GLUCÓLISIS
La glucólisis es la vía metabólica encargada de oxidar la glucosa con la finalidad
de obtener energía para la célula.
Es un proceso anaeróbico (no requiere Oxígeno).
Es una ruta metabólica común a casi todas las células, tanto procariotas como
eucariotas. En eucariotas, la glucólisis ocurre en el citosol.
La entrada de Glucosa a las células se produce por medio de la difusión
facilitada mediada por un grupo de receptores/transportadores de membrana
llamados Glut:
● Glut 1: en todos los tejidos
● Glut 2: hígado y páncreas
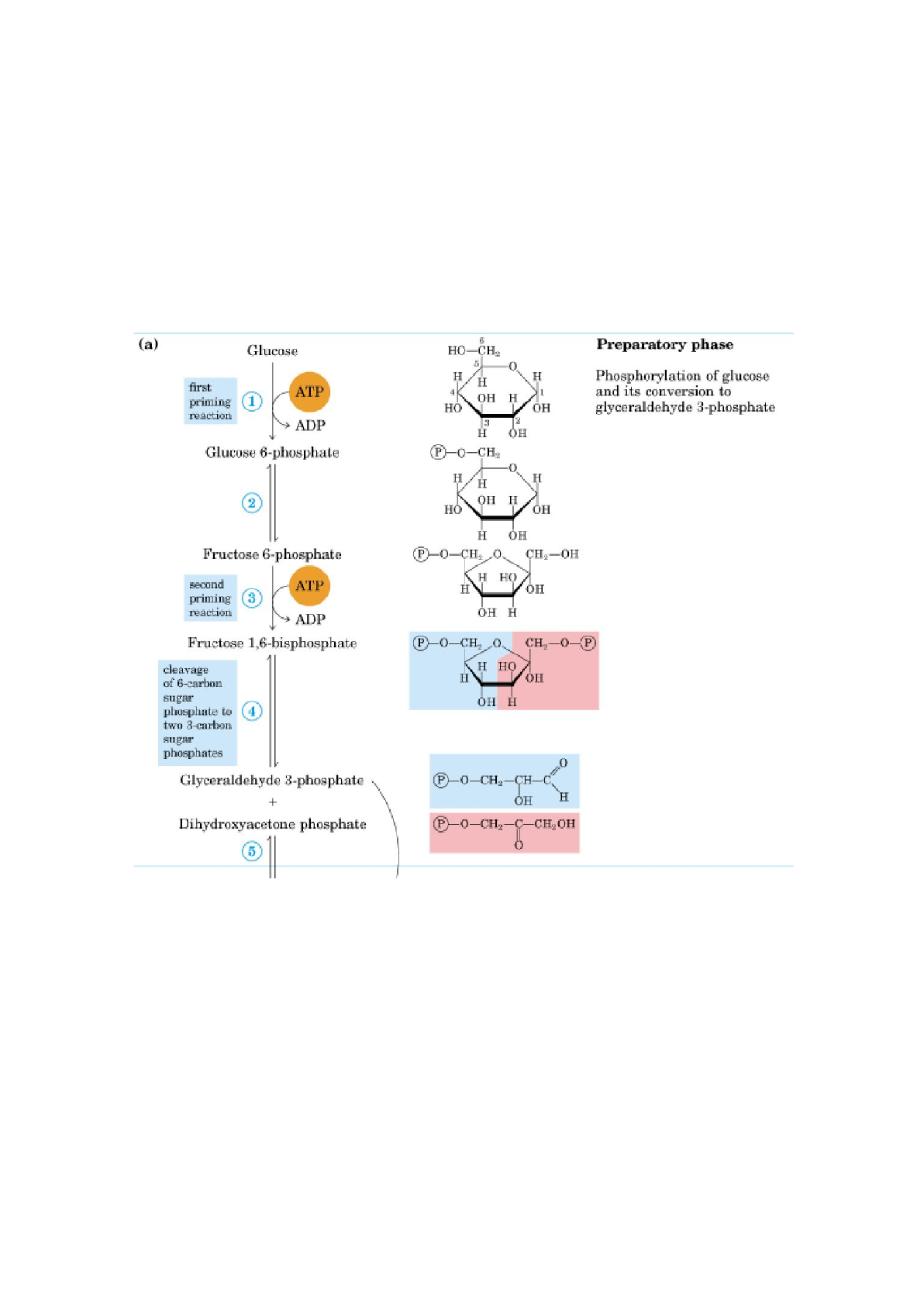
La rotura de la glucosa, que tiene 6 carbonos, en dos moléculas de piruvato,
formado por 3 carbonos, tiene lugar en 10 pasos, de los que los 5 primeros
constituyen la fase preparatoria, en la cual se invierte la energía del ATP,
elevando el contenido de energía libre de los intermediarios, y las cadenas de
carbono de tdas las hexosas metabolizadas se convierten en un producto común,
el gliceraldehído3-fosfato. El retorno energético tiene lugar en la fase de
beneficios de la glucólisis.
Fase preparatoria de la Glucólisis:
En ésta fase se invierten 2 moléculas de ATP y se rompe la cadena de hexosa en
2 triosas fosfato.
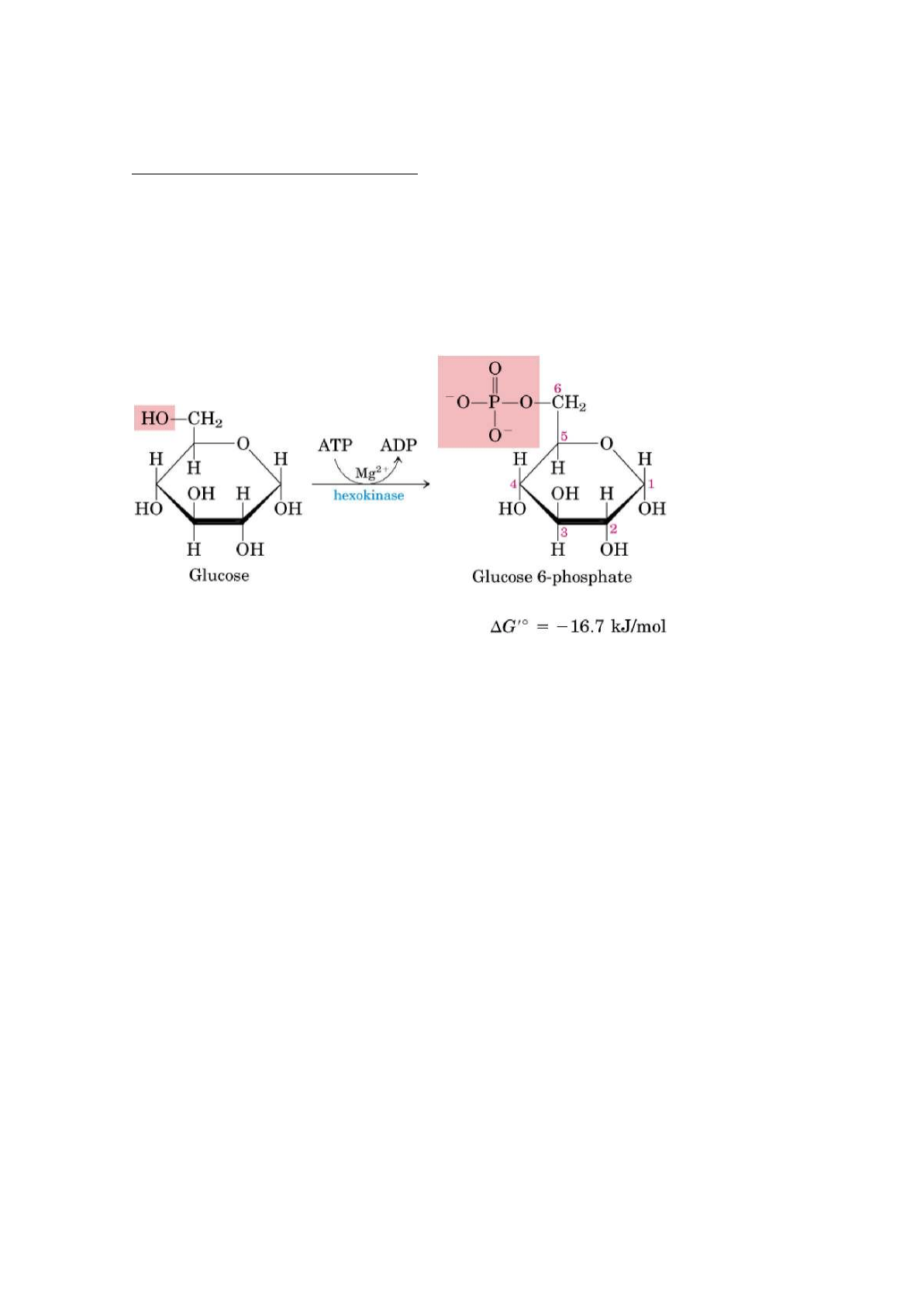
1. Fosforilación de la Glucosa:
En ésta reacción se fosforila la glucosa en el carbono 6 para formar
glucosa-6-fosfato. La fosforilación tiene como finalidad modificar a la molécula
de glucosa para que no pueda salir de la célula.
La fosforilación se hace a expensas de un fosfato qu ese extrae de una molécula
de ATP.
La enzima que actúa es la hexoquinasa (hexo= puede fosforilar a cualquier
hexosa; quinasa= transfiere fosfatos).
La hexoquinasa necesita Mg²⁺ para su actividad, porque el verdadero sustrato
del enzima es el complejo MgATP²ˉ.
Ésta enzima transfiere el fosfato terminal del ATP al carbono 6 de la glucosa,
formando glucosa-6-fosfato y ADP.
Existen 4 variedades de hexoquinasas en el organismo: las hexoquinasas I, II,
III y IV.
Las hexoquinasas I, II y III pueden fosforilar a cualquier hexosa, mientras que la
hexoquinasa IV, presente en el hígado, sólo puede fosforilar a la glucosa y se le
llama glucoquinasa.
Hexoquinasa: amplia especificidad de sustrato, Km=0,1mM.
Glucoquinasa: específica para glucosa, Km=10mM.
La hexoquinasa y la glucoqunasa son isozimas, es decir enzimas diferentes que
catalizan reacciones de fosforilación, pero poseen diferentes velocidades de
reacción, diferentes Km.
Ambas enzimas son quinasas, es decir, fosforilan; mediante este proceso se
aseguran que la glucosa no salga de la célula. Ambas son enzimas ubicadas en el
citosol, utilizan magnesio como cofactor, ambas realizan reacciones
irreversibles.
La glucoquinasa es específica para la D-glucosa, tiene baja afinidad por lo cual
alto Km y se localiza en el hígado y páncreas.
La hexoquinasa fosforila D-glucosa, D-manosa y D- fructosa, tiene alta afinidad
por lo tanto bajo Km y se localiza en todos los tejidos.
La hexoquinasa es inhibida por su producto, la glucoquinasa es inhibida por
fructosa-6-fosfato.
Ambas son enzimas alostéricas.
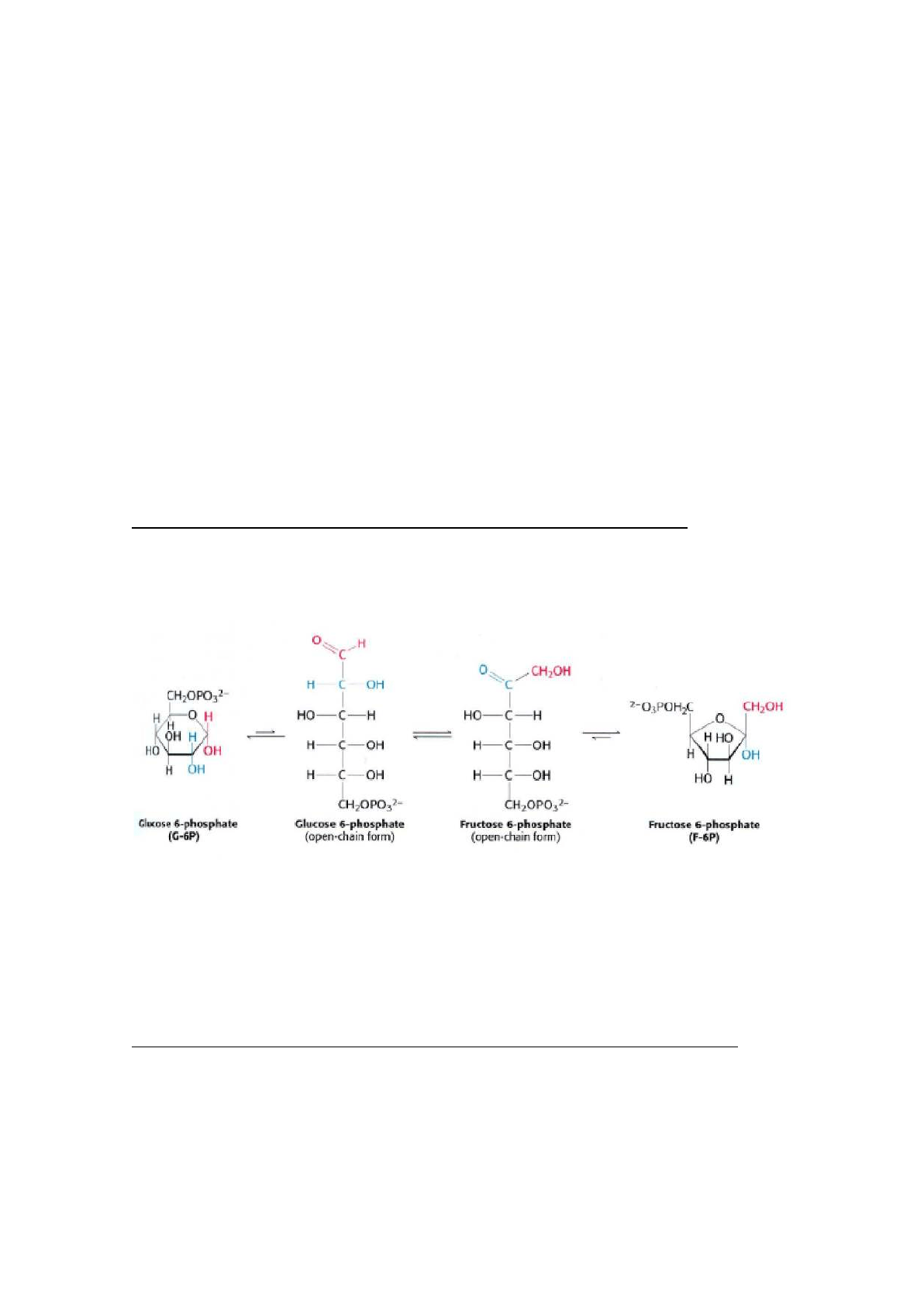
2. Conversión de la glucosa-6-fosfato en fructosa-6-fosfato
∆G= 1,7KJ/mol
El enzima fosfohexosa isomerasa (fosfoglucosa isomerasa) cataliza la
isomerización (conversión de una aldosa en una cetosa) reversible de la
glucosa-6-fosfato. El mecanismo de ésta reacción se realiza a través de un
intermedio enediol.
3. fosforilación de la fructosa-6-fosfato a fructosa 1,6-bisfosfato
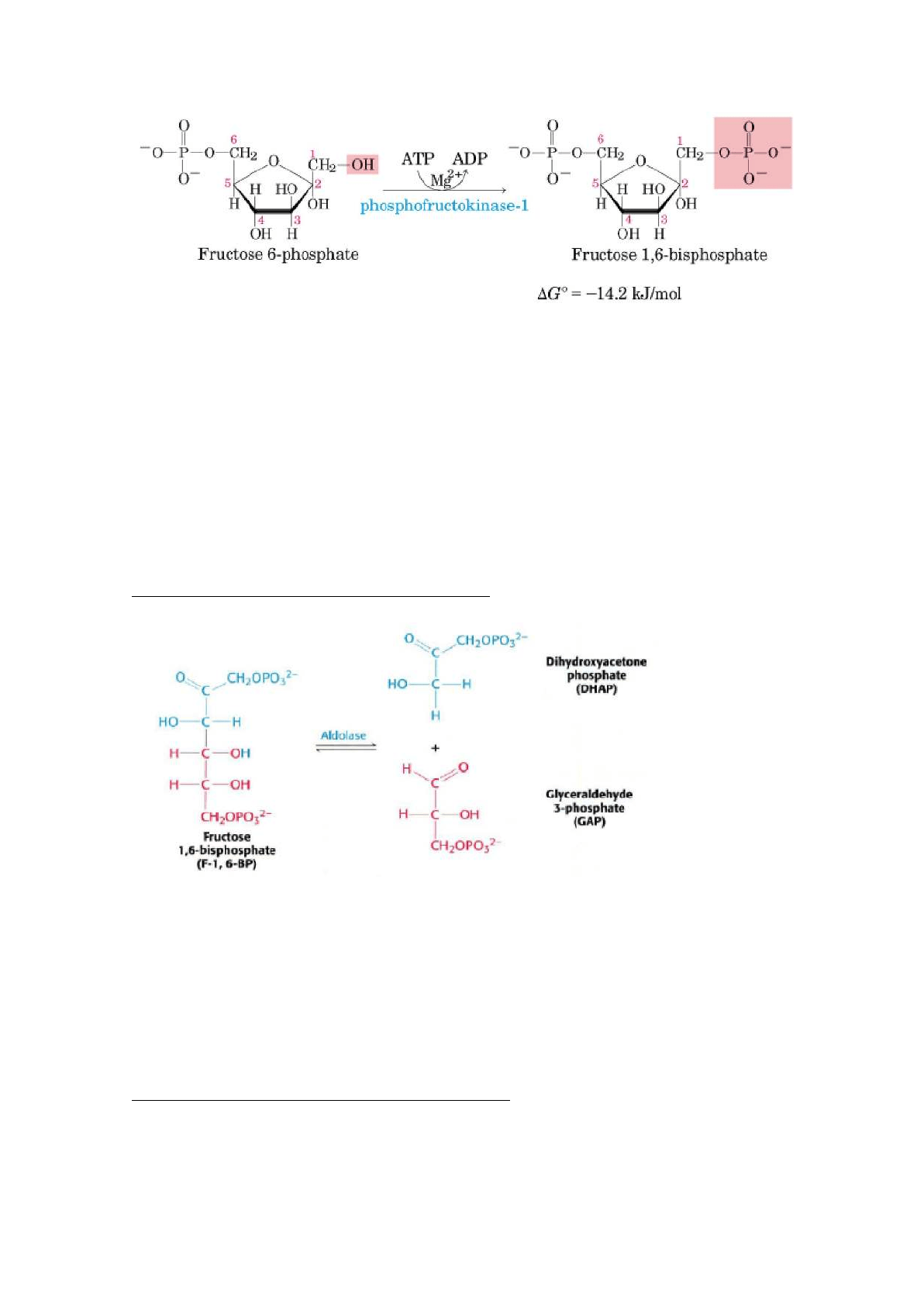
Reacción catalizada por la fosfogructoquinasa 1 (PFK-1), enzima alostérica. Esta
enzima cataliza la transferencia de un grupo fosforilo desde el ATP a la
fructosa-6-fosfato para dar fructosa 1,6-bisfosfato.
Es la principal reacción de regulación de la glucólisis.
Moduladores positivos: ADP, AMP, F-2,6DP
Moduladores negativos: ATP, citrato
Existe otra fosfofructoquinasa, llamada PFK-2 que se encuentra en el hígado y
que forma 2,6 bisfosfofructosa.
El papel de la PFK-2 es formar un regulador que estimula la PFK-1. Cuando el
hígado comienza la glucólisis, una proporción de fructosa-6-fosfato se convierte
en 2,6 BPF para estimular a la PFK-1-
4. Rotura de la fructosa 1,6-bisfosfato
∆G= 23,8 KJ/mol
El enzima aldolasa cataliza una condensación aldólica reversible. La fructosa
1,6-bisfosfato se rompe dando 2 triosas fosfato diferentes, el
gliceraldehído-3-fosfato, una aldosa, y la dihidroxiacetona fosfato, una cetosa.
El gliceraldehído-3-fosfato puede seguir la segunda etapa, mientras que la
dihidroxiacetona fosfato se debe isomerizar a gliceraldehído-3-fosfato.
5. Interconversión de las triosas fosfato
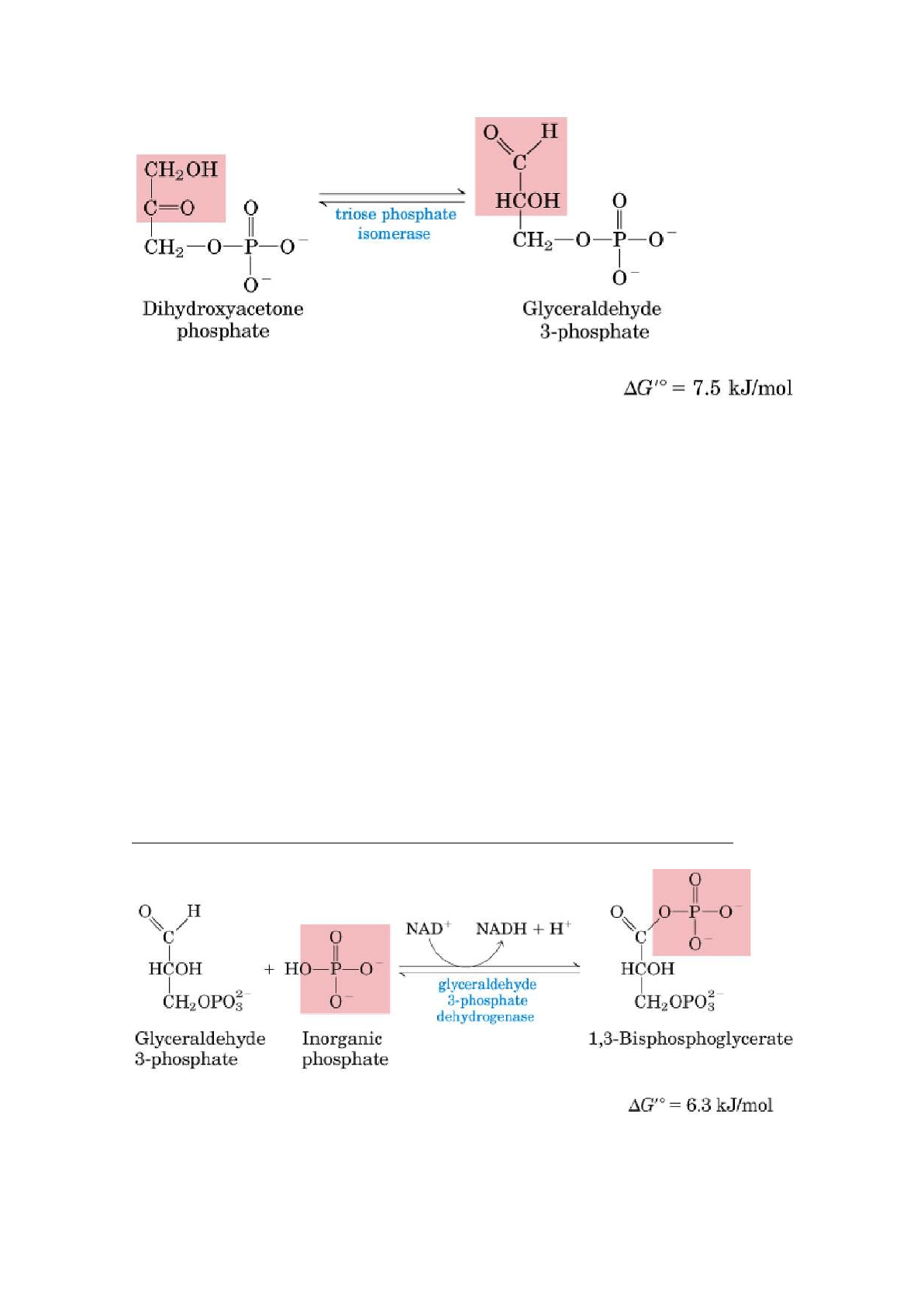
La dihidroxiacetona-fosfato se isomeriza a gliceraldehído-3-fosfato para poder
continuar la segunda etapa- la enzima es la triosa fosfato isomerasa y es una
reacción reversible. Esta reacción completa la fase preparatoria de la glucólisis.
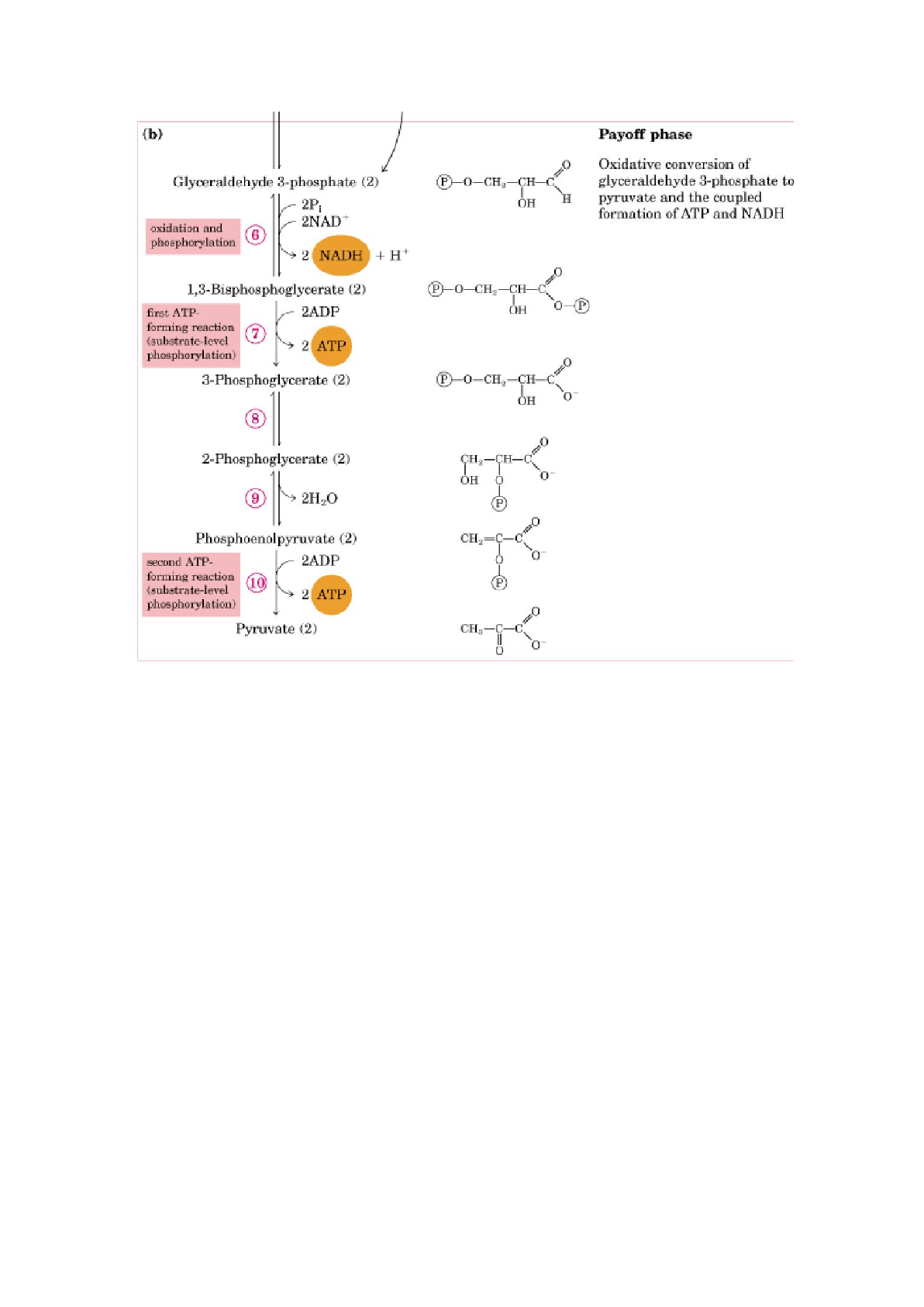
Fase de beneficios de la glucólisis:
La fase de beneficios de la glucólisis incluye los pasos de fosforilación
conservadores de energía en los que parte de la energía libre de la molécula de
glucosa se conserva en forma de ATP y NADH.
Una molécula de glucosa produce 2 moléculas de gliceraldehído-3-fosfato, las
dos mitades de la molécula de glucosa siguen la misima ruta en la segunda fase
de la glucólisis. La conversión de 2 moléculas de gliceraldehído-3-fosfato en 2
moléculas de piruvato se acompaña de la formación de 4 moléculas de ATP a
partir de ADP. Sin embargo, el rendimiento neto de ATP por molécula de
glucosa degradada es sólo de dos, porque se han invertido 2 moléculas de ATP
en la fase preparatoria de la glucólisis para fosforilar los 2 extremos de la
molécula de hexosa.
6. Oxidación del gliceraldehído 3-fosfato a 1,3-bisfosfoglicerato
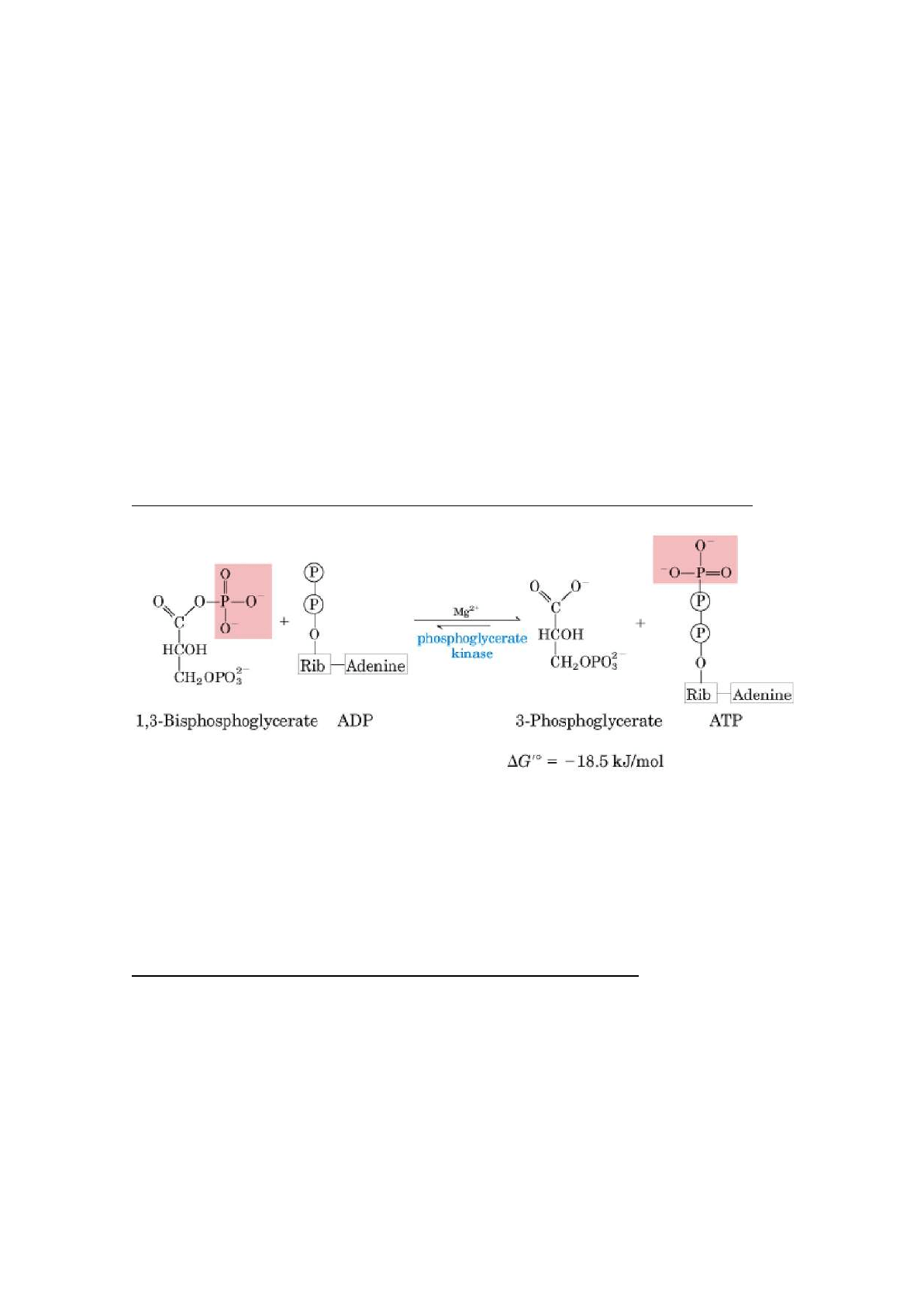
Reacción catalizada por el gliceraldehído-3-fosfato deshidrogenasa. Ésta es la
primera de las 2 reacciones conservadoras de energía de la glucólisis que
conducen en último término a la formación de ATP.
Mecanismo de acción de la gliceraldehído-3 fosfato deshidrogenasa: formación
de un intermediario de alta energía unido covalentemente al enzima como
mecanismo de acoplamiento energético. La G liberada en la oxidación de un C
se convierte en un grupo fosfato de alta energía o con alto potencial para la
transferencia del fosforilo.
7. Transferencia de fosforilo desde el 1,3-bisfosfoglicerato al ADP
El enzima fosfoglicerato quinasa transfiere el grupo fosforilo de alta energía
desde el grupo carboxilo del 1,3-bisfosfoglicerato al ADP, obteniéndose ATP y
3-fosfoglicerato.
Ésta reacción es la fosforilación del ADP para formar ATP a expensas de la
energía del sustrato que es el 1,3BPG. Por eso a ésta reacción se le llama
fosforilación a nivel de sustrato (FNS).
8. Conversión del 3-fosfoglicerato en 2-fosfoglicerato
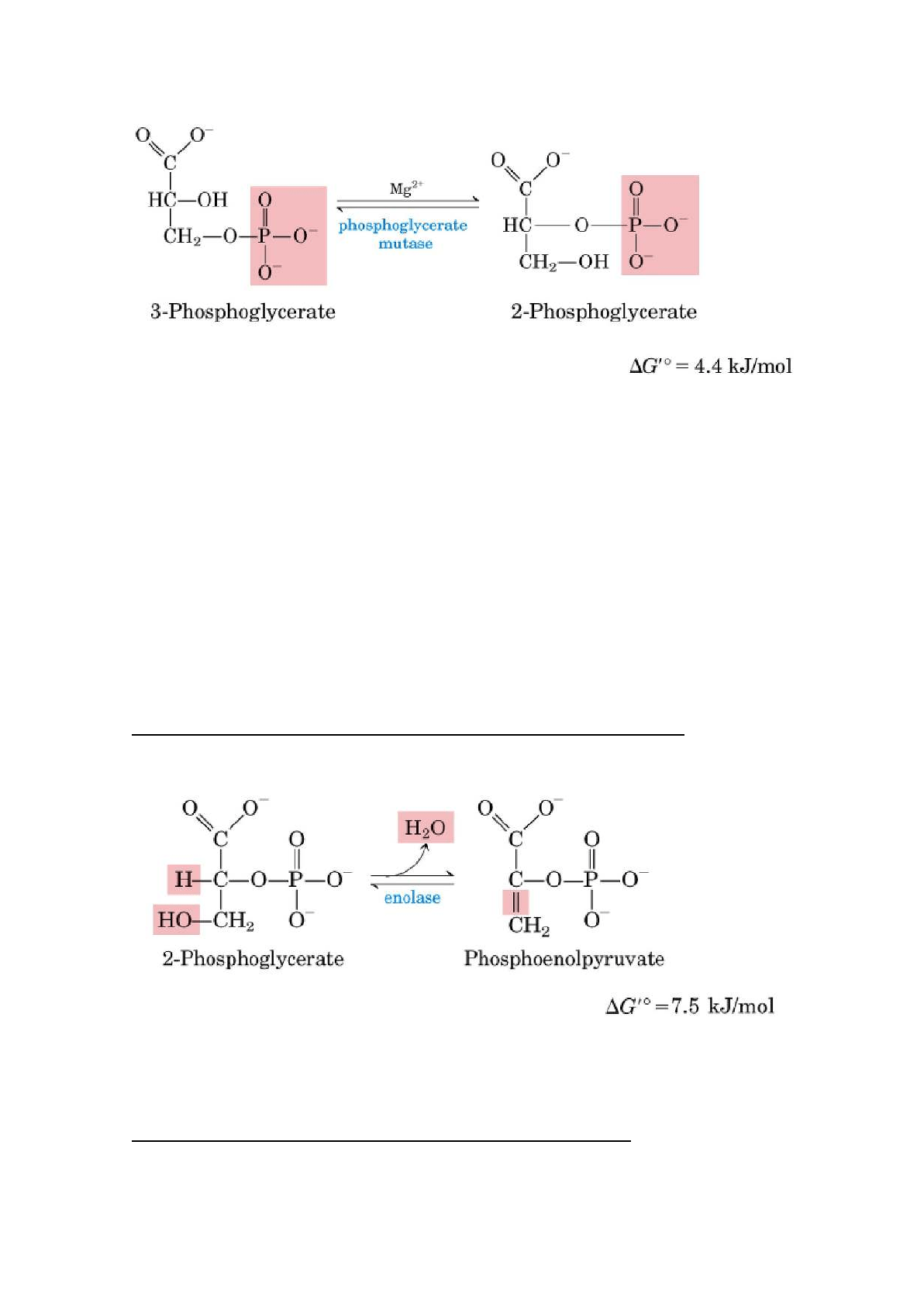
El enzima fosfoglicerato mutasa cataliza un desplazamiento reversible del grupo
fosforilo entre C-2 y C-3 del glicerato. El magnesio es esencial para ésta
reacción.
9. Deshidratación del 2-fosfoglicerato a fosfoenolpiruvato
La enzima enolasa promueve la eliminación reversible de una molécula de agua
del 2-fosfoglicerato, dando fosfoenolpiruvato (PEP).
10. Transferencia del grupo fosforilo del PEP al ADP
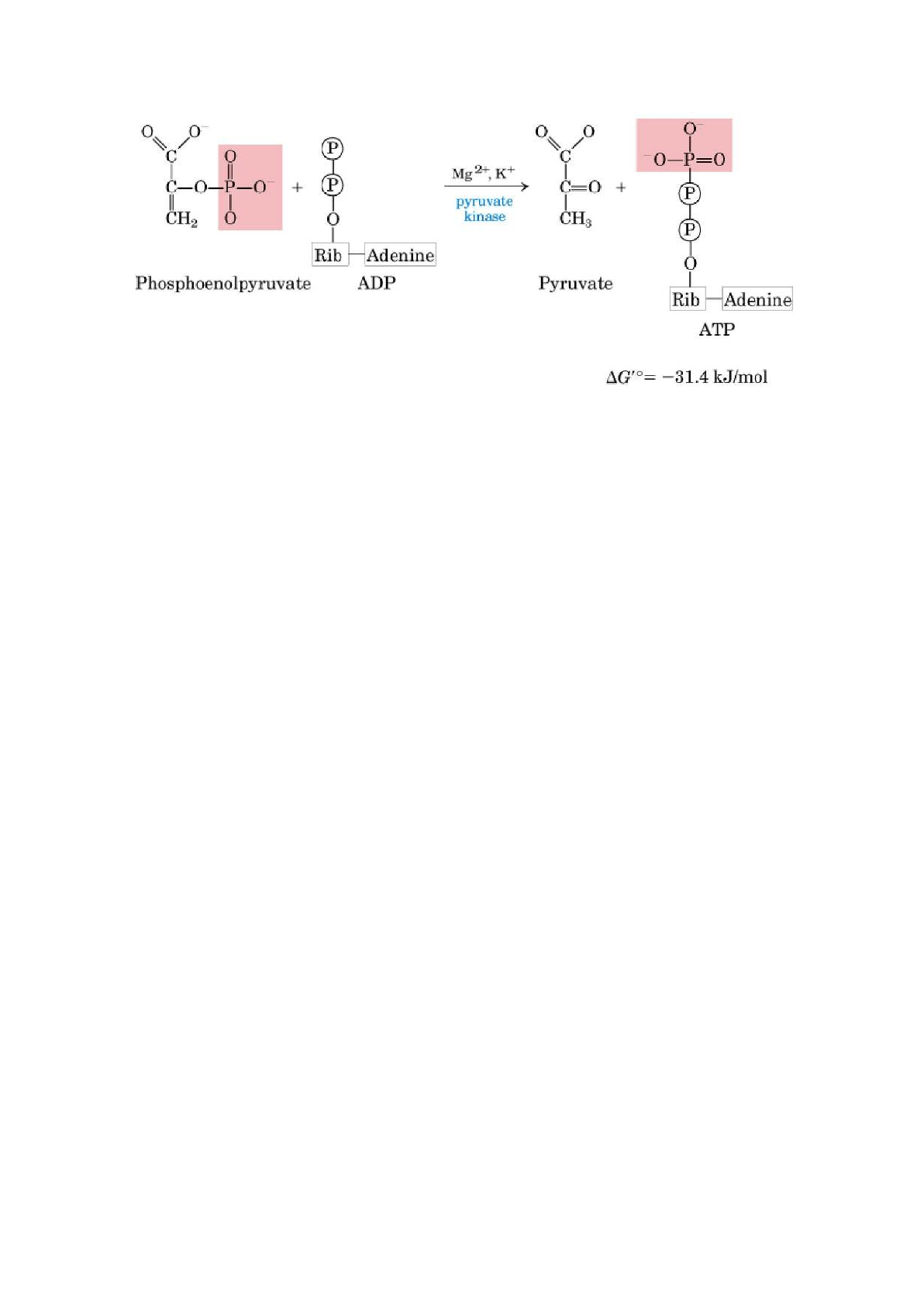
Reacción catalizada por la piruvato quinasa, que requiere potasio, magnesio o
manganeso. Es la segunda fosforilación a nivel de sustrato, por lo cual se forma
ATP.
Moduladores positivos: F-1,6DP
Moduladores negativos: ATP, acetil-CoA, ácidos grasos de cadena larga, alanina.
La piruvato quinasa también posee regulación covalente, se inhibe por
fosforilación (en hígado).
Balance energético de la glucólisis:
El primer miembro de la ecuación siguiente muestra todas las entradas de ATP,
NAD⁺, ADP y Pi; mientras que el segundo miembro muestra todo lo que se
produce.
Glucosa+2ATP+2NAD⁺+4ADP+2Pi→
2piruvato+2ADP+2NADH+2H⁺+4ATP+2H₂O
Simplificando,
Glucosa+2NAD⁺+2ADP+2Pi→ 2 Piruvato+2 NADH+ 2 H⁺+ 2ATP+ 2 H₂O
En el proceso glucolítico global, se convierte una molécula de glucosa en 2
moléculas de piruvato.
Dos moléculas de ADP y 2 de Pi se convierten en 2 moléculas de ATP.
Finalmente, 4 electrones, en forma de iones hidruro, se transfieren desde 2
moléculas de gliceraldehído-3.fosfato a 2 de NAD⁺.
Destinos del Piruvato:
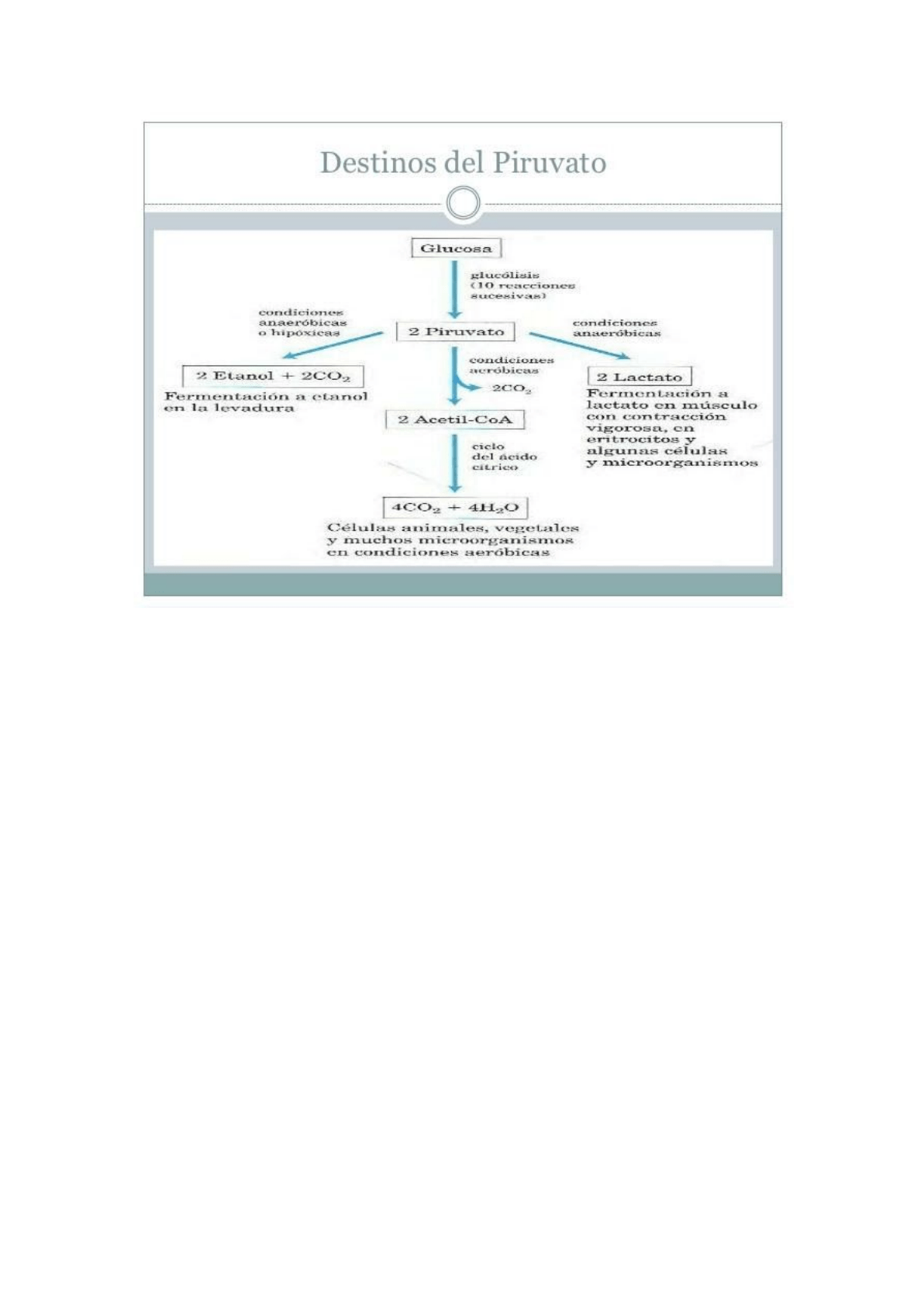
Cuando se produce la glucólisis anaerobia hasta lactato, no hay una oxidación
neta ni producción o consumo de NADH o NAD⁺. La glucólisis anaerobia hasta
lactato ocurre en el citosol, la enzima que convierte al piruvato en lactato es la
lactato deshidrogenasa y la reacción es reversible. La fermentación láctica es
una reducción necesaria para regenerar NAD⁺. La fermentación alcohólica es
para regenerar NAD⁺ en microorganismos.
En condiciones aeróbicas, la glucólisis sólo constituye el primer paso en la
degradación completa de la glucosa. El piruvato se oxida, con pérdida de su
grupo carboxilo en forma de CO₂, dando el grupo acetilo del acetil-coenzima A,
que es oxidado seguidamente a CO₂ por el ciclo de Krebs. Los electrones de
estas oxidaciones pasan al oxígeno a través de una cadena de transportadores en
la mitocondria, formando agua. La energía procedente de las reacciones de
transferencia electrónica impulsa la síntesis de ATP en la mitocondria.
Rutas alimentadoras de la glucólisis
Gran número de glúcidos (además de la glucosa) entran en la ruta glucolítica,
después de ser transformados en uno de los intermediarios glucolítocos. Los
más significativos son:
● Polisacáridos: glucógeno y almidón
● Disacáridos: maltosa, lactosa, tehalosa, sacarosa
● Monosacáridos: fructosa, manosa, galactosa

GLUCÓLISIS.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.