LÍPIDOS
Los lípidos biológicos constituyen un grupo químicamente heterogéneo de compuestos cuya
característica común y definitoria es su insolubilidad en agua. Las funciones biológicas de los
lípidos son tan diversas como su química. Son solubles en disolventes orgánicos (benceno,
cloroformo, éter, hexano, etc. ). Algunos de ellos tienen segmentos que sí son solubles en
agua, como los fosfolípidos.
En muchos organismos, las grasas y los aceites son las formas principales de almacenamiento
energético mientras que los fosfolípidos y los esteroles constituyen los principales elementos
estructurales de las membranas biológicas. Otros lípidos, aún estando presentes en cantidades
relativamente pequeñas, juegan papeles cruciales como cofactores enzimáticos,
transportadores electrónicos, pigmentos que absorben la luz, anclas hidrofóbicas para
proteínas, “chaperonas” que ayudan en el plegamiento de las proteínas de membrana,
agentes emulsionantes en el tracto digestivo, hormonas y mensajeros intracelulares.
CLASIFICACIÓN DE LOS LÍPIDOS
La heterogeneidad estructural de los
lípidos dificulta cualquier clasificación
sistemática. El componente lipídico
de una muestra biológica puede ser
extraído con disolventes orgánicos y
ser sometido a reacciones de
saponificación.
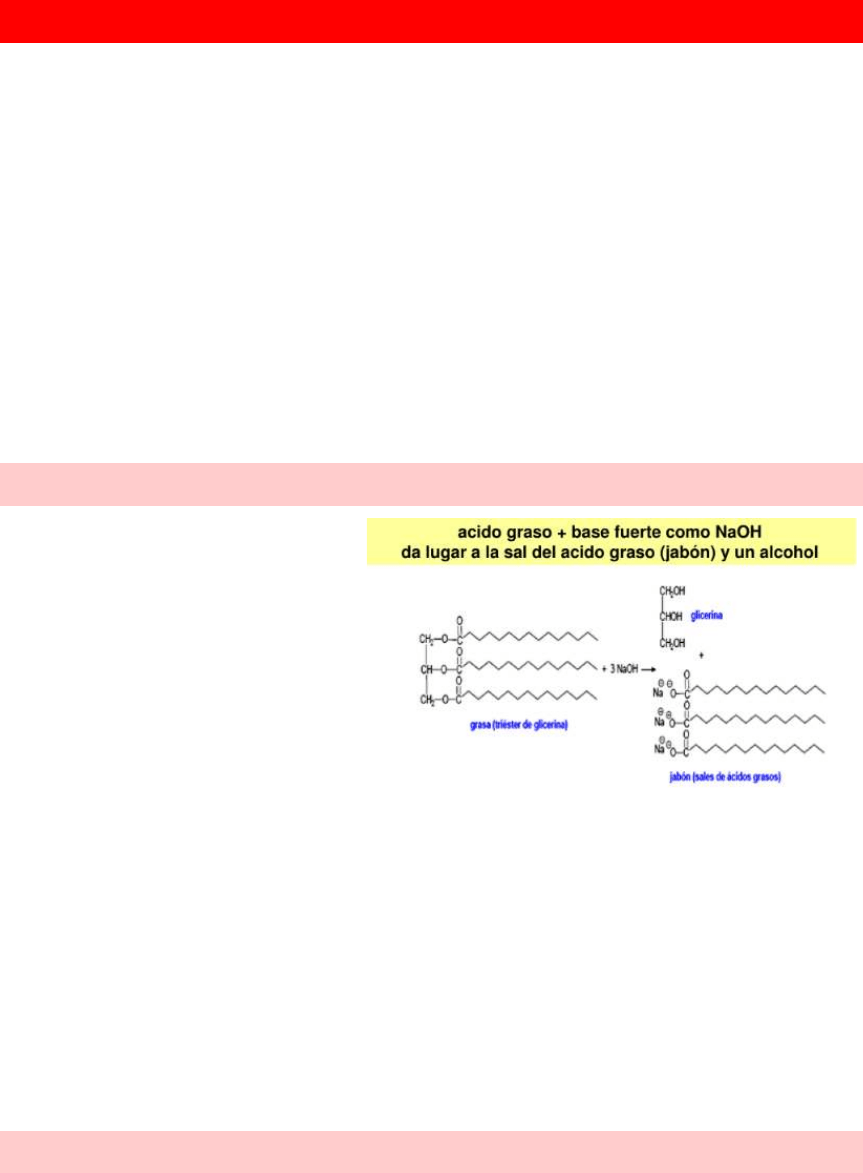
Lípidos saponificables: contienen ácidos grasos en su estructura y forman jabón
1- ácidos grasos
2- lípidos neutros (acilglicéridos y ceras)
3- lípidos anfipáticos (fosfolípidos y glucolípidos)
Lípidos insaponificables: no contienen ácidos grasos en su estructura y no forman jabón.
1- Terpenos
2- Esteroides
3- Eicosanoides
FUNCIÓN
1- Energética
2- Reserva de agua
3- Producción de calor
4- Estructural
5- Regulador
LÍPIDOS DE ALMACENAMIENTO
Las grasas y aceites, utilizados casi universalmente como formas de almacenamiento de
energía en los organismos vivos, son compuestos derivados de los ácidos grasos.
Los ácidos grasos son derivados hidrocarbonados con un nivel de oxidación tan bajo (esto es,
tan reducidos) como el de los hidrocarburos de los combustibles fósiles. La oxidación de los
ácidos grasos (a CO
2
y H
2
O) en las células, al igual que la oxidación explosiva de los carburantes
fósiles en los motores de combustión interno, es muy exergónica.
Los ácidos grasos son ácidos carboxílicos de cadena larga. Por lo general, contienen un número
par de átomos de carbono, normalmente entre 12 y 24. Son moléculas débilmente anfipáticas.
Según la naturaleza de la cadena carbonada, distinguimos.
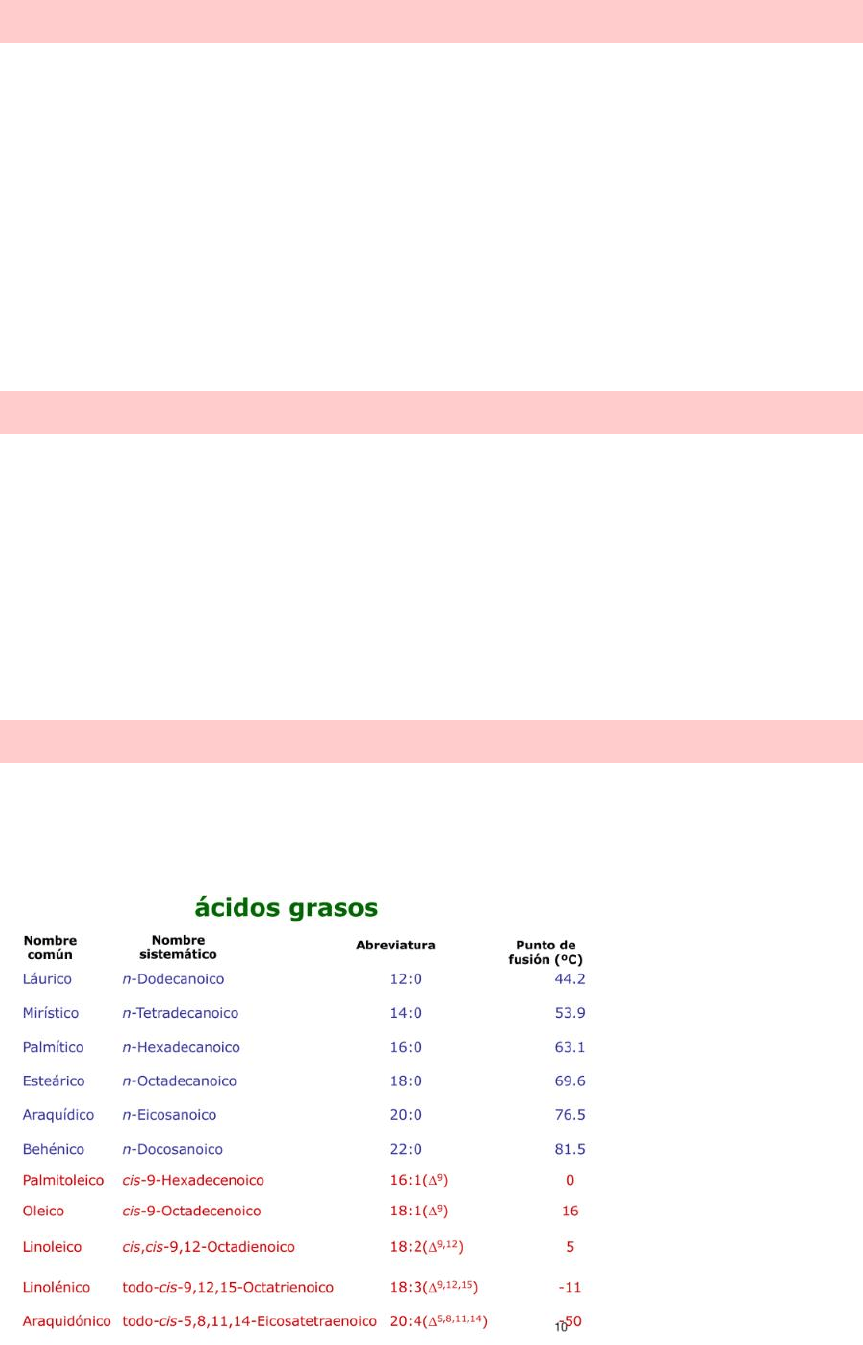
ÁCIDOS GRASOS SATURADOS
Presentan solo enlaces simples C-C y son muy poco reactivos.
Ejemplos:
1- Palmítico 16:0
2- Esteárico 18:0
3- Ácido miístico 14:0
4- Ácido araquídico 20:0
5- Ácido lignocérico 24:0
ÁCIDOS GRASOS INSATURADOS
Son aquellos que presentan al menos un doble enlace C=C (Cis). Este enlace produce un
quiebre en la molécula que disminuye su capacidad a empaquetarse. Tienen un punto de
fusión menor que los saturados, es decir que en su mayoría a temperatura ambiente son
líquidos.
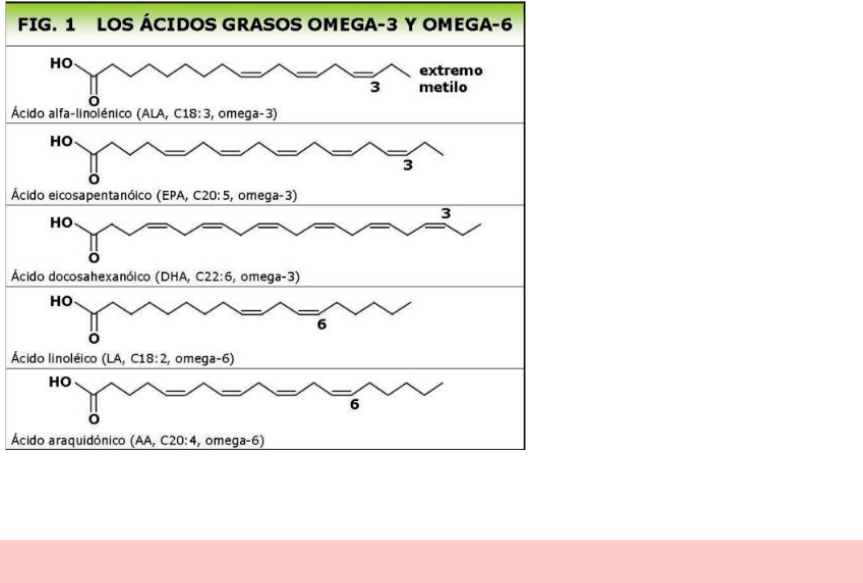
La serie omega hace referencia a un tipo de insaturación, pero el omega es contando desde el
otro lado. Según el ultimo a cuanto esté del carbono terminal
LOS ÁCIDOS GRASOS SON DERIVADOS DE HIDROCARBUROS
Los ácidos grasos son ácidos carboxílicos con cadenas hidrocarbonadas de 4 a 36 carbonos (C
4
a C
36
). En algunos ácidos grasos, esta cadena está completamente saturada y sin ramificar;
otros contienen uno o más dobles enlaces. Unos cuantos contienen anillos de tres carbonos,
grupo hidroxilo o grupos metilo ramificados.
Las propiedades físicas de los ácidos grasos y de los compuestos que los contienen vienen
determinados en gran parte por la longitud y el grado de insaturación de la cadena
hidrocarbonada. La cadena hidrocarbonada apolar explica la escasa solubilidad de los ácidos
grasos en agua. El ácido láurico (12:0, M, 200), por ejemplo, tiene una solubilidad de 0.063
mg/g de agua muy inferior a la de la glucosa (Mr180) que es de 1100 mg/g de agua. El grupo
ácido carboxílico es polar (y está ionizado a pH neutro) lo que justifica la ligera solubilidad en
agua de los ácidos grasos de cadena corta.
Los puntos de fusión están también muy influidos por la longitud y el grado de saturación de la
cadena hidrocarbonada. A temperatura ambiente (25°C), los ácidos grasos saturados desde
12:0 a 24:0 tienen una consistencia cérea, mientras que los ácidos grasos insaturados de estas
longitudes son líquidos oleosos. Esta diferencia en los puntos de fusión se debe a los diferentes
grados de empaquetamiento de las moléculas de los ácidos grasos. En los compuestos
totalmente saturados, la rotación libre alrededor de cada enlace Carbono-Carbono confiere
gran flexibilidad a la cadena hidrocarbonada; la conformación mas estable es la forma
totalmente extendida, en la que los impedimentos estéricos entre átomos vecinos están
reducidos al mínimo. Estas moléculas se pueden empaquetar fuertemente en ordenamientos
casi cristalinos con contactos por uniones de Van der Walls entre átomos a lo largo de la propia
cadena y átomos de cadenas vecinas. En los ácidos grasos insaturados, un doble enlace cis
provoca un doblamiento en la cadena hidrocarbonada. Los ácidos grasos con uno o más
doblamientos de este tipo no se pueden empaquetar tan fuertemente como los ácidos grasos
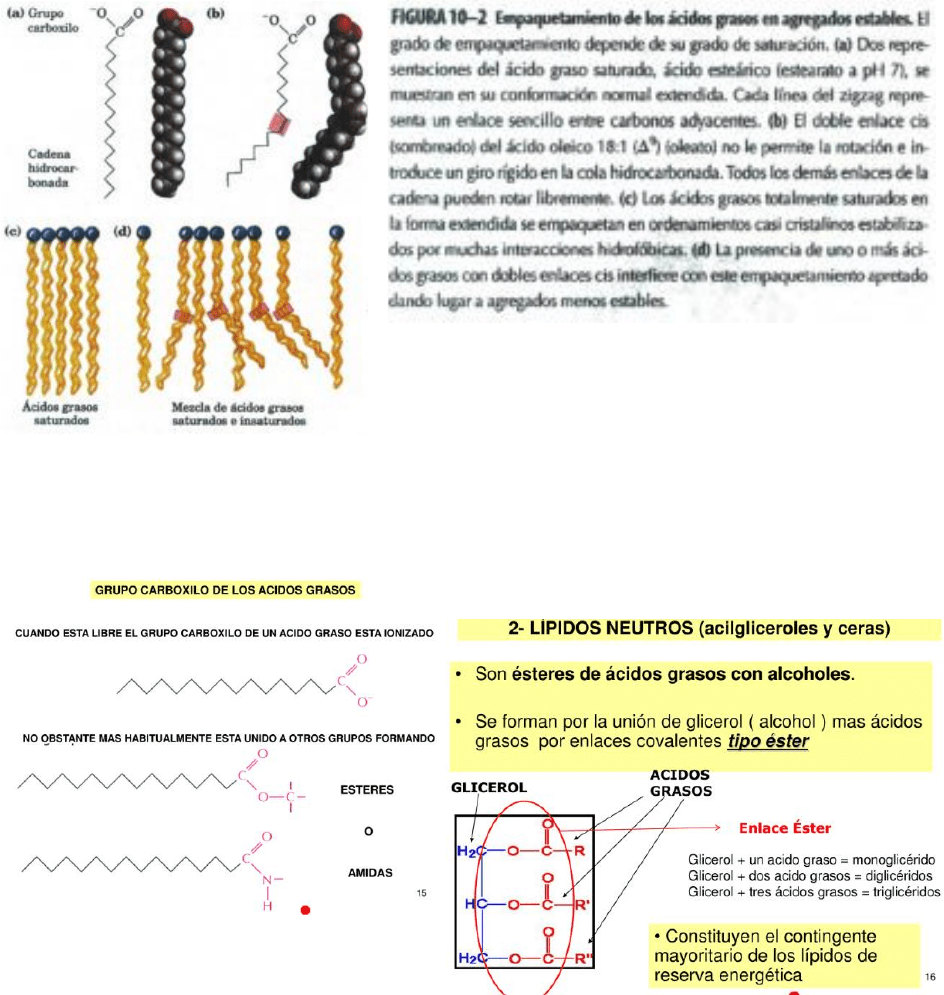
totalmente saturados, por lo que las interacciones entre ellos son más débiles. Dado que se
necesita menos energía térmica para desordenar estos conjuntos poco ordenados de ácidos
grasos insaturados, éstos tienen puntos de fusión claramente más bajos que los ácidos grasos
saturados de la misma longitud de cadena.
En los vertebrados los ácidos grasos libres (ácidos no esterificados, con un grupo carboxilato
libre) circulan por la sangre unidos de forma no covalente a una proteína portadora, la alúmina
sérica. No obstante, los ácidos grasos en su mayoría se encuentran presentes en el plasma
sanguíneo en forma de derivados del ácido carboxílico tales como ésteres o amidas. Al carecer
del grupo carboxilato cargado, estos derivados de los ácidos grasos son generalmente aún
menos solubles en agua que los ácidos grasos libres.

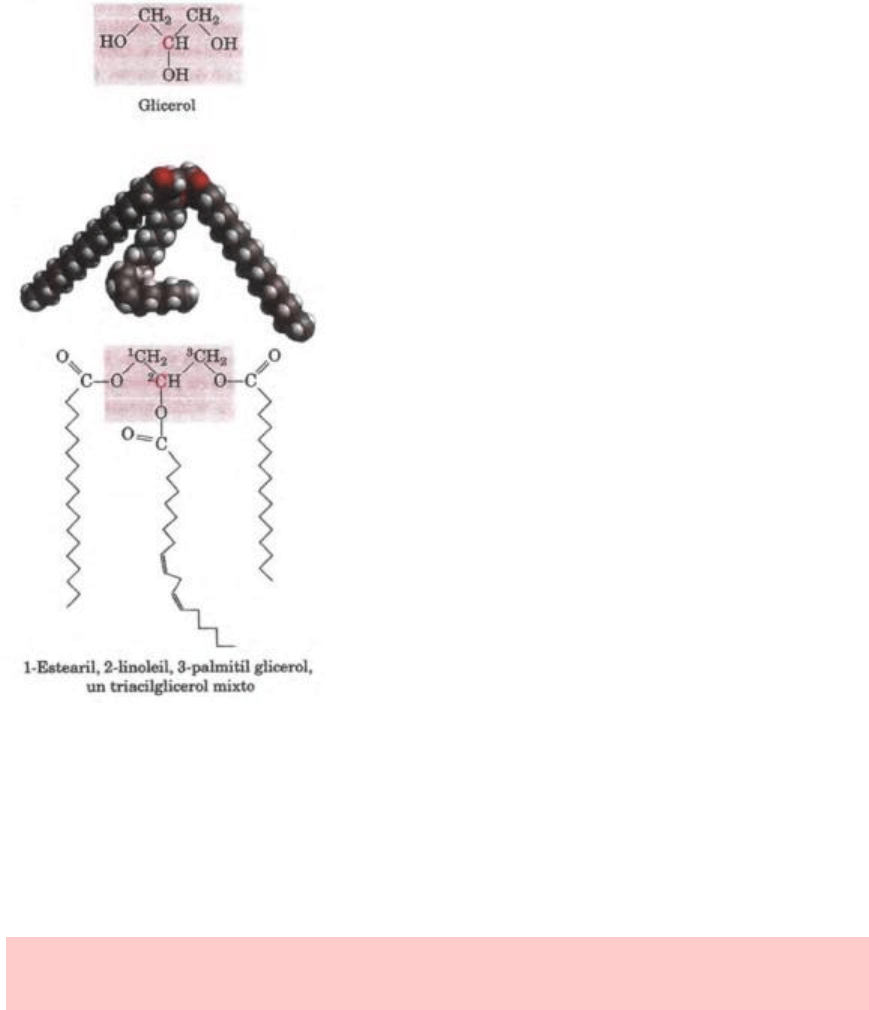
TRIACILGLICEROLES
Los triacilgliceroles son ésteres de ácidos grasos y
glicerol. Son reservas energéticas de uso tardío.
Los lípidos más sencillos obtenidos a partir de los
ácidos grasos son triacilgliceroles, también
denominados triglicéridos, grasas o grasas neutras.
Los triacilgliceroles están compuestos de tres ácidos
grasos unidos por enlaces éster con un solo glicerol.
Los que contienen el mismo tipo de ácido graso en las tres posiciones se denominan
triacilgliceroles simples y se denominan según el ácido graso que contienen. La triestearina, la
tripalmitina y la trioleína son, respectivamente, ejemplos de triacilglicéridos sencillos que
contienen 16:0, 18:0 y 18:1. La mayoría de los triacilglicéridos naturales son mixtos; éstos
contienen dos o más ácidos grasos diferentes. Se han de especificar el nombre y la posición de
cada ácido graso para designar sin ambigüedades estos compuestos.
Dado que los hidroxilos polares del glicerol y los carboxilatos polares de los ácidos grasos están
unidos por enlaces éster, los triacilgliceroles son moléculas apolares, hidrofóbicas,
prácticamente insolubles en agua. Los lípidos tienen densidades específicas menores que el
agua, lo que explica por qué las mezclas de agua y aceite (aceite y vinagre para aliñar la
ensalada, por ejemplo) tienen dos fases: el aceite, con menor densidad específica, flota sobre
la fase acuosa.
LOS TRIACILGLICEROLES APORTAN ENERGÍA ALMACENADA Y AISLAMIENTO
En la mayoría de células eucariotas, los triacilgliceroles forman una fase separada de gotitas
microscópicas oleosas en el citosol acuoso que sirven como depósito de combustible
metabólico. Las células especializadas de los vertebrados, denominadas adipocitos o células
grasas, almacenan grandes cantidades de triglicéridos en forma de gotitas de grasa que
ocupan casi totalmente la célula. Los triacilgliceroles se almacenan también, en forma de
aceite, en semillas de muchos tipos de plantas, proporcionando energía y precursores
biosintéticos durante la germinación de las semillas. Los adipocitos y las semillas en
germinación contienen lipasas, enzimas que catalizan la hidrólisis de los triacilgliceroles
almacenados, liberando ácidos grasos que son exportados a otros lugares donde se requieren
como combustible.
En tanto que combustibles almacenados, los triacilgliceroles tienen dos ventajas significativas
sobre los polisacáridos tales como el glucógeno o el almidón. En primer lugar, los átomos de
carbono de los ácidos grasos están más reducidos que los de los azucares por lo que la
oxidación de los triacilgliceroles proporciona más del doble de energía, gramo por gramo, que
la de los glúcidos. En segundo lugar, como los triacilgliceroles son hidrofóbicos y, por
consiguiente, no hidratados, el organismo que transporta combustible en forma de grasa no ha
de transportar el peso adicional del agua de hidratación asociada con los polisacáridos
almacenados (2g por gramo de polisacárido). El tejido graso en los humanos (formado
principalmente por adipocitos), se encuentra debajo de la
piel, en la cavidad abdominal y en las glándulas
mamarias. Las personas moderadamente obesas pueden
tener de 15 a 20 Kg de triacilgliceroles depositados en sus
adipocitos, lo que es suficiente para cubrir sus
necesidades energéticas durante varios meses solamente
con sus depósitos de grasa. Por el contrario, el cuero
humano no puede almacenar ni las necesidades
energéticas de un día en forma de glucógeno. Los
glúcidos tales como la glucosa y el glucógeno ofrecen
ciertas ventajas como fuentes rápidas de energía
metabólica siendo una de ellas su fácil solubilidad en
agua. En algunos animales, los triacilglicéridos
almacenados debajo de la piel no sólo sirven como
almacenes de energía sino como aislamiento contra las
bajas temperaturas. Las focas, las morsas, los pingüinos y
otros animales polares de sangre caliente están
ampliamente protegidos con triacilgliceroles. En los
animales hibernantes ( como los osos ) las enormes
reservas de grasa acumuladas antes de la hibernación
también tienen una doble función: aislamiento y depósito
de energía. La baja densidad de los triacilglicéridos
constituye la base de otra función notable de estos
compuestos. En los cachalotes, un almacén de triacilgliceroles y de ceras les permite a los
animales igualar la flotación de sus cuerpos con de la zona a su alrededor en las inmersiones
profundas en agua fría.
Figura 10-3: Glicerol y un triacilglicerol: el triacilglicerol mixto representado en la figura tiene
tres ácidos grasos diferentes unidos al armazón de glicerol. Cuando hay dos ácidos grasos
diferentes en C-1 y C-3 del glicerol, el C-2 se transforma en un centro quiral.
LA HIDROGENACIÓN PARCIAL DE LOS ACEITES DE COCINA PRODUCE ÁCIDOS
GRASOS
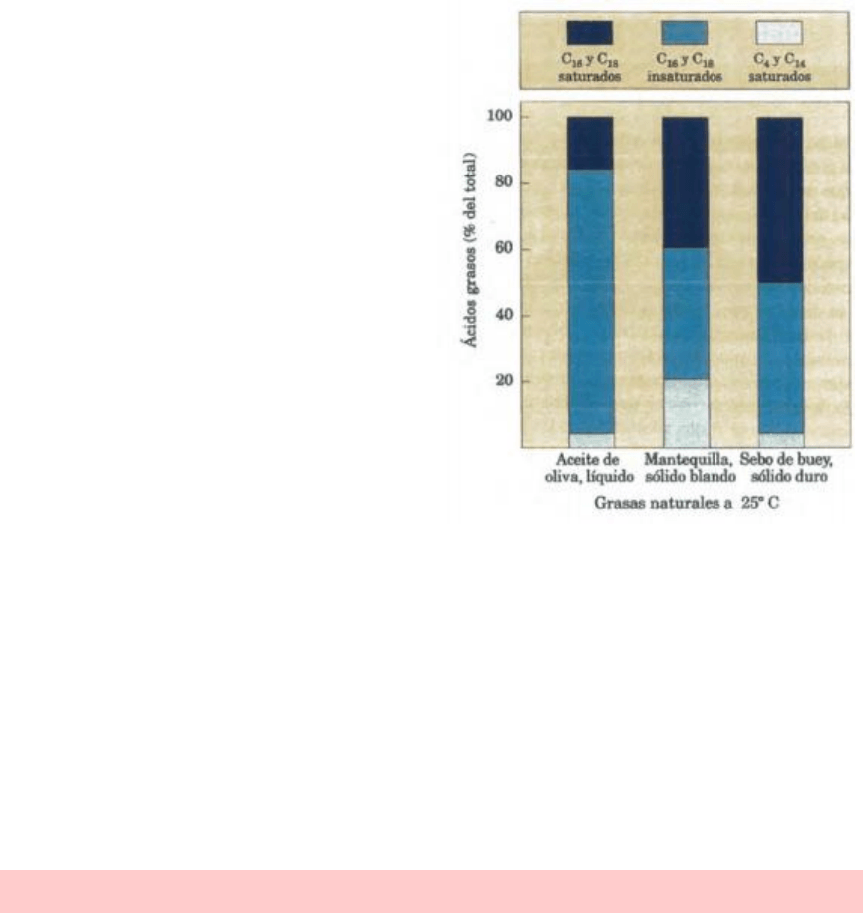
La mayoría de grasas naturales como los aceites vegetales, los productos lácteos y las grasas
animales son mezclas complejas de triacilgliceroles sencillos y mixtos. Estos últimos contienen
diversos ácidos grasos que difieren en la longitud de la cadena y grado de saturación.
Los aceites vegetales tales como el aceite de maíz y de oliva están compuestos
mayoritariamente por triacilgliceroles con ácidos grasos insaturados por lo que son líquidos a
temperatura ambiente. Los triacilgliceroles que sólo contienen ácidos grasos saturados, tales
como la triastearina, el componente principal de la grasa de buey, son sólidos blancos y grasos
a temperatura ambiente.
Cuando los alimento ricos en grasas se exponen demasiado tiempo al O del aire se pueden
estropear volviéndose rancios. El gusto y olor desagradables asociados con el ranciamiento
provienen de la rotura oxidativa de los dobles enlaces de ácidos grasos insaturados que
produce aldehídos y ácidos carboxílicos de cadena más corta y, por consiguiente, de mayor
volatilidad. Con el fin de mejorar las condiciones de caducidad de los aceites vegetales
utilizados para cocinar y con el fin de aumentar su estabilidad a las altas temperaturas de los
aceites vegetales usados para cocinar y con el fin de aumentar su estabilidad a las altas
temperaturas alcanzadas, los aceites vegetales comerciales se someten a hidrogenación
parcial. Este proceso convierte gran parte de
sus dobles enlaces cis en enlaces sencillos lo
que aumenta la temperatura de fusión de los
aceites, por lo que son casi sólidos a
temperatura ambiente. La hidrogenación
parcial tiene otro efecto, éste indeseable:
algunos dobles enlaces cis se convierten en
enlaces trans. Actualmente existen muchas
pruebas de que la ingestión con la dieta de
ácidos grasos trans (conocidos como grasas
trans) está en la base de una mayor
incidencia de enfermedades cardiovasculares,
por lo que evitando estas grasas en la dieta se
reduce el riesgo de enfermedad coronaria.
Los ácidos grasos trans de la dieta aumentan
la concentración de triacilgliceroles y de
colesterol ligado al LDL (“malo”) en la sangre
al tiempo que disminuyen la concentración
de colesterol ligado al HDL (“bueno”), cambios que por sí solos son suficientes para aumentar
el riesgo de enfermedad coronaria. Los ácidos grasos trans aún pueden tener más efectos
adversos. Parece que, por ejemplo, aumentan la respuesta inflamatoria corporal, lo cual es
otro riesgo de enfermedad cardíaca.
Los efectos perjudiciales de las grasas trans tienen lugar con ingestas de 2 a 7 g por día (20 a 60
kcal en una ingesta calórica diaria de 2000 Kcal).
la cadena de longitud corta e INSATURADA FAVORECE BAJOS PUNTOS DE FUSIÓN.
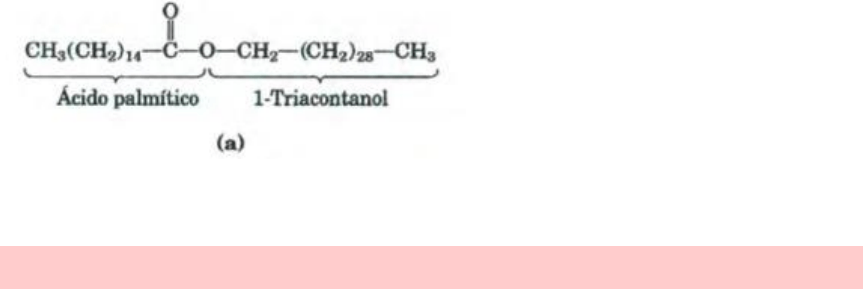
LAS CERAS
Las ceras sirven como almacenes de energía y como cubiertas impermeables al agua. Las ceras
biológicas son ésteres de ácidos grasos de cadena larga saturados e insaturados (C
14
a C
36
) con
alcoholes de cadena larga (C
16
a C
30
). Sus puntos de fusión (60 a 100°) son generalmente más
elevados que los de los triacilgliceroles. En plancton, constituidos por microorganismos
marinos flotantes de vida libre que se encuentran en la base de la cadena alimenticia para
animales marinos, las ceras son la forma de almacenamiento principal de combustible
metabólico.
Las ceras también realizan diversas funciones, que están relacionadas con sus propiedades
repelentes del agua y con su consistencia firme. Ciertas glándulas de la piel de los vertebrados
secretan ceras para proteger el pelo y la piel manteniéndose flexibles, lubricados e
impermeables.
Cera biológica: el triacontanilpalmitano,
componente mayoritario de la cera de
abeja, es un éster del ácido palmítico con
el alcohol triacontanol.
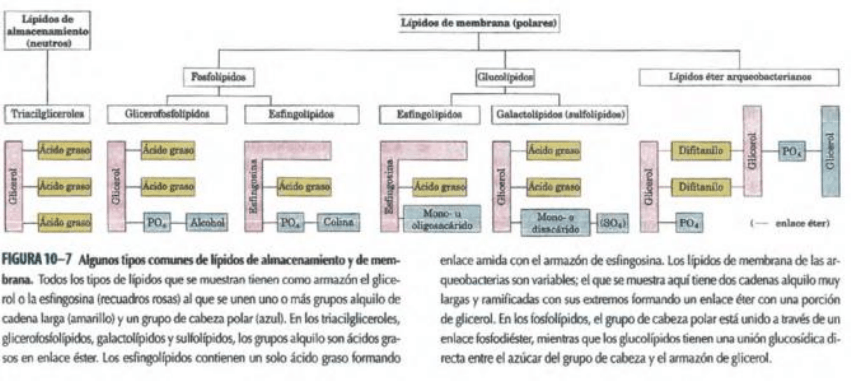
LÍPIDOS ESTRUCTURALES DE LAS MEMBRANAS
La característica arquitectónica central de las membranas biológicas es una doble capa lipídica
que constituye una barrera al paso de moléculas polares y de iones. Los lípidos de las
membranas son anfipáticos; un extremo de la molécula es hidrofóbico y el otro hidrofílico. Las
interacciones hidrofóbicas entre ellos y las hidrofílicas con el agua dirigen su
empaquetamiento hacia la formación de láminas llamadas bicapas membranosas. Cinco tipos
generales de lípidos de membrana son: glicerofosfolípidos, en los que las regiones hidrofóbicas
están compuestas por dos ácidos grasos unidos al glicerol; galactolípidos y sulfolípidos, que
también tienen dos ácidos grasos esterificados con el glicerol pero que carecen del fosfato
característico de los fosfolípidos, en los que las regiones hidrofóbicas están compuestas por
dos ácidos grasos unidos al glicerol; lípidos tetraéter de las arqueobacterias, en los que dos
cadenas alquílicas muy largas están unidas mediante enlace éter al glicerol de ambos
extremos; esfingolípidos, en los que se une un solo ácido graso a una amina grasa, la
esfingosina; y esteroles, que son compuestos que se caracterizan por tener un sistema rígido
de cuatro anillos hidrocarbonados fusionados.
Las partes de estos compuestos anfipáticos pueden ser muy sencillas, por ejemplo, un simple
grupo ---OH en un extremo del sistema anular de los esteroles, o pueden ser mucho más
complejas. Los glicerofosfolípidos y algunos esfingolípidos contienen un grupo polar que se
une a una porción hidrofóbica mediante un enlace fosfodiéster; son los fosfolípidos. Otros
esfingolípidos carecen de grupo fosfato, pero tienen un azúcar sencillo u oligosacáridos
complejos en sus extremos polares; son los glucolípidos. Dentro de estas clases de lípidos de
membrana existe una enorme diversidad debido a las diferentes combinación de “colas” de
ácidos grasos y “cabezas” polares.

ANEXO


GLÚCIDOS.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.