SISTEMA GENITAL
FEMENINO
GENERALIDADES
Los órganos reproductores femeninos internos están localizados en la pelvis, mientras que las estructuras genitales externas están
situadas en la parte anterior del periné y en conjunto reciben el nombre de vulva.
o Los órganos genitales internos son los ovarios, tubas uterinas, el útero y la vagina. Están situados sobre todo en la cavidad pélvica
y en el periné.
o Los órganos genitales externos incluyen el monte del pubis, los labios mayores y menores, el clítoris, el vestíbulo y el orificio de
la vagina, el himen y el orificio uretral externo.
Los ovarios, las tubas uterinas y el útero de una mujer sexualmente madura sufren cambios estructurales y funcionales relacionados
con la actividad nerviosa y las modificaciones de la concentración de las hormonas durante cada ciclo menstrual y durante el embarazo.
Estos mecanismos también regulan la embriogénesis del sistema genital femenino.
El inicio del ciclo menstrual, denominado menarca, ocurre entre los 9 y 14 años de edad y marca el final de la pubertad y el inicio de
la vida reproductiva. Durante esta fase de la vida, el ciclo menstrual tiene una duración promedio de entre 28 y 30 días. Entre los 45 y
55 años, el ciclo menstrual se torna infrecuente y finalmente cesa, la menopausia. Los ovarios dejan de producir ovocitos y detienen
su función endocrina de producción de hormonas que regulan la actividad endocrina. Otros órganos (vagina y glándulas mamarias)
disminuyen sus funciones, en particular la actividad secretora.
OVARIO
Los ovarios poseen dos funciones interrelacionadas: la gametogénesis y la esteroidogénesis. En las mujeres, la producción de gametos
se denomina ovogénesis. Los gametos en desarrollo se conocen como ovocitos y los gametos maduros se denominan óvulos.
Los ovarios secretan dos grupos principales de
hormonas esteroideas:
o Estrógenos: promueven el crecimiento y la
maduración de los órganos sexuales externos e
internos y producen las características sexuales
femeninas que se desarrollan en la pubertad.
También actúan sobre las glándulas mamarias
para promover el desarrollo de las mamas al
estimular el crecimiento de los conductos y el
estroma y la acumulación de tejido adiposo.
o Progestágenos: preparan los órganos sexuales
internos, sobre todo el útero, para el embarazo al
promover cambios secretores en el endometrio.
También preparan la glándula mamaria para la lactación al promover la proliferación de los lobulillos.
ESTRUCTURA OVÁRICA
En las nulíparas, los ovarios son estructuras
áreas blanco rosadas con forma de
almendra, miden unos 3cm de longitud,
1,5cm de ancho y 1cm de espesor. Cada
ovario está fijado a la superficie posterior del
ligamento ancho del útero a través de un
pliegue peritoneal, el mesoovario. El polo
superior (tubárico) del ovario está unido a la
pared de la pelvis mediante el ligamento
suspensorio del ovario, que conduce los
vasos y nervios. El polo inferior (uterino) está
unido al útero mediante el ligamento
ovárico. Éste es un resto del ligamento
genital caudal, cordón fibroso embrionario
que fija a la gónada en desarrollo al piso de
la pelvis. Antes de la pubertad, la superficie
del ovario es lisa, pero durante la vida
reproductiva adquiere cada vez más
cicatrices y se torna irregular debido a
las ovulaciones repetidas.
El ovario está compuesto por dos
regiones:
o La médula (región medular):
ubicada en la porción central
del ovario y contiene tejido
conectivo laxo, una masa de
vasos sanguíneos tortuosos
bastante grandes, vasos
linfáticos y nervios.
o La corteza (región cortical):
se encuentra en la porción
periférica del ovario y rodea
a la médula. Contiene los
folículos ováricos incluidos
en un tejido conjuntivo
celular. En el estroma que
rodea los folículos hay fibras
de músculo liso.
La superficie del ovario está cubierta por una capa individual de células cúbicas y, en algunas partes, casi planas. Esta capa celular se
conoce como epitelio germinativo y se continúa con el mesotelio que tapiza el mesoovario. Una capa de tejido conjuntivo denso, la
túnica albugínea, se localiza entre el epitelio germinativo y la corteza subyacente.
Los folículos ováricos de varios tamaños, cada uno con un solo ovocito, están distribuidos en el estroma de la corteza. El tamaño del
folículo indica el estado de desarrollo del ovocito. Los ovocitos presentes en el nacimiento permanecen detenidos en la primera
división meiótica de su desarrollo. Durante la pubertad, pequeños grupos de folículos sufren crecimiento y maduración cíclicos. La
primera ovulación, generalmente no ocurre hasta después de pasado un año de la menarca o incluso más tiempo. Luego se establece
un patrón cíclico de maduración folicular y ovulación que continúa en paralelo con el ciclo menstrual.
La mayoría de los 600.000a 800.000 ovocitos primarios que se estima están presentes al momento del nacimiento no completan la
maduración y se pierden en forma gradual a través de la atresia, muerte espontánea y reabsorción de los ovocitos inmaduros. Este
proceso comienza en el 5to mes de la vida fetal y es mediado por la apoptosis de células que rodean al ovocito.
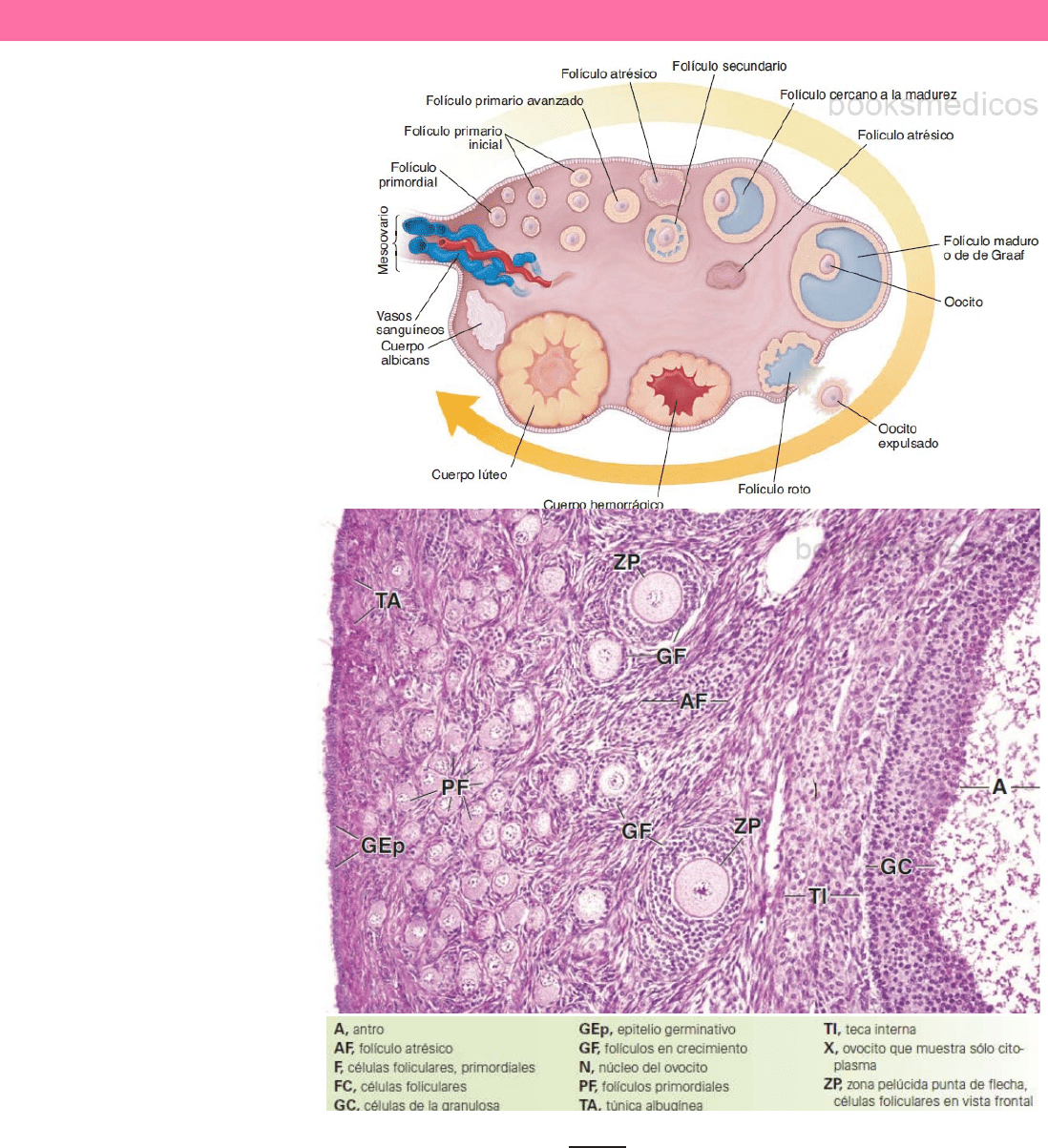
DESARROLLO FOLICULAR
Desde el punto de vista histológico, pueden identificarse 3 tipos básicos de
folículos ováricos, según su estado de desarrollo:
o Folículos primordiales.
o Folículos en crecimiento, que a su vez se subclasifican en folículos
primarios y secundarios.
o Folículos maduros o Folículos de de Graff.
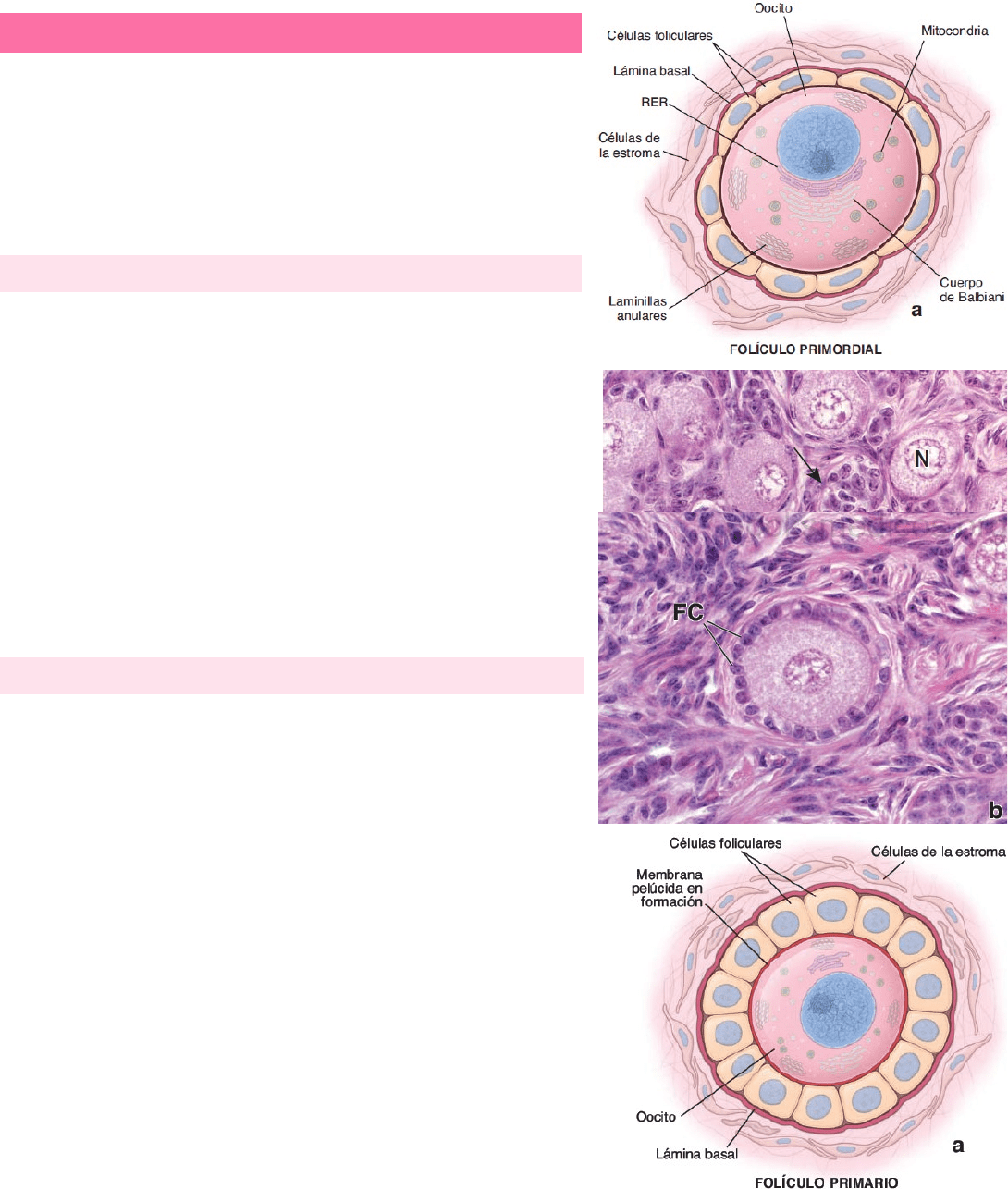
FOLÍCULOS PRIMORDIALES
Aparecen en el ovario durante el tercer mes de desarrollo fetal. El
crecimiento inicial de los folículos primordiales es independiente de la
estimulación por gonadotrofinas. En el ovario maduro, los folículos
primordiales se encuentran en el estroma de la corteza justo debajo de la
túnica albugínea. Una capa simple de células foliculares planas (F) rodea el
ovocito. La superficie externa de las células foliculares está limitada por una
lámina basal. El ovocito en el folículo posee un núcleo voluminoso que
contiene cromatina dispersa y uno o más nucléolos. El citoplasma del ovocito
(ovoplasma) contiene un cuerpo de Balbiani, que consiste en una
acumulación focalizada de vesículas y membranas de Golgi, retículo
endoplásmico, centriolos, mitocondrias y lisosomas. Los ovocitos humanos
contienen laminillas anulares y vesículas dispersas a lo largo del ovoplasma
junto con pequeñas mitocondrias.
FOLÍCULOS EN CRECIMIENTO: PRIMARIO
A medida que el folículo primordial se desarrolla en un folículo en
crecimiento, ocurren cambios en el ovocito, en las células foliculares y en el
estroma contiguo. El ovocito aumenta de tamaño y las células foliculares
aplanadas circundantes proliferan y se tornan cúbicas, es en este momento
donde el folículo recibe el nombre de folículo primario. A medida que el
ovocito crece, secreta proteínas específicas que se ensamblan en la zona pelúcida
(ZP) (cubierta extracelular), que aparece entre el ovocito y las células foliculares
contiguas. La zona pelúcida está compuesta por 3 clases de glucoproteínas de la
zona pelúcida, denominadas ZP-1, ZP-2 y ZP-3. La ZP1 no se sabe la función, la
ZP2 actúa como proteína secundaria para la unión del espermatozoide y la ZP3
(más importante) actúa como receptora para la unión del espermatozoide y como
inductora de la reacción acrosómica.
A través de una proliferación mitótica rápida, la capa simple de células foliculares
da origen a un epitelio estratificado, la membrana granulosa, que rodea al
ovocito. Las células foliculares ahora se identifican como células de la granulosa.
La lámina basal mantiene su posición entre la capa más externa de las células
foliculares, que se tornan cilíndricas, y el estroma de tejido conjuntivo. Durante el
crecimiento folicular, se desarrollan uniones de hendidura entre las células de la
granulosa.
A medida que las células de la granulosa proliferan, las células del estroma
perifolicular forman una vaina de células de tejido conjuntivo, conocida como teca
folicular, justo por fuera de la lámina basal.
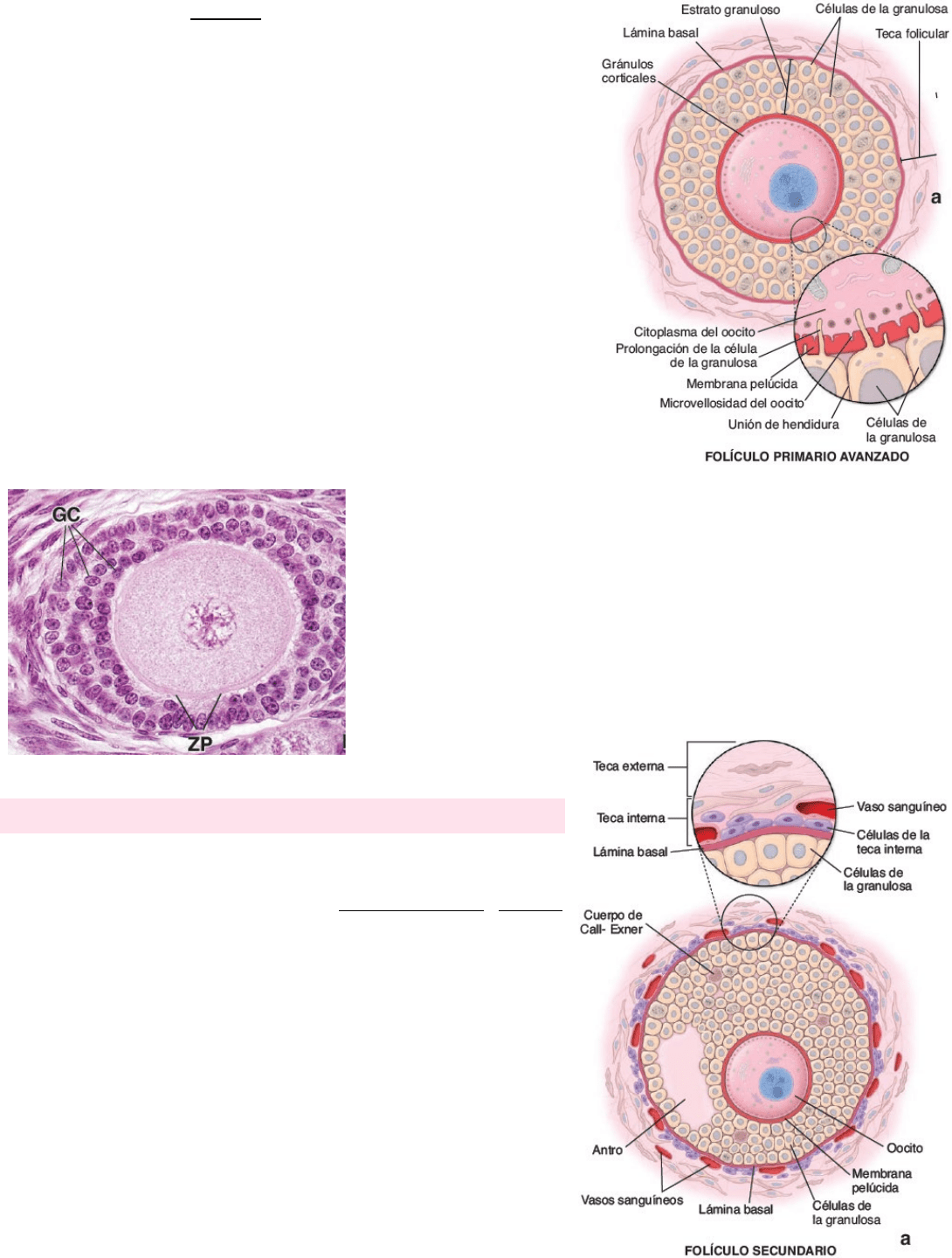
La teca folicular se diferencia en dos capas:
o Teca interna: capa interna, muy vascularizada, de células secretoras
cúbicas. Las células de la teca interna con diferenciación completa
poseen características típicas de células productoras de esteroides. Estas
células poseen una gran cantidad de receptores para la LH. En respuesta
a la estimulación por LH, sintetizan y secretan a los andrógenos
precursores de los estrógenos. La teca interna contiene fibroblastos,
haces de fibras colágenas y una red extensa de vasos pequeños (típico de
órganos endocrinos).
o Teca externa: capa más externa de células de tejido conjuntivo. Solo
contiene células de músculo liso y haces de fibras de colágeno.
La distribución de los orgánulos cambia a medida que el ovocito madura. Elementos
de Golgi que derivan del cuerpo del Balbiani y se dispersan en el citoplasma;
ribosomas, mitocondrias, vesículas pequeñas y cuerpos multivesiculares y la
cantidad de RER aumenta. Los ovocitos exhiben vesículas secretoras especializadas
conocidas como gránulos corticales, ubicados bajo el ovolema (MP del ovocito).
Estos gránulos contienen proteasas que se liberan por exocitosis cuando el óvulo es
activado por el espermatozoide. Muchas microvellosidades se proyectan desde el
ovocito hacia el espacio perivitelino entre el ovocito y las células de la granulosa
circundantes conforme se deposita la zona pelúcida. Al mismo tiempo, las células de
la granulosa desarrollan evaginaciones que se proyectan hacia el ovocito y se
entremezclan con las
microvellosidades ovocíticas.
FOLÍCULOS EN CRECIMIENTO: SECUNDARIO
El folículo primario inicialmente se hace más profundo en el estroma cortical a
medida que incrementa su tamaño, sobre todo por proliferación de las células
de la granulosa. Los factores necesarios para el crecimiento folicular y ovocítico
son:
o FSH
o Factores de crecimiento (epidérmico, IGF)
o Iones calcio (Ca2+)
Cuando el estrato granuloso alcanza un espesor de 6 a 12 capas celulares, entre
las células de la granulosa aparecen cavidades con contenido líquido y
hialuronato, llamado líquido folicular. A medida que este líquido se acumula
entre las células, las cavidades comienzan a confluir para formar una cavidad
con forma de semiluna: el antro.
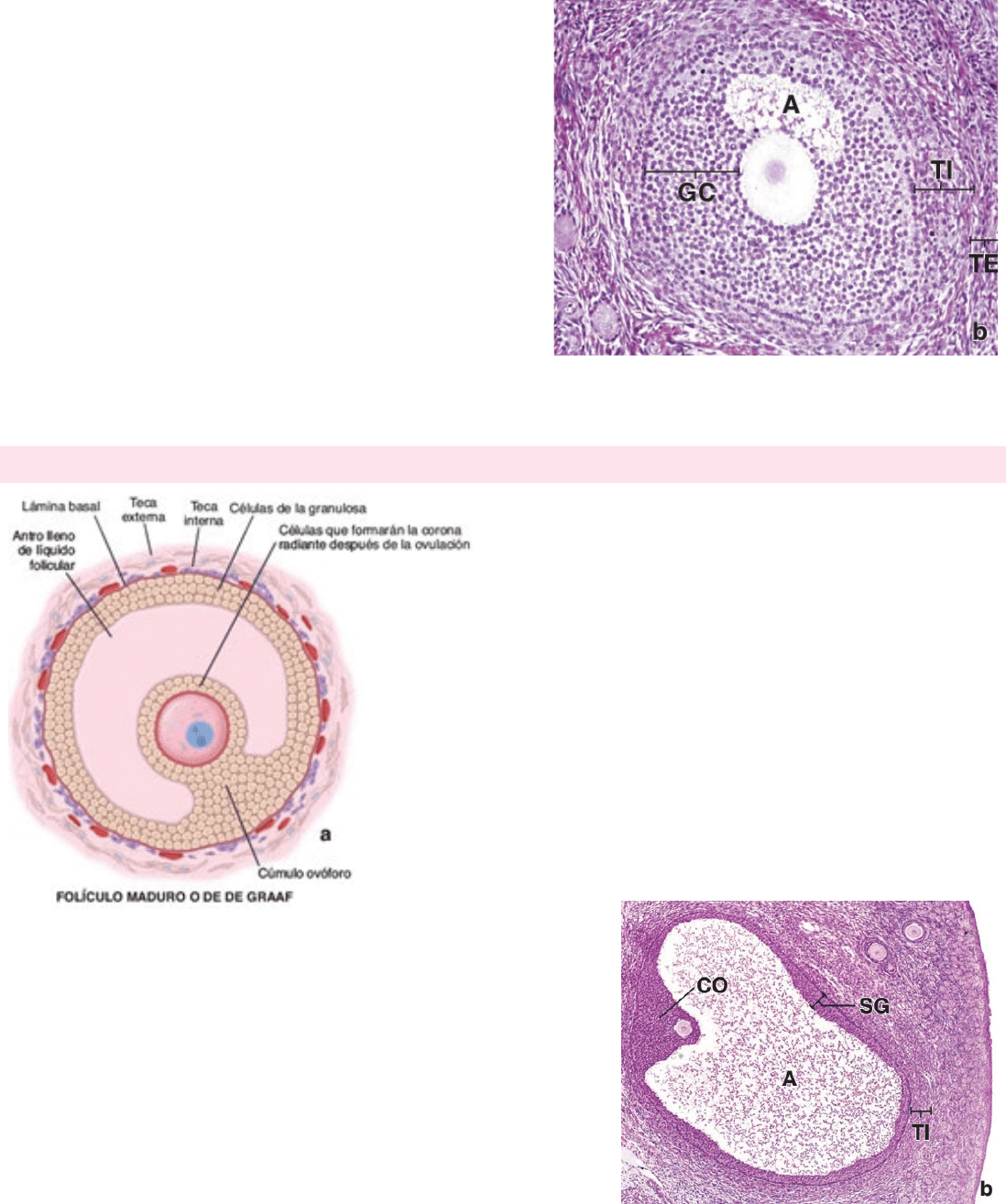
Este folículo ahora se denomina folículo secundario o folículo antral.
Ahora el ovocito ya no crece más gracias a la inhibición de su
crecimiento por el péptido inhibidor de la maduración ovocítica (OMI),
secretado por las células de la granulosa hacia el líquido antral.
Conforme el folículo secundario incrementa su tamaño, el antro,
revestido por varias capas de células de la granulosa, también se
agranda. El estrato granuloso posee un espesor uniforme excepto en la
región asociada al ovocito. Aquí, las células de la granulosa forman un
montículo abultado, el cúmulo ovóforo, que se proyecta dentro del
antro. Las células del cúmulo ovóforo que rodean al ovocito,
permanecen con él en la ovulación y forman la corona radiante. Ésta se
compone por células del cúmulo que envían microvellosidades
penetrantes a lo largo de la zona pelúcida para comunicarse a través de
las uniones de hendidura con las microvellosidades del ovocito. Durante
la maduración folicular, la cantidad de microvellosidades en la superficie de las células de la granulosa incrementan y se correlaciona
con un aumento en los receptores de LH en la superficie libre antral.
FOLÍCULOS MADUROS (DE GRAAF)
Debido a su gran tamaño, se extiende a través de todo el espesor de
la corteza ovárica y sobresale en la superficie del ovario. A medida
que el folículo se acerca a su tamaño máximo, la actividad mitótica
de las células de la granulosa disminuye. El antro y los espacios entre
las células de la granulosa se agrandan, el ovocito y las células del
cúmulo se separan del resto de las células de la granulosa en
preparación para la ovulación. Las células del cúmulo que rodean
inmediatamente al ovocito ahora forman la capa de células simple de
la corona radiante. Estas células y las células del cúmulo adheridas
con laxitud permanecen con el ovocito en la ovulación.
La LH estimula a las células de la teca interna para que secreten
andrógenos. Debido a la falta de la enzima aromatasa, las células de
la teca interna no son capaces de producir estrógenos. En cambio, las
células de la granulosa contienen esta enzima y, por ende, algunos
andrógenos son transportados a sus REL para un procesamiento
adicional. En
respuesta a la FSH, las células de la granulosa catalizan la conversión de los
andrógenos en estrógenos, los que a su vez estimulan a las células de la granulosa
para que proliferen y aumente el tamaño del folículo.
Unas 24hrs antes de la ovulación, en la adenohipófisis se induce una liberación
masiva de FSH o LH. En respuesta al aumento de LH, los receptores de esta
hormona en las células de la granulosa se inhiben y las células dejan de producir
estrógenos ante la estimulación por LH. Así se reanuda la primera división
meiótica del ovocito primario (pico de LH), causando la formación del ovocito
secundario y del primer cuerpo polar. Luego, tanto las células de la granulosa
como las células de la teca sufren luteinización y producen progesterona.
OVULACIÓN
Es el proceso por el cual se libera un ovocito secundario desde el folículo maduro. El folículo destinado a ovular en cualquier ciclo
menstrual se reclute entre varios folículos primarios en los primeros días del ciclo. Durante la ovulación, el ovocito atraviesa toda la
pared folicular, incluido el epitelio germinativo.
Una combinación de cambios hormonales y efectos enzimáticos produce la
liberación del ovocito secundario en la mitad del ciclo menstrual. Estos factores
comprenden:
o Aumento del volumen y de la presión del líquido folicular.
o Proteólisis enzimática de la pared folicular por plasminógeno activado.
o Depósito de GAG dirigido por hormonas entre el complejo ovocito-
cumulo ovóforo y el estrato granuloso.
o Contracción de las fibras musculares lisas en la capa de la teca externa
desencadenada por prostaglandinas.
Justo antes de la ovulación, el flujo sanguíneo cesa en una región de la superficie
ovárica sobre el folículo que sobresale. Esta región del epitelio germinativo,
conocida como mácula pelúcida (o estigma folicular) se eleva y luego se rompe. El
ovocito, rodeado por la corona radiante y las células del cúmulo ovóforo, se
expulsa del folículo abierto. En el momento de la ovulación, las franjas de la
trompa uterina entran en contacto con la superficie ovárica y dirigen
suavemente la masa cumular que contiene al ovocito hacia el interior de la
trompa uterina. La masa cumular se adhiere con firmeza a las franjas y es
transportada por las células ciliadas que revisten a la trompa. Después de
la ovulación, el ovocito secundario permanece viable durante unas 24hrs.
Si la fecundación no ocurre durante ese tiempo, el ovocito secundario se
degenera mientras atraviesa la trompa uterina.
Normalmente, un solo folículo completa la maduración en cada ciclo y se
rompe para liberar su ovocito secundario. En raras ocasiones, los ovocitos
son expulsados de otros folículos que alcanzaron la madurez durante el
mismo ciclo, lo que conlleva la posibilidad de que se formen cigotos
múltiples.
Los ovocitos primarios dentro de los folículos primordiales inician la
primera división meiótica en el embrión, pero el proceso se detiene en la
etapa de diploteno de la profase meiótica. La primera profase meiótica no
se completa hasta justo antes de la ovulación. Por lo tanto, los ovocitos
primarios permanecen detenidos en la primera profase meiótica entre 12 y
50 años. Este largo periodo de detención meiótica expone al ovocito
primario a influencias ambientales adversas y puede contribuir a errores en
la división, como las no disyunciones.
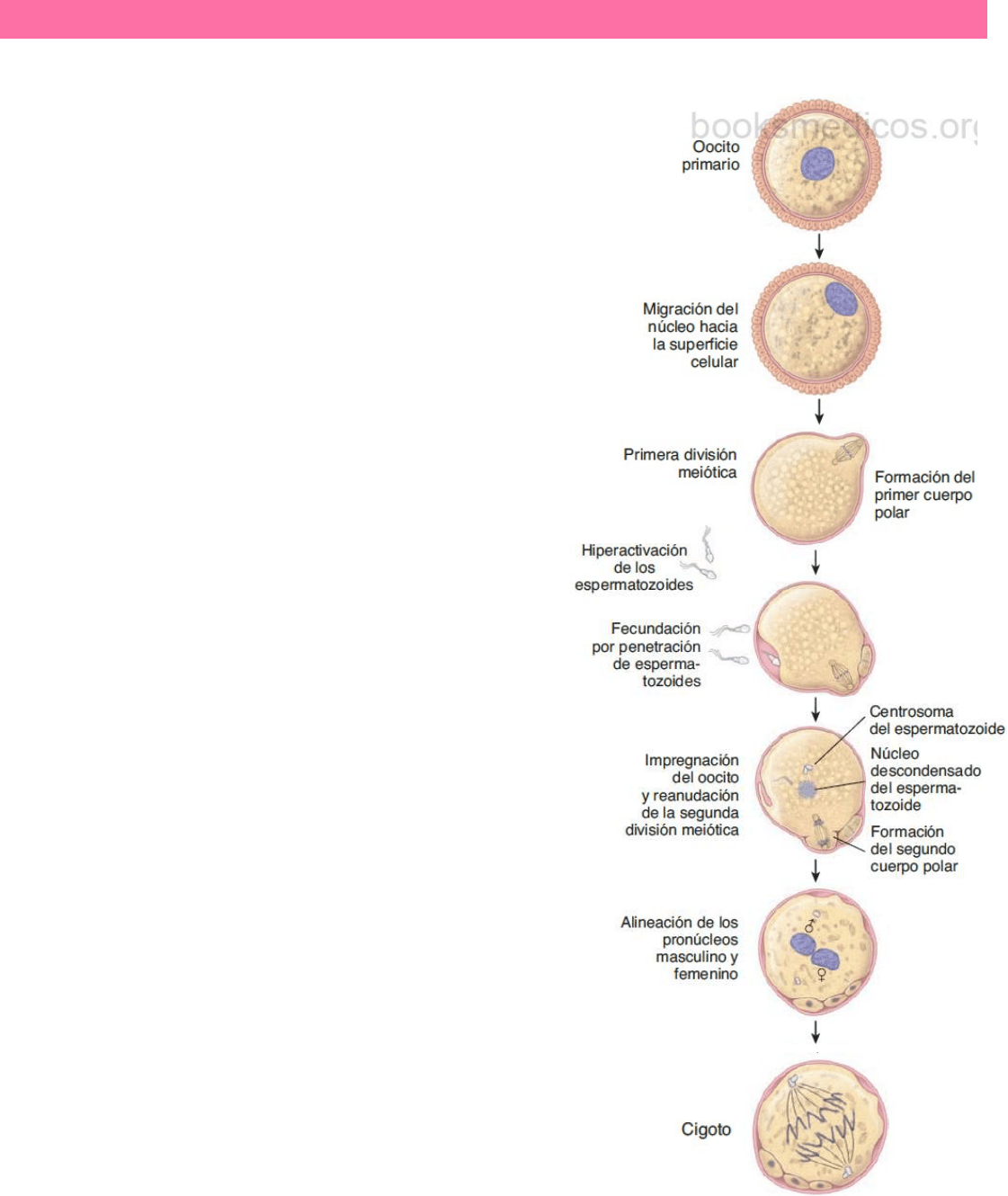
Una vez que se completa la primera división meiótica en el folículo madura, cada célula
hija del ovocito primario recibe una cantidad equivalente de cromatina, pero una de
ellas recibe la mayor parte del citoplasma y se convierte en el ovocito secundario. La
otra célula hija recibe una cantidad mínima de citoplasma y se convierte en el primer
cuerpo polar.
Tan pronto como se completa la primera división meiótica, el ovocito secundario inicia la segunda. A medida que el ovocito secundario
rodeado por las células de la corona radiante abandona el folículo en la ovulación, la segunda división está en progreso. Esta división
queda detenida en metafase y se completa sólo si el ovocito es penetrado por un espermatozoide. Si se produce la fecundación, el
ovocito secundario completa la segunda división meiótica y forma un óvulo maduro con el pronúcleo materno que contiene un juego
de 23 cromosomas. La otra célula producida en esta división es el segundo corpúsculo polar. El primer cuerpo polar es diploide y el
segundo es haploide. En algunos mamíferos, el primer cuerpo polar puede dividirse, por lo que el resultado final de la meiosis está
compuesto por un ovocito y 3 cuerpos polares haploide, que sufrirán apoptosis.
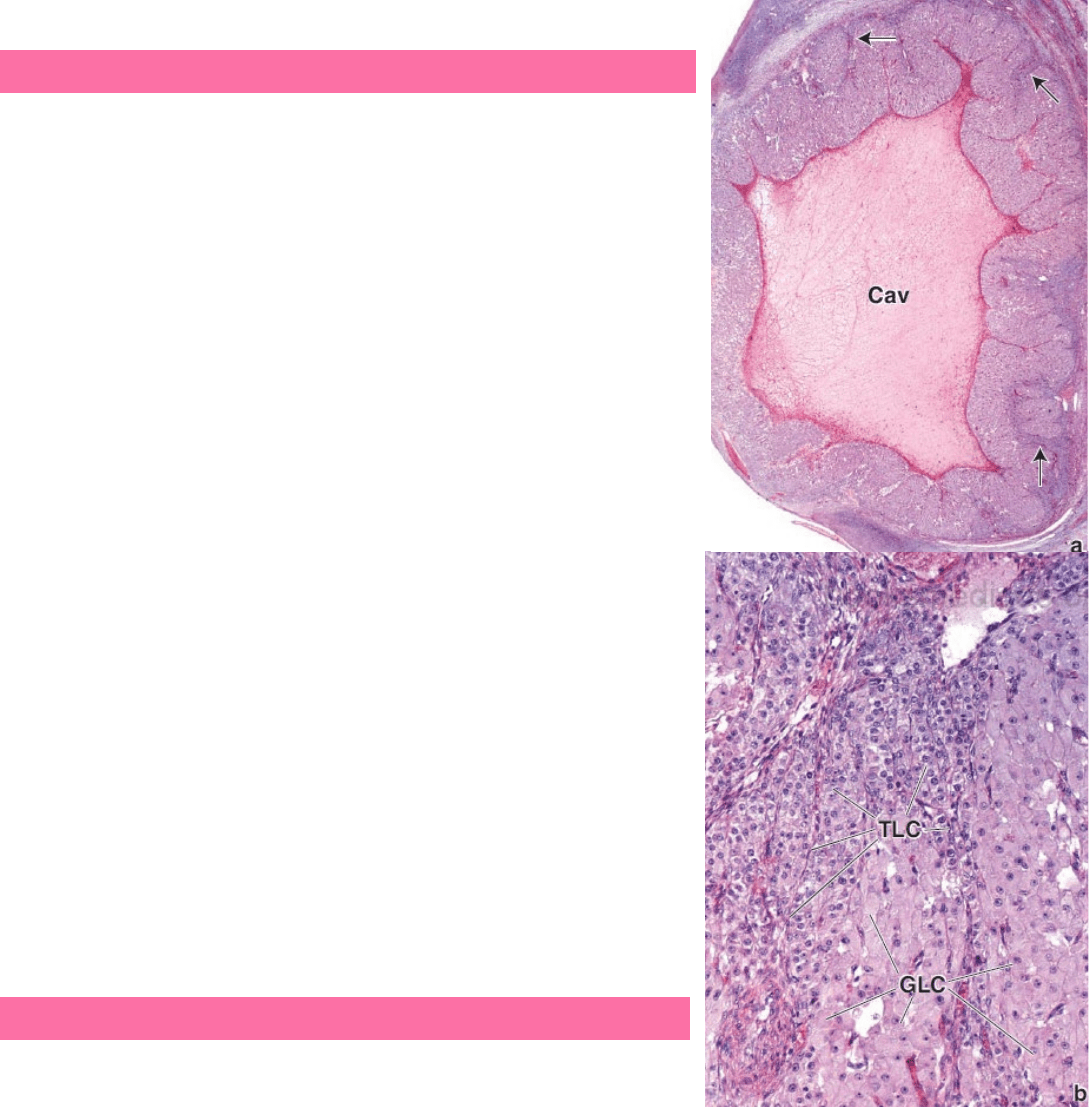
CUERPO LÚTEO
En la ovulación, la pared folicular, compuesta por las células de la granulosa y de la
teca remanentes, adquiere pliegues profundos a medida que el folículo colapsa y se
transforma en el cuerpo lúteo (cuerpo amarillo o glándula lútea). En un principio, la
hemorragia de los capilares en la teca interna hacia la luz folicular lleva a la formación
del cuerpo hemorrágico con un coágulo central. El tejido conjuntivo desde el estroma
invade entonces la antigua cavidad folicular. Las células de la capa granulosa y de la
teca interna se diferencian en células lúteas granulosas y células luteínicas de la teca
en el proceso denominado luteinización. El lipocromo en el citoplasma de las células
les da un aspecto amarillento.
o Células luteínicas de la granulosa: grandes, ubicadas en forma central derivadas
de las células de la granulosa. Sintetizan estrógenos, progesterona e inhibina.
Ésta regula la producción y secreción de la FSH desde la adenohipófisis.
Representan un 80% de las células del cuerpo lúteo.
o Células luteínicas de la teca: pequeñas, localizadas de forma periférica (en los
pliegues) y derivan de las células de la teca interna. Secretan andrógenos y
progesterona. Representan un 20% de las células del cuerpo lúteo.
A medida que el cuerpo lúteo comienza a formarse, los vasos sanguíneos y linfáticos
crecen rápidamente en la capa de la granulosa. Esta estructura muy vascularizada se
localiza en la corteza del ovario y secreta progesterona y estrógenos. Estas
hormonas estimulan el crecimiento y la actividad secretora del revestimiento del
útero, el endometrio, para prepararlo para la implantación del cigoto en desarrollo
en caso de que se produzca la fecundación.
Si la fecundación y la implantación no ocurren, el cuerpo lúteo permanece activo
durante 14 días, donde pasa a llamarse cuerpo lúteo de la menstruación. En
ausencia de hCG y otras luteotrofinas, el ritmo de secreción de progestágenos y
estrógenos declina y el cuerpo lúteo comienza a degenerarse alrededor de 10-12
días post-ovulación. Una cicatriz blanquecina, el cuerpo albicans, se forma a medida
que el material hialino intercelular se acumula entre las células del antiguo cuerpo
lúteo en degeneración.
CAPACITACIÓN Y FECUNDACIÓN
Tras su maduración en el epidídimo, los espermatozoides deben activarse dentro del
sistema genital femenino. Durante este proceso, denominado capacitación, en el
espermatozoide ocurren cambios estructurales y funcionales que aumentan su afinidad de unión a los receptores de la zona pelúcida.
Una capacitación exitosa se confirma por la hiperactivación de los espermatozoides, que se manifiesta por el batido vigoroso de sus
flagelos.
La capacitación comprende varias modificaciones bioquímicas en el espermatozoide y su MP:
o Aumento de la actividad de la adenilato ciclasa que conduce a una concentración elevada de AMPc.

o Aumento del ritmo de fosforilación de la tirosina.
o Activación de los conductos de Ca2+ que causa aumento de la concentración intracelular de este catión.
o Liberación de glucoconjugados de líquido seminal de la superficie de la cabeza del espermatozoide, que inhibían la unión a
los receptores de la membrana pelúcida.
o Modificación de la MP por la extracción del colesterol, el principal inhibidor de la capacitación y la posterior redistribución de
fosfolípidos y las moléculas de hidratos de carbono.
Sólo unos pocos centenares de los millones de espermatozoides que hay en el semen eyaculado alcanzan el sitio de fecundación, que
normalmente es la ampolla de la trompa uterina. A su llegada, los espermatozoides se encuentran con el ovocito secundario rodeado
por la corona radiante. Los espermatozoides deben penetrar la corona radiante para tener acceso a la zona pelúcida. Si bien varios
de pueden penetrarla, sólo un espermatozoide completa el proceso de la fecundación. La capacitación se completa cuando los
espermatozoides son capaces de unirse a los receptores de la zona pelúcida. La unión a los receptores ZP3 desencadena la reacción
acrosómica en la cual las enzimas liberadas del acrosoma permiten que un solo espermatozoide penetre en la ZP. La penetración se
logra por una proteólisis limitada de la zona pelúcida por delante del espermatozoide hipermóvil que avanza. Tras penetrar la
membrana pelúcida, el espermatozoide entra en el espacio perivitelino que hay entre ella y la MP del ovocito (ovolema). Aquí, la MP
del espermatozoide se fusiona con el ovolema, y el núcleo de la cabeza del espermatozoide se introduce en el ovocito. De este modo
se forma el pronúcleo masculino que contiene los 23 cromosomas paternos. Tras la alineación y disolución de las membranas
nucleares de los dos pronúcleos, el cigoto resultante, con su complemento diploide de 46 cromosomas, sufre una división mitótica o
primera segmentación, marcando el inicio del desarrollo embrionario.
CUERPO LÚTEO DEL EMBARAZO
Si la fecundación y la implantación ocurren, el cuerpo lúteo aumenta su tamaño para formar el cuerpo lúteo del embarazo. La
existencia y función del cuerpo lúteo dependen de una combinación de secreciones paracrinas y endocrinas, que en conjunto se
describen como luteotrofinas.
Las luteotrofinas paracrinas son producidas localmente por el ovario. Éstas incluyen:
o Estrógenos.
o IGF-I e IGF-II.
Las luteotrofinas endocrinas son producidas a distancia de su órgano diana (cuerpo lúteo). Éstas comprenden:
o hCG, secretada por el trofoblasto del corion, que estimula los receptores de LH en el cuerpo lúteo e impide su degeneración.
o LH y prolactina, secretadas por la hipófisis.
o Insulina, producida por el páncreas.
Las concentraciones elevadas de progesterona, producida a partir del colesterol por el cuerpo lúteo, bloquean el desarrollo cíclico de
los folículos ováricos.
La función del cuerpo lúteo comienza a declinar tras las 8 semanas de gestación, aunque persiste en todo el embarazo. La placenta
produce una cantidad suficiente de estrógenos y progestágenos de los precursores maternos y fetales como para hacerse cargo de la
función del cuerpo lúteo después de 6 semanas de gestación.
REGULACIÓN HORMONAL DEL CICLO OVÁRICO
Durante cada ciclo menstrual, el ovario sufre cambios cíclicos que comprenden dos fases:
o Fase folicular.
o Fase luteínica.
La ovulación ocurre entre las dos fases.
La fase folicular se inicia con el desarrollo de una pequeña cantidad de folículos primarios (entre 10 a 20) por la acción de la FSH y LH.
La selección de los folículos dominantes ocurre entre los días 5 y 7 del ciclo. Durante los primeros 8 a 10 días del ciclo, la FSH es la
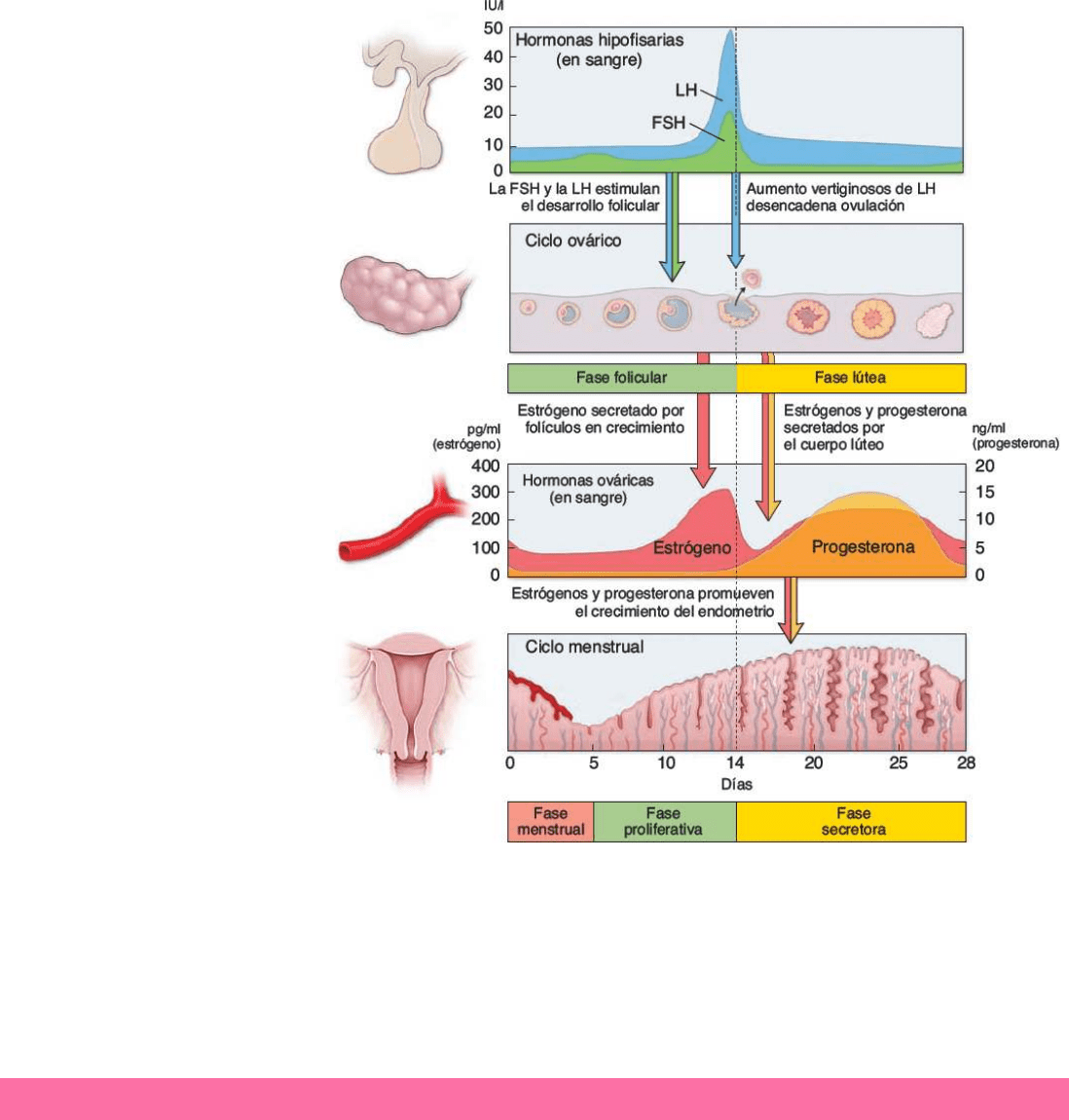
hormona principal que influye sobre el
crecimiento folicular. Estimula a las
células de la granulosa y de la teca, las
que comienzan a secretar hormonas
esteroideas, sobre todo estrógenos,
hacia la luz folicular. A medida que la
producción de estrógenos en el folículo
dominante se incrementa, la secreción
adenohipofisaria de FSH es inhibida por
feedback negativo. Los estrógenos
continúan acumulándose en la luz
folicular y finalmente alcanzan una
concentración que independiza el
folículo de la FSH para su crecimiento y
desarrollo continuos. Al final de esta
fase folicular, antes de la ovulación, la
concentración de progesterona
comienza a aumentar por efecto de la
LH. La cantidad de estrógenos en la
sangre circulante inhibe la producción
adicional de FSH por la adenohipófisis.
La ovulación es inducida por un gran
aumento de la LH, que ocurre en forma
paralela a un incremento menor de la
FSH. La expulsión del ovocito se
produce unas 34hrs a 36hrs después
del inicio del aumento de la LH o
alrededor de 10 a 12hrs después del
incremento máximo de esta hormona.
La fase luteínica se inicia justo después
de la ovulación, mientras las células de
la granulosa y de la teca del folículo roto sufren una rápida transformación morfológica para formar el cuerpo lúteo. El cuerpo lúteo
secreta estrógenos y grandes cantidades de progesterona. Por acción de ambas hormonas, pero por sobre todo de la progesterona,
el endometrio inicia su fase secretora, indispensable para que el útero esté preparado para la implantación. La LH tiene a su cargo el
desarrollo y mantenimiento del cuerpo lúteo durante el ciclo menstrual. Si la fecundación no ocurre, el cuerpo lúteo se degenera es
unos pocos días a medida que disminuyen las concentraciones hormonales. Si la fecundación ocurre, el cuerpo lúteo se mantiene y
continúa secretando progesterona y estrógenos. La hCG, inicialmente sintetizada por el embrión y luego por la placenta, estimula el
cuerpo lúteo y es responsable de su mantenimiento durante el embarazo.
ATRESIA
Muy pocos folículos ováricos que inician su diferenciación en el ovario embrionario podrán completar su maduración. La mayoría de
los folículos se degenera y desaparece a través de la atresia folicular ovárica. Ésta es mediada por la apoptosis de las células de la
granulosa. Una gran cantidad de folículos sufre atresia durante el desarrollo fetal, las primeras etapas de vida posnatal y la pubertad.
Tras la pubertad, grupos de folículos comienzan a madurar durante cada ciclo menstrual y, normalmente, un solo folículo completa su
maduración.
En la atresia de los folículos primordiales y de los folículos en crecimiento pequeños, el ovocito inmaduro reduce su tamaño y se
degenera. En las células de la granulosa ocurren cambios similares. Los folículos atrésicos se retraen y por último desaparecen del
estroma del ovario a causa de la repetición de episodios de apoptosis y fagocitosis de las células de la granulosa. A medida que las

células se reabsorben y desaparecen, las células circundantes del estroma migran al espacio que antes ocupaba el folículo, con lo que
se elimina todo rastro de su existencia.
En la atresia de grandes folículos en crecimiento, la degeneración del ovocito inmaduro se retrasa y ocurre en forma secundaria a las
alteraciones degenerativas de la pared folicular. Este retraso indica que una vez que el ovocito ha logrado su madurez y competencia,
ya no es sensible a los mismos estímulos que inician la atresia de las células de la granulosa. Los cambios foliculares comprenden:
1. Iniciación de la apoptosis dentro de las células de la granulosa, lo cual está indicado por el cese de las mitosis y la expresión
de las endonucleasas y otras enzimas hidrolíticas dentro de estas células.
2. Invasión de la capa granulosa por neutrófilos y macrófagos.
3. Invasión de la capa granulosa por franjas de tejido conjuntivo vascularizado.
4. Exfoliación de las células de las células de la granulosa dentro del antro folicular.
5. Hipertrofia de las células de la teca interna.
6. Colapso del folículo conforme la degeneración continúa.
7. Invasión de la cavidad del folículo por tejido conjuntivo.
Varios productos genéticos regulan el proceso de atresia folicular. Uno es la proteína nerviosa inhibidora de la apoptosis (NAIP)
inducida por gonadotrofinas, que inhibe y retrasa los cambios apoptósicos en las células de la granulosa. La expresión del gen de la
NAIP está presente en todas las etapas del folículo en crecimiento, pero falta en los folículos que sufren atresia. Una concentración
elevada de gonadotrofinas inhibe la apoptosis en los folículos ováricos porque aumenta la expresión de NAIP en los ovarios.
IRRIGACIÓN SANGUÍNEA Y DRENAJE LINFÁTICO
La irrigación sanguínea de los ovarios proviene de dos fuentes diferentes: las arterias ováricas y las arterias uterinas.
Las arterias ováricas son las ramificaciones de la aorta abdominal que pasan a los ovarios a través de los ligamentos suspensorios y
proporcionan la irrigación arterial principal a los ovarios y a las tubas uterinas. Estas se anastomosan con las ramificaciones ováricas
de las arterias uterinas, que se originan en las arterias ilíacas internas. Vasos bastante grandes que surgen de esta región de
anastomosis atraviesan el mesoovario y se introducen en el hilio ovárico; arterias denominadas arterias helicinas, debido a que se
ramifican y se enrollan a medida que pasan a la médula ovárica.
Las venas que acompañan a las arterias forman un plexo denominado plexo pampiniforme al abandonar el órgano por el hilio. La vena
ovárica se forma a partir de ese plexo.
En la región cortical del ovario, las redes de vasos linfáticos en las capas de la teca rodean a los folículos en desarrollo grandes y los
folículos atrésicos, así como los cuerpos lúteos. Los vasos linfáticos siguen el trayecto de las arterias ováricas conforme ascienden
hacia los ganglios linfáticos paraaórticos en la región lumbar.
INERVACIÓN
Las fibras nerviosas autónomas que inervan al ovario forman parte sobre todo del plexo ovárico autónomo. Las fibras nerviosas siguen
a las arterias en su paso por la médula y la corteza del ovario e inervan al músculo liso de las paredes vasculares. Las terminaciones
nerviosas sensitivas están dispersas en el estroma. Las fibras sensitivas envían impulsos a través del plexo ovárico que alcanzan los
ganglios espinales en los primeros nervios lumbares. Por lo tanto, el dolor ovárico es referido en la distribución cutánea de estos
nervios espinales. El dolor ovárico es producido por contracciones en las células musculares lisas tanto en el ovario como en sus
ligamentos. Estas contracciones responden a un aumento de la concentración de prostaglandinas mediado por un incremento masivo
de LH.
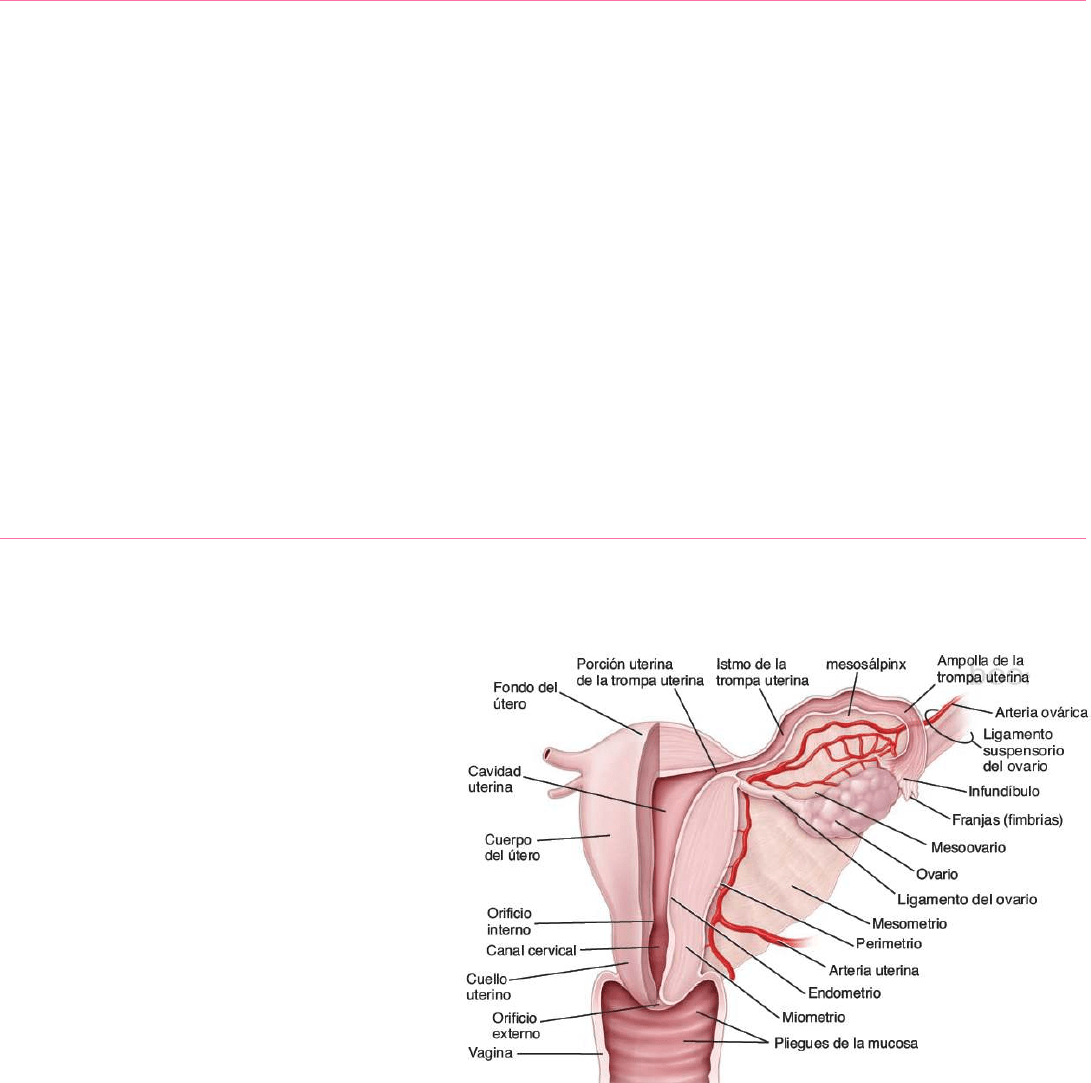
TROMPAS UTERINAS
Órganos pares con forma de tubo que se extienden en forma bilateral desde el útero hacia los ovarios. Estos órganos tubulares
transportan el óvulo desde el ovario hacia el útero y proveen el ambiente necesario para la fecundación y el desarrollo inicial del cigoto
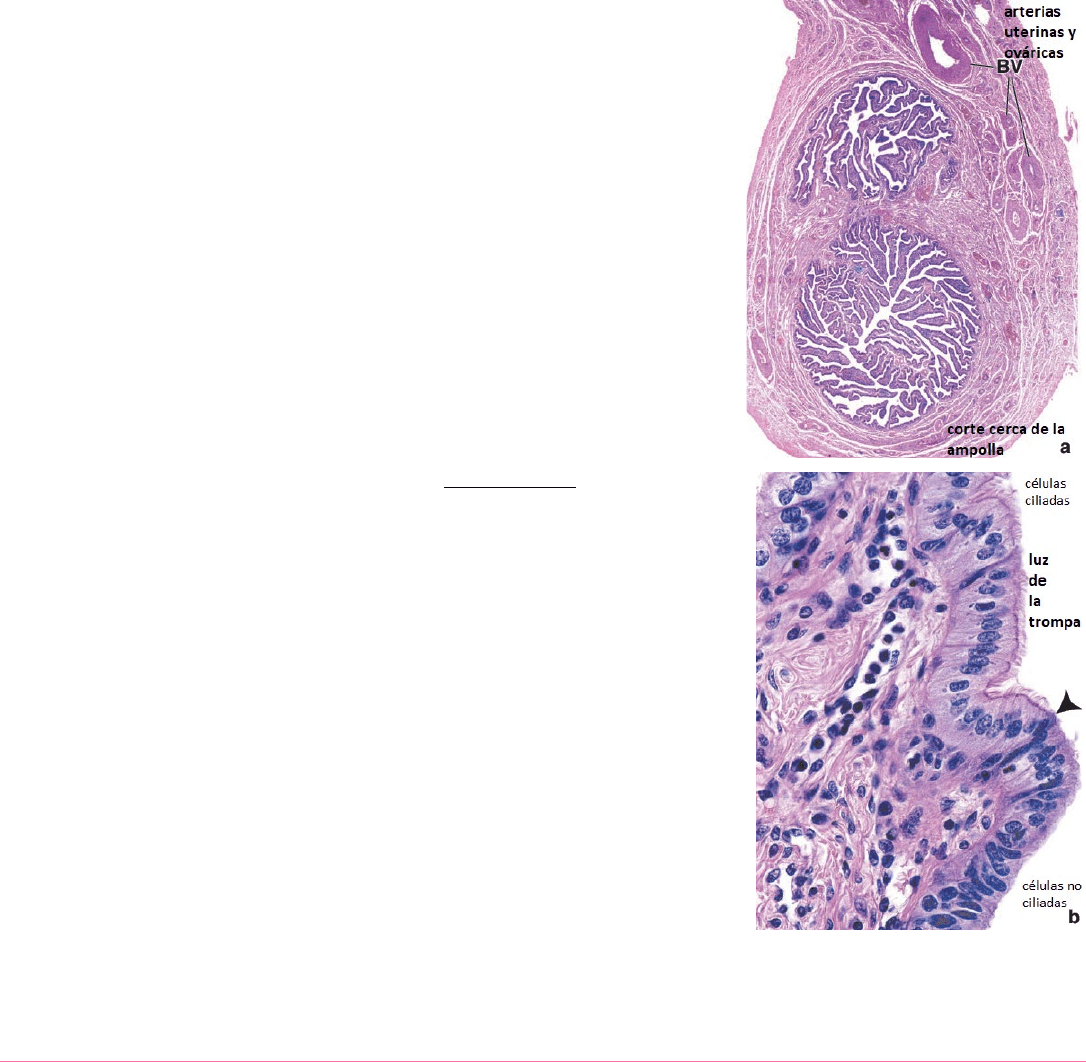
hasta su etapa de mórula. Uno de los extremos de la trompa contiguo al ovario se abre hacia la cavidad peritoneal, el otro se comunica
con la cavidad uterina.
Cada trompa uterina mide entre 10-12cm y puede dividirse en 3 segmentos macroscópicos:
o Infundíbulo: segmento de la trompa en forma de embudo contiguo al ovario. Extensiones de flecos, o fimbrias, se extienden
desde la boca del infundíbulo hacia el ovario.
o Ampolla: segmento más largo de la trompa y constituye alrededor de 2/3 partes
de su longitud total. Es el sitio donde ocurre la fecundación.
o Istmo: segmento medio, estrecho de la trompa uterina, contiguo al útero.
o Porción uterina: mide aprox. 1cm de longitud, se ubica dentro de la pared uterina
y se abre hacia la cavidad del útero.
La pared de la trompa se parece a la pared de otras vísceras huecas, y está compuesta por
una capa serosa externa, una capa muscular intermedia y una capa mucosa interna. Carece
de submucosa.
o Serosa: o peritoneo, es la capa más externa de la trompa y está compuesta por
mesotelio y una capa delgada de tejido conjuntivo.
o Muscular: organizada en una capa circular interna gruesa y una capa externa más
delgada.
o Mucosa: revestimiento interno de la trompa, exhibe pliegues longitudinales
delgados que se proyectan hacia la luz de la trompa en toda su longitud.
El revestimiento epitelial de la mucosa es un epitelio cilíndrico simple compuesto por dos
tipos de células, ciliadas y no ciliadas, que representan diferentes estados funcionales de
un único tipo celular:
o Las células ciliadas son más numerosas en el infundíbulo y en la ampolla. El batir
de los cilios está dirigido hacia el útero.
o Las células no ciliadas son células secretoras que producen el líquido que provee
sustancias nutritivas al óvulo.
Las células epiteliales sufren una hipertrofia cíclica durante la fase folicular y atrofia durante
la fase lútea en respuesta a los cambios de las concentraciones hormonales. El estrógeno
estimula la ciliogénesis y la progesterona incrementa la cantidad de células secretoras. En
el momento de la ovulación, el epitelio alcanza una altura de 30micrometros que después
se reduce casi a la mitad, justo antes de iniciarse la menstruación.
La trompa uterina ejecuta movimientos justo antes de la ovulación conforme las fimbrias
entran en contacto con el ovario y se ubican sobre la superficie ovárica donde ocurrirá la
rotura. Cuando se expulsa el ovocito, las células ciliadas del infundíbulo lo barren hacia la
abertura de la trompa uterina y así impiden que caiga en la cavidad peritoneal. El ovocito se desplaza a lo largo de la trompa mediante
contracciones peristálticas. La fecundación suele ocurrir en la ampolla, cerca del límite con el istmo. El cigoto permanece en la trompa
durante unos 3 días antes de ingresar en la cavidad uterina.
ÚTERO
Recibe la mórula en proliferación rápida que proviene de la trompa. Todo el desarrollo embrionario y fetal posterior ocurre dentro del
útero, el cual sufre un incremento drástico en tamaño y desarrollo.
Desde el punto de vista anatómico, se divide en dos regiones:

o Cuerpo: porción superior grande del útero. La parte superior redondeada del cuerpo que se expande por arriba de la
desembocadura de las tubas uterinas recibe el nombre de fondo uterino.
o Cuello o cérvix: es la porción inferior del útero, que se encuentra separada del cuerpo por el istmo. La luz del cérvix, el
conducto cervical, presenta una abertura estrecha en cada extremo. El orificio interno comunica con la cavidad del cuerpo
uterino y el orificio externo con la luz vaginal.
La pared uterina está compuesta por 3 capas, dese la luz hacia afuera son:
o Endometrio: mucosa del útero.
o Miometrio: capa muscular gruesa. Está en continuidad con la capa
muscular de la trompa uterina y la vagina. Las fibras de músculo liso
también se extienden dentro de los ligamentos fijados al útero.
o Perimetrio: capa serosa o cubierta peritoneal visceral del útero. Se
continúa con el peritoneo pélvico y abdominal y consiste en un
mesotelio y una capa delgada de tejido conjuntivo laxo. Bajo el
mesotelio, suele ser prominente una capa de tejido elástico. El
perimetrio cubre toda la superficie posterior del útero, pero sólo una
parte de la superficie anterior.
Tanto el miometrio como el endometrio sufren cambios cíclicos mensuales
cuya finalidad es preparar el útero para la implantación de un embrión. Estos
cambios constituyen el ciclo menstrual. Si un embrión se implanta, el ciclo se
detiene y ambas capas sufren un crecimiento y diferenciación considerables
durante el embarazo.
MIOMETRIO
Es la capa más gruesa de la pared uterina. Compuesto por 3 capas de músculo
liso:
o La capa media contiene abundantes vasos sanguíneos grandes y vasos
linfáticos, y se denomina estrato vascular. Posee haces de músculo
liso entrelazados con orientación circular o en espiral.
o Los haces musculares lisos en las capas externa e interna están
orientados predominantemente paralelos al eje longitudinal del
útero.
Comparado con el útero, el cérvix posee más tejido conjuntivo y menos músculo liso. Las fibras elásticas son abundantes en el cérvix,
pero en el cuerpo del útero sólo se encuentran en cantidades apreciables en la capa externa del miometrio.
ENDOMETRIO
Los cambios en la actividad secretora del endometrio durante el ciclo están relacionados con la maduración de los folículos ováricos.
El final de cada ciclo se caracteriza por la destrucción y desprendimiento parciales del endometrio, que se acompañan de hemorragia
desde los vasos de la mucosa. La eliminación de sangre y restos de tejido por la vagina, que suele durar entre 3 a 5 días se conoce
como menstruación o ciclo menstrual.
Durante la vida fértil, el endometrio está compuesto por dos capas o zonas que se diferencian en estructura y función:
o Estrato o capa funcional: es la porción gruesa del endometrio, que se desprende durante la menstruación.
o Estrato o capa basal: retenida durante la menstruación y sirve como fuente para la regeneración del estrato funcional.
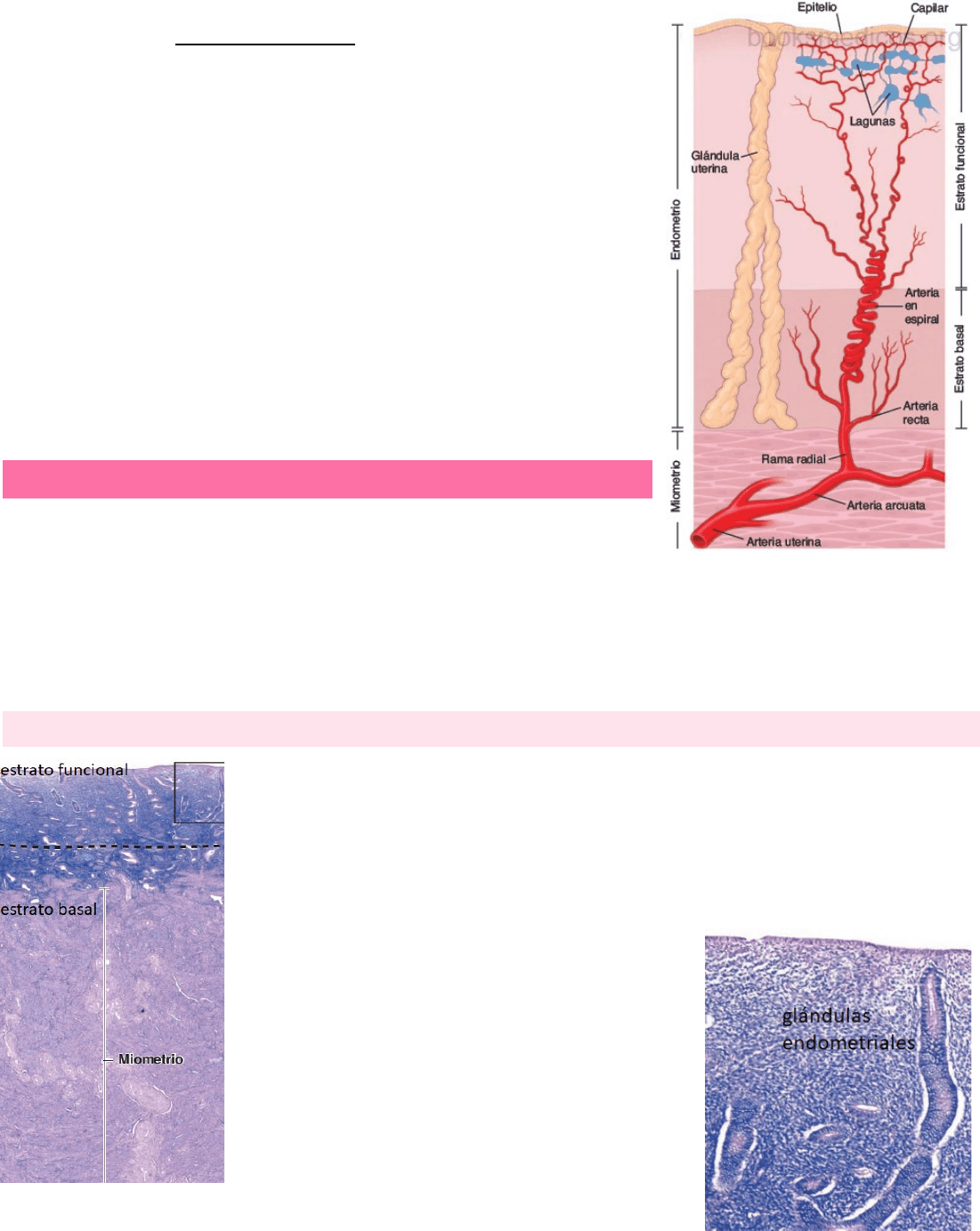
Durante las fases del ciclo menstrual, el espesor del endometrio varía entre 1mm y 6mm.
Está revestido por un epitelio cilíndrico simple con una mezcla de células secretoras y
ciliadas. El epitelio superficial se invagina en la lámina propia subyacente, el estroma
endometrial, para formar las glándulas uterinas. Éstas con glándulas tubulares simples
contienen menos células ciliadas. El estroma endometrial es muy celular y contiene
abundante sustancia fundamental intercelular. Aquí no hay submucosa que separe el
endometrio del miometrio.
El endometrio contiene un sistema singular de vasos sanguíneos. La arteria uterina emite
entre 6 a 10 arterias arcuatas que se anastomosan en el miometrio. Las ramas de estas
arterias, las arterias radiales, ingresan en la capa basal del endometrio donde dan origen
a pequeñas arterias rectas que irrigan esta región. La rama principal de la arteria radial,
la arteria en espiral, continúa su trayecto hacia arriba mientras se enrolla. Las arterias en
espiral emiten muchas arteriolas que con frecuencia se anastomosan y forman un lecho
capilar extenso, que incluye segmentos dilatados de paredes delgadas que se denominan
lagunas. Las lagunas pueden formar parte del sistema venoso que drena el endometrio.
La porción distal de las arterias en espiral, bajo la influencia de los estrógenos y la
progesterona, sufre una degeneración y una regeneración con cada ciclo menstrual.
CAMBIOS CÍCLICOS DURANTE EL CICLO MENSTRUAL
El ciclo normalmente se repite cada 28 días, durante los cuales el endometrio atraviesa
una secuencia de cambios morfológicos y funcionales.
Es conveniente describir el ciclo en 3 fases sucesivas:
o Fase proliferativa: ocurre al mismo tiempo que la maduración folicular y es afectada por la secreción de los estrógenos ováricos.
o Fase secretora: coincide con la actividad funcional del cuerpo lúteo y es afectada principalmente por la secreción de progesterona.
o Fase menstrual: comienza cuando la producción hormonal del ovario declina con la degeneración del cuerpo lúteo.
FASE PROLIFERATIVA
Al final de la fase menstrual, el endometrio está compuesto por una banda delgada de tejido
conjuntivo, que contiene las porciones basales de las glándulas uterinas y los segmentos proximales
de las arterias en espiral. Esta capa es el estrato basal, la capa desprendida durante la menstruación
es el estrato funcional.
La fase proliferativa se inicia por acción de los estrógenos. Las células epiteliales, las células del
estroma y las células endoteliales del estrato basal proliferan con rapidez y se observan los siguientes
cambios:
o Las células epiteliales en la porción basal de las glándulas
reconstituyen las glándulas y migran para cubrir la superficie
endometrial que se desprendió.
o Las células del estroma proliferan y secretan colágeno y
sustancia fundamental.
o Las arterias en espiral se alargan a medida que se
restablece el endometrio.
La fase proliferativa continúa hasta 1 día después de la
ovulación. En el final de esta fase, el endometrio tiene un espesor de unos 3mm, las glándulas
tienen una luz estrecha y son relativamente rectas. En la región basal de las células epiteliales
hay una acumulación de glucógeno.
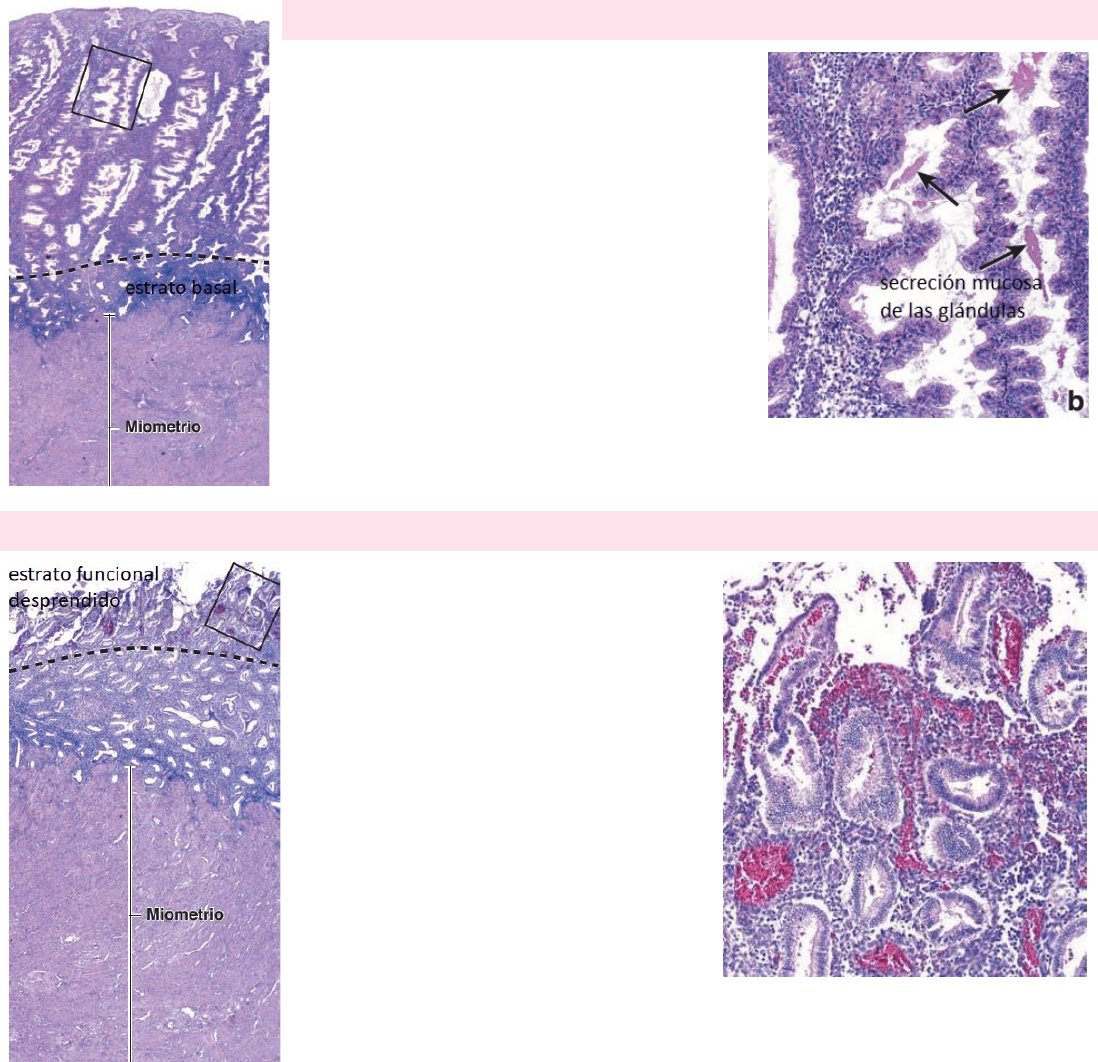
FASE SECRETORA
1 o 2 días después de la ovulación y por efecto de la
progesterona, ocurren cambios en el estrato funcional. El
endometrio se torna edematoso y puede alcanzar un
espesor de entre 5 a 6mm. Las glándulas crecen y la luz se
distiende a medida que se llena de productos de
secreción. El líquido mucoso producido por el epitelio
glandular tiene sustancias nutritivas que son necesarias
para sustentar el desarrollo en el caso de una
implantación. El crecimiento observado en esta etapa es
producto de la hipertrofia de las células epiteliales, el
aumento de la vascularización y el edema del endometrio.
Las arterias en espiral se alargan y enrollan más,
extendiéndose casi hasta la superficie del endometrio.
La acción secuencial de los estrógenos y progesterona sobre las células del estroma permite su
transformación en células deciduales. El estímulo transformador es la implantación del blastocisto.
FASE MENSTRUAL
El cuerpo lúteo produce hormonas en forma activa
durante unos 10 días si no se produce la
fecundación. Al disminuir con rapidez las
concentraciones hormonales ocurren cambios en
la irrigación del estrato funcional del endometrio.
En un principio, las concentraciones periódicas de
las paredes de las arterias en espiral, que duran
varias horas, causan isquemia del estrato
funcional. Las glándulas detienen su secreción y el
endometrio reduce su espesor conforme el
estroma se torna menos edematoso.
Después de unos 2 días, los períodos de
contracción arterial causan la destrucción del
epitelio de revestimiento superficial y la rotura de
los vasos sanguíneos. Cuando las arterias en espiral se ocluyen, la sangre fluye hacia el estrato
basal pero no hacia el estrato funcional. A medida que los fragmentos de tejido se separan del
endometrio, los extremos desgarrados de las venas, las arterias y glándulas quedan expuestos. El
desprendimiento continúa hasta que sólo queda la capa basal. La pérdida promedio de sangre durante la fase menstrual es de 35ml a
500ml. El flujo sanguíneo a través de las arterias rectas mantiene irrigada la capa basal.
Si no hay fecundación, el cese de la hemorragia acompañará el crecimiento y la maduración de nuevos folículos ováricos. Al comenzar
la fase proliferativa del ciclo siguiente, las células epiteliales proliferan con rapidez y migran hacia la superficie para restaurar el epitelio
de revestimiento.
Si no hay ovulación (ciclo anovulatorio), no se forma el cuerpo lúteo y, por ende, no se produce progesterona. En ausencia de
progesterona, el endometrio no ingresa en fase secretora y continúa en la fase proliferativa hasta la menstruación.
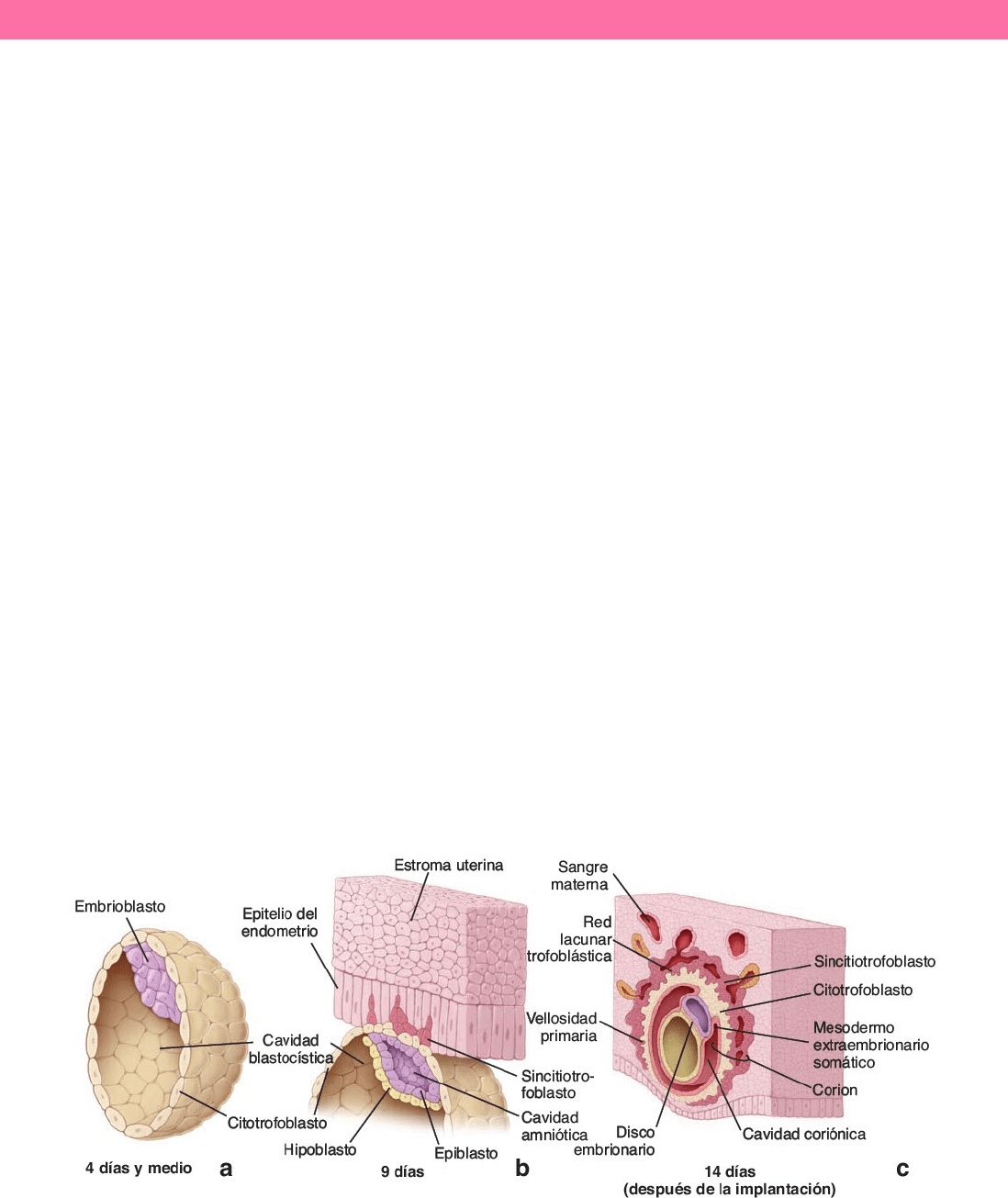
IMPLANTACIÓN
Si hay fecundación y el producto de la concepción se implanta, la involución del endometrio se retrasa hasta después del parto. Cuando
el blastocisto se implanta en la mucosa uterina al principio de la segunda semana, las células del corion de la placenta en desarrollo
comienzan a secretar hCG y otras luteotrofinas. Estas hormonas mantienen el cuerpo lúteo y lo estimulan para que continúe
produciendo progesterona y estrógenos.
El óvulo humano fecundado sufre una serie de cambios mientras atraviesa la trompa uterina y llega a la cavidad del útero que lo
prepara para su implantación en la mucosa uterina. Poco después de la fecundación, el embrión viable secreta el factor de
preimplantación (PIF), un péptido de 15 aminoácidos específico para el embrión que promueve la adhesión del embrión al
endometrio. En el momento de la implantación, el PIF estimula la proliferación y la invasión del trofoblasto en la decidua basal.
El cigoto sufre escisión, seguida de una serie de divisiones mitóticas sin crecimiento celular, cuya consecuencia es el aumento rápido
de la cantidad de células en el embrión. La masa celular resultante es la mórula y las células individuales son los blastómeros. Unos 3
días después de la fecundación, la mórula ingresa en la cavidad uterina. La mórula permanece libre en el útero alrededor de un día
mientras continúan las divisiones celulares y el desarrollo. El embrión inicial da origen a un blastocisto, una esfera celular hueca con
un cúmulo de células localizado en forma central. Este macizo celular interno da origen a los tejidos del embrión propiamente dicho,
mientras que la capa de células periféricas, el macizo celular externo forma el trofoblasto y después la placenta.
El líquido entra a través de la membrana pelúcida durante este proceso y forma una cavidad llena de líquido, la cavidad blastocística.
Este fenómeno define el comienzo de la etapa del blastocisto. Conforme éste permanece libre en la luz uterina durante uno o dos días
y sufre divisiones mitóticas adicionales, la membrana pelúcida desaparece. El macizo celular ahora se llama trofoblasto y el macizo
celular interno se denomina embrioblasto.
La adherencia del blastocisto al epitelio endometrial ocurre durante la ventana de implantación, que es el período durante el cual el
útero es receptivo para la implantación del blastocisto gracias a la acción de la progesterona y estrógeno sobre el endometrio. La
ventana de implantación comienza el día 6 después de la secreción masiva de LH y se completa alrededor del día 10.
Al entrar en contacto con la pared uterina por las células trofoblásticas del polo embrionario, el trofoblasto prolifera con rapidez y
comienza a invadir el endometrio. El trofoblasto invasor se diferencia en un citotrofoblasto y un sincitiotrofoblasto:
o El citotrofoblasto es una capa celular interna mitóticamente activa que produce células que se funden con el
sincitiotrofoblasto. La fusión de ambos puede desencadenarse por apoptosis.
o El sincitiotrofoblasto es una capa celular externa que no es mitóticamente activa y consiste en una masa citoplasmática
multinucleada. Invade activamente el epitelio y el estroma subyacente del endometrio.
Por actividad del trofoblasto, el blastocisto queda totalmente sumergido en el endometrio hacia el 11vo día de desarrollo. El
sincitiotrofoblasto posee aparatos de Golgi desarrollados, REL y RER abundantes, muchas mitocondrias y una cantidad grande de
inclusiones lipídicas. Estas características son consistentes con la función secretora de progesterona, estrógenos hCG y lactógenos que
cumple esta capa.
Este documento contiene más páginas...
Descargar Completo
Histología ap reproductor fem.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.