
Linfocito B
Estructura de las inmunoglobulinas:
Están formadas por dos cadenas pesadas
(H) y dos cadenas livianas (L), la región
aminoterminal es variable, en ella
encontramos porciones hipervariables
CDR1, CDR2 y CDR3 que constituyen el
PARATOPE que es la porción que se
conecta con el EPITOPE antigénico.
Existen 5 tipos de inmunoglobulinas:
• IgG, IgD, IgE y la Ig de membrana
BCR, son monomerica
• IgM es pentamerica
• IgA es dimerica
Ontogenia B
Empieza en la medula ósea con la liberación de factores de crecimiento y
diferenciación (IL-7, CXCL12 y SCF).
La recombinación somática ocurre de forma similar a los linfocitos t donde en
las células germinales hay recombinación de los genes VDJ con acción de las
RAG1 y2 para formar las porciones variables de las cadenas H (VDJ) y L(DJ).
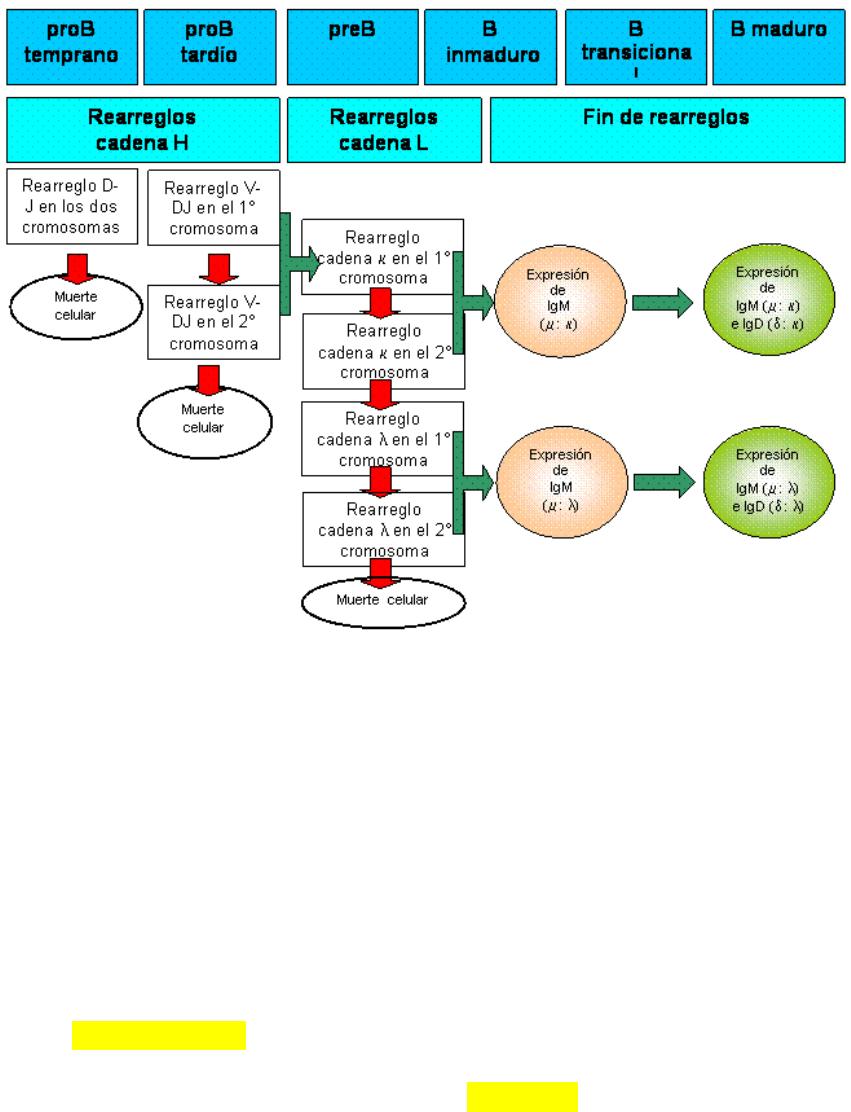
Estadios del LB
• PRO-B: reordenamiento de la cadena H, expresión de SSR (secuencia
de señales de recombinación), si el reordenamiento no es exitoso ocurre
apoptosis al final expresa una cena liviana sustituta.
o PRO-B temprano: asociación de fragmentos D-J
o PRO-B tardío: se une V a D-J, y ocurre exclusión alélica para
garantizar la formación de una sola cadena de los dos alelos del
genoma.
• PRE-B: empieza el reordenamiento de la cadena L, también gobernado
por la exclusión alélica. Y se evalúa se puede reconocer señales por las
células estromales de la medula.
• Estadio B inmaduro: una vez las cadenas H y L reordenadas se asocian
formando IgM (BCR) y empiezan los mecanismos de especificidad del
BCR
• Estadio B maduro: expresan IgM y IgD, como consecuencia de
SPLICING ALTERNATIVO (la porción variable es la misma, el splicing
ocurre en la porción constante de la cadena H) TODA Ig DE UN TB
RECONOCEN EL MISMO ANTIGENO.
Mecanismo de tolerancia central B:
Si el BCR no reconoce el antígeno presentado por las células estromales
emigran de la medula ósea para continuar su desarrollo.
Si la señal es de baja intensidad, se produce anergia e mueren en la periferia.
Si la señal es de alta intensidad se produce apoptosis o deleción.
Mecanismo de tolerancia periférica
Cuando salen de la medula ósea son denominado linfocitos b transicionales,
van al bazo donde son presentados a antígenos:
• Si no reconocen señal pasan de BTr1 A BTr2, con participación de la
citoquina BAFF.
• Los BTr2 evolucionan a LB maduros se reciben señal por su BCR.
Los linfocitos B maduros salen a la circulación presentado IgM IgD.

Ciclo de vida
Los linfocitos migran hacia el folículo gracias a la excreción de la citoquina
CXCL13.
Activación linfocitos B
Colaboración T-B: Es necesaria para la activación de los LB2
1. LB reconoce un epitope por su BCR y procesa el antígeno por la CMH2
vía exógena
2. El LT CD4fh reconoce el CMH2-peptido presentado por el LB. Para que
ocurra la activación el péptido presentado por LB debe ser del mismo
antígeno que el presentado al LT por la célula dendrítica.
3. El LB activado se diferencia a plasmocito y secreta Ac especifico.
*Haptenos: son moléculas que son reconocidas por BCR pero no generan
respuesta inmune (no generan péptido), para generar colaboración T-B de
debe conjugar al hapteno con un péptido, ese complejo puede ser presentado a
LT y así se genera anticuerpos para el hapteno. Ej.: vacunas conjugadas contra
polisacáridos bacterianos.

Diferenciación cd4 a TFH:
Requiere que el TCR reconozca con alta afinidad el CMH2 y las citoquinas IL-6
y IL-12. Lo LT empieza a secretar IL-21 y 19 que es esencial para la expresión
de CXCR5.
El aumento sostenido de CXCR5 junto con la disminución de CCR7 hace que
el TFH migre hacia el folículo primario atraído por CXCR13.
LB: el linfocito b naive atraviesan las VEH por expresaren CCR7 y migran al
folículo primario por expresaren CXCR5, en el folículo reciben señal de los
macrófagos subscapulares (1er señal de activación) lo que hace down
regulation con la molécula de CXCR5 lo que permite que el LB migre hacia el
borde del folículo (mayor expresión de CCR7) donde encuentra con el TFH.
ENCUENTRO: el encuentro desencadena la proliferación de ambos linfocitos y
forman el foco primario donde los LB se diferencian en:
• Plasmoblastos y luego en plasmocitos(IL-6) y quedan en la medula del
ganglio linfático secretando Ig, principalmente IgM (baja afinidad), que
son los primeros Ac de la respuesta humoral.
• Migran al centro del folículo primario y proliferan de forma muy activa
formando el folículo secundario o centro germinal, pasan a se llamar de
centroblastos y sufren hipermutación somática.
o Hipermutación somática: ocurre
en la región variable de ambas
cadenas CAMBIA EL
PARATOPE, o sea se forma la
misma Ig con diferentes grados
de afinidad.
o El centrocito migra hacia la
zona clara del foliculo donde tienen que reconocer el antigeno
presentado por la celula dendritica folicular(no es una celula
presentadora de antigeno, retienen el antigo nativo en su membrana), si
tienen gran afinidad por el antigeno o procesan por la via exogena
e lo presenta al TFH, esa segunda interacción T-B permite la
señal de supervivencia (Bcl-xl) al centrocito. Aquellos que tienen
poca afinidad no comletan ese proceso y sufren apoptosis.
o Cambio de isotipo de la inmunoglobulina: es necesario la segunda
interacción T-B, ocurre por recombinacion de su ADN. El LB pasa
expresar IgG, IgA, IgE (son mas especificas) y disminuye la
expresion de IgM. Al salir del centro germinativo se van a llamar
de linfocitos B de memoria.

Los linfocitos b de
memoria expresan
BCR, están en
circulación y en los
OLS, además producen
IgM, IgG, IgA y IgE.
Necesitan de PAX-5
para su diferenciación
Los plasmocitos
producen IgM, IgG, IgA
y IgE, no circulan, están
en la medula ósea o en
OLS, necesitan Blimp-1
y XBP-1 para su
diferenciación.
plasmocitos de vida corta: se ubican en la MO(B2) y en el bazo(BZM), secretan
anticuerpo por algunos días o semanas: principalmente IgM
plasmocitos de vida larga: se ubican en MO, secretan anticuerpos por años. Se
encuentran en un nicho de supervivencia que atrae a los plasmocitos por la
producción de CXCL12 que los atrae por quimiotaxis y es un factor de
supervivencia. Los plasmocitos nuevos desplazan a plasmocitos más viejos e
la medula ósea, esos mueren poco tiempo después.
Respuesta humoral primaria: Se produce ante el primer contacto con un
antígeno determinado. Se principalmente por la producción de IgM.
Respuesta humoral secundaria: cuando hay reinfección la respuesta es más
rápida y especifica con mayor participación de IgG que IgM, más intensa

Linfocitos B1
Se activan sin interacción T-B, inician su vida en el hígado fetal y a los 2 años
completan su maduración (por eso se da la vacuna conjugada a los niños menores de 2
años, ellos solo tienen LB1 funcional que necesita péptido), o sea expresan CD21, se
localizan mayoritariamente en la cavidad peritoneal y pleural.
Producen anticuerpos naturales y son los más importantes para el combate de
bacterias capsulas por su capacidad de reconocer antígenos no proteicos.
Linfocitos BZM
Zona marginal del bazo, secreta mayoritariamente IgM y en menor medida IgG,
también es relevante en la respuesta inmune contra bacterias encapsuladas, es
independiente de TFH

Linfocitos B reguladores
Inhiben la actividad de células dendríticas mieloides, monocitos, linfocitos t
citotóxicos, células TH1, TH17 Y TFH a través de la secreción de IL10, TGF,
IL35.
Promueve la actividad de células T regulatorias (FoxP3+ y Tr1) a través de
IL10 e IL35.
Anticuerpos
• IgG/D/E- son monomerica
• IgM es pentamerica
• IgA es dimerica en las mucosas
y monomerica en el suero
• Ig de membrana (parte del bcr)
siempre monomerica
• IgE es la única con 4 dominios
constantes
Función de los anticuerpos
1. Neutralización: de toxinas, son bloqueadas cuando entran en contacto
con los AC y luego ingeridas por los macrófagos.
2. Opsonización: una bacteria es opsonizada y luego fagocitada por el
sistema mononuclear fagocítico.
3. Activación del complemento: vía clásica del complejo inmune.
IgD- no es secretada por ningún plasmocito, se desconoce su función como Ig
secretoria
IgM sérica- tiene gran capacidad de ser unir a C1q por lo que activa la cascada
de complemento y es neutralizante. Es el primero Ig producido en respuesta
inmune es polirreactivo y de baja especificidad. Actúa mayoritariamente en los
vasos, pero en la inflamación con la mayor permeabilidad vascular llega a los
tejidos adyacentes.
IgE- es el de menor concentración en la sangre porque se asocia a los FcR
(receptores de alta afinidad) de los mastocitos, en personas con alergias el
entrecruzamiento de los FcR a consecuencia del reconocimiento de antígenos
específicos permite la activación del mastocito con la liberación de la histamina
de sus gránulos. Ademáslas IgE también se une a RFcR2 (baja afinidad) de
los eosinofilos y pueden inducir la citotoxicidad CCDA mediadas por
eosinofilos, monocitos y plaquetas.

IgG- mediante unión con RFc ( macrófagos…) media las funciones de
fagocitosis CCDA y liberación de mediadores inflamatorios ROL CRITICO EN
INFECCIONES VIRALES.
Receptor neonatal para IgG (FcRn) es el encargado de transportar la IgG de la
sangre materna hacia la sangre fetal a través de las células del
sinciotrofoblasto, él se une a la IgG a pH acido. La IgG de la sangre materna es
endocitada por la célula del sinciotrofoblasto, se une al receptor en el
endosoma, y en sangre fetal se disocia del receptor por el pH neutro de la
sangre. Además, cumple papel en la vida neonatal por la presencia de FcRn en
el lumen intestinal del neonato lo que permite la absorción de las IgG de la
leche materna.
Activa el sistema de complemento
IgA- neutraliza patógenos en el lumen, intracelular de virus y en la lámina
propia si los patógenos atraviesan el epitelio. Inhibe la adhesión de patógenos
al epitelio facilitando la acción de fuerzas mecánicas para la eliminación de
microorganismos.

linfocito B.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.