
634
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
testino delgado. Termina en la válvula ileocecal, la
unión del íleon distal y el ciego (lámina 61, pág. 668).
El intestino delgado es el sitio principal para la digestión
de alimentos y para la absorción de los productos de la di-
gestión.
El quimo del estómago ingresa en el duodeno, hacia donde
también se envían las enzimas del páncreas y la secreción bi-
liar hepática para continuar con el proceso de solubilización
y de digestión. Las enzimas, en particular las disacaridasas y
las dipeptidasas, también se localizan en el glucocáliz de las
microvellosidades de los
enterocitos, que son las células
absortivas intestinales
. Estas enzimas contribuyen al pro-
ceso digestivo al completar la degradación de la mayoría de
los sacáridos y proteínas en monosacáridos y aminoácidos,
que después se absorben (cuadro 17-4). El agua y los elec-
trolitos que llegan al intestino delgado con el quimo y las
secreciones pancreáticas y hepáticas también se reabsorben en
el intestino delgado, en particular en la porción distal.
Los pliegues circulares, las vellosidades y las microvellosi-
dades incrementan la extensión de la superfi cie absortiva
del intestino delgado.
La super cie absortiva del intestino delgado está ampli cada
por el tejido y las especializaciones celulares de la mucosa y de
la submucosa.
•
Los pliegues circulares, también conocidos como vál-
vulas de Kerckring, son repliegues transversales permanen-
tes que contienen un centro de submucosa. Cada pliegue
circular rodea entre la mitad y las dos terceras partes de la
circunferencia de la luz ( g. 17-17). Los pliegues comien-
zan a aparecer a unos 5 cm a 6 cm después del píloro. Son
más abundantes en la porción distal del duodeno y en el
comienzo del yeyuno y su tamaño y frecuencia se reducen
desde la mitad del íleon.
•
Las vellosidades son evaginaciones digitiformes o foliá-
ceas singulares de la mucosa, que se extienden dentro de la
luz intestinal en una distancia de 0,5 mm a 1,5 mm desde
la super cie mucosa teórica ( g. 17-18). Las vellosidades
cubren por completo la super cie del intestino delgado,
lo que le imparte un aspecto aterciopelado cuando se exa-
mina a simple vista.
•
Las microvellosidades de los enterocitos proporcionan
la principal ampliación de la super cie luminal. Cada cé-
lula posee varios miles de microvellosidades muy juntas
que son visibles con el microscopio óptico y le imparten
a la región apical de la célula un aspecto estriado, el así
llamado
borde estriado. Los enterocitos y sus microve-
llosidades se describen más adelante.
Las vellosidades y las glándulas intestinales, junto con la
lámina propia, el GALT asociado y la muscular de la mu-
cosa, constituyen los rasgos esenciales de la mucosa del
intestino delgado.
Las vellosidades, como se mencionó, son proyecciones o
evaginaciones de la mucosa. Están compuestas por un centro
de tejido conjuntivo laxo cubierto por un epitelio cilíndrico
simple. El centro de la vellosidad es una extensión de la lá-
mina propia, que contiene una abundancia de broblastos,
células musculares lisas, linfocitos, plasmocitos, eosinó los,
macrófagos y una red de capilares sanguíneos fenestrados,
ción tiende a ser en un patrón circular. Finos haces de células
musculares lisas, se extienden hacia la super cie a través de
la lámina propia desde la capa interna de la muscular de la
mucosa. Se cree que estas células musculares lisas en la lámina
propia contribuyen a la expulsión de las secreciones de las
glándulas gástricas.
Submucosa gástrica
La submucosa está compuesta por tejido conjuntivo denso
que contiene cantidades variables de tejido adiposo y vasos
sanguíneos, así como bras nerviosas y células ganglionares
que componen el
plexo submucoso (plexo de Meiss-
ner). Este último inerva los vasos de la submucosa y el mús-
culo liso de la muscular de la mucosa.
Muscular externa gástrica
La muscular externa del estómago tradicionalmente se
describe como compuesta por una capa longitudinal externa,
una capa circular media y una capa oblicua interna. Esta des-
cripción puede ser engañosa, ya que discernir las capas bien
de nidas no resulta fácil. Al igual que con otros órganos hue-
cos esferoidales (p. ej., la vesícula biliar, la vejiga urinaria y el
útero), el músculo liso de la muscular externa del estómago
está orientado en una forma más aleatoria que lo que el tér-
mino capa implica. Además, la capa longitudinal está ausente
en gran parte de las super cies gástricas anterior y posterior
y la capa circular está poco desarrollada en la región perieso-
fágica. Desde el punto de vista funcional, la disposición de
las capas musculares es importante, ya que está relacionada
con su papel en el mezclado del quimo durante el proceso
digestivo, así como con su capacidad para desplazar el con-
tenido con digestión parcial hacia el intestino delgado. Entre
las capas musculares se encuentran grupos de células ganglio-
nares y haces de bras nerviosas amielínicas. En conjunto for-
man el
plexo mientérico (de Auerbach), que inerva las
capas musculares.
Serosa gástrica
La serosa del estómago es como la que ya se describió para el
tubo digestivo en general. Es continua con el peritoneo parie-
tal de la cavidad abdominal a través del omento mayor y con
el peritoneo visceral del hígado a través del omento menor.
Aparte de eso, no exhibe características especiales.
INTESTINO DELGADO
El intestino delgado es el componente más largo del tubo
digestivo; mide más de 6 m y se divide en tres porciones
anatómicas:
•
El duodeno (⬃25 cm de longitud) es la primera porción
y la más corta y ancha del intestino delgado. Comienza a
la altura del píloro del estómago y termina en el ángulo
duodenoyeyunal (lámina 59, pág. 664).
•
El yeyuno (⬃2,5 m de longitud) comienza en el ángulo
duodenoyeyunal y constituye las dos quintas partes
proximales del intestino delgado. Cambia sus característi-
cas morfológicas en forma gradual hasta convertirse en el
íleon (lámina 60, pág. 666).
•
El íleon (⬃3,5 m de longitud) es una continuación del
yeyuno y constituye las tres quintas partes distales del in-
Pawlina_17_3R_Editado.indd 634Pawlina_17_3R_Editado.indd 634 15/07/15 12:4415/07/15 12:44
635
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
CUADRO 17-4
Consideraciones funcionales: funciones digestivas y
absortivas de los enterocitos
La membrana plasmática de las microvellosidades del ente-
rocito participa tanto en la digestión como en la absorción.
Las enzimas digestivas están ancladas en la membrana
plasmática y sus grupos funcionales se extienden hacia fuera
para formar el glucocáliz. Esta disposición acerca los produc-
tos fi nales de la digestión a su sitio de absorción. Entre las
enzimas se encuentran las peptidasas y las disacaridasas. La
membrana plasmática de las microvellosidades apicales tam-
bién contienen la enzima enteropeptidasa (enterocinasa),
la cual reviste particular importancia en el duodeno, donde
convierte tripsinógeno en tripsina. La tripsina puede continuar,
entonces, la conversión adicional de tripsinógeno en tripsina
y la tripsina convierte otros varios cimógenos pancreáticos en
sus enzimas activas (fi g. C17-4.1). en los párrafos que siguen,
se reseña digestión y la absorción de los tres tipos principales
de sustancias nutritivas.
La digestión fi nal de los hidratos de carbono la realizan
las enzimas unidas a las microvellosidades de los enterocitos
(fi g. C17-4.2). La galactosa, la glucosa y la fructosa son ab-
sorbidas directamente por los capilares venosos y son trans-
portadas al hígado a través de los vasos del sistema hepático
portal. Algunos infantes y un gran porcentaje de los adultos
no pueden tolerar la leche ni los productos lácteos no fer-
mentada debido a la ausencia de lactasa, la disacaridasa que
divide la lactosa en galactosa y glucosa. Si estas personas
ingieren leche, sufren distensión abdominal por gas producido
por la digestión bacteriana de la lactosa no procesada y pade-
cen diarrea. El trastorno se alivia por completo si se elimina
la lactosa (disácarido lácteo) de la dieta. En algunas personas,
la intolerancia a la leche también se puede aliviar en forma
parcial o total mediante el uso de productos lácteos reducidos
en lactosa o de tabletas de lactasa (enzima que digiere la
lactosa), que se expenden en el comercio como fármacos de
venta libre.
Los triglicéridos se degradan a glicerol, monoacilglicero-
les y ácidos grasos de cadenas corta, mediana y larga. Estas
sustancias son emulsionadas por las sales biliares y se in-
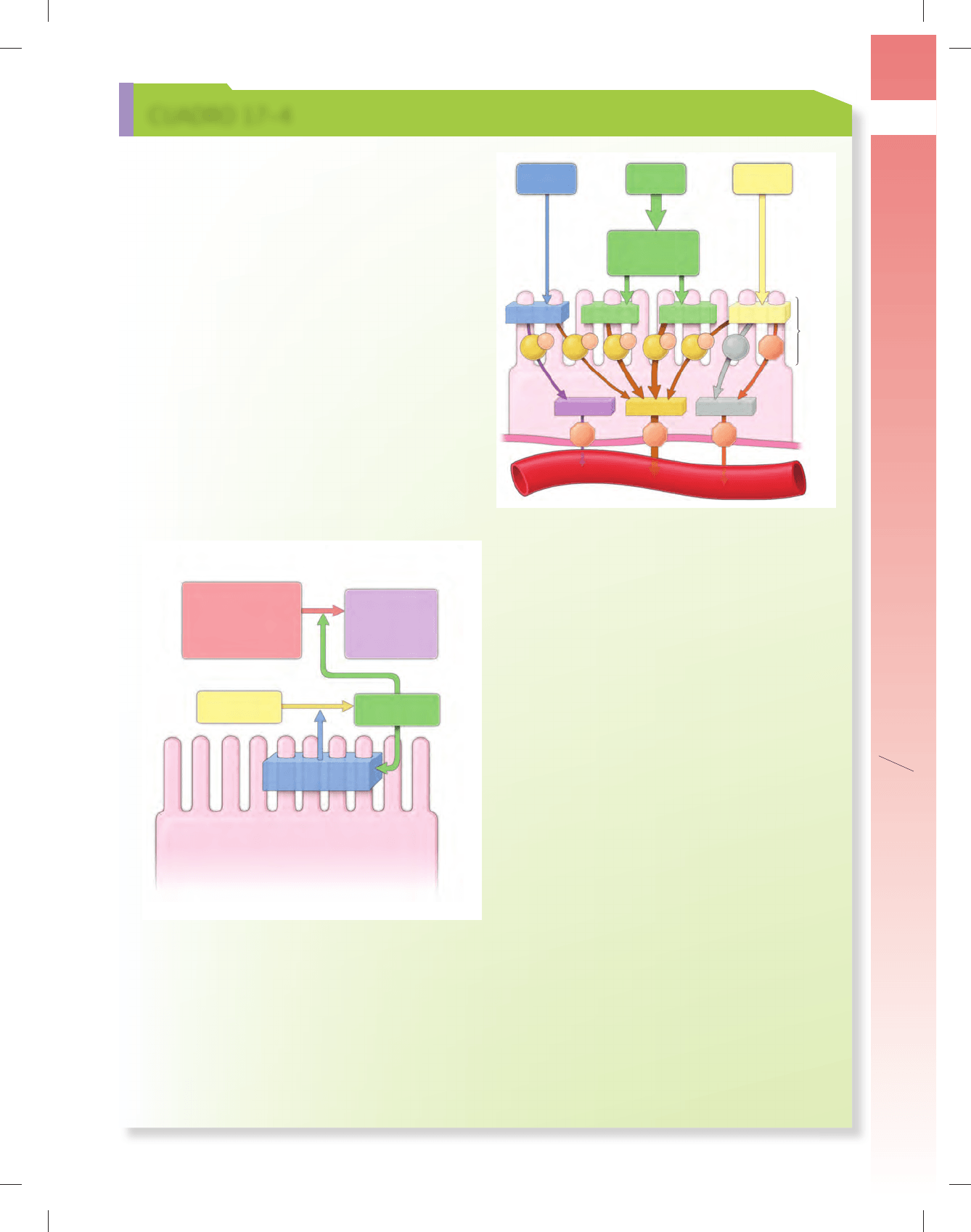
FIGURA C17-4-1 ▲ Diagrama que ilustra los fenóme-
nos en la activación de las enzimas proteolíticas del páncreas.
La mayoría de las enzimas pancreáticas (proteasas) se secretan como
proenzimas inactivas. Su activación se desencadena por la llegada del
quimo al duodeno. Esto estimula las células mucosas para que liberen y
activen la enterocinasa (en azul) dentro del glucocáliz. La enterocinasa
activa el tripsinógeno y lo convierte en su forma activa, la tripsina (en
verde). A su vez, la tripsina activa otras proenzimas pancreáticas (en rojo)
en sus formas activas (en púrpura). Las proteasas activas hidrolizan en-
laces peptídicos de proteínas y polipéptidos y los reducen a péptidos
pequeños y aminoácidos.
Cimógenos pancreáticos
(proenzimas inactivas)
Cimógeno pancreático
Enzimas activas
Enzima activa
Enterocinasa
Tripsina
Tripsinógeno
Quimiotripsinógeno
Proelastasa
Procarboxipeptidasa A
Procarboxipeptidasa B
Fosfofolipasa A
2
Tripsina
Quimiotripsina
Elastasa
Carboxipeptidasa A
Carboxipeptidasa B
Fosfofolipasa A
2
Enterocito
(continúa en página 636)
Sacarosa
Sacarosa
Maltasa
FructosaGlucosaGalactosa
Capilar
G
Na
⫹
G
Na
⫹
G
Na
⫹
GG
F
G
2
G
2
G
2
G
2
Na
⫹
Na
⫹
Lactasa ␣-dextrinasa
Lactosa
Glucógeno
Almidón
Amilasas
salivales
y pancreáticas
␣-dextrinas
Maltotriosas
Maltosas
Chapa
estriada
~70%
~25%~5%
Enterocito
FIGURA C17-4.2 ▲ Diagrama que ilustra la digestión y
la absorción de hidratos de carbono por el enterocito. Los hi-
dratos de carbono llegan al tubo digestivo como monosacáridos (p.
ej., glucosa, fructosa y galactosa), disacáridos (p. ej., sacarosa, lactosa y
maltosa) y polisacáridos (p. ej., glucógeno y almidón). Las enzimas que
participan en la digestión de hidratos de carbono se clasifi can como
amilasas salivales y pancreáticas. La digestión adicional se realiza en
la chapa estriada de los enterocitos por la acción de enzimas que de-
gradan oligosacáridos y polisacáridos en tres monosacáridos básicos
(glucosa, galactosa y fructosa). La glucosa y la galactosa son absorbidas
por el enterocito mediante un transporte activo que utiliza transporta-
dores de glucosa dependientes de Na
⫹
(SGLT1). Estos transportadores
se localizan en la membrana celular apical (círculos pardos con rótulos G
y Na
⫹
). La fructosa se introduce en la célula a través del transporte faci-
litado independiente de Na
⫹
que utiliza GLUT5 (círculo gris con rótulo F)
y transportadores de glucosa GLUT2 (octágono naranja con rótulo G2).
Los tres monosacáridos absorbidos entonces atraviesan la membrana
basal del enterocito, para lo que utilizan transportadores de glucosa
GLUT2, y pasan a los capilares subyacentes de la circulación portal que
los conducen hacia su destino fi nal en el hígado.
Pawlina_17_3R_Editado.indd 635Pawlina_17_3R_Editado.indd 635 15/07/15 12:4415/07/15 12:44

636
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
CUADRO 17-4
Consideraciones funcionales: funciones digestivas y
absortivas de los enterocitos
(cont.)
troducen en la región apical del enterocito. Aquí, el glicerol y
los ácidos grasos de cadena larga se resintetizan para formar
triglicéridos. Los triglicéridos resintetizados aparecen primero
en las vesículas apicales del REL (v. fi g. 17-21), después en el
aparato de Golgi (donde se convierten en quilomicrones, pe-
queñas gotas de grasa neutra) y por último en vesículas que
transportan los quilomicrones hacia el espacio intercelular. En
lugar de ser absorbidos directamente por los capilares veno-
sos, los quilomicrones se alejan del intestino a través de los
vasos linfáticos (quilíferos) que penetran en cada vellosidad.
La linfa con quilomicrones abundantes entonces drena en
el conducto torácico, que desemboca en el sistema venoso
sanguíneo. Cuando entran en la circulación sanguínea, los qui-
lomicrones se desintegran con rapidez y sus lípidos constitu-
yentes son utilizados en todo el cuerpo. Los ácidos grados de
cadenas corta y mediana y el glicerol atraviesan la membrana
celular apical y entran y salen del enterocito exclusivamente
a través de capilares tributarios de la vena porta que llega al
hígado.
La digestión y absorción de proteínas se ilustran en la fi -
gura C17-4.3. Los principales productos fi nales de la digestión
proteica son los aminoácidos (cerca del 30 %) y los oligopépti-
dos (cerca del 70 %), que son absorbidos por los enterocitos.
El mecanismo de absorción de aminoácidos es conceptual-
mente idéntico al de los hidratos de carbono. La membrana
plasmática apical de los enterocitos contiene al menos cuatro
cotransportadores de aminoácidos dependientes de Na
+
. Los
dipéptidos y los tripéptidos son transportados a través de la
membrana apical hacia el citoplasma celular por el cotranspor-
tador oligopéptido-H
+
(PepT1). La mayoría de los dipéptidos y
tripéptidos son degradados entonces por las peptidasas cito-
plasmáticas a aminoácidos libres, los que posteriormente son
transportados a través de la membrana basal (sin la necesidad
de un cotransportador) hacia los capilares subyacentes de la
circulación portal. En un trastorno de absorción de aminoáci-
dos (enfermedad de Hartnup), aparecen aminoácidos libres en
la sangre cuando a los pacientes se les administra dipéptidos
pero no cuando reciben aminoácidos libres. Esto sustenta la
conclusión de que los dipéptidos de ciertos aminoácidos se
absorben a través del cotransportador PepT1, que participa en
mecanismos diferentes de los utilizados por los aminoácidos
libres.
FIGURA C17-4.3 ▲ Diagrama que ilustra la digestión y la
absorción de proteínas por el enterocito. Las proteínas que ingre-
san en el tubo digestivo son digeridas completamente hasta aminoá-
cidos libres (aa) y pequeños fragmentos de dipéptidos y tripéptidos.
La digestión proteica comienza en el estómago con la pepsina, la cual
hidroliza proteínas en polipéptidos grandes. La siguiente etapa ocurre
en el intestino delgado por la acción de las enzimas proteolíticas pan-
creáticas. El proceso de activación se ilustra en la fi gura C17-4.1. Los
aminoácidos libres son transportados por cuatro cotransportadores
de aminoácidos Na
⫹
diferentes. Los dipéptidos y los tripéptidos son
transportados a través de la membrana apical hacia la célula por los
cotransportadores de oligopéptido H
⫹
(PepT1). La mayor parte de los
dipéptidos y tripéptidos son degradados por peptidasas citoplasmáti-
cas, y los aminoácidos libres son transportados a través de la membrana
basal hacia los capilares subyacentes de la circulación portal.
Aminoácidos libres
Neutros Básicos
Ácidos
Na
⫹
H
⫹
Na
⫹
Na
⫹
Na
⫹
Aminoácidos
Capilar
Proteínas
Pepsina
Proteasas
pancreáticas
Oligopéptidos
2-3 aa
Polipéptidos
grandes
Oligopéptidos
2-3 aa
Transporta-
dores de
polipéptidos
Amino-
ácido
neutro
Transportador
de aminoácidos
Amino-
ácido
básico
Amino-
ácido
ácido
Iminoácido/
glicina
Amino-
ácido
neutro
Amino-
ácido
básico
Amino-
ácido
ácido
Iminoácido/
glicina
Peptidasa citoplasmática
Enterocito
Oligopéptido
cotransporta-
dorPepT1
Chapa
estriada
~75%
~25%
Imino y
glicina
a la altura de la base de la vellosidad (v. g. 17-18). Las glán-
dulas están compuestas por un epitelio cilíndrico simple que
es continuo con el epitelio de las vellosidades.
Al igual que en el estómago, la lámina propia rodea las
glándulas intestinales y contiene abundantes células del sis-
tema inmunitario (linfocitos, plasmocitos, mastocitos, ma-
crófagos y eosinó los), en particular en las vellosidades. La
lámina propia también contiene muchos nódulos de
tejido linfático que son uno de los componentes principales
del GALT. Los nódulos son particularmente grandes y abun-
dantes en el íleon, donde se localizan de manera preferencial
en el lado del intestino opuesto a la jación mesentérica, es
decir, en el borde antimesentérico del intestino ( g. 17-20).
Estas aglomeraciones nodulares se conocen como
conglo-
ubicados justo debajo de la lámina epitelial basal. Además,
la lámina propia de la vellosidad contiene un capilar linfático
central de fondo ciego, el
vaso quilífero central ( g. 17-19
y lámina 60, pág. 666). Las células musculares lisas derivadas
de la muscular de la mucosa se extienden hacia la vellosidad
y acompañan al vaso quilífero. Estas células musculares lisas
podrían ser la causa de la contracción y el acortamiento in-
termitente de las vellosidades, una acción que impulsaría la
linfa desde el vaso quilífero hacia la red de vasos linfáticos que
rodean a la muscular de la mucosa.
Las
glándulas intestinales o criptas de Lieberkühn,
son estructuras simples tubulares que se extienden desde la
muscular de la mucosa a través del espesor de la lámina pro-
pia, donde desembocan en la super cie luminal del intestino,
Pawlina_17_3R_Editado.indd 636Pawlina_17_3R_Editado.indd 636 15/07/15 12:4415/07/15 12:44
637
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
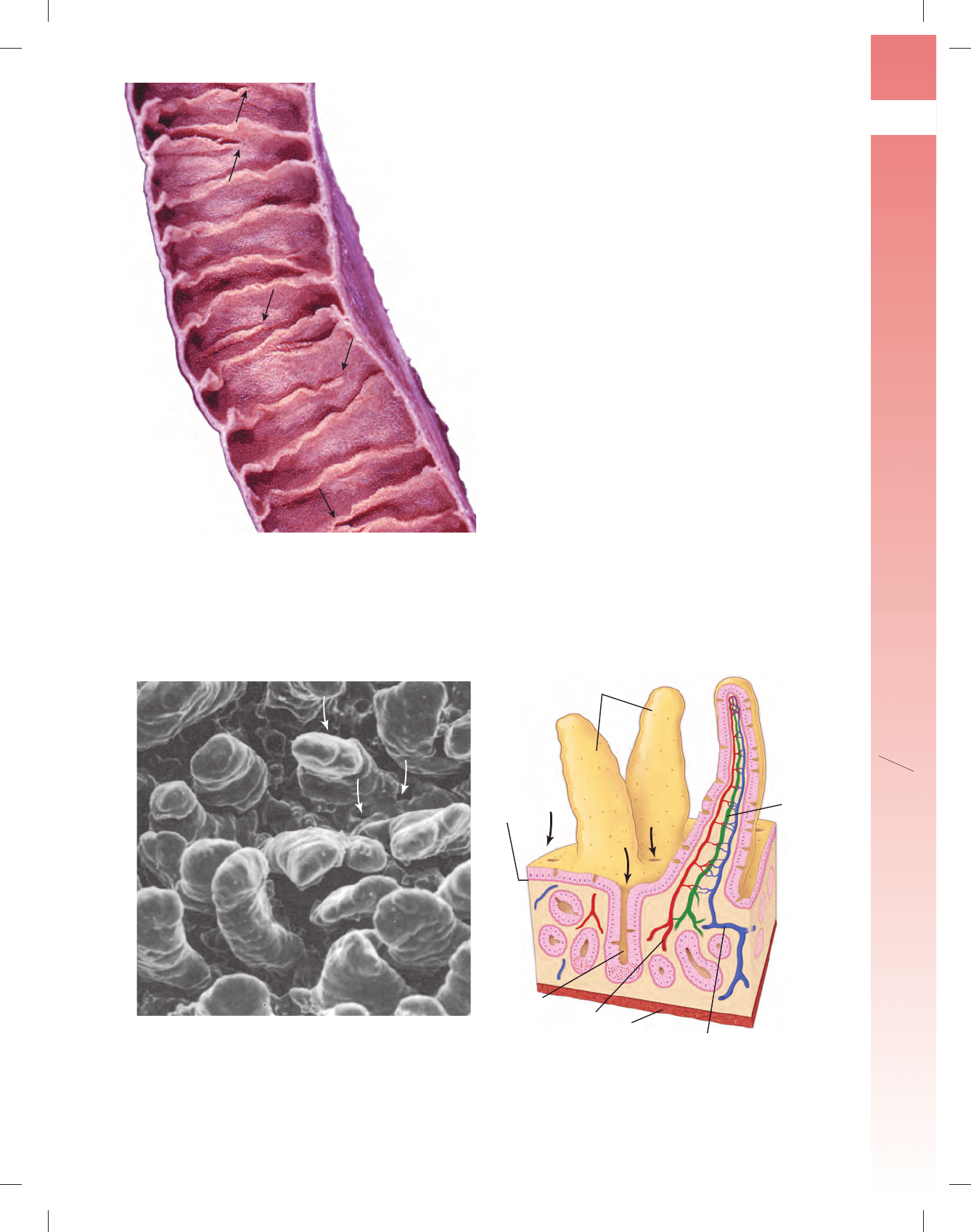
FIGURA 17-17 ▲ Fotografía de la superfi cie mucosa del in-
testino delgado. Esta fotografía de un segmento de un yeyuno humano
muestra la superfi cie mucosa. Los pliegues circulares (válvulas conniven-
tes) aparecen como una serie de crestas orientadas en forma transversal
que se extienden parcialmente alrededor de la luz. En consecuencia, algu-
nos pliegues circulares parecen terminar (o comenzar) en varios sitios a lo
largo de la superfi cie luminal (fl echas). Toda la mucosa presenta un aspecto
aterciopelado debido a la presencia de vellosidades.
a
b
Vellosidades intestinales
Membrana
basal
Glándula
intestinal
Arteria
Vena
Muscular de
la mucosa
Vaso
quilífero
central
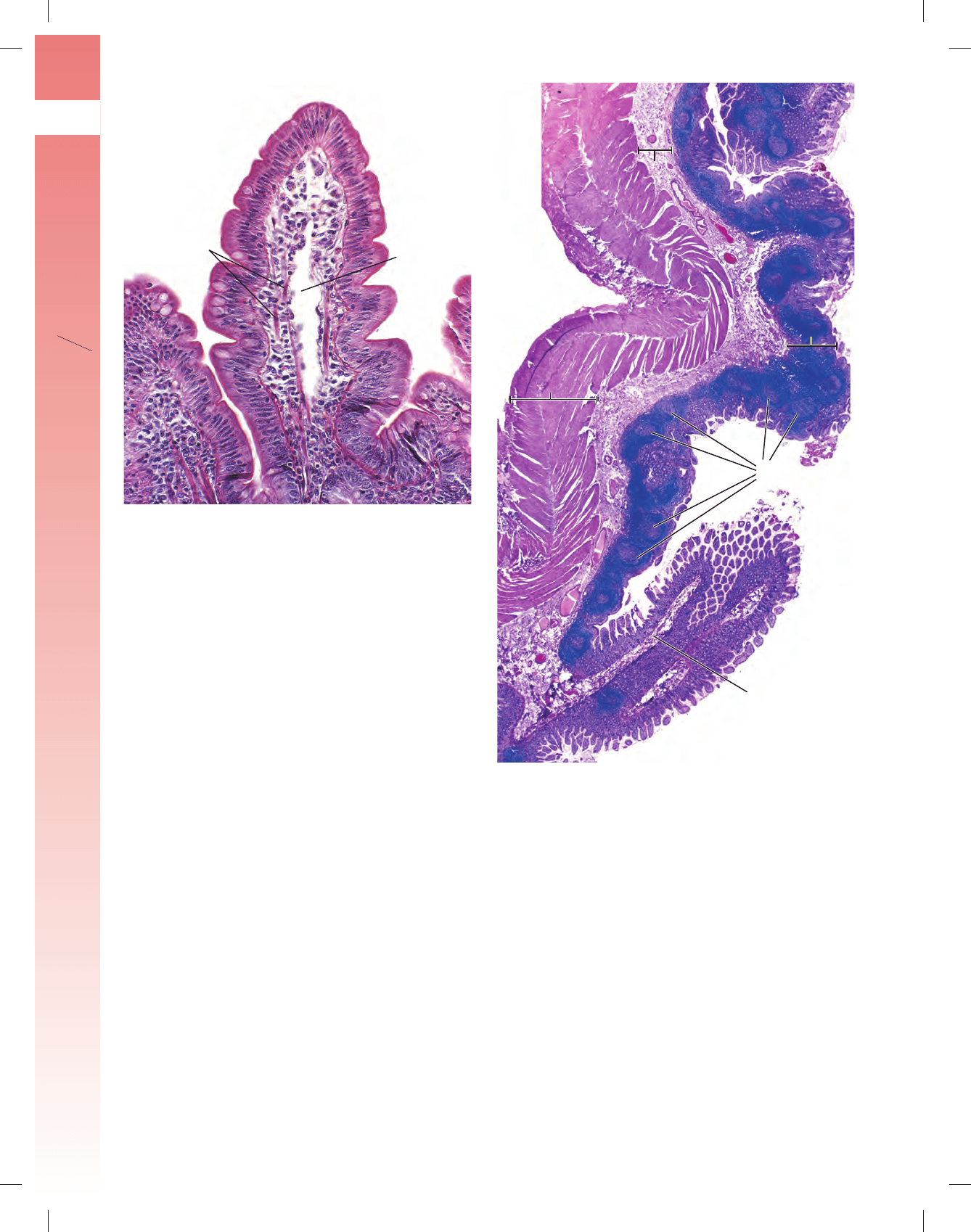
FIGURA 17-18 ▲ Vellosidades de la mucosa del intestino delgado. a. Fotomicrografía electrónica de barrido de la mucosa intestinal en la que
se ven sus vellosidades. Nótese los orifi cios (fl echas) ubicados entre las bases de las vellosidades que comunican con las glándulas intestinales (criptas de
Lieberkühn). 800 ⫻. b. Este diagrama tridimensional de las vellosidades intestinales muestra la continuidad del epitelio que las tapiza con el epitelio que
tapiza las glándulas intestinales. Nótense los vasos sanguíneos y el capilar linfático de terminación ciega, denominado vaso quilífero central, en el centro
de la vellosidad. Entre las bases de las vellosidades, se pueden ver los orifi cios de las glándulas intestinales (fl echas). Además, los orifi cios pequeños que
aparecen en la superfi cie de las vellosidades indican la ubicación de las células caliciformes que han liberado sus gránulos.
merados linfonodulares ileales o placas de Peyer.
En la anatomía macroscópica, aparecen como conjuntos de
motas blanquecinas en la mucosa.
La
muscular de la mucosa está compuesta por dos capas
delgadas de células musculares lisas, una capa circular interna y
una capa longitudinal externa. Como ya se mencionó, las haces
nos de células musculares lisas se extienden desde la muscu-
lar de la mucosa hacia la lámina propia de las vellosidades.
En el epitelio de la mucosa intestinal se encuentran al
menos cinco tipos de células.
Las células maduras del epitelio intestinal se encuentran en
las glándulas intestinales y en la super cie de las vellosidades.
Estas células consisten en:
•
Enterocitos, cuya función primaria es la absorción
•
Células caliciformes, que son glándulas unicelulares
secretoras de mucina
•
Células de Paneth, cuya función principal es mantener
la inmunidad innata de la mucosa mediante la secreción
de sustancias antimicrobianas.
•
Células enteroendocrinas, que producen varias hor-
monas endocrinas y paracrinas.
•
Células M (células con micropliegues), que son cé-
lulas especializadas (enterocitos) en el epitelio que cubren
los nódulos linfáticos en la lámina propia.
Los enterocitos son células absortivas especializadas para
el transporte de sustancias desde la luz del intestino hacia
el sistema circulatorio.
Los enterocitos son células cilíndricas altas con un núcleo
posicionado en forma basal (v. g. 17-18 y 17-21). Las micro-
vellosidades incrementan la super cie apical hasta 600 veces;
Pawlina_17_3R_Editado.indd 637Pawlina_17_3R_Editado.indd 637 15/07/15 12:4415/07/15 12:44
638
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
en los cortes para la microscopia óptica se reconocen en su
conjunto como un
borde o chapa estriada en la super cie
luminal.
Cada microvellosidad tiene un centro de micro lamentos
de actina orientados en forma vertical, que están anclados a
la villina ubicada en la punta de la microvellosidad y tam-
bién están adheridos a la membrana plasmática de toda la
estructura por moléculas de miosina I. Los micro lamen-
tos de actina se extienden dentro del citoplasma apical y se
insertan en el
velo terminal, una red de micro lamentos
contráctiles orientados en forma horizontal que forman una
capa en el citoplasma más apical y se unen a la densidad in-
tracelular asociada con la zonula adherens. La contracción del
velo terminal determina que las microvellosidades se separen,
lo cual aumenta el espacio entre ellas para permitir una mayor
super cie de exposición para que ocurra la absorción. Ade-
más, la contracción del velo terminal contribuiría a “cerrar”
las brechas dejadas en la lámina epitelial por la exfoliación de
las células envejecidas. Los enterocitos están unidos entre sí
y a las células caliciformes, a las células enteroendocrinas y a
otras células del epitelio por complejos de unión.
Las uniones herméticas establecen una barrera entre la luz
intestinal y el compartimento intercelular epitelial.
Las uniones herméticas entre la luz intestinal y el compar-
timento de tejido conjuntivo del organismo permiten la re-
Vaso
quilífero
central
Células de
músculo
liso
FIGURA 17-19 ▲ Fotomicrografía de una vellosidad intesti-
nal. La superfi cie de la vellosidad consiste en células epiteliales cilíndricas,
sobre todo enterocitos que están provistos de chapa estriada apical. Tam-
bién hay células caliciformes que pueden identifi carse con facilidad por la
acumulación apical de gránulos de mucinógeno. Debajo del epitelio se en-
cuentra la lámina propia que consiste en tejido conjuntivo laxo muy celu-
lar. La lámina propia contiene grandes cantidades de células redondeadas,
en su mayoría linfocitos. Además, pueden identifi carse células musculares
lisas. Un capilar linfático denominado vaso quilífero central ocupa el centro
de la vellosidad. Cuando el vaso quilífero está dilatado, como en esta mues-
tra, se identifi ca con facilidad. 160 ⫻.
tención selectiva de sustancias absorbidas por los enterocitos.
Como se mencionó en la sección sobre uniones ocluyentes, el
“hermetismo” de estas uniones puede variar.
En las uniones herméticas (zonulae occludens) bastante im-
permeables, como son las del íleon y el colon, para mover
solutos a través de la barrera se requiere el transporte activo.
En términos simples, los sistemas de transporte activos, por
ejemplo, las bombas de sodio (ATPAsa Na
⫹
/K
⫹
) localizadas
en la membrana plasmática lateral, reducen en forma transi-
toria la concentración citoplasmática de Na
⫹
, al transportarlo
a través de esta membrana hacia el espacio extracelular por
debajo del nivel de la zonulae occludens. Este transporte de
Nódulos
linfáticos
Pliegue
circular
Submucosa
Submucosa
Submucosa
Mucosa
Mucosa
Mucosa
Muscular
Muscular
externa
externa
Muscular
externa
FIGURA 17-20 ▲ Fotomicrografía de placas de Peyer. Esta
fotomicrografía muestra un corte longitudinal a través de la pared de un
íleon humano. Cabe notar la gran cantidad de nódulos linfáticos localiza-
dos en la mucosa y el corte de un pliegue circular que se proyecta hacia la
luz del íleon. Los nódulos linfáticos de la placa de Peyer se ubican princi-
palmente dentro de la lámina propia, aunque muchos se extienden dentro
de la submucosa. Están cubiertos por el epitelio intestinal que contiene
enterocitos, células caliciformes ocasionales y células M transportadoras de
antígeno especializadas. 40 ⫻.
Pawlina_17_3R_Editado.indd 638Pawlina_17_3R_Editado.indd 638 15/07/15 12:4515/07/15 12:45
639
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
Otros mecanismos de transporte también aumentan las
concentraciones de sustancias especí cas, como monosacári-
dos, aminoácidos y otros solutos, en el espacio intercelular.
Estas sustancias luego se difunden o uyen a favor de sus gra-
dientes de concentración dentro del espacio intercelular, para
cruzar la lámina basal epitelial e ingresar a los capilares fenes-
trados en la lámina propia, ubicados justo debajo del epitelio.
Las sustancias que son demasiado grandes para entrar en los
vasos sanguíneos, como las partículas lipoproteicas, ingresan
en el vaso quilífero linfático.
La super cie celular lateral de los enterocitos exhiben eva-
ginaciones citoplasmáticas complejas aplanadas (pliegues)
que se interdigitan con las evaginaciones de las células conti-
guas (v. g. 5-24). Estos pliegues incrementan la extensión de
super cie lateral de la célula, con lo que aumentan la cantidad
de membrana plasmática que contiene enzimas de transporte.
Durante la absorción activa, en especial de solutos, agua y
lípidos, estos
pliegues laterales se separan y agrandan el
compartimento intercelular. El aumento de la presión hidros-
tática de los solventes y los solutos acumulados causa un ujo
Aparato
de Golgi
Quilomicrones
CÉLULAS ABSORTIVAS
RER
REL
Complejos
de unión
Velo
terminal
Microvellosidades
ab
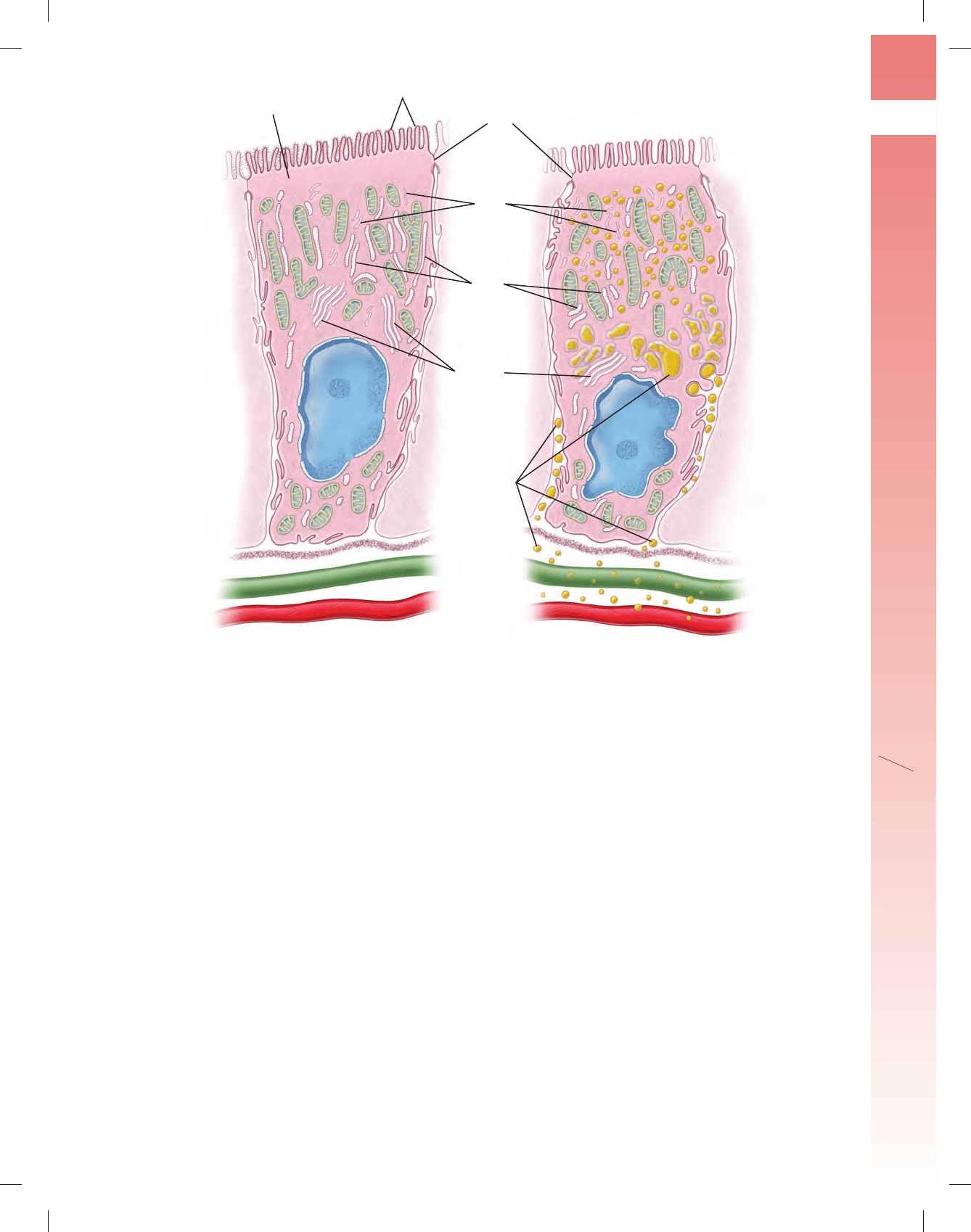
FIGURA 17-21 ▲ Diagramas de un enterocito en diferentes fases de absorción. a. Esta célula tiene una chapa estriada (borde estriado) en
su superfi cie apical y complejos de unión que aíslan la luz del intestino del espacio intercelular lateral. En el diagrama se observa el complemento de orgá-
nulos característico. b. Esta célula muestra la distribución de lípidos durante la absorción de grasas, tal como se observa con el MET. Al principio, los lípidos
aparecen en asociación con las microvellosidades de la chapa estriada. Son captados, entonces, por la célula y se ven dentro de las vesículas del retículo
endoplásmico liso (REL), en la región apical del citplasma. Los lípidos limitados por membrana pueden rastrearse hasta el centro de la célula, donde se
fusionan muchas de las vesículas que los contienen. Después se expulsan hacia el espacio intercelular. Los lípidos extracelulares, que se conocen como
quilomicrones, atraviesan la lámina basal para ser transportados hacia los vasos linfáticos (verdes), los vasos sanguíneos (rojos) o ambos.
Na
⫹
crea una concentración intercelular alta del catión, lo
cual determina que el agua de la célula salga hacia el espacio
intercelular y se reduzcan así las concentraciones de agua y
Na
⫹
en la célula. En consecuencia, el agua y el Na
⫹
ingresan
a la célula por su super cie apical, la atraviesan y salen por su
membrana plasmática lateral siempre que la bomba de sodio
continúe funcionando. El aumento de la osmolaridad en el
espacio intercelular atrae el agua hacia este espacio y esto crea
una presión hidrostática que impulsa el Na
⫹
y el agua a través
de la lámina basal hacia el tejido conjuntivo.
En los epitelios con uniones herméticas más permeables,
como las que se encuentran en el duodeno y en el yeyuno,
una bomba de sodio también crea una baja concentración de
Na
⫹
intracelular. Sin embargo, cuando el contenido que pasa
hacia el duodeno y el yeyuno es hipotónico, se produce una
absorción considerable de agua junto con Na
⫹
adicional y
otros pequeños solutos, directamente a través de las uniones
herméticas de los enterocitos en los espacios intercelulares.
Este mecanismo de absorción se conoce como
arrastre del
solvente.
Pawlina_17_3R_Editado.indd 639Pawlina_17_3R_Editado.indd 639 15/07/15 12:4515/07/15 12:45
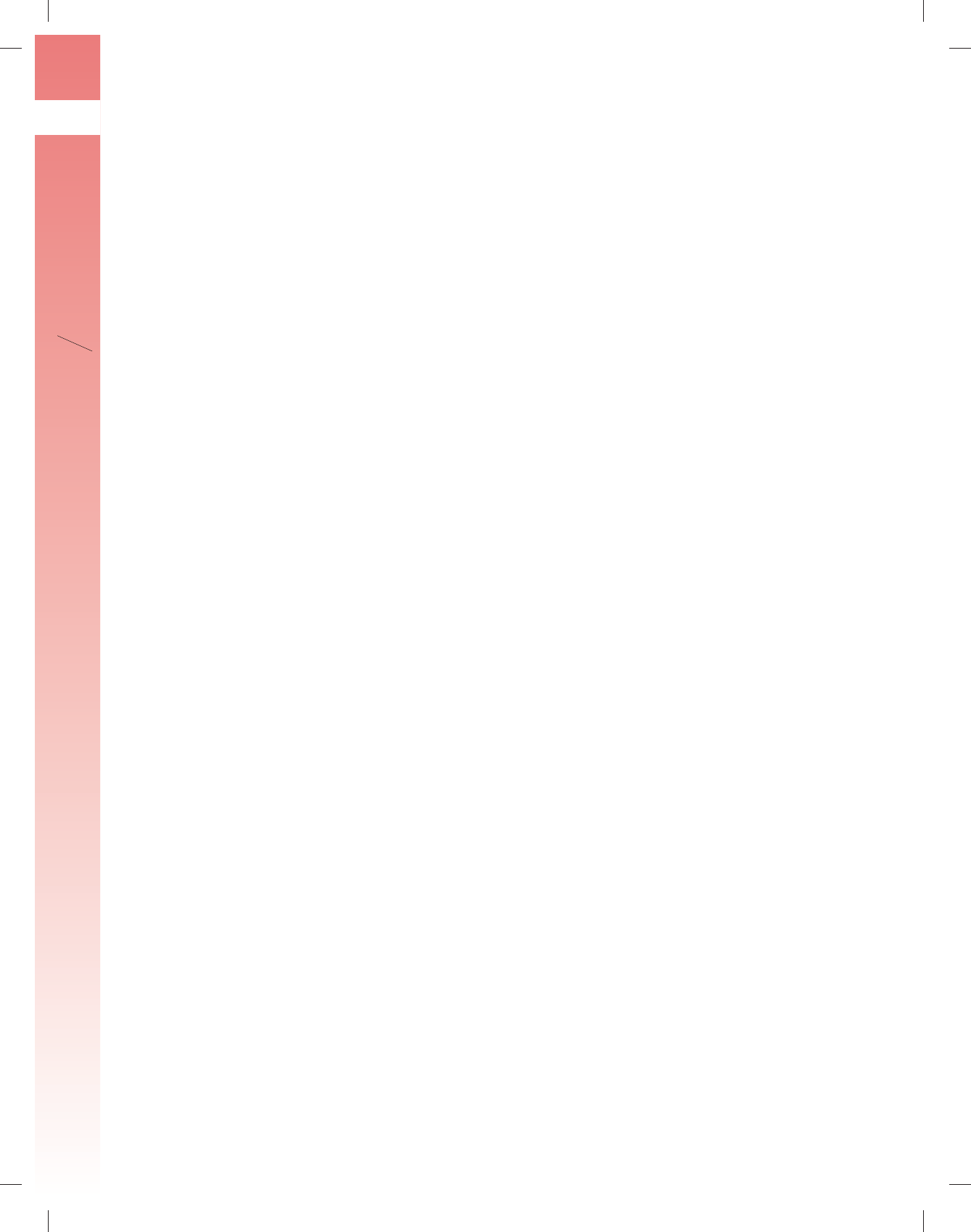
640
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
de los gránulos de mucinógeno recién formados, que es con-
tigua a la porción basal de la célula (v. g. 17-22a). Las micro-
vellosidades de las células caliciformes están restringidas en el
reborde delgado de citoplasma (la teca) que rodea la porción
apicolateral de la acumulación de gránulos de mucinógeno.
Las microvellosidades son más obvias en las células calicifor-
mes inmaduras que hay en la mitad profunda de las glándulas
intestinales.
Las células de Paneth cumplen una función en la regula-
ción de la fl ora bacteriana normal del intestino delgado.
Las células de Paneth se ubican en las bases de las glándu-
las intestinales. (A veces, también se encuentran en pequeñas
cantidades en el colon normal; su cantidad puede incremen-
tarse en ciertos estados patológicos). Estas células tienen un
citoplasma basal basó lo, un aparato de Golgi supranuclear y
grandes gránulos de secreción apicales que son muy acidó los
y refráctiles. Estos gránulos permiten su fácil identi cación en
los cortes histológicos de rutina ( g. 17-23). Los gránulos de
secr
eción contienen la enzima antibacteriana lisozima, ␣-de-
fensinas, otras glucoproteínas, una proteína con abundancia
de arginina (que puede ser la causa de la acido lia intensa)
y cinc. La
lisozima digiere las paredes celulares de ciertos
grupos de bacterias. Las ␣
-defensinas son homólogas de los
péptidos que funcionan como mediadores en los linfocitos T
CD8
⫹
citotóxicos. Su acción antibacteriana y su capacidad
para fagocitar ciertas bacterias y protozoos indican que las cé-
lulas de Paneth desempeñan un papel en la regulación de la
ora bacteriana normal del intestino delgado.
Las células enteroendocrinas en el intestino delgado pro-
ducen casi todas las mismas hormonas peptídicas que en
el estómago.
Las células enteroendocrinas en el intestino delgado se
parecen a las que se encuentran en el estómago (v. g. 17-12).
Las “células cerradas” se concentran en la porción basal de la
glándula intestinal, mientras que las “células abiertas” pueden
encontrarse en todos los niveles de cada vellosidad. La activa-
ción de los receptores de gusto que hay en la membrana celular
apical de las “células abiertas”, activan la
cascada de seña-
lización iniciada por proteínas G, que produce la libera-
ción de péptidos que regulan una gran variedad de funciones
gastrointestinales. Estas funciones comprenden la regulación
de la secreción pancreática, la inducción de la digestión y de
la absorción y el control de la homeostasis energética al actuar
sobre mecanismos nerviosos del
eje encefaloenteroadi-
poso. Casi todas las mismas hormonas peptídicas identi -
cadas en este tipo celular en el estómago, pueden detectarse
en las células enteroendocrinas del intestino (v. tabla 17-1).
La
colecistocinina (CCK), la secretina, el polipéptido
inhibidor gástrico (GIP) y la motilina son los reguladores
más activos de la siología gastrointestinal que se liberan en
esta porción del intestino (v. g. 17-13). La CCK y la secretina
incrementan la actividad del páncreas y de la vesícula biliar e
inhiben la función secretora y la motilidad gástricas. El GIP
estimula la liberación de insulina por el páncreas y la motilina
induce la motilidad gástrica e intestinal. Si bien se han aislado
otros péptidos producidos por las células enteroendocrinas,
todavía no se consideran hormonas y, por lo tanto, se los de-
nomina
candidatos hormonales (pág. 632). Las células
enteroendocrinas también producen al menos dos hormonas,
la somatostatina y la histamina, que actúan como
hormonas
direccionado a través de la lámina basal del epitelio hacia la
lámina propia (v. g. 5-1).
Además de las especializaciones de membrana asociadas
con la absorción y el transporte, el citoplasma del enterocito
también está especializado en estas funciones. Las mitocon-
drias alargadas que suministran energía están concentradas en
el citoplasma apical entre el velo terminal y el núcleo. Los
túbulos y las cisternas del retículo endoplásmico liso (REL),
que participa en la absorción de ácidos grasos y glicerol y en la
resíntesis de las grasas neutras, se encuentran en el citoplasma
apical debajo del velo terminal.
Los enterocitos también son células secretoras que pro-
ducen las enzimas necesarias para la digestión terminal y
la absorción, así como para la secreción de agua y electro-
litos.
La función secretora de los enterocitos, que consiste princi-
palmente en la síntesis de enzimas glucoproteicas que se inser-
tarán en la membrana plasmática apical, tiene como correlato
morfológico los rimeros de cisternas del Golgi en la región
supranuclear inmediata y los ribosomas libres y el RER a los
lados del aparato de Golgi (v. g. 17-21). En el citoplasma
apical, justo debajo del velo terminal y a lo largo de la mem-
brana plasmática lateral, hay pequeñas vesículas de secreción
que contienen glucoproteínas destinadas a la super cie celu-
lar. Sin embargo, para distinguir estas vesículas de secreción
de vesículas endocíticas o de lisosomas pequeños, es necesario
utilizar métodos histoquímicos o autorradiográ cos.
El intestino delgado también secreta agua y electrolitos.
Esta actividad ocurre principalmente en las células dentro
de las glándulas intestinales. Se cree que esta secreción que
ocurre en estas glándulas contribuye al proceso de digestión
y de absorción al mantener un estado líquido adecuado del
quimo intestinal. En situaciones normales, la absorción de
líquidos por el enterocito de una vellosidad está equilibrada
con la secreción de líquido por el enterocito de una glándula
intestinal.
Las células caliciformes representan glándulas unicelula-
res dispersas entre las otras células del epitelio intestinal.
Al igual que en otros epitelios, las células caliciformes pro-
ducen moco. En el intestino delgado, las células caliciformes
incrementan su cantidad desde el duodeno hasta la porción
terminal del íleon.
Además, como en otros epitelios, dado que el mucinó-
geno hidrosoluble se pierde durante la preparación de cortes
teñidos con H&E de rutina, la parte de la célula que nor-
malmente contiene gránulos de mucinógeno aparece vacía.
El examen con el MET permite observar una gran acumula-
ción de gránulos de mucinógeno en el citoplasma apical que
distiende esa región de la célula y distorsiona la forma de las
células vecinas ( g. 17-22). Con la región celular apical re-
pleta de gránulos de mucinógeno, la porción basal de la célula
parece una columna delgada. Esta porción basal es muy basó-
la en los preparados histológicos debido a que está ocupada
por un núcleo heterocromático, un RER extenso y ribosomas
libres. Las mitocondrias también están concentradas en el ci-
toplasma basal. La forma característica de esta célula, con su
dilatación apical por la acumulación de gránulos y su región
basal muy estrecha, es la causa del nombre “caliciforme”, por
su semejanza con un cáliz. Una estructura extensa de cisternas
de Golgi aplanadas forma una concavidad amplia alrededor
Pawlina_17_3R_Editado.indd 640Pawlina_17_3R_Editado.indd 640 15/07/15 12:4515/07/15 12:45
641
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
tan micropliegues en lugar de microvellosidades y una capa
delgada de glucocáliz. La super cie apical expresa abundancia
de receptores de la glucoproteína 2 (GP2) que jan macromo-
léculas especí cas y bacterias gramnegativas (p. ej., escherichia
coli). Las sustancias unidas a los receptores GP2 son captadas
por las vesículas endocíticas y transportadas a la super cie ce-
lular basolateral del receso con forma de bolsillo. Dentro del
receso, el contenido liberado se trans ere de inmediato a las
células inmunitarias que están en este espacio. Por lo tanto,
las células M funcionan como
células transportadoras de
antígeno altamente especializadas que relocalizan antígenos
intactos desde la luz intestinal a través de la barrera epitelial.
Los antígenos que alcanzan las células inmunitarias de este
modo, estimulan una respuesta en el GALT que se describe
más adelante.
Las células intermediarias constituyen el compartimento
de amplifi cación del nicho de células madre intestinales.
Las células intermediarias constituyen la mayoría de las
células del nicho de células madre intestinales que se localiza
en la mitad basal de la glándula intestinal. Estas células com-
ponen el compartimento de ampli cación de las células que
mantienen la capacidad de dividirse y que suelen sufrir una
paracrinas (v. pág. 632) (es decir, hormonas que tienen un
efecto local y no circulan en el torrente sanguíneo). Además,
varios péptidos son secretados por las células nerviosas locali-
zadas en la submucosa y la muscular externa. Estos péptidos,
denominados
hormonas neurocrinas, están representados
por el péptido intestinal vasoactivo (VIP, vasoactive intestinal
peptide), la bombesina y las encefalinas. Las funciones de estos
péptidos se reseñan en la tabla 17-2.
Las células M transportan microorganismos y otras macro-
moléculas desde la luz intestinal hacia las placas de Peyer.
Las células M son células epiteliales que cubren las placas
de Peyer
y otros nódulos linfáticos grandes; son muy dife-
rentes de las células epiteliales intestinales circundantes (cua-
dro 17-5). Las células M tienen una forma muy interesante
debido a que cada célula desarrolla un receso profundo con
forma de bolsillo conectado al espacio extracelular. Las células
dendríticas, los macrófagos y los linfocitos T y B se localizan
en este espacio. Debido a esta forma única, la super cie celu-
lar basolateral de la célula M se ubica a unos pocos micrones
de su super cie apical, con lo que se reduce mucho la distan-
cia que las vesículas endocíticas deben recorrer para cruzar la
barrera epitelial. En su super cie apical, las células M presen-
Gránulos de
mucinógeno
Aparato de
Golgi
Núcleo
Retículo
Retículo
endoplásmico
endoplásmico
rugoso
rugoso
*
*
Lámina
basal
a
Gránulos de
mucinógeno
Aparato de
Golgi
Núcleo
Retículo
endoplásmico
rugoso
Retículo
Retículo
endoplásmico
endoplásmico
rugoso
rugoso
Retículo
endoplásmico
rugoso
*
*
Lámina
basal
CÉLULA CALICIFORME
Microvellosidad
b
a
FIGURA 17-22 ▲ Fotomicrografía electrónica y diagrama de una célula caliciforme. a. Esta fotomicrografía electrónica muestra la región
basal de una célula caliciforme que se ilustra en el diagrama contiguo. La célula está apoyada en la lámina basal. La región basal de la célula contiene el
núcleo, el retículo endoplásmico rugoso y las mitocondrias. Justo encima del núcleo se observa una cantidad abundante de dictiosomas del aparato de
Golgi. A medida que el producto mucoso se acumula en las cisternas de Golgi, éstas se dilatan (asteriscos). Los gránulos de mucinógeno grandes ocupan
casi toda la región apical de la célula y en conjunto constituyen el “cáliz mucoso” que se observa con el microscopio óptico. 15 000 ⫻. b. Este diagrama
muestra una célula caliciforme completa. La región incluida en el cuadro de este diagrama corresponde a la región de la que probablemente se obtuvo
la fotomicrografía electrónica contigua. El núcleo está ubicado en la porción basal de la célula. La mayor parte de la célula está llena de gránulos de muci-
nógeno que le imparten la forma de copa o cáliz mucoso que se observa con el microscopio electrónico. En la base y en la parte inferior de los lados del
cáliz mucoso, se encuentran los sáculos aplanados del gran aparato de Golgi. Los otros orgánulos se distribuyen en el resto del citoplasma, en especial en
el citoplasma perinuclear en la base de la célula.
Pawlina_17_3R_Editado.indd 641Pawlina_17_3R_Editado.indd 641 15/07/15 12:4515/07/15 12:45
642
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
una capa de organización laxa que contiene nódulos linfáticos,
linfocitos, macrófagos, plasmocitos y eosinó los en la lámina
propia (lámina 55, pág. 656). Los linfocitos también se loca-
lizan entre las células epiteliales. Este
GALT actúa como una
barrera inmunitaria en toda la extensión del tubo digestivo. En
cooperación con las células epiteliales subyacentes, en parti-
cular las células M, el tejido linfático toma muestras de los an-
tígenos que hay en los espacios intercelulares del epitelio. Los
linfocitos, los macrófagos y otras células presentadoras de antí-
genos procesan los antígenos y migran hacia los nódulos linfá-
ticos de la lámina propia donde sufren activación (v. pág. 491),
lo cual conduce a la secreción de anticuerpos por los plasmo-
citos recién diferenciados.
La superfi cie mucosa está protegida por respuestas media-
das por inmunoglobulina.
La superfi cie mucosa del tubo intestinal se encuentra
en desafío constante por la presencia de microorganismos
(p. ej., virus, bacterias, parásitos) y toxinas ingeridas, los que
después de afectar la barrera epitelial pueden causar infeccio-
nes o enfermedades. Un ejemplo de un mecanismo de defensa
especí co es la respuesta mediada por inmunoglobulinas en
la que participan anticuerpos IgA, igM e IgE. La mayoría de
los plasmocitos de la lámina propia del intestino secretan
an-
ticuerpos dIgA
diméricos en lugar de IgG que son más co-
munes. Otros plasmocitos producen IgM pentamérica e IgE
(v. pág. 600). La dIgA dimérica está compuesta por dos subu-
nidades de IgA monoméricas y una cadena J de polipéptidos
(v. g. 16-28). Las moléculas dIgA secretadas se unen al
re-
ceptor de inmunoglobulina polimérica (pIgR)
locali-
zado en el dominio basal de las células epiteliales ( g. 17-24).
El receptor pIgR es una glucoproteína transmembrana
(75 kDa) sintetizada por enterocitos y expresada en la mem-
brana plasmática basal. El complejo pIgR–dIgA luego sufre
endocitosis y se transporta a través del epitelio hasta la super-
cie apical del enterocito (este tipo de transporte se conoce
como transcitosis). Después de que el complejo pIgR–dIgA
alcanza la super cie apical, la pIgR se escinde proteolítica-
mente y la porción extracelular del receptor que está unida a
la dIgA se libera en la luz intestinal (v. g. 17-24). Este domi-
nio extracelular de unión escindido del receptor es conocido
como
componente secretor (SC); la dIgA secretada en
asociación con el SC se denomina
IgA secretora (sIgA).
La liberación de inmunoglobulinas sIgA es decisiva para la
vigilancia inmunitaria adecuada por el sistema inmunita-
rio de la mucosa. En la luz, la sIgA se une a antígenos, toxinas
y microorganismos. La IgA secretora impide la adherencia y
la invasión de virus y bacterias a la mucosa ya sea por inhibir
su motilidad, causar aglomeración microbiana o enmascarar
los sitios de adhesión de los agentes patógenos en la super -
cie epitelial. Por ejemplo, la sIgA se une a una glucoproteína
situada en la envoltura del
virus VIH (virus de inmunode -
ciencia humana) e impide su adherencia, su incorporación y
su replicación ulterior en la célula.
La IgA secretora es la principal molécula inmunitaria de
la mucosa. S
in embargo, las moléculas IgM utilizan meca-
nismos similares de transcitosis mediada por receptores para
alcanzar la super cie de la mucosa. Algunas IgE se jan a las
membranas plasmáticas de los mastocitos de la lámina propia
(v. pág. 215-217), lo cual sensibiliza en forma selectiva a antí-
genos especí cos provenientes de la luz intestinal.
o dos mitosis antes de comprometerse a la diferenciación en
células absortivas o caliciformes. Estas células tienen micro-
vellosidades irregulares cortas con lamentos centrales largos
que se extienden en profundidad hacia el citoplasma apical y
establecen muchas uniones maculares (desmosomas) con las
células contiguas. Pequeños gránulos secretores similares a la
mucina forman una columna en el centro del citoplasma su-
pranuclear. Las células intermedias que están predestinadas a
convertirse en células caliciformes desarrollan una pequeña
colección redondeada de gránulos secretores justo debajo de
la membrana plasmática apical; en tanto que las células pre-
destinadas a convertirse en células absortivas pierden los grá-
nulos secretores y comienzan a acumular mitocondrias, RER
y ribosomas en el citoplasma apical.
El GALT es prominente en la lámina propia del intestino
delgado.
Como ya se mencionó, la lámina propia del tubo digestivo
está superpoblada por elementos del sistema inmunitario; al-
rededor de una cuarta parte de la mucosa está compuesta por
Muscular
Muscular
de la mucosa
de la mucosa
Muscular
de la mucosa
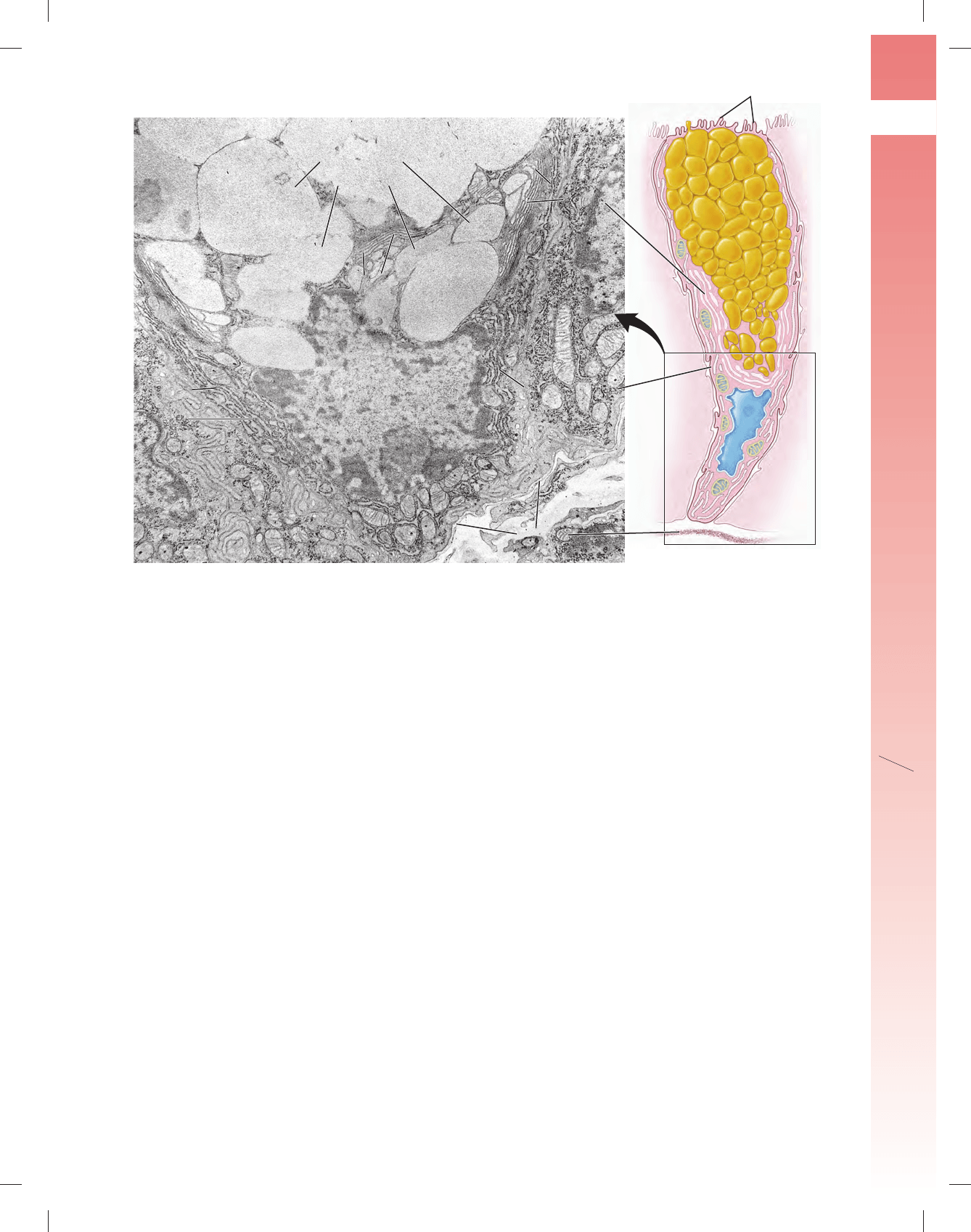
FIGURA 17-23 ▲ Fotomicrografía de las glándulas intestina-
les en las que se ven las células de Paneth. Esta fotomicrografía mues-
tra la base de las glándulas intestinales (yeyunales) en un preparado teñido
con H&E. La glándula de la derecha aparece en corte longitudinal; a la iz-
quierda de la fotografía hay otra glándula seccionada en sentido transversal
que aparece como una silueta circular. Las células de Paneth normalmente
se ubican en la base de las glándulas intestinales y se ven bien con el mi-
croscopio óptico debido a la intensa tinción con eosina de sus gránulos. La
lámina propia contiene abundantes plasmocitos, linfocitos y otras células
del tejido conjuntivo. Cabe notar que hay varios linfocitos en el epitelio de
la glándula (fl echas). 240 ⫻. Recuadro. Este aumento mayor de la región
contenida en el rectángulo muestra el citoplasma basófi lo característico de
la porción basal de las células y las grandes acumulaciones de gránulos de
secreción refráctiles muy eosinófi los en la porción apical de la célula. Es
probable que una proteína rica en arginina que se encuentra en los gránu-
los sea la causa de la intensa reacción eosinófi la. 680 ⫻.
Pawlina_17_3R_Editado.indd 642Pawlina_17_3R_Editado.indd 642 15/07/15 12:4515/07/15 12:45

643
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
CUADRO 17-5
Consideraciones funcionales: funciones
inmunitarias del tubo digestivo
Los inmunólogos han demostrado que el GALT (tejido linfático
asociado con el intestino) no sólo responde a estímulos anti-
génicos sino que también posee capacidad de vigilancia inmu-
nitaria. Esta función se ha esclarecido en forma parcial para
los nódulos linfáticos del tubo digestivo. Las células M que
cubren las placas de Peyer y los nódulos linfáticos poseen mi-
cropliegues superfi ciales distintivos que podrían confundirse
con microvellosidades gruesas en los cortes. Las células se
identifi can con facilidad con el microscopio electrónico de
barrido porque los micropliegues de su superfi cie contrastan
mucho con las microvellosidades que componen la chapa es-
triada de los enterocitos contiguos.
Se ha demostrado con la glucoproteína GP2 (que se uti-
liza como marcador molecular para las células M) que las cé-
lulas M captan proteínas y bacterias por endocitosis desde la
luz intestinal, las transportan en vesículas a través de la célula
y expulsan su contenido por exocitosis hacia recesos profun-
dos que son continuos con el espacio extracelular (fi g. C17-
5.1). Las células dendríticas y los linfocitos que hay dentro de
los recesos del espacio extracelular toman muestras de las
proteínas, incluso antígenos, provenientes de la luz intestinal
y así tienen la oportunidad de estimular el desarrollo de anti-
cuerpos específi cos contra los antígenos. El destino de estos
linfocitos expuestos aún no se ha determinado con certeza.
Algunos se mantienen dentro del tejido linfático local, pero
otros pueden estar destinados a otros sitios del organismo,
como las glándulas salivales y mamarias. Se debe recordar
que en las glándulas salivales, las células del sistema inmu-
nitario (plasmocitos) secretan IgA, que el epitelio glandular
después convierte en sIgA. Algunos estudios experimentales
señalan que el contacto con el antígeno necesario para la pro-
ducción de IgA por los plasmocitos, ocurre en los nódulos lin-
fáticos de los intestinos. Los hallazgos recientes de estudios
que utilizaron ratones carentes de GP2 muestran que la inte-
racción de la GP2 con la bacteria cumple un papel importante
en las respuestas inmunitarias específi cas contra antígeno en
las placas de Peyer. Esto puede conducir al desarrollo no sólo
de nuevas vacunas orales para enfermedades infecciosas sino
también del tratamiento innovador de tumores y enfermeda-
des infl amatorias del intestino.
b
b
Célula M
Linfocitos
Células
absortivas
Lámina
basal
a
Célula dendrítica
Macrófago
FIGURA C17-5.1 ▲ Diagrama de células M que cubren un nódulo linfático del intestino. a. Este diagrama muestra la relación de
las células M (células con micropliegues) y las células absortivas en el epitelio que cubre un nódulo linfático. La célula M es una célula epitelial que
muestra micropliegues en lugar de microvellosidades en su superfi cie apical. Tiene recesos profundos dentro de los cuales, los linfocitos, los macró-
fagos y las evaginaciones de las células dendríticas se acercan a la luz del intestino delgado. Un antígeno intacto proveniente de la luz intestinal se
transfi ere a través de la capa delgada del citoplasma apical de la célula M a los linfocitos y a otras células presentadoras de antígenos que ocupan
los recesos. b. Fotomicrografía electrónica de barrido de un nódulo linfático de una placa de Peyer que sobresale en la luz del íleon. Cabe notar
que el área del folículo cubierta por células M está rodeada por proyecciones digitiformes de la vellosidad intestinal. La superfi cie de las células
M tiene un aspecto liso. La falta de células absortivas y células caliciformes productoras de moco en la región cubierta por células M, facilitan las
reacciones inmunitarias ante los antígenos. 80 ⫻. (Reproducido con autorización de Owen RL, Johns AL. Epithelial cell specialization within human
Peyer’s patches: an ultrastructural study of intestinal lymphoid follicles. Gastroenterology 1974;66:189-203).
glándulas submucosas, también denominadas glándu-
las de Brunner.
Las glándulas submucosas tubulares rami cadas del duo-
deno, poseen células secretoras con características tanto de cé-
lulas productoras de cimógeno como de células productoras
de moco ( g. 17-25).
Submucosa
Una característica distintiva del duodeno es la presencia de
glándulas submucosas.
La submucosa está compuesta por tejido conjuntivo denso
y sitios localizados que contienen acumulaciones de adipoci-
tos. Una característica obvia en el duodeno es la presencia de
Pawlina_17_3R_Editado.indd 643Pawlina_17_3R_Editado.indd 643 15/07/15 12:4515/07/15 12:45
644
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO DELGADO
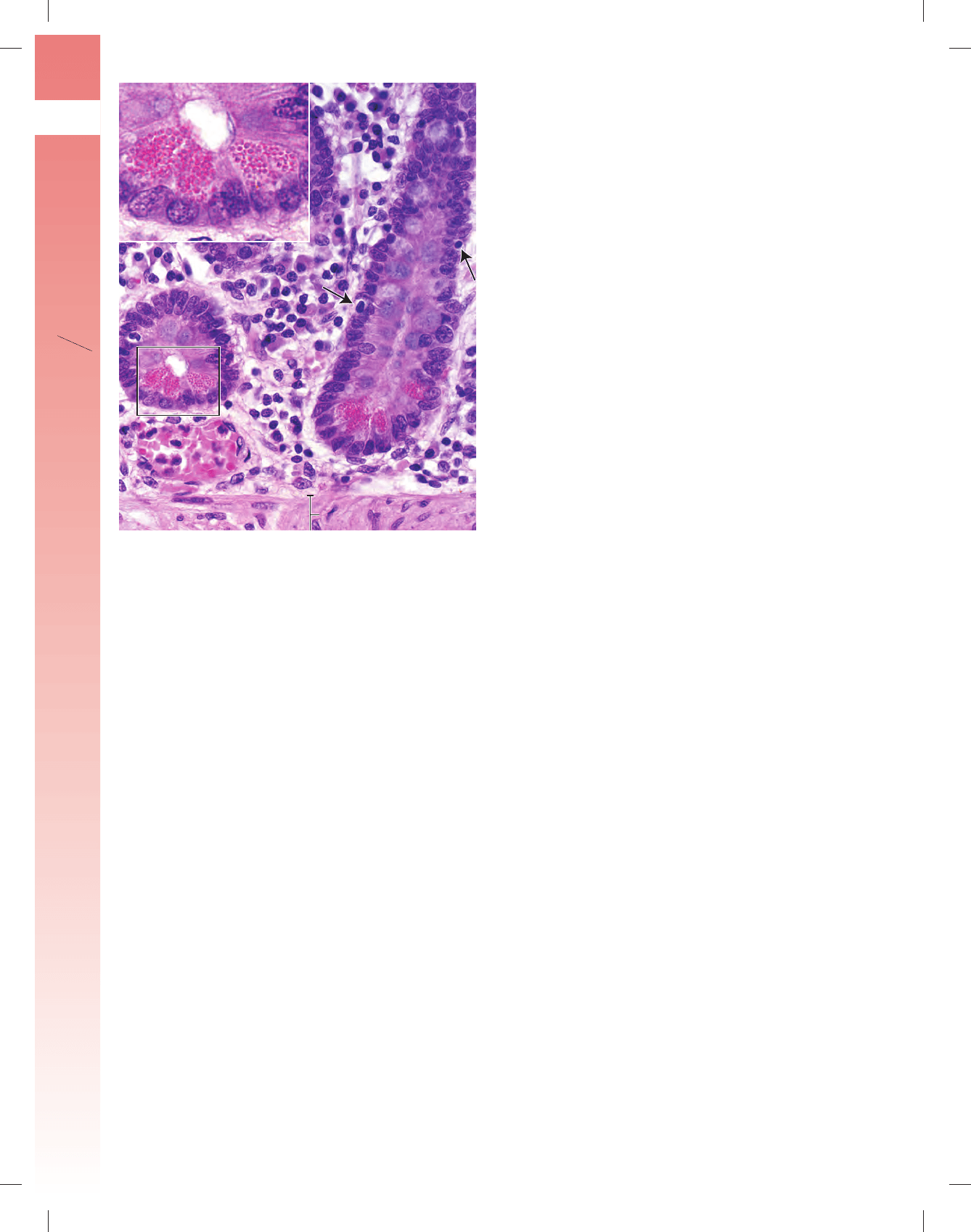
FIGURA 17-24 ▲ Diagrama de la secreción y el transporte de
la inmunoglobulina A (IgA). El plasmocito sintetiza una forma monomé-
rica de la inmunoglobulina A (IgA). La IgA se secreta hacia la lámina propia
en la forma dimérica dlgA. La dIgA está compuesta por dos subunidades
IgA monoméricas y una cadena J de polipéptidos, también sintetizada por
el plasmocito. En la lámina propia, la dIgA se une al receptor de la inmu-
noglobulina polimérica (pIgR) situado en la membrana celular basal del
enterocito. El complejo pIgR-IgA ingresa en la célula por endocitosis y se
transporta dentro de las vesículas endocíticas hasta el compartimento en-
dosómico temprano y después hasta la superfi cie apical (un proceso lla-
mado transcitosis). Las vesículas endocíticas se fusionan con la membrana
plasmática apical, el pIgR se escinde en forma proteolítica y la dIgA se libera
con la porción extracelular del receptor pIgR. Esta porción del pIgR perma-
nece con el dímero IgA y se convierte en el componente secretor (SC) de
la IgA secretora (sIgA).
LUZ INTESTINAL
LÁMINA
PROPIA
Cadena J
plgR
Endosoma
temprano
Plasmocito
Lámina
basal
dIgA
IgA
sIgA
Componente
secretor (SC)
La secreción de estas glándulas tiene un pH entre 8,1 y 9,3
y contiene glucoproteínas neutras y alcalinas e iones bicarbo-
nato. Es probable que esta secreción muy alcalina sirva para
proteger el intestino delgado proximal al neutralizar el quimo
ácido proveniente del estómago. También acerca el pH del
contenido intestinal a valores casi óptimos para la acción de
las enzimas pancreáticas que también llegan al duodeno.
Glándulas
Glándulas
de Brunner
de Brunner
Glándulas
de Brunner
Submucosa
Submucosa
Submucosa
Muscular
Muscular
de la mucosa
de la mucosa
Muscular
de la mucosa
FIGURA 17-25 ▲ Fotomicrografía de las glándulas de Brunner
en el duodeno. Esta fotomicrografía muestra parte de la pared duodenal
en un preparado teñido con H&E. Una característica distintiva del duodeno
es la presencia de las glándulas de Brunner. La línea de puntos marca el lí-
mite entre las vellosidades y las glándulas intestinales típicas (criptas de
Lieberkühn). Estas últimas se extienden hacia la muscular de la mucosa. De-
bajo de la mucosa se encuentra la submucosa, que contiene las glándulas
de Brunner. Éstas son glándulas tubulares ramifi cadas cuyos componentes
secretores están formados por células cilíndricas. El conducto de las glán-
dulas de Brunner se abre en la luz de la glándula intestinal (fl echas). 120 ⫻.
Muscular externa
La muscular externa está compuesta por una capa in-
terna de células musculares lisas dispuestas en forma circular
y una capa externa de células musculares lisas dispuestas en
forma longitudinal. Los componentes principales del
plexo
mientérico (plexo de Auerbach) se localizan entre estas
dos capas musculares ( g. 17-26). En el intestino delgado se
producen dos clases de contracción muscular. Las contraccio-
nes locales desplazan el contenido intestinal en forma tanto
proximal como distal y reciben el nombre de
contracciones
de segmentación. Estas contracciones son causadas sobre
todo por la capa muscular circular. Sirven para movilizar el
quimo en forma local al mezclarlo con los jugos digestivos y
hacer que entre en contacto con la mucosa para la absorción.
La
peristalsis, la segunda clase de contracción, comprende
la acción coordinada de las capas musculares circular y lon-
gitudinal y desplaza el contenido intestinal en forma distal.
Serosa
La serosa de las partes del intestino delgado que están cu-
biertas por peritoneo dentro de la cavidad abdominal, se co-
Pawlina_17_3R_Editado.indd 644Pawlina_17_3R_Editado.indd 644 15/07/15 12:4515/07/15 12:45

645
CAPÍTULO 17 Sistema digestivo II: esófago y tubo digestivo INTESTINO GRUESO
rresponde con la descripción general presentada al comienzo
de este capítulo.
Renovación celular epitelial
en el intestino delgado
Todas las células maduras del epitelio intestinal provienen
de una sola población de células madre.
Las células madre están situadas en la base de la glándula in-
testinal. Este
nicho de células madre intestinales (zona
de replicación celular) está restringido en la mitad basal de
la glándula y contiene células intermedias muy proliferati-
vas (como ya se explicó) y células en diversas etapas de dife-
renciación. Una célula destinada a convertirse en una célula
caliciforme o absortiva suele sufrir varias mitosis adicionales
después de abandonar el reservorio de células madre. Las cé-
lulas epiteliales migran hacia arriba en la glándula intestinal
y ascienden por la vellosidad hasta que sufren apoptosis y
se exfolian hacia la luz. Los estudios autorradiográ cos han
mostrado que el tiempo de renovación para las células ab-
sortivas y las
células caliciformes en el intestino delgado
humano es de 4 a 6 días.
Las
células enteroendocrinas y las células de Paneth
también derivan de los células madre de la base de la glándula
intestinal. Al parecer, las células enteroendocrinas se dividen
sólo una vez antes de sufrir diferenciación. Migran junto con
las células absortivas y las células caliciformes pero con un
ritmo más lento. Las
células de Paneth migran hacia abajo
y permanecen en la base de la glándula intestinal. Viven alre-
dedor de 4 semanas y después son reemplazadas por diferen-
ciación de una célula “predestinada” cercana en la glándula
intestinal. Las células que se reconocen como células de Pa-
CB
SM
N
SM
CB
BV
SC
CB
SM
N
SM
CB
BV
SC
CB
SM
N
SM
CB
BV
SC
CB
SM
N
SM
CB
BV
SC
CB
SM
N
SM
CB
BV
SC
FIGURA 17-26 ▲ Fotomicrografía electrónica del plexo mientérico (de Auerbach). El plexo está ubicado entre las dos capas de músculo liso
(SM) de la muscular externa. Está compuesto por somas neuronales (CB) y una gran red de fi bras nerviosas (N). Junto a los somas neuronales se observa
una célula satélite (SC), también conocida como célula neuróglica entérica. Estas células tienen características estructurales y químicas en común con las
células gliales del sistema nervioso central. BV, vaso sanguíneo. 3 800 ⫻.
neth no se dividen más. Como ya se mencionó en el capítulo
sobre tejido epitelial (pág. 171), la expresión del
factor de
transcripción Math1 parece determinar el destino de las
células en diferenciación en el nicho de células madre intes-
tinales. Las células destinadas al linaje secretor (es decir, las
células que se diferenciarán en células caliciformes, enteroen-
docrinas y de Paneth) sufren un aumento de la expresión de
Math1. La inhibición de la expresión de Math1 caracteriza
el mecanismo de desarrollo por defecto que da origen a las
células intestinales absortivas (enterocitos).
INTESTINO GRUESO
El intestino grueso comprende el ciego con su apéndice
vermiforme, el colon, el recto y el conducto anal. El
colon a su vez se subdivide según su ubicación anatómica en
colon ascendente, colon transverso, colon descen-
dente y colon sigmoide. Las cuatro capas características
del tubo digestivo también aparecen en todo el intestino
grueso. Sin embargo, en el nivel macroscópico se comprue-
ban varios rasgos distintivos ( g. 17-27):
•
Tenias del colon, que representan tres bandas equidis-
tantes, estrechas y gruesas formadas por la capa longitudi-
nal externa de la muscular externa. Se ven sobre todo en el
ciego y en el colon y están ausentes en el recto, el conducto
anal y el apéndice vermiforme.
•
Haustras colónicas, que son saculaciones visibles entre
las tenias del colon en la super cie externa del ciego y del
colon.
•
Apéndices omentales, que son pequeñas proyecciones
adiposas de la serosa que se observan en la capa externa
del colon.
Pawlina_17_3R_Editado.indd 645Pawlina_17_3R_Editado.indd 645 15/07/15 12:4515/07/15 12:45

intestino.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.