
HORMONAS
Hipófisis: también llamada glándula pituitaria es un órgano neuro - glandular situado en la cara inferior del
cerebro, en el piso medio de la base del cráneo. Se ubica en la parte inferior del cerebro, por fuera de éste, más
precisamente detrás del quiasma óptico, debajo del diencéfalo y del piso del tercer ventrículo del cerebro, a
continuación del hipotálamo, al cual está unido por medio del tallo pituitario. Se aloja en la silla turca, concavidad
central y media de la cara superior del hueso esfenoides y limitada por las cuatro apófisis clinoides (salientes
óseas, dos anteriores y dos posteriores a la izquierda y derecha). Hacia arriba la duramadre se extiende sobre
la silla turca formando el diafragma de la silla, tienda de la hipófisis que en la parte central presenta un orificio
por el que pasa el tallo pituitario.
Relaciones anatómicas: adelante con el trasfondo de las fosas nasales, abajo con el seno esfenoidal,
lateralmente con los órganos que contiene el seno cavernoso (lagos venosos, carótida interna, y nervios
oculomotores, nervio oftálmico), atrás con el tronco basilar que asienta sobre la protuberancia y más hacia atrás
la celda cerebelosa, arriba, con la cara inferior del cerebro, a través del tallo pituitario que atraviesa la tienda de
la hipófisis. Así, el tallo se relación atrás con los tubérculos mamilares, arriba con el tercer ventrículo y adelante
con el quiasma óptico mediante la tienda de la hipófisis.
Constitución anatómica: la hipófisis se compone de dos lóbulos: uno posterior o nervioso la neurohipófisis y otro
anterior o glandular, la adenohipófisis. Estos lóbulos se diferencian a nivel anatómico, histológico y funcional.
Adenohipófisis: es la glándula principal del sistema endocrino, ya que regula a la mayoría de las demás
glándulas endocrinas. Se compone de tres partes: la porción tuberal, la porción distal y la porción intermedia.
La porción tuberal es una prolongación superior de la adenohipófisis que se dispone alrededor del tallo neural
(de la neurohipófisis), con lo cual forma parte del tallo pituitario o infundíbulo (de la hipófisis), está formada por
cordones y grupos celulares pequeños que se localizan entre los sinusoides de la red capilar primaria.
La porción distal es la más anterior y voluminosa de la hipófisis, segrega hormonas que controlan y regulan el
funcionamiento de la mayoría de las glándulas endocrinas, está envuelta por una capsula de tejido conectivo,
rico en fibras colágenas y vasos sanguíneos que acompañan al estroma, las células de esta porción se clasifican
de acuerdo a la afinidad por los colorantes hematoxilina – eosina en células cromófilas (sus gránulos se tiñen
intensamente, pueden ser acidófilas cuando se tiñen con eosina o basófilas cuando se tiñen con hematoxilina),
y células cromófobas (sus gránulos no tienen afinidad por los colorantes). El parénquima en esta porción se
compone de células basófilas (10%), acidófilas (40%) y cromófobas (50%), entre las que transcurren capilares
sinusoides (de sinuoso, que se curvan de un lado al otro). Las células acidófilas son células redondeadas, de
menor tamaño que las basófilas y más numerosas, con vesículas citoplasmáticas que se colorean de rojo o
anaranjado con eosina, hay de 2 tipos células somatotropas que segregan la hormona del crecimiento, son
numerosas, se agrupan a los lados de los capilares en zonas laterales de la porción distal de la adenohipófisis,
presentan un RER y complejo de Golgi muy desarrollados ya que la hormona que sintetizan es de naturaleza
proteica; y las células lactotropas o mamotropas que segregan prolactina, se ubican en el interior de los
cordones celulares del parénquima, aisladas, contienen gránulos voluminosos, el RER y el complejo de Golgi
aumentan durante la lactancia. Las células basófilas hay 3 tipos células tirotropicas que secretan la TSH, son
grandes y poligonales, se ubican en el interior de los cordones, contienen pequeños gránulos con hormona
tirotrofina; las células gonadotropas que segregan las gonadotrofinas LH y FSH, son esféricas, con gránulos de
tamaño variable, RER y complejo de Golgi muy desarrollados, se disponen aisladas o en grupos entre los otros
tipos celulares; y las células corticotropas que segregan ACTH que índice a la corteza suprarrenal a que
produzca corticoesteroides, se localizan en zona cercana a la porción distal, son grandes, esféricas u ovoides,
con escasos gránulos y de baja tinción.

Las células cromófobas son pequeñas, están ubicadas en el interior de los cordones celulares, sus gránulos
no presentan afinidad por hematoxilina – eosina.
La porción intermedia es una lámina epitelial delgada que separa los dos lóbulos, estrechamente adosado al
lóbulo posterior.
Hipófisis intermedia: está formada por células basófilas que rodean una serie de cavidades quísticas que se
extienden hasta el interior de la parsnervosa. Las células basófilas poseen gránulos que contiene alfa o
betaendorfinas, es un producto secundario del proceso de postraduccion de la hormona lipotrofica (LPH)
Neurohipófisis: se origina de neuroectodermo, es de color amarillo grisáceo, se compone de dos partes, el tallo
infundibular o neural y el proceso infundibular o porción nerviosa. El tallo neural se prolonga desde arriba, a
continuación del tuber cinereum (del hipotálamo, en el piso del diencéfalo), por medio de la eminencia media,
hasta la porción nerviosa. Es el sitio de depósito de las neuro secreciones de las neuronas de los núcleos
supraópticos y paraventricular del hipotálamo. Es una vía nerviosa cuyas terminaciones almacenan y liberan
productos de secreción del hipotálamo y contiene axones amielíticos y terminaciones nerviosas de neuronas
neurosecretoras cuyos cuerpos se encuentran en los núcleos supraópticos y paraventriculares del hipotálamo.
El resto de la estructura de la neurohipófisis presenta células semejantes a gliales (pituicitos)
Irrigación de la hipófisis: estructura vascular que tiene la importante
función de asegurar la irrigación de los componentes del sistema
nervioso central que se en cuentan en la cavidad craniana, el polígono
de Willis. Este circuito arteria resulta de la anastomosis en forma
poligonal de tres ramas arteriales importantes, las arterias carótidas
internas izquierda y derecha, y el tronco basilar. El polígono de Willis
se encuentra por encima de la silla turca, en el espacio subaracnoideo
de la base del cerebro, rodeando el quiasma óptico y el tallo pituitario,
se compone de las siguientes estructuras vasculares las arterias
carótidas internas izquierda y derecha, cerebrales anteriores
(izquierda y derecha, siendo cada una rama de las carótidas internas),
las cerebrales posteriores izquierdas y derechas (estas nacen de la
bifurcación del tronco basilar) y tres pequeñas arterias que cierran el
polígono, dos comunicantes posteriores (que unen las cerebrales
posteriores con las carótidas internas, atrás a ambos lados), y una
arteria comunicante anterior (la cual comunica las dos cerebrales
anteriores, por delante del quiasma óptico).
Las arterias hipofisarias superiores (AHS) derivan de las arterias
carótida interna y comunicante posterior del polígono de Willis e irrigan
las pars tuberalis, la eminencia media y el tallo infundibular formando
una corona alrededor del infundíbulo. Las AHS dan origen a capilares
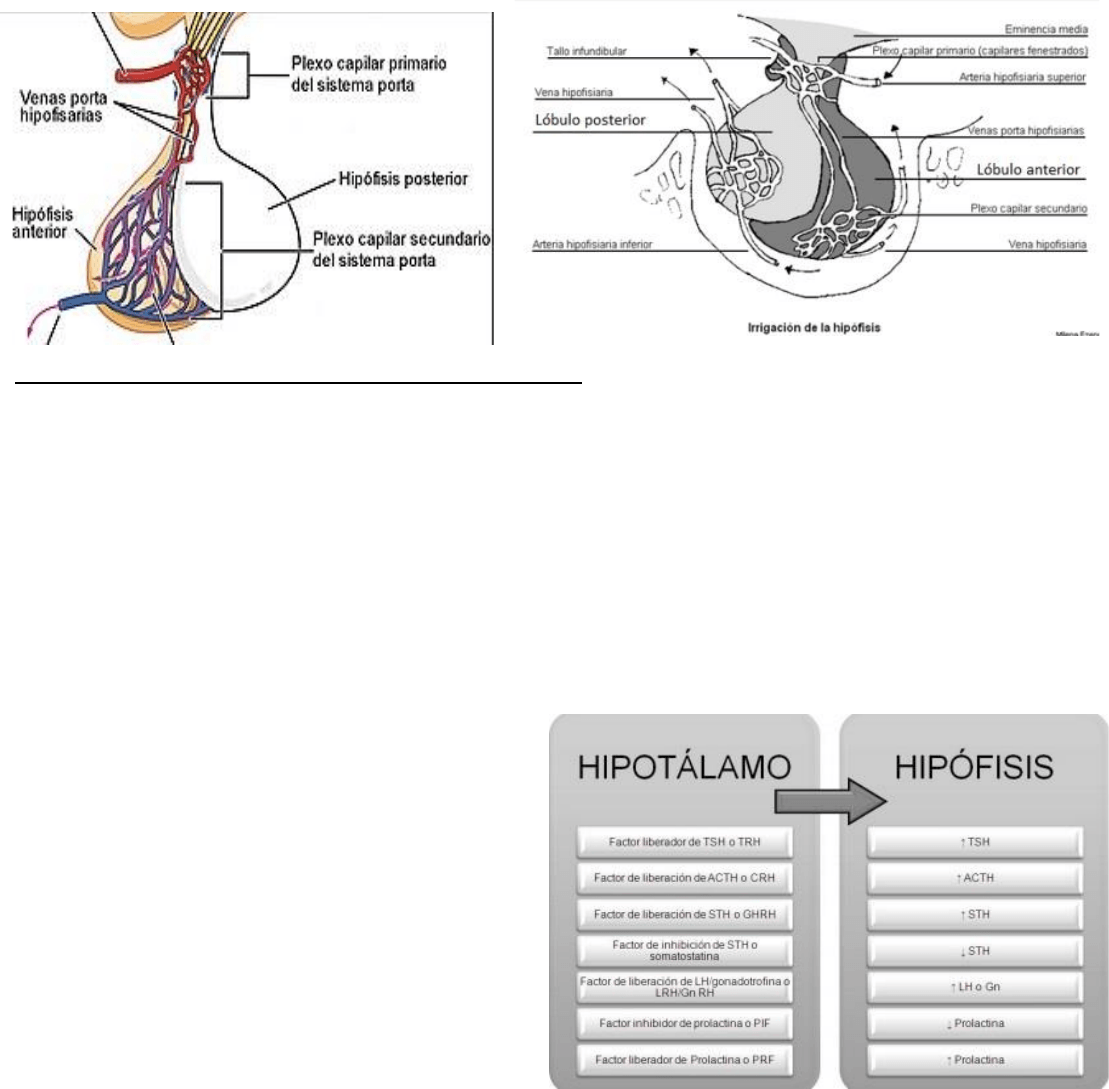
fenestrados (plexo capilar primario). Luego de esto los capilares drenan en las venas porta hipofisarias que
corren a lo largo de la pars tuberlis y dan origen a una segunda red capilar (plexo capilar secundario). Se
establece así el sistema porta hipofisiario, constituido por una vía venosa interpuesta entre dos red de capilares.
El sistema porta transporta las secreciones neuroendocrinas de los axones hipotalámicos.
Las arterias hipofisarias inferiores derivan solo de la arteria carótida interna e irrigan la neurohipófisis y envían
algunas ramas que terminan en los capilares de la adenohipófisis. La sangre de los capilares tanto de la
adenohipófisis y de la neurohipófisis, es drenada por las venas hipofisarias hacia el seno cavernoso y de allí a
las venas yugulares internas.
Regulación hipotalámica sobre la secreción de la hipófisis: el hipotálamo contiene grupos de neuronas capaces
de sintetizar hormonas y factores que controlan la secreción hipofisaria. La hipófisis posterior, las fibras
nerviosas que la constituyen se originan en células nervios ubicadas en los núcleos supraópticos y
paraventricular del hipotálamo. La hipófisis anterior es controlada por “factores hipotalámicos de liberación o
inhibición” sintetizados por neuronas hipotalámicas cuyo axón los vuelca en los vasos que constituyen el
“sistema porta hipotalámico - hipofisario” para actuar en las células glandulares de la hipófisis y controlar su
secreción. El hipotálamo es un área pequeña del diencéfalo que sirve de enlace entre la psiquis y el sistema
endocrino. El dolor, los sentimientos, sensaciones visuales, auditivas, la concentración de agua, nutrientes,
mensajeros químicos, son señales reconocidos por el hipotálamo para generar después diferentes respuestas.
El hipotálamo al sintetizar y secretar los factores de liberación o inhibición controla de manera indirecta a todos
los órganos blanco de la hipófisis: tiroides, corteza suprarrenal y gónadas. Controla el funcionamiento de la
mayoría de las células del organismo y así se constituye un área integrada y de unión entre la psiquis y el
cuerpo.
Los factores hipotalámicos que regulan la secreción
de las hormonas:
• Factor de liberación de TSH o TRH
• Factor de liberación de ACTH o CRH
• Factor de liberación de STH o GHRH
• Factor de inhibición de STH o somatostatina
• Factor de liberación de LH o gonadotrofina o LRH
• Factor inhibidor de prolactina o PIF
• Factor liberador de prolactina o PRF
El hipotálamo sintetiza y secreta encefalinas,
endorfinas, sustancia P, colecistoquinina y angiotensina.

Hormonas de la hipófisis anterior (adenohipófisis): La STH es la única hormona de la Adenohipófisis que actúa
sobre la mayoría de los tejidos. La TSH, LH, FSH, ACTH, y prolactina actúan sobre la glándula tiroides, ovarios
o testículos y mamas respectivamente.
1. Hormona del crecimiento o STH: es una hormona de naturaleza proteica y su función es provocar el
crecimiento de todos los tejidos, estimulando la reproducción celular y el aumento de su tamaño, tiene efectos
sobre los metabolismos más importantes.
• Efectos de la STH sobre el crecimiento: su efecto no es por acción directa sino a través de las
somatomedinas secretados por el hígado bajo la estimulación de STH, las somatomedinas actúan sobre el
cartílago y el hueso fomentando su crecimiento a través de una aceleración de la condrogénesis,
ensanchamiento de los discos cartilaginososepifisiarios y aumento de los depósitos en la matriz ósea, en los
extremos de los huesos largos, las somatomedinas pueden producción los mismos efectos metabólicos que
la STH.
• Efectos metabólicos de la STH:
✓ Incrementa el anabolismo proteico en todo el cuerpo (estimula el transporte de aa a través de las
membranas celulares, estimula la síntesis proteica en los ribosomas, incrementa la transcripción de ADN
para formar ARN, disminuye el catabolismo de proteínas y aa y aumenta la eritropoyesis),
✓ Aumenta la liberación y aprovechamiento de los ácidos grasos (provoca la liberación de ac. grasos del
tejido adiposos lo que provoca un aumento de estos en los líquidos corporales, estimula la conversión de
ac. graso en acetilCoA para la formación de ATP, al utilizar los ac. grasos como energía se ahorra aa que
sean utilizados para la síntesis proteica, la utilización de ac. grasos incrementa la formación de cuerpos
cetónicos)
✓ Disminuye el índice de utilización de glucosa (disminuye la utilización de glucosa para obtener energía,
aumento de los depósitos de glucógeno, disminuye la captación de glucosa por las células, y esto provoca
un aumento de la glucemia, el aumento de la glucemia provoca la estimulación en la secreción de insulina)
✓ Acción sobre el balance de electrolitos (aumenta la absorción intestinas de Ca y reduce la excreción de
Na y K.).
• Regulación de la secreción de STH: la secreción de STH está controlada por el hipotálamo, a través del
factor liberador de STH y la somatostatina que llegan a la adenohipófisis por el sistema porta hipotálamo –
hipofisario. La STH es secretada en la niñez y después de la adolescencia cuando se detiene el crecimiento.
La secreción de STH en el niño y en el adulto está sujeta a fluctuaciones rápidas y de magnitud en respuesta
a numerosos estímulos.
• Estímulos que aumentan la secreción de STH:
✓ Deficit de sustrato energético
❖ Hipoglucemia
❖ Ayuno
❖ Ejercicio
✓ Aumento de ciertos aa en plasma
❖ Comida proteica
❖ Infusión de arginina y otros aa
✓ Productos de estrés
❖ Fiebre
❖ Stress psíquico
✓ Glucagón
✓ Dormir
✓ Estrógenos
✓ Agonistas o adrenérgicos
✓ L. DOPA
✓ Apomorfina

• Estimulo que disminuyen la secreción de STH: STH, glucosa, cortisol, ac. grasos libres, sueños con
movimientos oculares rápidos, progesterona
El núcleo del hipotálamo que secreta factor liberador de STH es el ventromedial, la misma área que provoca el
hambre.
El control de la secreción de STH está sometido a un mecanismo de retroalimentación negativa.
2. Prolactina:
• Efectos de la prolactina:
✓ Estimulación y mantenimiento de la secreción de leche durante la lactación de la mujer.
✓ Los estrógenos y la progesterona inhiben la secreción láctea durante la gestión.
✓ La secreción hipofisaria de prolactina empieza a aumentar en forma constante a partir de la quinta
semana de embarazo.
✓ Después del nacimiento la mama secreta unos milímetros de calostro.
✓ El descenso de la secreción de estrógenos y progesterona placentarios que ocurren tras el parto, permite
que el efecto lactógeno de la prolactina no sea inhibido.
✓ La STH, cortisol y la parathormona son necesarias para la producción de leche con las cantidades
suficientes de aa, ac. grasos, glucosa y calcio.
✓ Durante la lactancia la prolactina tiene un efecto antigonadotrofico que ocasiona la suspensión del ciclo
ovárico.
• Regulación de la prolactina: la secreción de prolactina esta inhibida por el hipotálamo a través de un factor
inhibidor o PIH, menos en el periodo de lactancia, la secreción de prolactina esta estimulada cada vez que
la madre da de amamantar al bebe, la succión de los pezones emite señales al hipotálamo que secreta un
factor liberador que incrementa en forma intermitente la secreción de prolactina.

Hormonas de la hipófisis posterior (neurohipófisis): constituida por pituicitos, células gliares de sostén y
paraxones provenientes de núcleos hipotalámicos, secreta 2 hormonas, antidiurética o HAD y la oxitocina, son
sintetizadas por las neuronas de los núcleos hipotalámicos y luego son transportadas en combinación con una
proteína (neurofisina) hacia la neurohipófisis en un trayecto que requiere varios días, el núcleo supraópticos
sintetiza HAD, el núcleo paraventricular sintetiza oxitocina, la neurofisina se separa de la neurohormona
después de su liberación.
1. Hormona antidiurética o HAD: actúa en el tejido blanco fijándose como receptor de membrana plasmática,
y los receptores de HAD se encuentran en la membrana basal de los túbulos colectores y la unión hormona –
receptor activa la adenilciclasa y forma AMPc, que abre los poros de las membranas celulares permitiendo la
difusión de agua hacia el espacio peritubular.
• Efectos de la HAD: acción antidiurética al actuar sobre los túbulos colectores y distales del nerón renal,
haciéndolos más permeables al agua, la cual se reabsorbe pasando luego a los capilares, a
concentraciones normales de HAD no ejerce ningún efecto vasopresor, pero a mayor concentración
produce constricción de las arteriolas.
• Regulación de su secreción: el principal estímulo para la secreción es la hiperosmolaridad del plasma,
cuando el líquido extracelular es hipotónico, los osmorreceptores se hinchan y se inhibe la liberación de
HAD, cuando el líquido extracelular es hipertónico, disminuyen de volumen los osmorreceptores y se
provoca la secreción de HAD, la disminución de la presión arterial estimula la secreción e HAD.
• Factores que afectan la producción de HAD: aumenta su secreción: traumatismos, morfina, dolor, nicotina,
tranquilizantes, ansiedad y disminuyen su secreción: alcohol.
2. Oxitocina
• Efectos sobre las mamas:
✓ Produce la eyección de leche a través de un reflejo neuroendocrino.
✓ Estimulación que produce la succión del pezón, genera un reflejo que viaja hasta los núcleos
paraventriculares del hipotálamo, se estimula la secreción de oxitocina que ocasiona la concentración de
células mioepiteliales. Provoca la expulsión de leche hacia los conductos y de allí al pezón
✓ En mujeres lactantes, la estimulación genital y las emociones pueden producir secreción de oxitocina.
• Efectos sobre el útero:
✓ Produce contracciones en la musculatura uterina, durante el comienzo del parto
✓ Al final del embarazo aumenta el número de receptores de esta hormona que permite que el útero se
vuelva muy sensible a su acción.
✓ La dilatación del cuello uterino al comenzar el trabajo de parto genera un reflejo que lleva una importante
secreción de oxitocina para la contracción de las paredes uterinas y facilitar la expulsión del feto.
Glándula tiroides:
Anatomía macroscópica: es una glándula impar, casi simétrica, está ubicada en la base del cuello, por delante
y a ambos lados de la laringe y la tráquea, es de color marrón rojiza, es de consistencia blanda, está rodeada
por una capsula externa de tejido conectivo, separada del tejido conectivo laxo por una capa de tejido conectivo
denso unido al tejido glandular, está constituida por 2 lóbulos, derecho e izquierdo, reunidos por un istmo
transversal, con el que adopta forma de H.
Lóbulos: tienen forma piramidal, la parte inferior es más gruesa que la superior que se afina hacia arriba hasta
terminar en un vértice a la altura del cartílago tiroides, se distinguen 3 caras y 2 polos. Caras: la anteroexterna
que es convexa superficial cubierta por una capa muscular, la posterointerna cóncava aplicada contra la cara
anterolateral de la laringe y de la traque, y la posterior que es menos extensa, orientada hacia atrás y afuera.

Polos: el inferior que es convexo, hacia abajo y adentro, desciende hasta el 5° o 6° anillo de la tráquea, y el
superior que es más afinado, orientado hacia arriba y atrás en contacto con el borde inferior del cartílago tiroides.
Istmo: reúne los 2 lóbulos en el centro, con los que continua cerca de los polos inferiores. No presenta un límite
definido, pero se distinguen dos bordes y dos caras. Caras: la anterior que es convexa y la posterior que es
cóncava, y abraza los primeros cartílagos traqueales. Bordes: uno superior que es cóncavo arriba y responde
al cartílago criocoides y otro inferior que es corto y cóncavo.
Relaciones de la tiroides: anterolaterales: corresponden a cara anterior del istmo y anteroexterna de los lóbulos
recubiertos por, piel, tejido subcutáneo y el musculo cutáneo del cuello; aponeurosis superficial, dispuesta entre
ambos músculos esternocleidomastoideos; músculos infrahioideos. Posterior: la línea media el istmo se apoya
y corresponde al 2° o 3° anillo de la tráquea y puede llegar al cartílago cricoides, la cara posterior se relaciona
con el plexo venoso profundo, origen de venas tiroideas inferiores, el borde superior se relaciona con el espacio
pre laríngeo que contiene ganglios prelaringeos, de este borde se desprende el lóbulo piramidal, la cara posterior
de los lóbulos lateralmente esta delante del paquete vasculonervioso yugulocarotideo, con la arteria carótida
primitiva por dentro, la vena yugular interna por fuera y entre ambas y atrás el nervio vago, los ganglios linfáticos
yugulocarotideos son laterales a la vena yugula interna, la cara posterior interna de los lóbulos se amolda al
conducto aerodigestivo, responde a las glándulas paratiroides, a la arteria tiroidea inferior y a los nervios
recurrentes. Polos: polo superior llega a la mitad del cartílago tiroides mientras que el polo inferior se ubica
detrás y adentro del confluente yugolosubclavio a la altura del 5° anillo de la tráquea.
Vascularización: ricamente vascularizada por 4 arterias, 2 a cada lado, arterias tiroideas superiores e inferiores.
Arterias: tiroidea superior son colaterales de la arteria carótida externa, se dirige en sentido medial y luego
hacia abajo, hacia el polo superior de la glándula tiroides donde termina, las ramas colaterales son arterias
laríngea superior, inferior y externa, y las ramas terminales hacen anastomosis unas con las homologas
contralateral y otra con la tiroides inferior del mismo lado; la tiroidea inferior se origina en la arteria subclavia,
presenta 3 segmentos: vertical, transversal y terminal, las ramas colaterales se dirigen al esófago, a la tráquea,
a la laringe y a los músculos cercanos, y las ramas terminales son 3 y hacen anastomosis unas con la homologa
contralateral y otra con la tiroidea superior del mismo lado; y la tiroidea media la anastomosis arteriales
longitudinales reúnen las arterias del mismo lado, y las transversales reúnen las arterias del lado opuesto
Venas: venas tiroideas superiores siguen el trayecto de la arteria tiroidea superior y terminan en el tronco
venoso tirolinguofacial, en forma directa en la vena yugular interna; venas tiroideas inferiores se forman en el
polo inferior de cada lóbulo y terminan en la vena yugular interna; venas tiroideas laterales medias emergen
de la parte media del lóbulo lateral, terminan en la vena yugular interna, es inconstante; y las venas ístmicas
venas inferiores que terminan en el tronco venoso braquiocefálico izquierdo.

Linfáticos: se originan en el interior de la glándula, forman un plexo subescapular, los colectores eferentes son
medios y laterales, linfáticos medios presentan colectores superiores que se dirigen al ganglio prelaringeo y
colectores inferiores que siguen las venas media hasta llegar a los ganglios intratoracicos y ganglios
pretraqueales, y linfáticos laterales: emergen en toda la extensión de los lóbulos, alcanzan los ganglios de la
cadena yugular y siguen las venas.
Nervios: el pedículo superior e inferior con fibras simpáticas y parasimpáticas mezcladas, no siguen el trayecto
de las arterias
Anatomía microscópica (histología): está rodeada por 2 envolturas de tejido conectivo denso, separadas entre
si por tejido conectivo laxo, la envoltura mas interna o capsula tiroidea emite tabiques compuestos por fibras
reticulares, capilares sanguíneos fenestrados, capilares linfáticos y nerviosos, que dividen a la glándula en
lobulillos, estos últimos contienen estructuras vesiculares que miden 0.2 a 1 mm llamados folículos tiroideos
que son unidades funcionales de la glándula, que están revestidos por epitelio cubico simple cuya cavidad
contiene coloide donde la glándula almacena su producto de secreción, el coloide es de consistencia viscosa,
se tiñe con colorantes ácidos y básicos. Los folículos están rodeados de tejido conectivo laxo muy vascularizado.
Se pueden observar 2 tipos celulares, las células foliculares que son células cubicas, de citoplasma basófilo,
núcleo ovoide, RER y complejo de Golgi supranuclear y microvellosidades en la membrana apical, presenta

lisosomas, mitocondrias filamentosas y vesículas, se mantienen unidas entre si por complejos de unión y
delimitan el folículo, producen triyodotironina y tiroxina, están en contacto con la luz del folículo; y las células
parafoliculares o células C que son células que se encuentran intercaladas entre los folículos pero no se
relacionan con el interior de los folículos ni tienen contacto con el coloide, son más grandes y están en menor
número que las foliculares, nunca entran en contacto con la luz del folículo, aparecen aisladas o en grupos
pequeños de 3 o 4 células, tienen forma ovalada, núcleo esférico, complejo de Golgi prominente y citoplasma
claro, con vesículas esféricas que contienen la hormona calcitonina que regula el metabolismo del calcio. La
glándula tiroides es la única glándula que tiene reservorio extracelular.
Fisiología: la tiroides produce 3 tipos de hormona, la triyodotironina o T3, la tiroxina o T4, y la calcitonina
Triyodotironina y tiroxina
• Formación y secreción: la secreción tiroidea depende de la producción de TSH, se secreta 90% de T4 y 10%
de T3.
✓ Etapas de la formación de T3 y T4:
❖ Captación de yoduro: la membrana basal de las células foliculares tiroideas posee una bomba de
yoduro que transporta el yodo activamente al interior de la célula.
❖ Formación y secreción de tiroglobulina: el RER y el Aparato de Golgi sintetizan y secretan una
glucoproteína (tiroglobulina) tiene 140 aa tirosina, forman la T3 y T4.
❖ Oxidación de yoduro: yoduro es oxidado por una peroxidasa que se encuentra en la membrana apical
o en el citoplasma cercano, es la encargada de proporcionar yodo oxidado en el sitio donde emerge la
molécula de tiroglobulina del Aparato de Golgi.
❖ Iodacion de tirosina y formación de T3 y T4, organificación de tiroglobulina: yodo oxidado se une al
aa tirosina, forma monoiodotirosina y después diyodotirosina por la adicción de otro yodo. Acoplamiento
de 2 diyodotirosina forman la T4 y de monoiodotirosina y diyodotirosina forman la T3.
❖ Almacenamiento de tiroglobulina: cada molécula de tiroglobulina contiene 5 o 6 moléculas de T4 y un
promedio de T3 de 1/10 con respecto a la T4.
✓ Liberación de T3 y T4: la superficie apical de las células foliculares encierra por pinocitosis vesículas de
coloide, luego se fusiona con los lisosomas quienes van a liberar T3 y T4 por acción de una proteinasa,
estas hormonas van a difundir por la membrana basal a los capilares, el 90% de hormona liberada es T4 y
10% T3, luego mientras circulan por sangre pequeñas cantidades de T4 pierden un yodo y se transforman
en T3.
• Transporte de T3 y T4: las hormonas tiroideas se combinan con proteínas plasmáticas para su transporte a
los diferentes tejidos, de la siguiente manera: 2/3 se combina con globulina fijadora de T4, 1/4 con prealbúmina
fijadora de T4, 1/6 con albúmina, y la T3 es transportada por una globulina y la albumina
Las proteínas transportadoras tienen una gran afinidad con las hormonas tiroideas, por esta razón se liberan
hacia los tejidos con lentitud. Cuando estas hormonas quedan libres, entran en las células blanco y se fijan a
proteínas intracelulares. De esta manera quedan almacenadas para luego ejercer su acción en forma lenta
durante días o semanas. La T4 tiene un período de latencia antes de iniciar su actividad de 2 a 3 días, luego
su acción aumenta en forma progresiva, alcanzando su máxima en 10 a 12 días, y persistiendo hasta 6
semanas. En cambio, la T3 tiene un período de latencia más corto, de 6 a 12 horas. Y su actividad máxima se
observa en 2 a 3 días.
• Mecanismo de acción: las hormonas tiroideas ejercen su acción una vez que se introducen al interior de la
célula, donde penetran al núcleo para unirse a receptores específicos, que a su vez se fijarán a secuencias
específicas del ADN para regular la síntesis de diversas proteínas. Además de esta acción central, activa las
mitocondrias produciendo un efecto calorígeno, probablemente a través de la unión de estas hormonas a una
proteína mitocondrial. También tienen un efecto primario sobre la membrana plasmática, regulando el flujo
transcelular de cationes y sustratos
• Efectos de las hormonas tiroideas en los tejidos, las hormonas tiroideas presentan dos grandes funciones
aumento del índice metabólico y estimulación del crecimiento y maduración de los tejidos.
✓ Aumento del índice metabólico: la T3 y T4 aumentan las actividades metabólicas en casi todos los tejidos
excepto cerebro, retina, bazo, testículo y pulmón.
❖ Efectos sobre las proteínas aumenta la síntesis proteica en casi todos los tejidos por una estimulación
del proceso de traducción (formación de proteínas por los rizomas) y un aumento de la transcripción de
ADN a ARN, que origina un incremento generalizado de la síntesis de casi todos los tipos de proteínas
celulares
❖ Efecto sobre los sistemas enzimáticos al aumentar la síntesis proteica se incrementa el pool de
enzimas intracelulares.
❖ Efecto sobre las mitocondrias aumenta del número, tamaño y superficie total de las membranas de
las mitocondrias en forma proporcional al incremento del metabolismo basal.
❖ Efecto sobre el transporte de iones: intensifica la actividad de la Na-K ATPasa. Esta enzima a su vez
estimula el transporte de Na-K a través de la membrana celular.
✓ Estimulación del crecimiento se manifiesta sobre todo en la infancia, fomenta el desarrollo del cerebro
en la vida fetal y los primeros años de vida. Este efecto estimulante depende posiblemente de su capacidad
para promover la síntesis de proteínas.
✓ Efectos sobre el metabolismo de los glúcidos aumenta la captación de glucosa por las células, aumenta
la glucolisis y la gluconeogénesis, aumenta la absorción intestinal de glucosa, incrementa la secreción de
insulina
✓ Efectos sobre el metabolismo de los lípidos moviliza los lípidos almacenados en el tejido adiposo,
aumenta la concentración de ácidos grasos libres en sangre, acelera la betaoxidación, disminuye los niveles
de colesterol, fosfolípidos y triglicéridos en sangre.
✓ Efectos sobre el metabolismo de vitaminas como produce un incremento de las enzimas y las vitaminas
son constituyentes de sistemas enzimáticos, se acrecientan las necesidades de vitaminas
✓ Aumenta el metabolismo basal en la mayor parte de los tejidos, excepto en cerebro, retina, bazo,
testículos y pulmón.
✓ Efecto sobre el peso corporal: en el hipertiroidismo disminuye el peso, en cambio el hipotiroidismo se
aumenta de peso.
✓ Efectos sobre el aparato cardiovascular: acrecienta el flujo sanguíneo por un incremento de la utilización
de oxígeno a nivel tisular, en el hipertiroidismo se incrementa el gasto cardíaco, de la frecuencia cardíaca
y de la fuerza de contracción del miocardio. También se incrementa el volumen sanguíneo y la presión
arterial diferencial (gran amplitud entre presión arterial sistólica y diastólica).
✓ Efectos en aparato respiratorio: acrecienta la utilización de oxígeno y la formación de dióxido de carbono,
y aumento de la frecuencia y la profundidad de la ventilación.
✓ Efectos en el tubo digestivo: aumenta la absorción de nutrientes a nivel intestinal, e incrementa la
motilidad y las secreciones digestivas.
✓ Efectos sobre el S.N.C.: acelera la encefalización, en el hipertiroidismo: mayor excitabilidad y
nerviosismo.
✓ Efectos sobre el tejido muscular: en el hipertiroidismo: temblor fino por mayor excitabilidad.
✓ Efectos sobre el sueño: en el hipertiroidismo insomnio a pesar del cansancio, y en el hipotiroidismo
aumento de las horas de sueño.
✓ Efectos sobre el sistema endócrino incrementa la secreción de la mayoría de las glándulas endócrinas y
también las necesidades de las hormonas en los tejidos.
✓ Efectos sobre el aparato reproductor: en el hombre el hipotiroidismo produce pérdida de la libido. El
hipotiroidismo puede ocasionar impotencia, y en la mujer hipertiroidismo e hipotiroidismo ocasionan
trastornos menstruales.

• Regulación de la secreción: las hormonas tiroideas deben secretarse de manera constante y en la cantidad
adecuada para conservar la normalidad de la actividad metabólica. Esto es posible gracias a la regulación por
retroalimentación negativa que ejercen el hipotálamo y la hipófisis.
Efectos de la T.S.H, la tirotrofina es una hormona de naturaleza glicoproteica que incrementa la secreción deT3
y T4. Sus efectos son aumenta la proteólisis de la tiroglobulina y libera T3 y T4, aumenta la actividad de la
bomba de yoduro, aumento de lodación de tirosina y formación de hormona, aumento del tamaño y secreción
de las células tiroideas, aumento del número de células tiroideas. La T.S.H. incrementa todas las actividades
de las células foliculares de la tiroides. El hipotálamo estimula la secreción de T.S.H. a través del Factor
Liberador de T.S.H. o T.R.H. Este factor actúa directamente sobre las células hipofisarias que secretan T.S.H.
Estimulan la liberación de T.R.H el frío y las emociones.
Calcitonina: es una hormona polipeptídica, compuesta por 32 aminoácidos, secretada por las células
parafoliculares de la tiroides.
• Efectos de la calcitonina es una hormona que disminuye la calcemia a través de los siguientes mecanismos
efecto inmediato disminuye la osteólisis, efecto mediato (en 1 hora) aumenta la actividad osteoblástica, efecto
prolongado disminuye la formación de osteoclastos; en el riñón tiene un efecto menor, contrario al de la PTH;
los efectos de la calcitonina en el adulto sobre la concentración de Ca son débiles, en cambio en el niño es
más intenso.
• Regulación de la calcitonina un incremento de la concentración plasmática de Ca produce un aumento de la
secreción de calcitonina pero este mecanismo tiene dos diferencias con el mecanismo de regulación de PTH
el mecanismo calcitonina es más rápido, y el mecanismo calcitonina es regulador a corto plazo de la calcemia.
Glándula paratiroides: son 4 pequeñas glándulas de color amarillo castaño y de forma circular aplanada,
ubicadas en la parte posterior de la glándula tiroides, adosada al cuerpo de los lóbulos tiroideos en sus caras
posterointernas, se disponen en 2 paratiroideas superiores y 2 paratiroideas inferiores a la izquierda y a la
derecha. Cada glándula esta cubierta por una capsula delgada de tejido conectivo que envía tabiques hacia su
interior que la dividen en lobulillos. El parénquima esta formado por cordones entre ellos el estroma conectivo
con fibras reticulares, capilares sanguíneos fenestrados, capilares linfáticos, nervios y células adiposas. Hay 2
tipos celulares, las células principales que son las más abundantes, pequeñas, de forma poligonal, con núcleo
vesiculoso y central, rodeado de citoplasma levemente acidófilo. Tienen a cargo la regulación de la síntesis,
almacenamiento y secreción de la hormona paratiroidea o parathormona (PTH). Cuando están en actividad,
presentan aumentadas las mitocondrias, RER, complejo de Golgi y vesículas de secreción, que contienen la
PTH. Las células principales poseen receptores sensibles al calcio que detectan la baja concentración de calcio
sérico y aumentan la secreción de PTH; por el contrario, la alta concentración de calcio sérico inhibe su
secreción; y las células oxífilas que son más grandes que las células principales, pero en menor número,
aisladas o formando grupos. Poseen citoplasma eosinófilo, escaso RER y complejo de Golgi, mitocondrias y
gránulos de glucógeno. No poseen vesículas de secreción.
Irrigación: las arterias tiroideas superiores irrigan las glándulas paratiroideas superiores y las arterias tiroideas
inferiores a las glándulas paratiroideas inferiores. Son muy voluminosas con relación al tamaño de las glándulas
que irriga. Las venas y linfáticos son tributarias de las venas tiroideas correspondientes.

Metabolismo de calcio y fosforo
• Hipocalcemia: produce excitabilidad a nivel del sistema nervioso por aumento de la permeabilidad neuronal
al Na, también tetania y muerte.
• Hipercalcemia: deprime el sistema nervioso, altera la contractibilidad cardíaca, estreñimiento y disminución
del apetito.
• La absorción de calcio en el tubo digestivo es escasa porque la mayoría de los compuestos son insolubles y
los cationes bivalentes son difíciles de absorber. La vitamina D produce un incremento en la absorción de Ca.
• La eliminación de Calcio se hace fundamentalmente a través de las heces (7/8 partes), el resto por orina. El
Calcio se reabsorbe a en la nefrona a nivel del túbulo proximal y asa de Henle. Puede ocurrir una resorción
adicional de Calcio en los túbulos contorneados distales y colectores que es selectiva y depende de las
concentraciones de Calcio (Ca) en sangre y de la hormona paratiroidea.
• Las modificaciones en la concentración de fósforo (P) inorgánico plasmática y en líquido extracelular no
producen efectos inmediatos importantes en el organismo.
• El P se absorbe fácilmente en intestino delgado, salvo cuando la alimentación es rica en Ca y se forma
Ca3(PO4)2que es prácticamente insoluble y se elimina en heces.
• La eliminación de fósforo se realiza por orina y depende de la concentración plasmática, a mayor
concentración mayor excreción. La parathormona interviene en su excreción.

• Efectos de la Vitamina D
✓ Sobre la absorción de Ca el 1,25 (OH) D3 funciona como una hormona al fomentar la absorción intestinal
de Ca por los siguientes mecanismos: forma una proteína fijadora de Ca en las células epiteliales
intestinales que funciona en las microvellosidades, para transportar Ca al citoplasma, el Ca pasa luego al
líquido extracelular por la membrana basolateral, la intensidad de absorción depende de la concentración
de esta proteína y su efecto persiste durante varias semanas; forma una ATPasa estimulada por Ca; y
formación de fosfatasa alcalina en las células epiteliales intestinales.
✓ Sobre la absorción de P es de menor importancia que la anterior por que el P se absorbe con facilidad
en intestino.
✓ Sobre el tejido óseo en pequeñas cantidades fomenta la calcificación ósea, en cantidades excesivas
produce una resorción ósea semejante al efecto de la PTH y en ausencia de esta hormona reduce e
inclusive impide el efecto de resorción.
• Regulación de la formación de 1,25 (OH)2 D3. La conversión de 25 (OH) D3 en 1,25 (OH) D3 en riñón
requiere de PTH. Esta hormona es secretada por las glándulas paratiroideas cuando disminuye la calcemia.
Como ya se describió, la PTH estimula la formación del principio activo de vitamina D y por lo tanto aumenta
la absorción de Ca a nivel intestinal. Este efecto junto con otros de la PTH restablece la calcemia normal
Parathormona: La parathormona tiene una naturaleza química proteica y está compuesta por 84 aminoácidos.
• Efectos de PTH: esta hormona aumenta la calcemia por los siguientes mecanismo
✓ Aumenta la resorción .de Ca y P en hueso a través de tres acciones que se ejecutan en diferentes
tiempos, fase rápida (en minutos) produce osteoliosis, que es la remoción de sales óseas desde la matriz
ósea vecina a los osteocitos y osteoblastos sin resorción de la matriz, y fase lenta (de semanas a meses):
inicia una activación y aumento del número de los osteoclastos que digieren la matriz ósea. También
disminuye la actividad de los osteoblastos.
✓ Disminuye la excreción de Ca y aumenta la excreción de P por orina produce una pérdida rápida de P
por orina por una disminución de la resorción tubular de éste. En cambio, aumenta la resorción de Ca en
túbulos distales y colectores.
✓ Aumenta la absorción de Ca a nivel del intestino delgado por un efecto indirecto a través de la vitamina
D.
✓ Sobre la vitamina D estimula la transformación en riñón de 25 (OH)D3 en 1,25(OD)D3, principio activo
de la vitamina D.
• Regulación de la secreción de PTH La disminución de la calcemia estimula la paratiroides para secretar PTH.
También aumenta su secreción el embarazo y la lactancia. Un aumento de las concentraciones séricas de
calcio inhibe la secreción de PTH. La hipercalcemia puede ser originada por un exceso de Ca en la dieta, un
aumento de vitamina D en la alimentación o un incremento de la resorción ósea.
Glándulas suprarrenales
Anatomía macroscópica: las glándulas suprarrenales son dos órganos endócrinos
bilaterales, uno izquierdo y otro derecho, ubicados en la región posterosuperior del
abdomen, arriba de cada riñón donde se asientan, limitadas atrás y arriba por el
diafragma, aplicadas sobre la cara anteroexterna de la porción superior de la
columna lumbar (delante y afuera de las vértebras D12 y L1), son de color castaño
amarillento y pesan en conjunto alrededor de 12 g, cada suprarrenal presenta forma
diferente, la derecha tiene forma triangular más definida, mientras que la izquierda
adapta una forma semilunar. En las dos glándulas se observa, una base cóncava
que se aplica sobre el polo superior de cada riñón; dos caras, una anteroexterna
donde se encuentra el hilio y una posteromedial en relación con los cuerpos
vertebrales y el diafragma; y un borde medial interno, que mira hacia la columna

lumbar y los grandes vasos (vena cava inferior y aorta abdominal), llegando abajo hasta el pedículo renal
correspondiente. La cara anteroexterna de las glándulas es tapiza por peritoneo. La suprarrenal izquierda es
retrogástrica (ubicada detrás del estómago); adelante responde a la transcavidad de los epiplones donde se
relaciona con el páncreas y a veces con el bazo (borde posterior), la derecha hace lo propio con la cara posterior
del hígado
Relaciones: afuera con el polo superior del riñón, adentro con la columna vertebral y vasos (a la izquierda la
arteria aorta; a la derecha la vena cava inferior), adelante con el hígado (derecha) y estómago (izquierda), abajo
con el pedículo renal, y atrás y arriba con la pared posterior del diafragma.
Medios de fijación: las glándulas suprarrenales mantienen su posición gracias a la fascia prerrenal por delante
de la glándula hasta el diafragma, y la fascia retrorrenal por detrás de la glándula hasta el diafragma. La glándula
no sigue al riñón en sus desplazamientos.
Vascularización
Arterias: las adrenales reciben sangre de tres principales ramas superiores o capsulares que son rama de la
arteria diafragmática inferiores y ésta a su vez de la aorta abdominal, abordan las glándulas por su parte
superior, penetrando la capsula de tejido conectivo que las cubre dando dos tipos de vasos, los capilares
corticales, que irrigan las capas de la corteza y las arterias medulares que descienden directamente hasta la
médula donde recién se capilarizan (capilares medulares); mediales que son rama directas de la aorta
abdominal ingresan a las suprarrenales por el hilio, llegando a la médula; las inferiores que son rama de la
arteria renal, son las más voluminosas, alcanzan la glándula por su parte inferior y se distribuyen de igual
manera a las arterias suprarrenales superiores; y por ultimo las accesorias que provienen de las arterias
lumbares, renales, gonádicas o perirrenales.
Venas: son dos los drenajes venosos en estas glándulas, las más importantes son las venas centrales que
recogen casi la totalidad de sangre con los principales productos metabólicos, emergiendo del hilio alcanzando
la vena cava inferior, directamente a la derecha y por intermedio de la vena renal, a la izquierda; y las venas
periféricas son pequeñas y terminan en las venas frénicas en ambos casos.
Inervación: la inervación de estas glándulas es proporcionalmente mayor que la de otros órganos y lógicamente
corresponde al SN autónomo, la mayoría provienen del nervio esplénico mayor que corresponde a la vía
simpática, son fibras mielinizadas preganglionares que llegan a la medula para hacer sinapsis con las células
cromafines.

Anatomía microscópica: las glándulas están cubiertas por una cápsula de tejido conectivo (TC) que las rodea y
emite tabiques al interior conformando el estroma de las suprarrenales. En el parénquima se distinguen dos
porciones de origen embriológico diferente, la corteza es mesodérmica y la médula deriva del ectodermo. La
corteza (periferia de la glándula), representa la mayor parte de la glándula, compuesta por cordones epiteliales
en distinta disposición que permite distinguir tres capas de afuera hacia adentro son capa glomerular,
fasciculada y reticular, que elaboran hormonas esteroideas diferentes. La capa glomerular está constituida por
cordones celulares plegados sobre sí mismos formando cordones arqueados o grupos ovoides, entre los que
se dispone la red capilar, las células presentan un núcleo bien basófilo mientras que el citoplasma es acidófilo,
REL muy desarrollado, gotas de lípidos pequeñas, mitocondrias y cúmulos de RER. Elaboran
mineralocorticoides (aldosterona y desoxicorticosterona), hormonas que controlan el metabolismo mineral a
través del aumento de la reabsorción de Sodio y de la excreción de Potasio. Esta capa tiene gran poder
regenerativo a partir del cual puede recomponer la masa celular de las capas fasciculada y reticular. La capa
fasciculada es la más ancha y sus células poliédricas forman cordones organizados en columnas de a dos
células de espesor, con disposición radial, entre las que se transcurren largos capilares fenestrados, el núcleo
de las células es bastante claro, el citoplasma es débilmente basófilo o acidófilo, contiene un REL muy
desarrollado y, posee abundantes inclusiones de lípidos que le otorgan un aspecto esponjoso, las mitocondrias
presentan un aspecto particular con crestas largas tubulares, complejo de Golgi bien desarrollado asociado a
cúmulos de RER. En esta capa se producen glucocorticoides (cortisol, cortisona, hidrocortisona) que afectan el
metabolismo de los hidratos de carbono, proteínas y lípidos. La capa reticular está formada por hileras de
célula que se anastomosan como una red entre capilares fenestrados, poseen escasas y pequeñas gotas de
lípidos, su citoplasma es eosinófilo y en el límite con la médula presentan células oscuras con un núcleo
pequeño y cromatina condensada. Elaboran hormonas sexuales (estrógenos y andrógenos) en escasa
cantidad. Las características en común que presentan las células de estas tres capas radican en que todas las
hormonas que producen son esteroideas, se producen a partir de colesterol que en gran parte captan de
lipoproteínas plasmáticas, gracias a receptores proteicos de membrana (una pequeña cantidad lo pueden
sintetizar las propias células), lo almacenan para luego metabolizarlo en el REL y las mitocondrias. La médula
representa aproximadamente el 10 % de la glándula, está constituida por trabéculas epiteliales que rodean un
rico entramado vascular, formado por capilares medulares y vénulas que provienen de los capilares corticales,
para drenar finalmente en la vena central, las células epiteliales tienen un citoplasma levemente basófilo con
gránulos limitados por membrana, que contienen las catecolaminas (adrenalina; 90% y noradrenalina 10 %),
estos se observan oscuros al tratar con una solución de sales de cromo por lo que son células cromafines,
abundante RER y aparato de Golgi bien desarrollado. Encontramos células ganglionares simpáticas aisladas o
agrupadas pero que son difíciles de distinguir con métodos comunes de coloración.
Este documento contiene más páginas...
Descargar Completo
Hormonas.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.