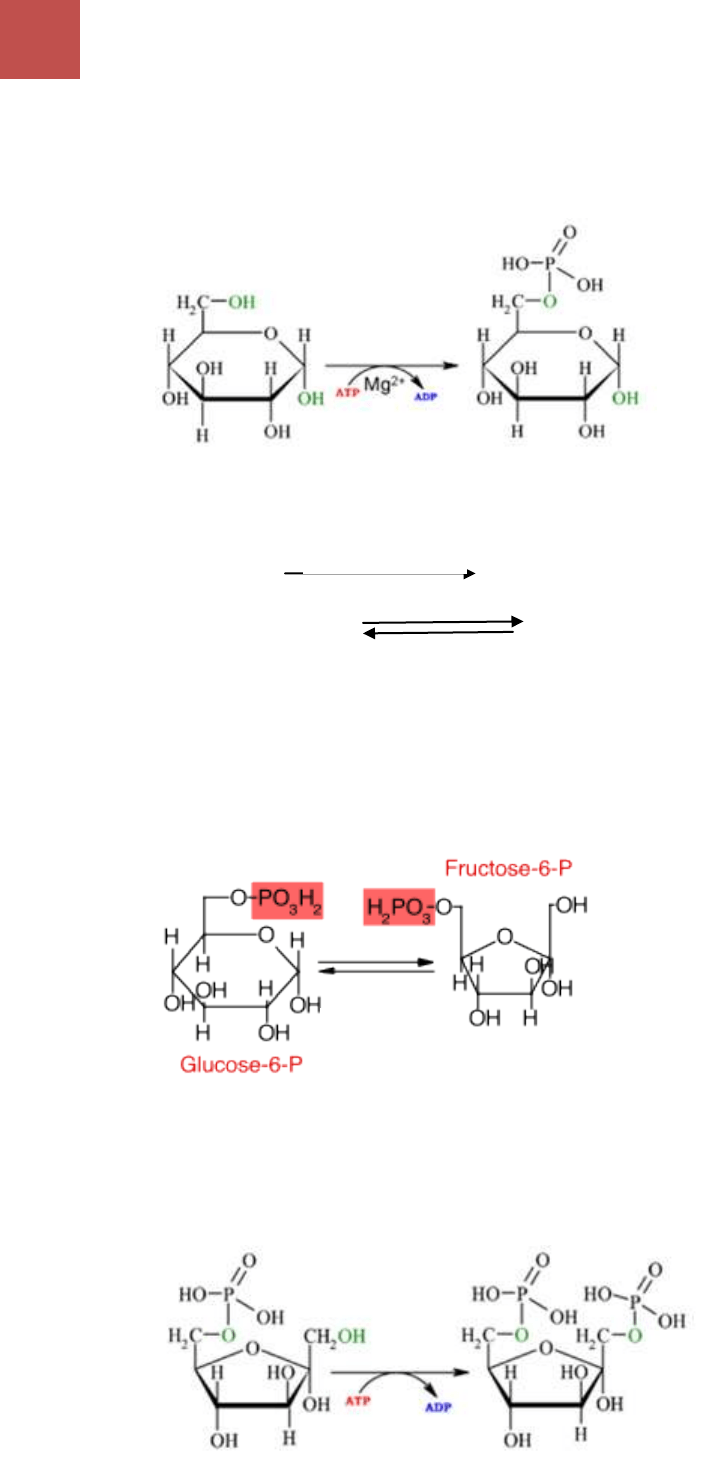
9
I.S.U Instituto Superior Universitario
hexoquinasas. La reacción es reversible en las condiciones reinantes de la
célula. Cuando se parte de glucógeno, la degradación hasta glucosa-6-fosfato
se cumple en dos etapas, catalizadas por fosforilasa y fosfoglucomutasa.
GLUCOSA GLUCOSA-6-FOSFATO.
GLUCOGENO GLUCOSA-1-FOSFATO
GLUCOSA-1-FOSFATO GLUCOSA-6-FOSFATO
B. Formación de fructosa-6-fosfato: es un proceso de isomeración, la glucosa-6-
fosfato es convertida en fructosa-6-fosfato (F6P). la reacción es catalizada por
fosfoglucoisomerasa, es reversible. La fosfoglucoisomerasa requiere de iones
de Mg
+2
o Mn
+2
.
C. Fosforilación de fructosa-6-fosfato: la fructosa-6-fosfato es fosforilada en el
carbono 1 y se transforma en fructosa-1,6-bifosfato (F1,6-bisP). La reacción
exige la transferencia de un grupo fosforilo cedido por ATP. Cataliza esta
transformación la fosfofructoquinasa, en presencia de iones de magnesio.
hexoquinasa
FOSFORILASA
FOSFOGLUCOMUTASA
fosfoglucoisomeras
a
fosfofructoquinasa

10
I.S.U Instituto Superior Universitario
El acoplamiento con la hidrólisis de ATP hace posible la síntesis del éster- fosfórico en
carbono1. La reacción, esencialmente irreversible, es muy importante en la regulación de
esta vía metabólica. La enzima es activada por AMP, ADP y fructosa 2,6-bifosfato, e inhibida
por ATP y citrato.
D. Formación de triosas-fosfato: la fructosa-1,6-bifosfato es escindida en dos
triosas fosfato, gliceraldehído-3-fosfato (G3P) y dihidroxiacetonafosfato
(DHAP). La reacción es catalizada en ambos sentidos por una liasa, la aldosa A.
La reacción de ruptura de fructosa-1,6-bifosfato transcurre con toda facilidad porque los
productos (triosas-fosfato) son eliminados rápidamente por las reacciones siguientes.
E. Interconversión de triosas-fosfato: de las dos triosas-fosfato producidas en la
reacción anterior, sólo D-gliceraldehído-3-fosfato continúa directamente la vía
metabólica. La DHAP sigue el camino de la glucólisis, pero para ello debe ser
convertida en G3P. esto es posible gracias a la conversión reversible de las
triosas por acción de triosa-fosfato isomerasa.
SEGUNDA FASE DE LA GLUCÓLISIS: en la reacción anterior, DHAP se convierte en G3P.
por esta razón se considera que cada molécula de glucosa da lugar a dos moléculas de
G3P.
F. Oxidación y fosforilación del gliceraldehído-3-fosfato: es una etapa de gran
importancia en la glucólisis. En ella se produce deshidrogenación del G3P. la
energía liberada es utilizada para introducir ortofosfato (Pi) del medio y formar
1,3-bisfosfoglicerato. La reacción es catalizada por gliceraldehído-3-fosfato
deshidrogenasa oxidoreductasa que utiliza NAHD como coenzima.

11
I.S.U Instituto Superior Universitario
G. Fosforilación a nivel de sustrato: el fosfato de alta energía es transferido a 1,3-
bifosfoglicerato a ADP, por acción de fosfogliceratoquinasa, se produce 3-
fosfoglicerato y ATP. Las reacciones F y G sumadas resultan reversibles en las
condiciones de la célula. ES ÉSTA UNA FOSFORILACIÓN A NIVEL DE SUSTRATO
Y LA PRIMERA REACCIÓN DE LA GLUCÓLISIS EN LA CUAL HAY CONSERVACIÓN
DE ENERGÍA.
H. Formación de 2-fosfoglicerato: el 3-fosfoglicerato es convertido en 2-
fosfoglicerato por transferencia intramolecular de fosforilo. Ésta reacción es
catalizada, en ambos sentidos, por fosfoglicerato mutasa en presencia de
iones de magnesio.

12
I.S.U Instituto Superior Universitario
I. Formación de fosfoenolpiruvato: se produce una deshidratación y
redistribución intramolecular en el 2-fosfoglicerato para generar un
compuesto rico en energía, el fosfoenolpiruvato. Cataliza esta reacción
reversible la enalasa, que requiere magnesio o manganeso.
J. Segunda fosforilación a nivel de sustrato: el fosfoenolpiruvato tiene un
potencial de transferencia suficiente para cederé fosfato a ADP y formar ATP.
La reacción es catalizada por piruvato quinasa y necesita iones de magnesio o
manganeso. El catión potasio tiene efecto activador sobre esta enzima. El
enolpiruvato se transforma espontáneamente en piruvato.

13
I.S.U Instituto Superior Universitario
El enolpiruvato se
transforma espontáneamente en piruvato.
Se ha generado otra molécula de ATP por fosforilación a nivel de sustrato.
K. Formación de lactato: el piruvato formado puede seguir distintas vías: cuando
la disponibilidad de oxígeno es escasa a nula, el piruvato es reducido a lactato
por acción de la lactato deshidrogenasa, enzima que utiliza NAD como
coenzima. El proceso es fácilmente reversible.
En ausencia o deficiencia de oxígeno el NADH formado durante la oxidación del G3P^en
la reacción F no puede oxidarse a NAD cediendo sus equivalentes de reducción a la cadena
respiratoria, pues ésta no funciona (en anaerobiosis, esa transferencia se realiza por vía
indirecta).
La glucólisis está limitada por la disponibilidad de NAD, se detiene cuando todo el NAD
existente en el citosol se reduce a NADH. La conversión de piruvato en lactato es un

14
I.S.U Instituto Superior Universitario
mecanismo que asegura la reoxidación del NADH y permite el funcionamiento sostenido de
la glucólisis.
Ésta reacción explica porqué el lactato es el producto final de la glucólisis en tejidos que
funcionan en relativa anaerobiosis como por ejemplo el músculo esquelético.
BALANCE ENERGÉTICO DE LA GLUCÓLISIS:
Cada mol de glucosa ingresado en la vía da origen a dos moles de triosa fosfato y finalmente se
convierte en dos moles de lactato. Hay dos etapas en las cuales se consume ATP.
Un mol de glucosa requiere de ATP en la fosforilación inicial para formar G6P y otro mol en la
segunda fosforilación de fructosa-6-p a fructosa-1,6,bifosfato. Cuando se parte de glucógeno
no se consume ATP en la fosforilación inicial. En la segunda fase de la glucólisis, dos etapas
producen ATP por fosforilación a nivel de sustrato. Son las reacciones catalizadas por
fosfogliceroquinasa y piruvatoquinasa. Cada mol de 1,3-bifosfoglicerato genera uno de ATP a
partir de ADP y cada mol de fosfoenolpiruvato engendra otro de ATP. Por lo tanto, como una
glucosa da lugar a dos triosas-fosfato, EL RENDIMIENTO POR MOL DE GLUCOSA ES DE 4
MOLES DE ATP.
EL BALANCE FINAL DE LA GLUCÓLISIS ES DE UNA GANANCIA NETA DE DOS MOLES DE ATP
POR MOL DE GLUCOSA UTILIZADA.
La hidrólisis de un mol de ATP a ADP cede aproximadamente 30,5 KJ (7,3 Kcal) en
condiciones estándar. El rendimiento energético de la glucólisis por mol de glucosa es de
61KJ (14,6 Kcal).

15
I.S.U Instituto Superior Universitario
Si se tiene en cuenta que el total de energía contenida en la glucosa es 2870 KJ/mol (686
Kcal/mol), el rendimiento logrado por la glucólisis es ínfimo.
PAPEL FUNCIONAL DE LA GLUCÓLISIS
La glucólisis es la principal vía inicial de utilización de glucosa en todos los tejidos, pero en
algunos tiene significado especial.
MÚSCULO ESQUELÉTICO: la glucólisis es la vía de generación de ATP requerido por la
contracción muscular durante ejercicios intensos.
TEJIDO ADIPOSO: una de las principales funciones de la glucólisis en este tejido, especializado
en el almacenamiento de triacilglicéridos, es proveer dihidroxiacetonafosfato, precursora del
glicerolfosfato utilizado en la síntesis de esos compuestos.
GLÓBULOS ROJOS: no tienen mitocondria y no pueden generar ATP por vías oxidativas.
Dependen de la glucólisis para la síntesis de ATP.
DESCARBOXILACIÓN OXIDATIVA DEL PIRUVATO
Cuando existe adecuada provisión de oxígeno, el piruvato producido en la vía glucolítica, es
oxidado a dióxido de carbono y agua. Incluso el lactato formado en anaerobiosis sigue el
mismo camino cuando hay disponibilidad de oxígeno; para ello debe ser convertido en
piruvato por acción de la lactato deshidrogenasa. De esta manera el lactato resultante de la
actividad muscular intensa puede ser utilizada como combustible.
El piruvato formado en el citosol como producto de la glucólisis es degradado oxidativamente
dentro de las mitocondrias. Para ello atraviesa la membrana interna de estas organelas gracias
a un transportador que lo introduce en la matriz. Aquí se cumple el primer paso de su
degradación por descarboxilación oxidativa, en la cual pierde un grupo carboxilo, se
desprende dióxido de carbono y queda de dos restos carbono (acetilo o acetato).
Ésta descarboxilación es catalizada por un sistema multienzimático denominado complejo
piruvato deshidrogenasa. Éste complejo está constituido por tres enzimas:
a) Piruvato descarboxilasa o E1
b) Dihidrolipoil transacetilasa o E2
c) Dihidrolipoil deshidrogenasa o E3
Participan 5 coenzimas:
a) Pirofosfato de tiamina (PPT)
b) Ácido lipoico
c) Coenzima A.
d) FAD
e) NAD
El ordenamiento de los componentes dl sistema en el complejo asegura su funcionamiento
eficiente.
Este documento contiene más páginas...
Descargar Completo
proteinas mio.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.