
5
Capítulo
Estructura y función
de la membrana celular
Una serpiente de cascabel se prepara para atacar. (Detalle)
Araña reclusa parda.
Estudio de caso
Venenos nocivos
DESDE MUCHO ANTES DE LAS VACACIONES ESCOLARES,
Justin Schwartz, de 13 años, esperaba su estancia de tres
semanas en el Parque Nacional de Yosemite. El 21 de
julio de 2002, después de una caminata de más de siete
kilómetros, Justin se sentó a descansar sobre unas rocas
soleadas, con las manos sueltas a sus costados. De pronto,
sintió un dolor que le perforaba la palma de la mano
derecha. Una serpiente de cascabel de metro y medio, que
probablemente se sintió amenazada por el brazo de Justin,
lo atacó sin advertencia.
Sus compañeros vieron alarmados cómo la serpiente
se escurría por entre la maleza, pero Justin estaba
concentrado en su mano, pues el dolor se había vuelto
insoportable y la palma estaba hinchada. De pronto, se
sentía débil y mareado. Los consejeros y compañeros lo
cargaron durante cuatro horas por el sendero de regreso.
Entre tanto, el dolor se extendía por todo el brazo de
Justin, cada vez más decolorado; además, sentía como
si su mano fuera a reventar. Un helicóptero los trasladó
a un hospital, donde cayó inconsciente. Un día después,
recuperó la conciencia en el University of California Davis
Medical Center (Centro Médico Davis de la Universidad
de California). Ahí, durante un mes, Justin se sometió a
10 cirugías destinadas a aliviar la enorme presión por la
inflamación del brazo, a retirar tejido muscular muerto
e inició un largo proceso para reparar los extensos daños
de mano y brazo.
El suplicio de Diane Kiehl comenzó cuando se vestía
para una ceremonia conmemorativa informal con su
familia el 27 de mayo de 2006. Al enfundarse los jeans
que había dejado tirados en el baño la noche anterior,
sintió una picadura en el muslo derecho. Se sacó los jeans
y miró con angustia como huía una Loxosceles reclusa
parda. Como vivía en una vieja casa de la parte rural de
Kansas, Diane se había acostumbrado a la visita frecuente,
aunque indeseable, de arañas. Los dos pequeños puntos
de la mordedura le parecieron una molestia menor,
pero el día siguiente apareció una erupción extensa y
molesta. Al tercer día el dolor intermitente la atacaba
como si le clavaran un cuchillo en el muslo. Un médico le
dio analgésicos, esteroides para reducir la inflamación y
antibióticos para combatir las bacterias introducidas por
la mordedura de la araña. Los siguientes 10 días fueron
una pesadilla de dolor por la llaga creciente, que se cubrió
de ampollas supurantes y sangre reseca. La lesión tardó
cuatro meses en sanar. Un año después, a veces Diane
siente dolor en la gran cicatriz que quedó.
¿Por qué el veneno de la serpiente de cascabel y de
la araña reclusa parda lastiman los vasos sanguíneos,
desintegran piel y tejidos, y en ocasiones producen en
todo el cuerpo síntomas que ponen en peligro la vida?
¿Qué tienen que ver los venenos con las membranas
celulares?

78 UNIDAD 1 La vida de la célula
5.1
¿CÓMO SE RELACIONA LA ESTRUCTURA
DE UNA MEMBRANA CON SU FUNCIÓN?
Las membranas celulares aíslan el contenido
de la célula y permiten la comunicación
con el entorno
Como se sabe, todas las células y organelos de las células euca-
riontes están rodeados por membranas. Las membranas celulares
cumplen varias funciones cruciales:
• Aíslan de forma selectiva el contenido de la célula del
ambiente externo, de modo que se producen gradientes de
concentración de sustancias disueltas producidas en diversas
partes de la membrana.
• Regulan el intercambio de compuestos esenciales entre la
célula y el medio acuoso extracelular o entre los organelos
envueltos en membranas y el citoplasma del entorno.
• Permiten la comunicación entre células.
• Permiten las uniones en el interior de las células y entre ellas.
• Regulan muchas reacciones bioquímicas.
Son tareas formidables para una estructura tan delgada que 10 mil
membranas, una sobre otra, apenas igualarían el grosor de esta pá-
gina. La clave del funcionamiento de la membrana radica en su
estructura. Las membranas no son unas puras láminas uniformes,
sino que son estructuras complejas y heterogéneas cuyas moléculas
que las conforman cumplen funciones distintas. Las membranas
varían según el tipo de tejido y cambian de manera dinámica en
reacción al entorno.
Todas las membranas de una célula tienen una estructura
básica parecida: proteínas que flotan en una bicapa de fosfolí-
pidos (véanse las páginas 44-45). Los fosfolípidos realizan la
función aislante de las membranas, mientras que las proteínas
intercambian sustancias selectivamente y se comunican con el
entorno, controlan las reacciones bioquímicas de la membrana
celular y forman enlaces.
Las membranas son “mosaicos fluidos”
en los que las proteínas se desplazan
en capas de lípidos
Antes de la década de 1970, aunque los biólogos sabían que las
membranas celulares constan principalmente de proteínas y lípi-
dos, no se sabía cómo estas moléculas originan la estructura y la
función de la membrana. En 1972, los investigadores de la célula
S.J. Singer y G.L. Nicolson desarrollaron el modelo del mosaico
fluido de la membrana celular, que ahora se sabe que es correcto.
Según este modelo, cada membrana consta de un mosaico o “par-
che” de diferentes proteínas que cambian de manera constante y
se mueven en un fluido viscoso (grueso y pegajoso) constituido
por una bicapa de fosfolípidos (
FIGURA 5-1). Aunque los com-
ponentes de la membrana plasmática se mantienen relativamente
constantes, la distribución general de las proteínas y los tipos de
fosfolípidos cambia con el tiempo. Veamos con más detalle la es-
tructura de las membranas.
La bicapa de fosfolípidos es la parte fluida
de la membrana
Como vimos en el capítulo 3, un fosfolípido consta de dos partes
muy diferentes: una “cabeza” que es polar e hidrofílica (atraída por
el agua) y un par de “colas” de ácidos grasos no polares que son
hidrofóbicas (que no las atrae el agua). Las membranas contienen
muchos fosfolípidos del tipo general que se muestra en la
FIGURA
5-2
. Observa que en este fosfolípido particular, un enlace doble
(que hace al ácido graso insaturado) crea una flexión en la cola
del ácido graso que ayuda a mantener la fluidez de la membrana.
Estudio de caso Venenos nocivos
5.1 ¿Cómo se relaciona la estructura de una
membrana con su función?
Las membranas celulares aíslan el contenido de la célula
y permiten la comunicación con el entorno
Las membranas son “mosaicos fluidos” en los que las
proteínas se desplazan en capas de lípidos
La bicapa de fosfolípidos es la parte fluida de la membrana
De cerca Forma, función y fosfolípidos
Estudio de caso continuación Venenos nocivos
Diversas proteínas forman un mosaico dentro
de la membrana
5.2 ¿Cómo pasan las sustancias por las membranas?
Las moléculas de los fluidos se mueven en respuesta
a gradientes
El movimiento a través de las membranas ocurre por
transporte pasivo y activo
De un vistazo
El transporte pasivo es por difusión simple, difusión
facilitada y ósmosis
Investigación científica El descubrimiento
de las acuaporinas
El transporte que requiere energía es transporte activo,
endocitosis y exocitosis
El intercambio de materiales por las membranas influye
en el tamaño y la forma de la célula
Membrane Transport (disponible en inglés)
5.3 ¿Cómo las uniones especializadas permiten
a las células establecer conexiones y comunicarse?
Los desmosomas unen a las células
Las uniones estrechas impiden las filtraciones
en las células
Las uniones en hendidura o gap y plasmodesmos permiten
la comunicación entre células
Estudio de caso otro vistazo Venenos nocivos
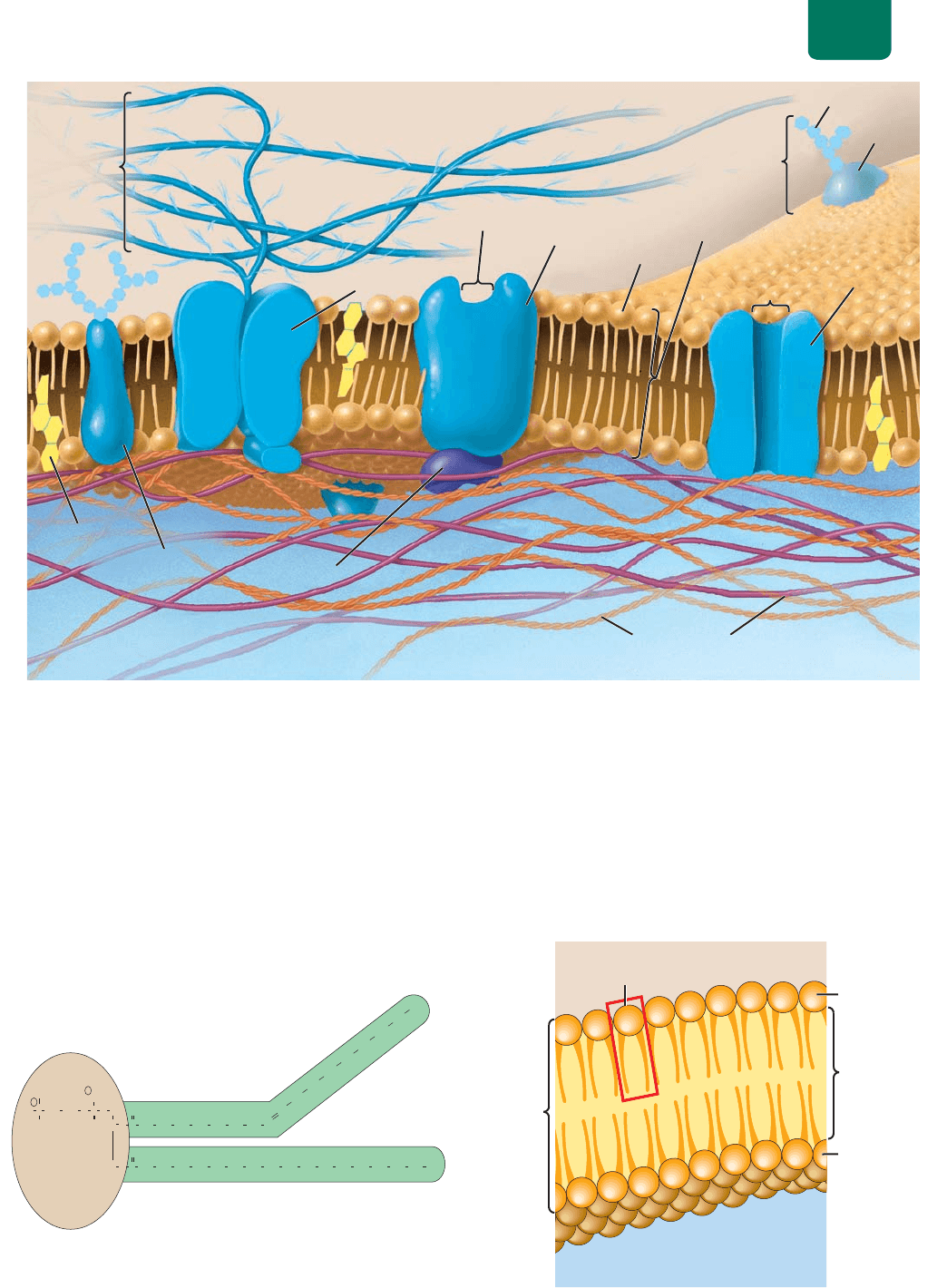
Estructura y función de la membrana celular Capítulo 5 79
Todas las células están rodeadas por agua. Los organismos
unicelulares viven en agua dulce o en el mar, el agua satura las
paredes celulares de las plantas y las células animales están baña-
das por un medio acuoso extracelular débilmente salino que sale
de la sangre. El citosol (la parte líquida del citoplasma) es prin-
cipalmente agua. Así, las membranas plasmáticas separan el cito-
sol acuoso del entorno acuoso exterior y las membranas internas
rodean compartimentos acuosos dentro de la célula. Rodeados
por agua, los fosfolípidos se organizan de forma espontánea en
una doble capa llamada bicapa lipídica (
FIGURA 5-3). El hidró-
geno forma enlaces entre el agua y las cabezas hidrofílicas de los
fosofolípidos, de modo que éstas se orientan hacia afuera, hacia
JOXFRSURWHķQD
SRUR
SxX\PKVL_[YHJLS\SHYL_[LYPVY
JP[VWSHZTHPU[LYPVY
FROHVWHURO
ELFDSDGH
IRVIROķSLGRV
SURWHķQD
GHWUDQVSRUWH
SURWHķQD
UHFHSWRUD
IRVIROķSLGR
SURWHķQDGH
UHFRQRFLPLHQWR
PDWUL]
H[WUDFHOXODU
HQ]LPD
FLWRHVTXHOHWR
FDUERKLGUDWR
SXQWRGHXQLʼnQ
SURWHķQD
SURWHķQDGHXQLʼnQ
FIGURA 5-1 La membrana plasmática La membrana plasmática es una bicapa de fosfolípidos
entremezclados con moléculas de colesterol que forman una matriz fluida en la que están inmersas
varias proteínas (azul). Muchas proteínas llevan unidos carbohidratos para formar glucoproteínas. Se
ilustran las proteínas de reconocimiento, unión, recepción y transporte.
cabeza
(hidrofílica)
colas
(hidrofóbica)
2
CH
CH
2
CH
2
CH
2
CH
2
CH
2
CH
2
CH
2
CH
3
CH
CH
2
CH
2
CH
2
CH
2
CH
CH
2
C
O
CH
2
N
CH
2
CH
2
OP
3
CH
CH
3
CH
3
O
O
–
O
CH
C
2
OH
+
CH
2
CH
2
CH
2
CH
2
CH
2
CH
2
CH
2
CH
C
CH
2
CH
2
CH
2
CH
2
CH
2
CH
2
CH
2
CH
2
CH
3
2
O
C
O
H
2
FIGURA 5-2 Un fosfolípido Observa que un doble enlace
en una de las colas de ácidos grasos, hace que se flexione.
fosfolípido
cabezas
hidrofílicas
colas
hidrofóbicas
cabezas
hidrofílicas
líquido extracelular
(medio acuoso)
citoplasma
(medio acuoso)
bicapa
FIGURA 5-3 Bicapa de fosfolípidos de la membrana celular

80 UNIDAD 1 La vida de la célula
más fluido menos fluido
FIGURA 5-4 Las flexiones aumentan la fluidez
el medio acuoso a ambos lados de la membrana. Las colas de
los fosfolípidos, como son hidrofóbicas, se colocan dentro de la
bicapa.
Las moléculas individuales de los fosfolípidos no están
unidas unas con otras y las membranas contienen fosfolípidos
con ácidos grasos insaturados cuyos enlaces dobles flexionan sus
colas (véase la figura 5-2). Gracias a estas propiedades, los fosfo-
lípidos se mueven dentro de las capas, con lo que la bicapa es
bastante fluida. Las membranas con más dobles enlaces en las co-
las de sus fosfolípidos son más fluidas que las membranas cuyos
fosfolípidos tienen menos dobles enlaces (
FIGURA 5-4).
Las células tienen diferentes grados de saturación (y, por
consiguiente, de fluidez) en la bicapa de fosfolípidos, lo que
les permite realizar diferentes actividades o funcionar en distin-
tos ambientes. Por ejemplo, las membranas tienden a volverse
más fluidas a temperaturas más altas (porque las moléculas se
mueven con mayor rapidez) y son menos fluidas con las tem-
peraturas bajas (cuando las moléculas son más lentas). Por con-
siguiente, las membranas celulares de los organismos que viven
en ambientes fríos tienen abundantes fosfolípidos insaturados,
para que con las colas flexionadas, la membrana conserve la flui-
dez que se necesita (véase el apartado “De cerca: Forma, función
y fosfolípidos”).
Casi todas las moléculas biológicas, incluyendo las sales,
aminoácidos y carbohidratos, son hidrofílicas, es decir, son po-
lares y solubles en agua. Estas sustancias no pasan de forma fácil
por entre las colas hidrofóbicas y no polares de los ácidos grasos
de la bicapa de fosfolípidos. Esta capa cumple una de las primeras
cinco funciones que se describieron en párrafos anteriores: aislar
de forma selectiva el contenido de la célula del ambiente exterior.
Sin embargo, el aislamiento que crea la membrana plas-
mática no es completo. Como se verá después, moléculas muy
pequeñas (de agua, oxígeno y dióxido de carbono), así como mo-
De cerca Forma, función y fosfolípidos
Para funcionar de forma adecuada, las membranas celulares
deben mantener la mejor fluidez. Así como la mantequilla se
funde en la sartén y el aceite se solidifica en el congelador, la
fluidez de la membrana celular es sensible a los cambios de
la temperatura. Muchos organismos, incluidos los protistas,
bacterias, plantas, animales de “sangre fría” (ranas, peces
y serpientes) y mamíferos que hibernan, sufren grandes
fluctuaciones de temperatura corporal. Las cantidades relativas
de colas de ácidos grasos saturados e insaturados en los
fosfolípidos de la membrana afectan la fluidez de ésta. Las
células pueden modificar la composición de su membrana
para mantener la fluidez a diferentes temperaturas. A mayores
temperaturas, se insertan fosfolípidos con más ácidos grasos
saturados, mientras que con temperaturas más bajas se
agregan ácidos grasos insaturados.
Algunos mamíferos hibernantes de sangre caliente tienen
adaptaciones parecidas. Por ejemplo, los caribúes (que viven
en regiones al norte) mantienen una temperatura básica
cercana a 38 C, pero la temperatura de sus patas puede bajar
casi hasta congelarse (0 C), de modo que conservan el calor
corporal al estar sobre la nieve de invierno (
FIGURA E5-1). En
las patas del caribú, las membranas de las células cerca de las
pezuñas congeladas tienen muchos ácidos grasos insaturados,
mientras que las células cercanas al tronco tibio poseen más
ácidos grasos saturados. De esta manera, las membranas
plasmáticas tienen la fluidez necesaria a lo largo de las patas,
pese a las grandes diferencias de temperatura.
La presión elevada reduce la fluidez de las membranas.
Los moradores de las profundidades del mar sufren presiones
inmensas, así como temperaturas cercanas a la congelación,
una combinación que volvería a la membrana de células
humanas demasiado rígida para funcionar. Como es fácil
anticipar, la membrana celular de los animales abisales
tienen una proporción elevada de ácidos grasos insaturados
(más como el aceite vegetal que como la mantequilla). Estos
animales, como están adaptados a la presión elevada y a las
bajas temperaturas, no sobreviven si los sacan a la superficie.
Los investigadores postulan la hipótesis de que a la presión de
la superficie, la membrana de sus células se vuelve demasiado
fluida y permeable, con lo que las células no pueden mantener
su gradiente normal y mueren.
FIGURA E5-1 Caribúes en busca de alimento
Estructura y función de la membrana celular Capítulo 5 81
tos unidos a la parte expuesta de la membrana celular (véase la
figura 5-1) se llaman glucoproteínas (gluco viene del término
griego que significa “dulce” y se refiere a la parte de los carbohi-
dratos con sus unidades de azúcar). Casi todas las proteínas de la
membrana plasmática están insertadas, al menos parcialmente,
en la bicapa de fosfolípidos, pero algunas se adhieren a la su-
perficie. Las proteínas de membrana pueden clasificarse en cinco
grandes categorías basadas en su función: receptoras, de recono-
cimiento, enzimáticas, de unión y de transporte.
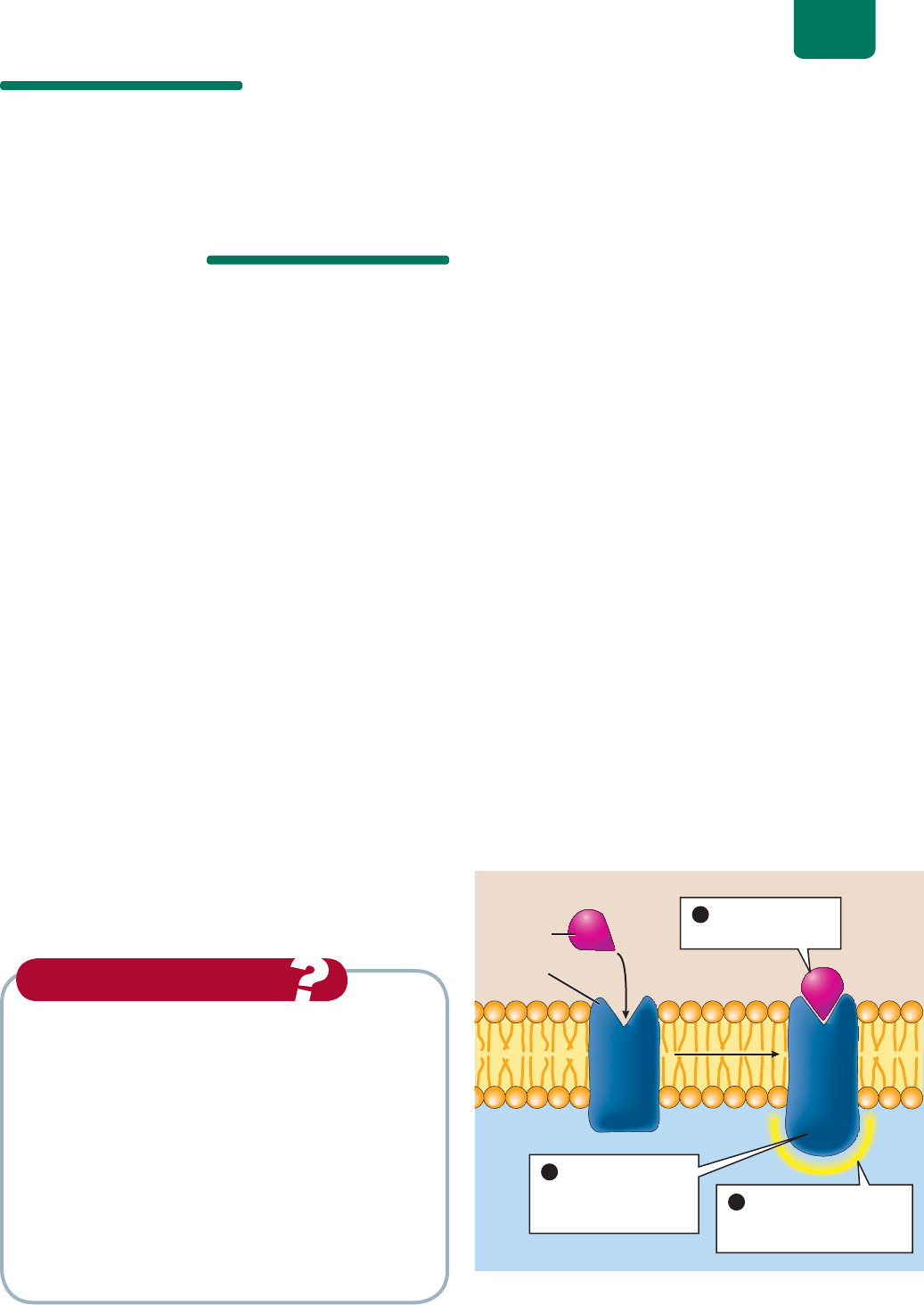
Casi todas las células llevan docenas de tipos de proteínas
receptoras (algunas de las cuales son glucoproteínas) repartidas
por la membrana plasmática. Para realizar sus funciones, las cé-
lulas tienen que responder a los mensajes enviados por otras. Es-
tos mensajes son moléculas (como las hormonas) transportadas
por el torrente sanguíneo. Después de penetrar por difusión en el
líquido extracelular, estas moléculas mensajeras se unen en pun-
tos específicos de las proteínas receptoras, las cuales comunican
el mensaje al interior de la célula (
FIGURA 5-5). Cuando la mo-
lécula apropiada se une a la receptora, ésta se “activa” (a menudo
con un cambio de forma), lo que produce una respuesta dentro
de la célula.
La respuesta puede ser muy diversa. La forma de una pro-
teína dentro de una célula puede modificarse y pasar de inactiva
a activa. Este cambio estimula una secuencia de reacciones quími-
cas en la célula que alteran su actividad. Por ejemplo, cuando la
hormona epinefrina (adrenalina) se enlaza a un receptor especí-
fico de la membrana en las células musculares, las estimula para
que degraden el glucógeno en glucosa y aporten más energía
para la contracción de los músculos. Cuando otras proteínas re-
ceptoras se unen a moléculas mensajeras, abren los canales ióni-
cos o inician secuencias de reacciones que estimulan a las células
para que se dividan o produzcan hormonas. La comunicación
entre las células nerviosas depende también de los receptores y
gracias a las proteínas receptoras las células del sistema inmune
reconocen y atacan a los invasores que causan enfermedades.
léculas más grandes sin carga y solubles en lípidos, pueden pasar
por la bicapa.
En la mayoría de las células animales, la bicapa de fosfolí-
pidos de la membrana contiene también colesterol (véase la figu-
ra 5-1). Algunas membranas celulares tienen pocas moléculas de
colesterol; otras tienen tantas de colesterol como de fosfolípidos.
El colesterol afecta la estructura y el funcionamiento de la mem-
brana de varias maneras: estabiliza la bicapa de fosfolípidos, de
modo que sea menos fluida a temperaturas elevadas y menos só-
lida con las bajas, además de ser menos permeable a sustancias
solubles en agua, como iones o monosacáridos.
La naturaleza flexible y un tanto fluida de la bicapa es muy
importante para el funcionamiento de la membrana. Cada vez
que respiras, mueves los ojos o pasas las páginas de este libro, las
células de tu cuerpo cambian de forma. Si las membranas plas-
máticas fueran rígidas en lugar de flexibles, las células podrían
romperse y morir. Además, como se expuso en el capítulo 4, las
membranas de las células eucariontes están en movimiento cons-
tante. Los compartimentos envueltos en membranas transportan
sustancias dentro de la célula, toman y expulsan material al exte-
rior y en estas actividades fusionan sus membranas. Este flujo y
fusión de las membranas es posible gracias a la naturaleza fluida
de la bicapa de fosfolípidos.
Diversas proteínas forman un mosaico
dentro de la membrana
Miles de proteínas de membrana están insertadas o unidas a la
superficie de la bicapa de fosfolípidos de la membrana celular.
Las proteínas de la membrana plasmática que llevan carbohidra-
Estudio de caso continuación
Venenos nocivos
Algunos de los efectos más devastadores del veneno de
ciertas serpientes y arañas se deben a que contienen enzimas,
llamadas fosfolipasas, que degradan los fosfolípidos de la
membrana celular y producen la muerte de las células.
¿Te has preguntado...,
qué hacer si te muerde una serpiente venenosa?
Si hay especies de serpientes venenosas en la región donde
vives, fíjate en los colores y el tamaño de la serpiente que te
mordió para poder identificarla cuando llames a los servicios
de urgencias. El doctor David Erk del Wyoming Medical Center
(Centro Médico de Wyoming), interesado particularmente
en mordidas de serpiente, ofrece el siguiente consejo: en
primer lugar, pide ayuda médica de inmediato. Para retrasar
la propagación del veneno y la inflamación que causa, mantén
inmovilizado el miembro mordido, idealmente apenas por
debajo de la altura a la que se encuentra el corazón. Quita
adornos y accesorios apretados alrededor de la herida. No
cortes la herida ni la succiones. No apliques un torniquete, calor
ni hielo. No bebas café ni alcohol, ni tomes medicina alguna.
Conserva la calma lo más que puedas.
UHFHSWRU
KRUPRQD
JP[VWSHZTH
MS\PKVL_[YHJLS\SHY
8QDKRUPRQD
VHXQHDOUHFHSWRU
/DXQLʼnQGHOD
KRUPRQDDFWLYDDO
UHFHSWRUTXHFDPELD
GHIRUPD
(OUHFHSWRUDFWLYDGR
HVWLPXODXQDUHVSXHVWD
GHODFÒOXOD
FIGURA 5-5 Activación de proteínas receptoras

82 UNIDAD 1 La vida de la célula
Las proteínas de reconocimiento son glucoproteínas que
sirven como etiquetas de identificación (véase la figura 5-1). Las cé-
lulas de cada individuo llevan glucoproteínas únicas que las iden-
tifican como “yo”. Las células del sistema inmunitario ignoran al
yo y atacan a las células invasoras, como las bacterias, que tienen
diferentes células de reconocimiento en la membrana. Las pro-
teínas de reconocimiento de la superficie de los glóbulos rojos
llevan distintos grupos de carbohidratos y determinan si la san-
gre es tipo O, A, B o AB (véase tabla 10-1, en la página 187). Las
transfusiones, así como los órganos trasplantados, deben tener
glucoproteínas que concuerden con las del receptor para reducir
al mínimo los ataques del sistema inmunitario.
Las enzimas son proteínas que catalizan las reacciones quí-
micas que sintetizan o degradan las moléculas biológicas, como
se verá en el capítulo 6. Aunque muchas enzimas están situadas
en el citoplasma, algunas se extienden a la membrana celular y
otras están unidas a la superficie de las membranas (véase la figu-
ra 5-1). Las enzimas de la membrana plasmática incluyen las que
sintetizan las proteínas y carbohidratos de la matriz extracelular
(una red de fibras de proteínas y glucoproteínas que llena los es-
pacios entre las células animales).
Un grupo variado de proteínas de unión ancla las mem-
branas celulares de diversas maneras. Algunas se extienden por
la membrana plasmática y sostienen el citoesqueleto dentro de la
célula, con la matriz extracelular fuera (véase la figura 5-1), de
modo que la célula conserve su lugar en un tejido. Algunas pro-
teínas de unión mantienen la forma de la célula al enlazar la
membrana plasmática al citoesqueleto y otras adhieren a la cé-
lula y la mueven por las superficies. Otras proteínas de unión
establecen conexiones entre células contiguas, como se verá más
adelante (véanse las figuras 5-17 y 5-18).
Las proteínas de transporte regulan el movimiento de
las moléculas hidrofílicas por la membrana plasmática. Algunas
proteínas de transporte, llamadas proteínas de canal, forman
canales por cuyos poros centrales las moléculas de agua o iones
específicos atraviesan la membrana siguiendo el gradiente de
concentración (véase la figura 5-1). Otras proteínas de transpor-
te, llamadas proteínas portadoras, tienen lugares de enlace en
los que se unen temporalmente a las moléculas en un lado de la
membrana. Enseguida, estas proteínas cambian de forma (a veces
tomando energía del ATP de la célula), pasan la molécula por la
membrana y la depositan del otro lado. En secciones posteriores
se describirán estas proteínas de transporte.
5.2
¿CÓMO PASAN LAS SUSTANCIAS
POR LAS MEMBRANAS?
Las moléculas de los fluidos se mueven
en respuesta a gradientes
Ya sabes que las sustancias atraviesan las membranas por difusión
en la bicapa de fosfolípidos o pasan por proteínas de transporte es-
pecializado. Para entender mejor este fenómeno se requieren defi-
niciones y conocimientos previos. Como la membrana plasmática
separa el medio acuoso citoplasmático del extracelular, se iniciará
el estudio del transporte en las membranas con un análisis de las
características de los fluidos, empezando con algunas definiciones:
• Un fluido es toda sustancia cuyas moléculas pueden desli-
zarse unas en otras; como resultado, los fluidos no tienen
forma propia. Son fluidos los gases, los líquidos y también
las membranas celulares, cuyas moléculas pueden deslizarse
unas sobre otras.
• Un soluto es una sustancia que puede disolverse (dispersarse
en átomos, moléculas o iones individuales) en un disol-
vente, que es un fluido (normalmente un líquido) capaz
de disolver el soluto. El agua, en la que ocurren todos los
procesos biológicos, disuelve tantos solutos que es llamada
“el disolvente universal”.
• La concentración de una sustancia define la cantidad de
soluto en una cantidad dada del disolvente. Por ejemplo, la
concentración de la solución de azúcar es una medida del
número de moléculas de azúcar contenidas en un volumen
dado de la solución.
• Un gradiente es una diferencia física en propiedades como la
temperatura, presión, carga eléctrica o concentración de una
sustancia en un fluido entre dos espacios contiguos. Se requiere
energía para formar gradientes. Con el tiempo, los gradientes
se disuelven, salvo que se aporte energía para conservarlos o los
separe una barrera eficaz. Por ejemplo, los gradientes de tem-
peratura causan un flujo de energía de la región de temperatura
alta a la región de menor temperatura. Los gradientes eléctricos
pueden impulsar el movimiento de iones. Los gradientes de
concentración o presión hacen que se muevan iones o mo lécu-
las de una región a otra en el sentido en que se equilibra la
diferencia. Las células utilizan energía y las propiedades
únicas de la membrana para generar gradientes de concen-
tración de varias moléculas y iones disueltos en su citosol en
relación con el entorno acuoso.
Es importante tener presente que a temperaturas sobre el cero
absoluto (–273 °C), átomos, moléculas y iones se encuentran
en constante movimiento. Conforme aumenta la temperatura,
el movimiento se incrementa y a las temperaturas a las que es
posible que se desarrolle la vida, estas partículas se mueven con
mucha rapidez. Como resultado de este movimiento, moléculas
y iones en solución chocan de forma constante unas con otras y
con las estructuras del medio. Con el tiempo, los movimientos
azarosos producen un movimiento neto de las regiones de alta
concentración a las de baja concentración que se llama difusión.
En un sistema inerte, si nada se opone al movimiento (los facto-
res que se oponen son la carga eléctrica, diferencia de presión o
barreras físicas), la agitación aleatoria de las moléculas continúa
hasta que se encuentren dispersas uniformemente por todo el
fluido.
Para imaginar cómo el movimiento aleatorio de moléculas
o iones de un fluido termina por deshacer los gradientes de con-
centración, piensa en un cubo de azúcar que se disuelve en café
caliente o las moléculas de un perfume que salen al aire de un
frasco abierto. En cada caso, hay un gradiente de concentración.
Si dejas el perfumero abierto o te olvidas del café, al final queda
un frasco vacío y una habitación aromatizada o un café frío y dul-
ce. En una analogía con la gravedad, decimos que las moléculas
que pasan de regiones de mayor a menor concentración “bajan”
por su gradiente de concentración.
Estructura y función de la membrana celular Capítulo 5 83
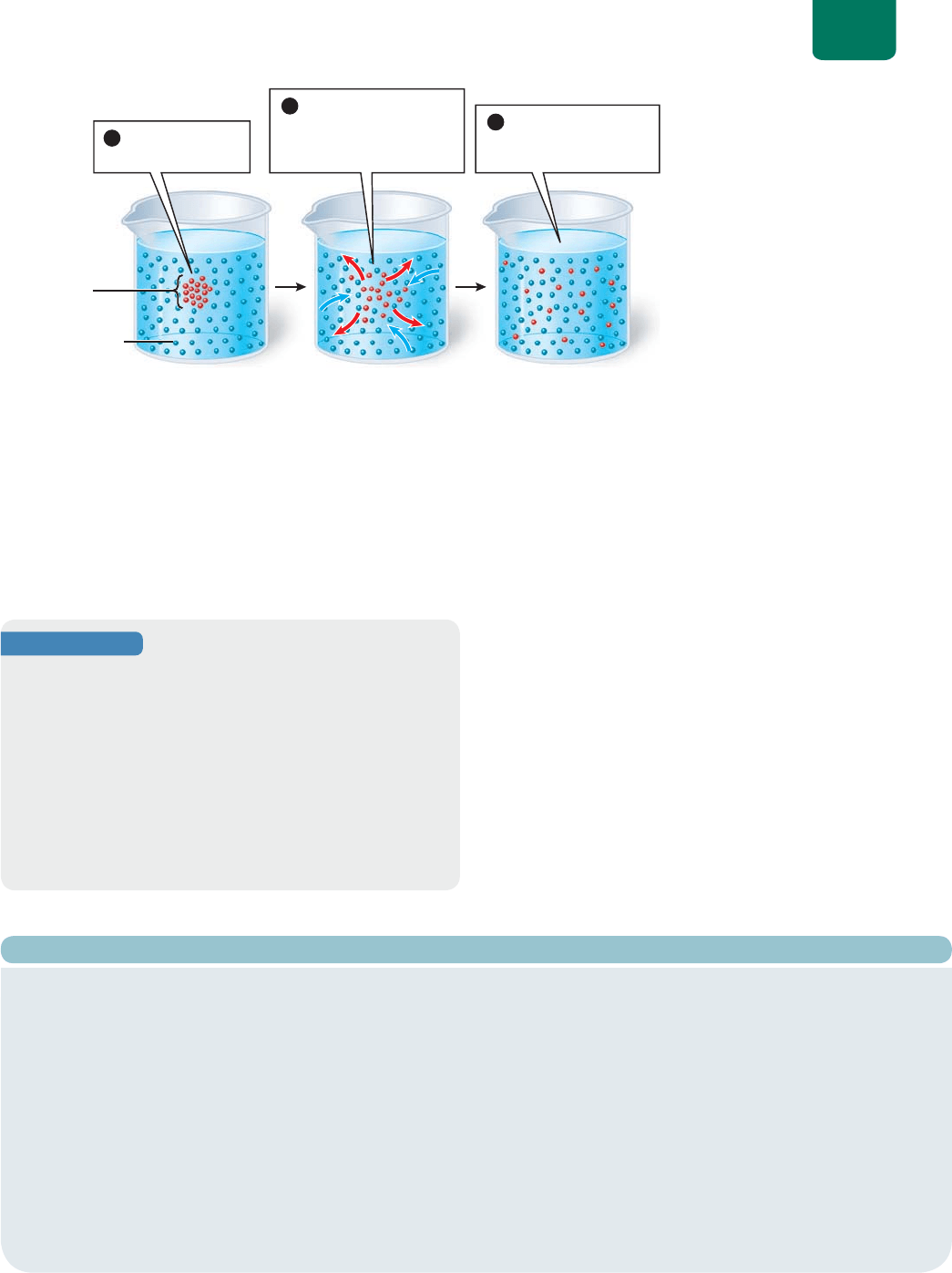
Para observar la difusión en acción, pongamos una gota
de colorante vegetal en una jarra de vidrio (
FIGURA 5-6). El mo-
vimiento aleatorio impulsa a las moléculas del tinte a entrar y
salir de la gota de color, pero hay una transferencia neta de tinte
al agua y del agua al tinte. El movimiento neto del tinte conti-
nuará hasta que esté disperso uniformemente en el líquido. Si se
compara la difusión del tinte en agua caliente y fría, se ve que el
calor aumenta la velocidad de la difusión, lo que se debe a que
el calor acelera el movimiento de las moléculas.
En resumen Los principios de la difusión
• La difusión es el movimiento neto de moléculas de
un gradiente de mayor a menor concentración.
• Cuanto mayor es el gradiente de concentración, la
velocidad de la difusión es mayor.
• Cuanto más alta es la temperatura, la velocidad de la
difusión es mayor.
• Si no intervienen otros mecanismos, la difusión
continúa hasta que las concentraciones se igualan
en todas sus partes, es decir, hasta que se pierde el
gradiente de concentración.
El movimiento a través de las membranas
ocurre por transporte pasivo y activo
Los gradientes de iones y moléculas por la membrana celular son
cruciales para la vida; una célula sin gradientes está muerta. Las
proteínas de las membranas celulares gastan energía para crear y
mante ner estos gradientes de concentración porque muchos de los
procesos bioquímicos de la vida dependen de ellos. Por ejemplo,
las neuronas dependen del flujo de iones específicos por su gradien-
te de concentración para producir las señales eléctricas que fundan
la sensación y el movimiento. Se dice que las membranas plasmá-
ticas son selectivamente permeables porque sólo permiten que
pasen (que permeen) ciertos iones o moléculas. La permeabilidad
selectiva de la membrana plasmática crea una barrera que ayuda a
mantener los gradientes que caracterizan a todas las células.
En su función de guardiana de la célula, la membrana plas-
mática favorece el movimiento de las sustancias de dos formas:
transporte pasivo y transporte que requiere de energía (
Tabla 5-1).
El transporte pasivo consiste en la difusión de las sustancias a
través la membrana celular de un medio de mayor a uno de me-
nor concentración, mientras que en el transporte que requiere
energía, la célula consume energía para que entren y salgan las
sustancias.
Se pone una gota
de tinte en agua
1
Las moléculas del tinte
se difunden en el agua; las
moléculas de agua se
difunden en el tinte
2
Las moléculas del
tinte y del agua se
dispersan uniformemente
3
gota de tinte
molécula de agua
FIGURA 5-6 Difusión de un
tinte en agua
Tabla 5-1 Transporte por la membrana
Transporte pasivo Difusión de las sustancias a través de una membrana, por un gradiente de concentración, presión o carga
eléctrica; no requiere energía celular.
Difusión simple Difusión del agua, gases disueltos o moléculas solubles en lípidos por la bicapa lipídica de una membrana.
Difusión facilitada Difusión de agua, iones o moléculas solubles en agua por una membrana, a través de una proteína de canal
o portadora.
Ósmosis Difusión de agua a través de una membrana selectivamente permeable de una región de mayor a una de
menor concentración de agua libre.
Transporte que requiere energía Entrada y salida de las sustancias de una célula, con la energía suministrada por el ATP.
Transporte activo Movimiento de pequeñas moléculas o iones individuales contra sus gradientes de concentración, por medio
de proteínas que atraviesan la membrana.
Endocitosis Entrada de partículas o moléculas grandes a una célula; ocurre cuando la membrana plasmática envuelve la
sustancia en un saco membranoso que se introduce en el citoplasma.
Exocitosis Salida de partículas o moléculas grandes a una célula; ocurre cuando la membrana plasmática envuelve el
material en un saco membranoso que se desplaza a la superficie de la célula, se fusiona con la membrana
y se abre hacia el exterior expulsando su contenido y permitiendo que se difunda en el medio extracelular.
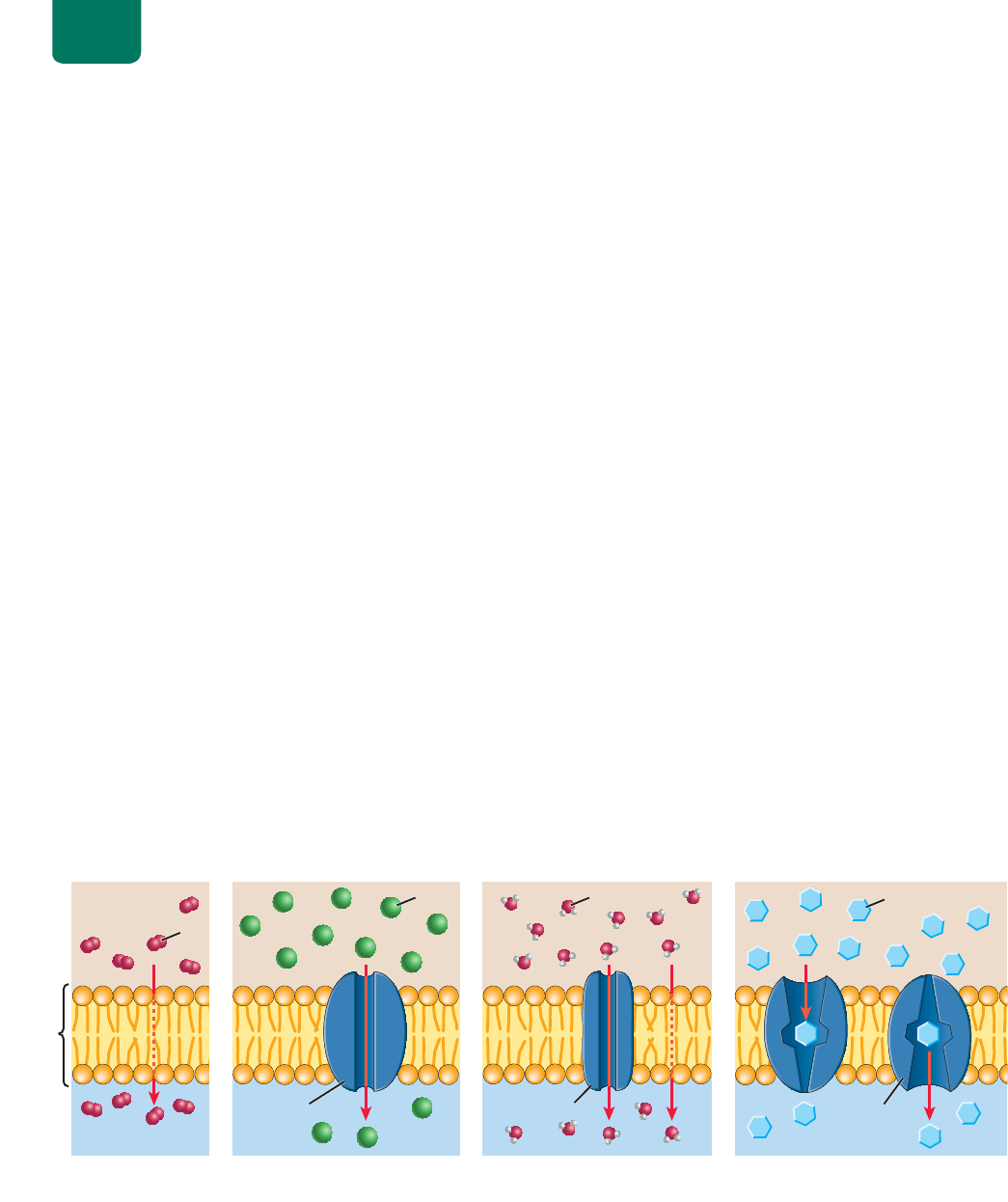
84 UNIDAD 1 La vida de la célula
agua
glucosa
proteína
portadora
acuaporina
proteína
de canal
bicapa
lipídica
JP[VWSHZTH
MS\PKV
L_[YHJLS\SHY
Cl
⫺
O
2
H+PM\ZP}UZPTWSLWVY
SHIPJHWHSPWxKPJH
I+PM\ZP}UMHJPSP[HKHWVY
JHUHSLZKLWYV[LxUH
JÔZTVZPZWVYHJ\HWVYPUHZ
VWVYSHIPJHWHSPWxKPJH
K+PM\ZP}UMHJPSP[HKHWVYWYV[LxUHZ
WVY[HKVYHZ
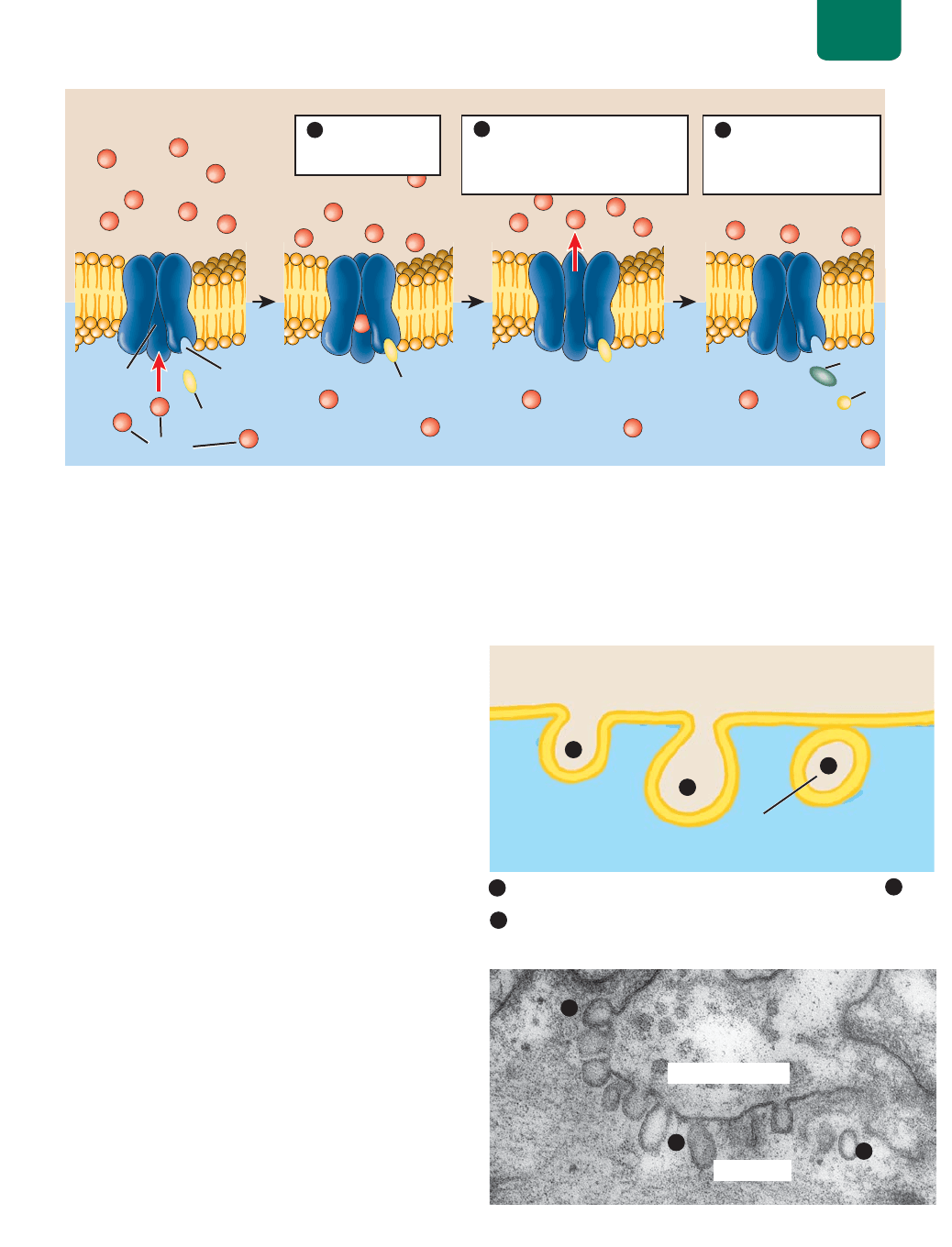
FIGURA 5-7 Tipos de difusión por la membrana plasmática (a) Las moléculas que son muy
pequeñas, que no cambian o que son liposolubles atraviesan directamente la bicapa de fosfolípidos por
difusión simple. Aquí, las moléculas de oxígeno se difunden del líquido extracelular a la célula por su
gradiente de concentración (flecha roja). (b) Por difusión facilitada en el canal de proteínas, los iones cruzan
las membranas. Aquí, iones de cloro descienden por su gradiente de concentración a la célula a través de
canales de cloro. (c) La ósmosis es la difusión de agua. Las moléculas de agua cruzan la bicapa lipídica por
difusión simple o pasan más rápidamente en la difusión facilitada por canales de agua llamados acuaporinas.
(d) Las proteínas portadoras tienen sitios de enlace para moléculas específicas (como la de glucosa que se
muestran aquí). Al enlazarse con la molécula transportada, la proteína portadora cambia de forma y lanza la
molécula a través de la membrana, por su gradiente de concentración.
El transporte pasivo es por difusión simple,
difusión facilitada y ósmosis
La difusión se produce en un fluido o a través de una membrana
que es permeable a la sustancia y que separa dos compartimentos
del fluido. Muchas moléculas cruzan la membrana plasmática por
difusión, impulsadas por diferencias de concentración entre el cito-
plasma y el fluido extracelular.
Algunas moléculas atraviesan las membranas
por difusión simple
Las moléculas muy pequeñas sin carga neta, como las de agua,
oxígeno y dióxido de carbono (lo mismo que moléculas liposolu-
bles, como el alcohol, las vitaminas A, D y E, y las hormonas este-
roides) se difunden a través de la bicapa de fosfolípidos por sus
gradientes de concentración. Este proceso se llama difusión sim-
ple (
FIGURA 5-7a). Un gradiente de concentración mayor, tempe-
ratura elevada, tamaño molecular pequeño y mayor solubilidad en
lípidos aumentan la velocidad de la difusión simple.
¿Cómo se difunde el agua (una molécula polar) a través de
la bicapa lipídica que es hidrofóbica (literalmente, “que teme al
agua”)? Como las moléculas de agua son tan pequeñas y abun-
dantes en el citoplasma y el fluido extracelular, algunas se pier-
den entre la abundancia de colas de los fosfolípidos, donde sus
movimientos aleatorios las llevan al otro lado de la membrana.
La difusión simple del agua por la bicapa lipídica es relativamen-
te lenta, pero en muchos tipos de células, las moléculas de agua
atraviesan la membrana mucho más deprisa gracias a la difusión
facilitada, como se verá en la siguiente sección.
Algunas moléculas atraviesan las membranas por difusión
facilitada, mediante proteínas de transporte
Muchas sustancias no pueden difundirse solas por la bicapa lipídi-
ca. Entre éstas se encuentran los iones (como K
, Na
, Cl
y Ca
2
),
que forman enlaces de hidrógeno con las moléculas de agua. Es-
tas moléculas retienen a los iones, ya sea dentro o fuera de la célula.
Moléculas como los monosacáridos (azúcares simples) son polares
y también forman enlaces de hidrógeno con el agua; así su atrac-
ción al agua y su tamaño inhibe su paso a la bicapa. Estos iones y
moléculas sólo pueden difundirse a través de las membranas con
la ayuda de proteínas de transporte específicas: proteínas de canal
o proteínas portadoras. El movimiento a través la membrana celu-
lar por gradiente de concentración y con la ayuda de proteínas de
transporte se llama difusión facilitada.
Las proteínas de canal forman poros (canales) en la bicapa
lipídica por la que ciertos iones o agua pueden descender por su
gradiente de concentración (
FIGURA 5-7b). Las proteínas de canal
tienen un diámetro interior relacionado con el diámetro del ión
cuyo movimiento facilitan y cargas eléctricas que atraen el ión de
los aminoácidos que revisten el poro. Por ejemplo, el canal para el
Na
está revestido con cargas negativas para atraer Na
. Como las
células deben mantener gradientes de muchos iones a través de sus
membranas, numerosos canales de iones tienen “puertas” de pro-
teínas que se abren o cierran, dependiendo de las necesidades de
la célula.
Muchas células tienen proteínas de canal especiales para
agua llamadas acuaporinas (literalmente, “de poros de agua”;
FIGURA 5-7c). El pequeño tamaño de estos canales, aunado a la
Estructura y función de la membrana celular Capítulo 5 85
carga positiva de los aminoácidos (que atraen el polo negativo de
las moléculas de agua dentro del poro), los hacen selectivos de las
moléculas de agua. El movimiento del agua en una membrana a
través de las acuaporinas, por difusión simple o facilitada, tiene
un nombre especial: ósmosis. Para saber más sobre los canales de
agua, véase la sección “Investigación científica: El descubrimiento
de las acuaporinas” de la página 86).
Las proteínas portadoras tienen sitios activos que enlazan
moléculas específicas del citosol o del extracelular, como ciertos
azúcares o proteínas pequeñas. El enlace produce un cambio en
la forma de la proteína de carga que permite pasar a las molécu-
las y cruzar la membrana. Estas proteínas no usan energía de la
célula y sólo pueden transferir moléculas a favor de su gradiente
de concentración (
FIGURA 5-7d).
La ósmosis es la difusión de agua por las membranas
selectivamente permeables
La ósmosis es el movimiento de agua a través de una membrana
selectivamente permeable en respuesta a gradientes de concentra-
ción, presión o temperatura. Aquí vamos a centrarnos en la ósmo-
sis de una región de mayor concentración de agua a una de menor
concentración. La ósmosis puede ocurrir directamente a través de
la bicapa de fosfolípidos o (más deprisa) por canales de acuapori-
na compuestos de proteínas que se extienden sobre la membrana.
¿Qué significa que una solución tiene una “concentración
elevada de agua” o una “concentración baja de agua”? La res-
puesta es sencilla: el agua pura tiene la mayor concentración po-
sible de agua. Toda sustancia que se disuelve en el agua (todo so-
luto) desplaza algunas moléculas de agua en un volumen dado
y también forma enlaces de hidrógeno con muchas otras molé-
culas de agua, con lo que impide que pasen por una membrana
permeable al agua. Por tanto, cuanto mayor es la concentración
de sustancias disueltas, menor es la concentración de agua dis-
ponible para atravesar la membrana. Por consiguiente, habrá
un movimiento neto de la solución con más moléculas de agua
libres (con menos soluto) a la solución con menos moléculas li-
bres (con más soluto). Por ejemplo, el agua se mueve por ósmo-
sis de una solución con menos azúcar disuelta a una solución
con más azúcar disuelta. Como hay más moléculas de agua libres
en la solución con azúcar menos concentrada, más moléculas
chocan y atraviesan la membrana permeable al agua de ese lado.
La concentración del soluto en el agua determina la “fuerza os-
mótica”; cuanto mayor es la concentración del soluto, mayor es
la fuerza osmótica.
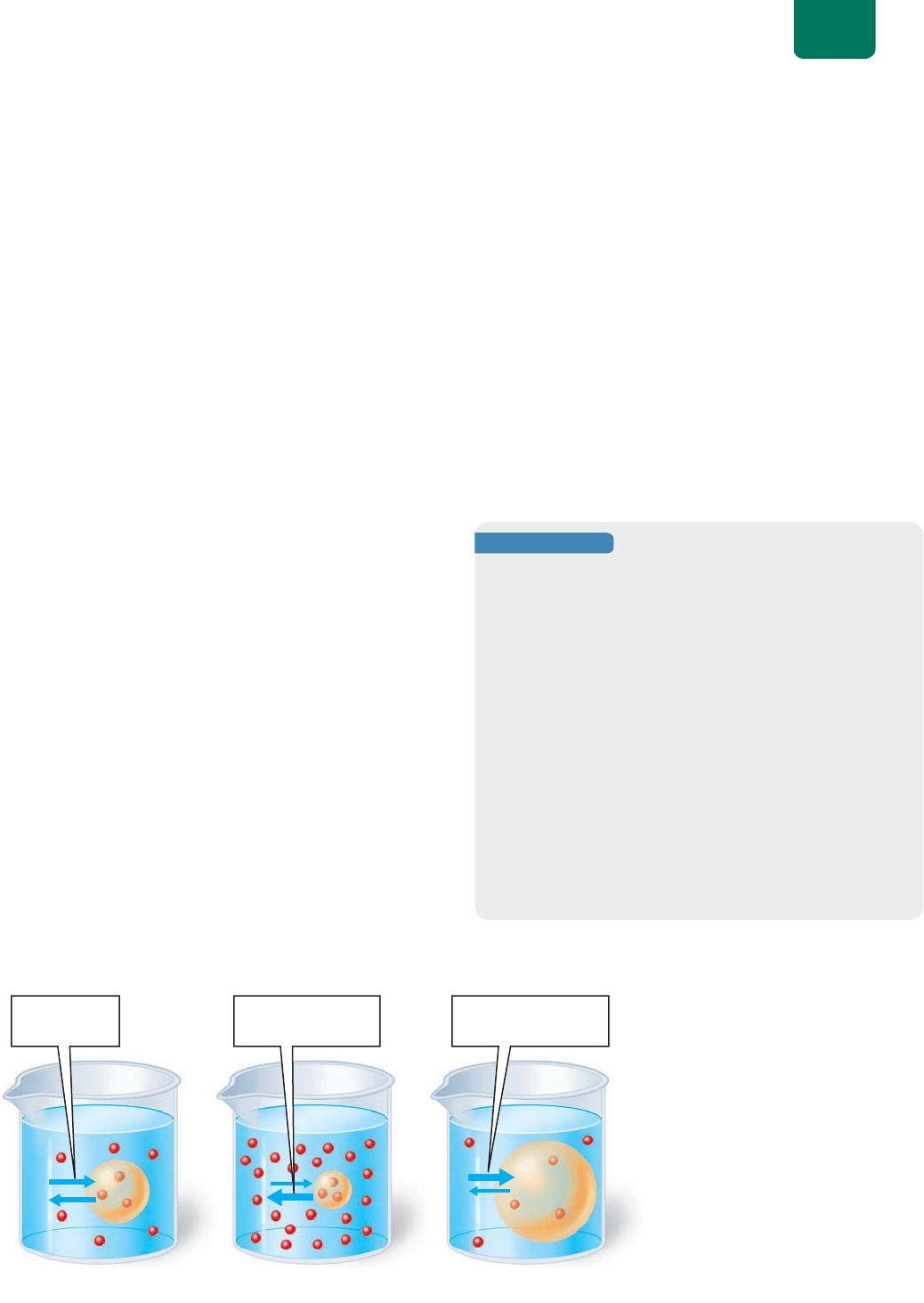
Los científicos usan la palabra “tonicidad” para comparar
las concentraciones de sustancias disueltas en el agua a través de
una membrana que es selectivamente permeable al agua. Se dice
que las soluciones con concentraciones iguales de un soluto (y,
por tanto, con concentraciones iguales de agua) son isotónicas
una de la otra (el prefijo iso- significa “igual”). Cuando las solu-
ciones isotónicas están separadas por una membrana permeable
al agua, no hay movimiento neto de agua entre ellas (
FIGURA
5-8a
). Cuando una membrana selectivamente permeable al agua
separa soluciones con diferentes concentraciones de soluto, la
solución que contiene una mayor concentración del soluto es
hipertónica (el prefijo hiper- significa “mayor que”) respecto de
la solución menos concentrada (
FIGURA 5-8b). La solución más
diluida se llama hipotónica (hipo- significa “debajo de”;
FIGU-
RA 5-8c
). El agua se mueve de las soluciones hipotónicas a las
hipertónicas.
En resumen Los principios de la ósmosis
• La ósmosis es el movimiento de agua por difusión
a través de una membrana selectivamente permea-
ble al agua simple o por difusión facilitada por
acuaporinas.
•
El agua cruza una membrana selectivamente permea
-
ble al agua por su gradiente de concentración, del
lado con la mayor concentración de moléculas de
agua libre al lado con menor concentración.
• Las sustancias disueltas reducen la concentración de
agua libre de las moléculas de una solución.
• Si dos soluciones están separadas por una membrana
que es selectivamente permeable al agua, la solución
con la mayor concentración de soluto es hipertóni-
ca y tiene mayor fuerza osmótica, mientras que la
solución con la menor concentración del soluto es
hipotónica y tiene menor fuerza osmótica.
Sin flujo
neto de agua
El agua sale;
el globo se encoge
El agua entra;
el globo se hincha
H.SVIVLU\UH
ZVS\JP}UPZV[}UPJH
I.SVIVLU\UH
ZVS\JP}UOPWLY[}UPJH
J.SVIVLU\UH
ZVS\JP}UOPWV[}UPJH
FIGURA 5-8 Efecto de la
concentración del soluto en la
ósmosis
Comenzamos con tres
globos hechos de una membrana que
es selectivamente permeable al agua
(pero no al azúcar; esferas rojas).
Colocamos volúmenes iguales de la
misma concentración de agua azucarada
en cada globo. Cuando sumergimos los
globos en un vaso de precipitados lleno
con diferentes concentraciones de agua
azucarada —(a) isotónica, (b) hipertónica
o (c) hipotónica— que en los globos. En
la figura se muestra el resultado al cabo
de una hora. Las flechas azules indican la
entrada y salida del agua por el globo.

86 UNIDAD 1 La vida de la célula
Investigación científica
El descubrimiento de las acuaporinas
El comentario del microbiólogo francés Louis Pasteur de que “la
suerte favorece a la mente preparada” es tan válido hoy como
cuando lo pronunció en la década de 1880. Desde hace mucho,
los científicos saben que la ósmosis directa por la bicapa de
fosfolípidos es demasiado lenta para explicar el movimiento del
agua por ciertas membranas celulares, como los túbulos renales
(que reabsorben cantidades enormes de agua que el riñón filtra
de la sangre) y los glóbulos rojos (véase la figura 5-9). Pero los
intentos por identificar proteínas de transporte selectivo del
agua fracasaron repetidamente, por una parte porque el agua
es abundante a ambos lados de la membrana plasmática y, por
otra, porque el agua también puede pasar de forma directa por la
bicapa lipídica.
Entonces, como suele ocurrir en las ciencias, se encontraron
la suerte y una mente preparada. A mediados de la década de
1980, el doctor Peter Agre (FIGURA E5-2), que por ese tiempo
trabajaba en la Johns Hopkins School of Medicine (Escuela de
Medicina Johns Hopkins) de Maryland, trataba de determinar
la estructura de una glucoproteína de los glóbulos rojos. La
proteína que aisló estaba contaminada con grandes cantidades
de otra proteína. En lugar de desechar la proteína desconocida,
Agre y sus colaboradores la investigaron, para lo cual hicieron
que huevos de rana (que son significativamente permeables
al agua) insertaran la proteína en su membrana plasmática.
Mientras que los huevos sin la proteína misteriosa se hinchaban
poco cuando se colocaban en una solución hipotónica, los
huevos con la proteína se hinchaban muy rápido y reventaban en
la misma solución (FIGURA E5-3a). En nuevos estudios se mostró
que por este canal no pasaban otros iones ni moléculas, así
que fue llamado acuaporina. En 2000, Agre y otros equipos de
investigadores hicieron pública la estructura tridimensional de la
acuaporina y describieron cómo aminoácidos polares específicos
del interior atraen el agua y permiten el paso por el canal de
miles de millones de moléculas de agua, una tras otra, en un
segundo, al tiempo que repelen otros iones y moléculas (FIGURA
E5-3b
).
Ahora ya se han identificado muchos tipos de acuaporinas,
pues se encuentran en todas las formas de vida que se han
investigado. Por ejemplo, la membrana plasmática de la vacuola
central de las células vegetales tiene numerosas acuaporinas
(véase la figura 5-10). En 2003, Agre compartió el premio
Nobel de Química por su descubrimiento: el resultado de la
oportunidad, observación atenta, persistencia y, quizá, un poco
de lo que Agre llama, con modestia, “pura buena suerte”.
FIGURA E5-2 Peter Agre
(fluido extracelular)
canal de
acuaporina
(a) Huevos de rana
huevo sin
acuaporinas
huevo con
acuaporinas
agua
agua
(citoplasma)
(b) canal de acuaporina
FIGURA E5-3 Función y estructura de las acuaporinas
(a) El huevo de rana de la derecha tiene acuaporinas en la
membrana plasmática; el huevo de la izquierda, no. Los dos
fueron sumergidos en una solución hipotónica durante 30
segundos. El huevo de la derecha reventó, mientras que el de la
izquierda sólo se hinchó levemente. (b) El canal de acuaporina
(mostrado en corte transversal) está formado por una proteína
que abarca la membrana. Dentro del canal, aminoácidos
cargados interactúan con las moléculas de agua y favorecen su
movimiento en cualquier dirección, al tiempo que repelen otras
sustancias.

Estructura y función de la membrana celular Capítulo 5 87
La ósmosis por la membrana plasmática cumple
una función importante en la vida de las células
Normalmente, el fluido extracelular de los animales es isotónico
con el líquido citoplasmático de sus células, así que no hay nin-
guna tendencia a que el agua salga o entre. Aunque las concentra-
ciones de solutos específicos casi nunca son los mismos dentro y
fuera de las células, la concentración total de todas las partículas
disueltas es igual; por tanto, la concentración de agua es igual den-
tro y fuera de las células.
Si se sumergen glóbulos rojos en soluciones salinas de di-
versas concentraciones de soluto, se observan los efectos de la
entrada y salida de agua de las membranas celulares. En una so-
lución salina isotónica, el tamaño de los glóbulos se mantiene
constante (
FIGURA 5-9a). Si la solución salina es hipertónica con
relación al citosol de los glóbulos, el agua sale por ósmosis y éstos
se encogen (
FIGURA 5-9b). Por el contrario, si la solución de sal
está muy diluida y es hipotónica en relación con el citosol de los
glóbulos rojos, el agua entra en éstos y los hincha (
FIGURA 5-9c).
Si los glóbulos rojos están colocados en agua pura (solución hi-
potónica), se hinchan hasta reventar.
La ósmosis a través de la membrana plasmática es crucial
para entender muchos procesos biológicos, incluyendo la capta-
ción de agua por las raíces de las plantas, la absorción en el in-
testino del agua ingerida y la reabsorción de agua en los riñones.
Los organismos que viven en agua dulce deben consumir ener-
gía para contrarrestar la ósmosis. Protistas como los paramecios
tienen vacuolas contráctiles que eliminan el agua que entra de
forma continua al citosol, el cual es hipertónico con relación al
agua dulce de los estanques en que viven. Los paramecios aprove-
chan la energía celular para bombear sales del citosol a la vacuola
contráctil. Así pasa el agua por ósmosis y llena la vacuola, que
entonces se contrae y lanza el agua por un poro de la membrana
(véase la figura 4-15).
Casi toda célula vegetal viva está sostenida por el agua que
entra por ósmosis. Como se vio en el capítulo 4, la mayor par-
te de las células vegetales tienen una membrana grande que en-
globa la vacuola central y que tiene abundantes acuaporinas. Las
sustancias disueltas guardadas en la vacuola hacen que el conte-
nido sea hipertónico respecto del citosol celular, el cual, por su
parte, también es hipertónico con relación al fluido extracelular
que baña las células. Por tanto, el agua pasa al citosol y luego a la
vacuola por ósmosis. La presión del agua en la vacuola, llamada
presión de turgencia, empuja al citoplasma contra la pared ce-
lular con fuerza considerable (
FIGURA 5-10a).
Las paredes celulares son flexibles, así que las células ve-
getales dependen de la turgencia para sostenerse. Cuando no se
riega una planta de interiores, la vacuola central y el citosol de
las células pierde agua y la membrana plasmática se encoge y se
aleja de la pared celular. Así como una pelota se desinfla cuando
se queda sin aire, la planta se desploma si sus células pierden
la presión de turgencia (
FIGURA 5-10b). Ahora ya sabes por qué
en la sección de frutas y verduras de tiendas y mercados siempre
rocían con agua los productos: para que se vean frescos y en buen
estado, con vacuolas centrales llenas.
El transporte que requiere energía es transporte
activo, endocitosis y exocitosis
Sin gradientes de concentración en las membranas, una célula
muere. Al formar gradientes y, en ciertas circunstancias, dejar que
desaparezcan, las células regulan sus reacciones bioquímicas, res-
H.S}I\SVZYVQVZLU\UH
ZVS\JP}UPZV[}UPJH
I.S}I\SVZYVQVZLU\UH
ZVS\JP}UOPWLY[}UPJH
J.S}I\SVZYVQVZLU\UH
ZVS\JP}UOP
W
V[}UPJH
FIGURA 5-9 Efectos de la ósmosis en los glóbulos rojos (a) Las células se sumergen en una
solución isotónica y conservan su forma normal con un hoyuelo. (b) Las células se encogen cuando se
colocan en una solución hipertónica, pues sale más agua de la que entra. (c) Las células se hinchan cuando
se colocan en una solución hipotónica.
PREGUNTA Una estudiante vierte agua destilada en una muestra de sangre. Al volver más tarde, observa la
sangre al microscopio y no ve ningún glóbulo rojo. ¿Qué ocurrió?

88 UNIDAD 1 La vida de la célula
&XDQGRKD\SRFDDJXDOD
YDFXRODFHQWUDOVHHQFRJH\OD
SDUHGFHOXODUSLHUGHVRSRUWH
6LQHOVRSRUWHGHODJXD
ODSODQWDVHPDUFKLWD
FLWRSODVPD
H3H[\YNLUJPHWYVWVYJPVUHZVWVY[L
YDFXRODFHQWUDO
SDUHGFHOXODU PHPEUDQDSODVPÀWLFD
6LKD\DEXQGDQWHDJXDOOHQD
ODYDFXRODFHQWUDOSUHVLRQD
HOFLWRSODVPDFRQWUDODSDUHG
FHOXODU\PDQWLHQHODIRUPD
GHODFÒOXOD
/DSUHVLʼnQGHO
DJXDVRVWLHQH
ODVKRMDVGHHVWD
SODQWDEDOVDPLQD
I3HWtYKPKHKL[\YNLUJPHTHYJOP[HSHWSHU[H
FIGURA 5-10 Turgencia en células vegetales Las
acuaporinas permiten al agua entrar y salir con rapidez
de las vacuolas centrales en células vegetales. La célula
y planta de la parte (a) están sostenidas por la turgencia
del agua, mientras que en la parte (b) perdieron esa
presión por deshidratación.
PREGUNTA Si una célula vegetal se pone en agua sin
solutos, ¿acabará por reventar? ¿Y una célula animal?
Explica.
ponden a los estímulos externos y obtienen energía química. Las
señales eléctricas de las neuronas, la contracción de los músculos
y la generación de ATP en mitocondrias y cloroplastos (véanse los
capítulos 7 y 8) dependen de los gradientes de concentración de
los iones. Pero los gradientes no se forman de manera espontánea,
sino que requieren el transporte activo a través de la membrana.
El transporte activo utiliza energía para mover moléculas
contra su gradiente de concentración
En el transporte activo, las proteínas de membrana utilizan ener-
gía celular para mover moléculas o iones por la membrana celular,
en contra de su gradiente de concentración (
FIGURA 5-11). Todas
las células necesitan trasladar materiales “corriente arriba”, en con-
tra de su gradiente de concentración. Por ejemplo, toda célula re-
quiere nutrimentos que están menos concentrados en el entorno
que en el citoplasma de la célula. Otras sustancias, como los iones
de sodio y calcio, se mantienen en concentraciones mucho me-
nores dentro de la célula que en el fluido extracelular. Las células
nerviosas mantienen gradientes de concentración de iones grandes
porque sus señales eléctricas requieren un flujo rápido y pasivo de
los iones cuando los canales se abren. Cuando estos iones se difun-
den dentro (o fuera) de la célula, sus gradientes de concentración
deben reconstituirse por transporte activo.
Las proteínas de transporte activo abarcan todo el ancho
de la membrana y tienen dos sitios activos. Uno (que puede estar
hacia dentro o hacia fuera de la membrana plasmática, depen-
Estructura y función de la membrana celular Capítulo 5 89
diendo de la proteína) enlaza a una molécula o ión, como los
iones de calcio. Como se ve en la
FIGURA 5-11 ❶, el segundo
sitio, que siempre está dentro de la membrana, une a la molé-
cula transportadora de energía, que por lo regular es el adenosín
trifosfato (ATP; véase la página 51). El ATP cede energía a la pro-
teína, con lo que ésta cambia de forma y mueve el ión de calcio
por la membrana (
FIGURA 5-11 ❷). La energía para el transporte
activo viene de un enlace energético que une el último de los tres
grupos fosfato del ATP. Al liberar su energía guardada, el ATP se
convierte en ADP (adenosín difosfato) más un fosfato libre (
FI-
GURA 5-11
❸). Las proteínas del transporte activo se llaman a
veces bombas (por analogía con las bombas de agua), porque usan
energía para mover iones o moléculas en contra de un gradiente
de concentración.
La célula engloba partículas o líquidos por endocitosis
Una célula puede necesitar materiales de su entorno extracelu-
lar que son demasiado grandes para pasarlos por la membrana.
Entonces, la membrana plasmática los engloba en vesículas que
transporta dentro de la célula. Este proceso que requiere energía se
llama endocitosis (en griego significa “dentro de la célula”). Aquí
vamos a describir tres formas de endocitosis basadas en el tamaño
y tipo del material adquirido y del método de adquisición: pinoci-
tosis, endocitosis mediada por receptor y fagocitosis.
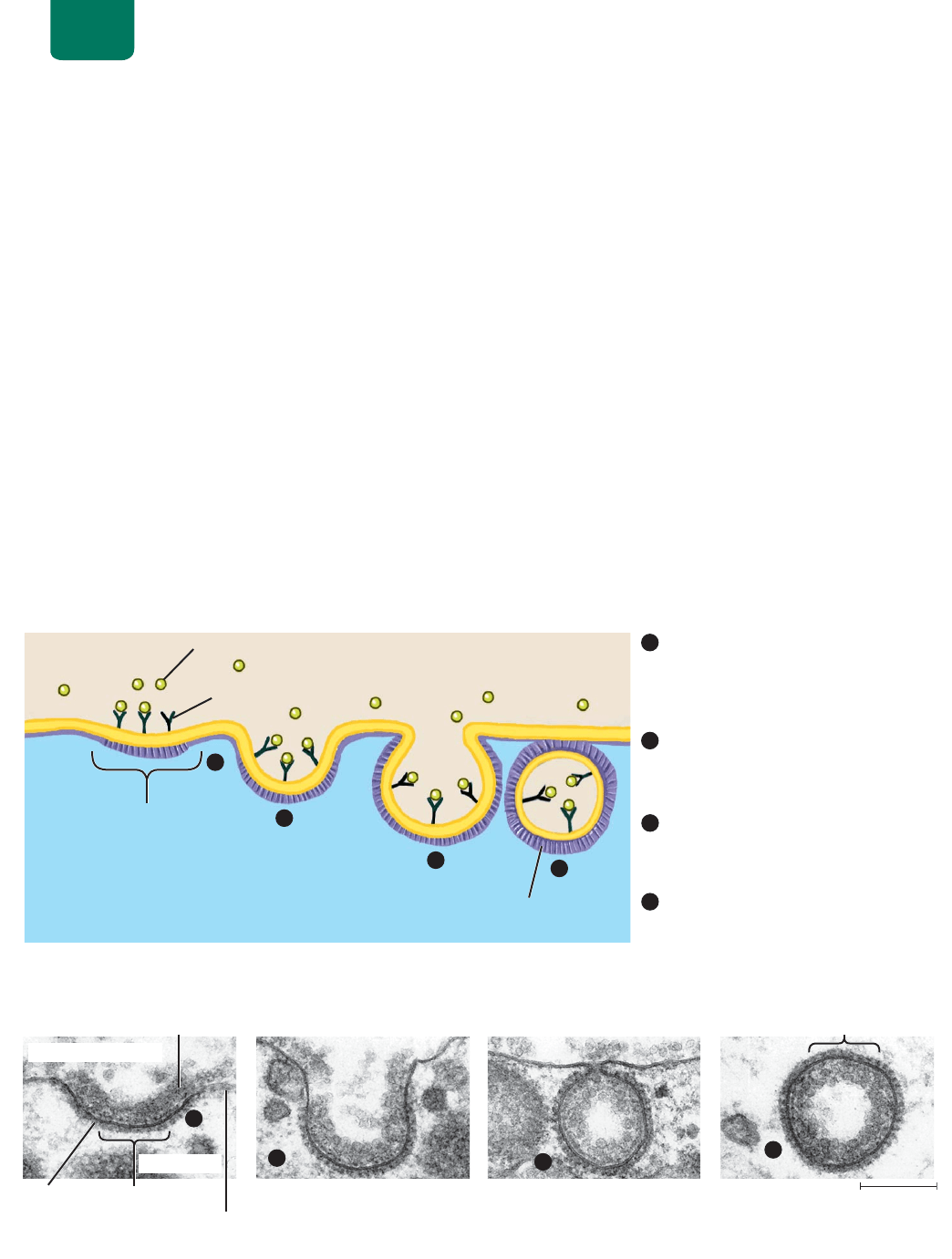
La pinocitosis lleva líquidos al interior de la célula En la pino-
citosis (“bebida de la célula”), un segmento muy pequeño de
la membrana plasmática sufre una depresión que se hace más
profunda conforme se llena de fluido extracelular y sigue hun-
diéndose hasta extrangularse dentro del citosol para formar una
vesícula diminuta que se desprende en el citoplasma (
FIGURA
5-12
). La pinocitosis introduce en la célula una gota de fluido ex-
/DSURWHķQDGH
WUDQVSRUWHXQH
HO$73\HO&D
⫹
/DHQHUJķDGHO$73
FDPELDODIRUPDGHODSURWHķQD
GHWUDQVSRUWH\KDFHSDVDU
HOLRQSRUODPHPEUDQD
/DSURWHķQDOLEHUD
HOLʼnQ\ORVUHPDQHQWHV
GHO$73$'3\3\
VHFLHUUD
VLWLRGH
HQODFH
GHO$73
VLWLRGH
UHFRQR
FLPLHQWR
$73
3
$'3
&D
⫹
fluido extracelular
citoplasma
$73
FIGURA 5-11 Transporte activo El transporte activo utiliza energía celular para mover moléculas a
través de la membrana plasmática y en contra del gradiente de concentración. Una proteína de transporte
(azul) tiene un sitio de enlace de ATP y un sitio de reconocimiento de las moléculas que se van a transportar;
en este caso, iones de calcio (Ca
2
). Observa que cuando el ATP dona su energía, pierde su tercer grupo
fosfato y se convierte en ADP P.
(uido extracelular)
(citoplasma)
YHVķFXODFRQIOXLGR
H[WUDFHOXODU
H7PUVJP[VZPZ
I0THNLUKL4,;KLSHWPUVJP[VZPZ
6HIRUPDXQDGHSUHVLʼnQHQODPHPEUDQDSODVPÀWLFDTXHVH
KDFHPÀVSURIXQGDVHHQVDQFKD\VHOOHQDGHIOXLGRH[WUDFHOXODU
/DPHPEUDQDHQJOREDDOIOXLGRH[WUDFHOXODUIRUPDQGRXQDYHVķFXOD
IOXLGRH[WUDFHOXODU
FLWRSODVPD
FIGURA 5-12 Pinocitosis Los números encerrados en
círculos corresponden a (a) el diagrama y (b) la micrografía de un
microscopio electrónico de transmisión.
90 UNIDAD 1 La vida de la célula
recubrimiento
de proteínas
fosa recubierta
vesícula recubierta
partículas extracelulares
unidas a receptores
membrana plasmática
(fluido extracelular)
(citoplasma)
0.1 micra
(citoplasma)
molécula nutriente
(fluido extracelular)
receptor
vesícula recubierta
fosa recubierta
(a) Endocitosis mediada por receptor
Las proteínas receptoras de moléculas
o complejos de moléculas específicas se
encuentran dentro de fosas recubiertas.
Se desprende en el citoplasma una
vesícula (“vesícula recubierta”) que
contiene las moléculas enlazadas.
La región de la fosa recubierta engloba
las moléculas unidas a los receptores.
Los receptores se unen a las
moléculas y la membrana se forma una
depresión.
1
2
3
4
1
2
3
4
4
3
2
1
(b) Imagen de TEM de endocitosis mediada por receptor
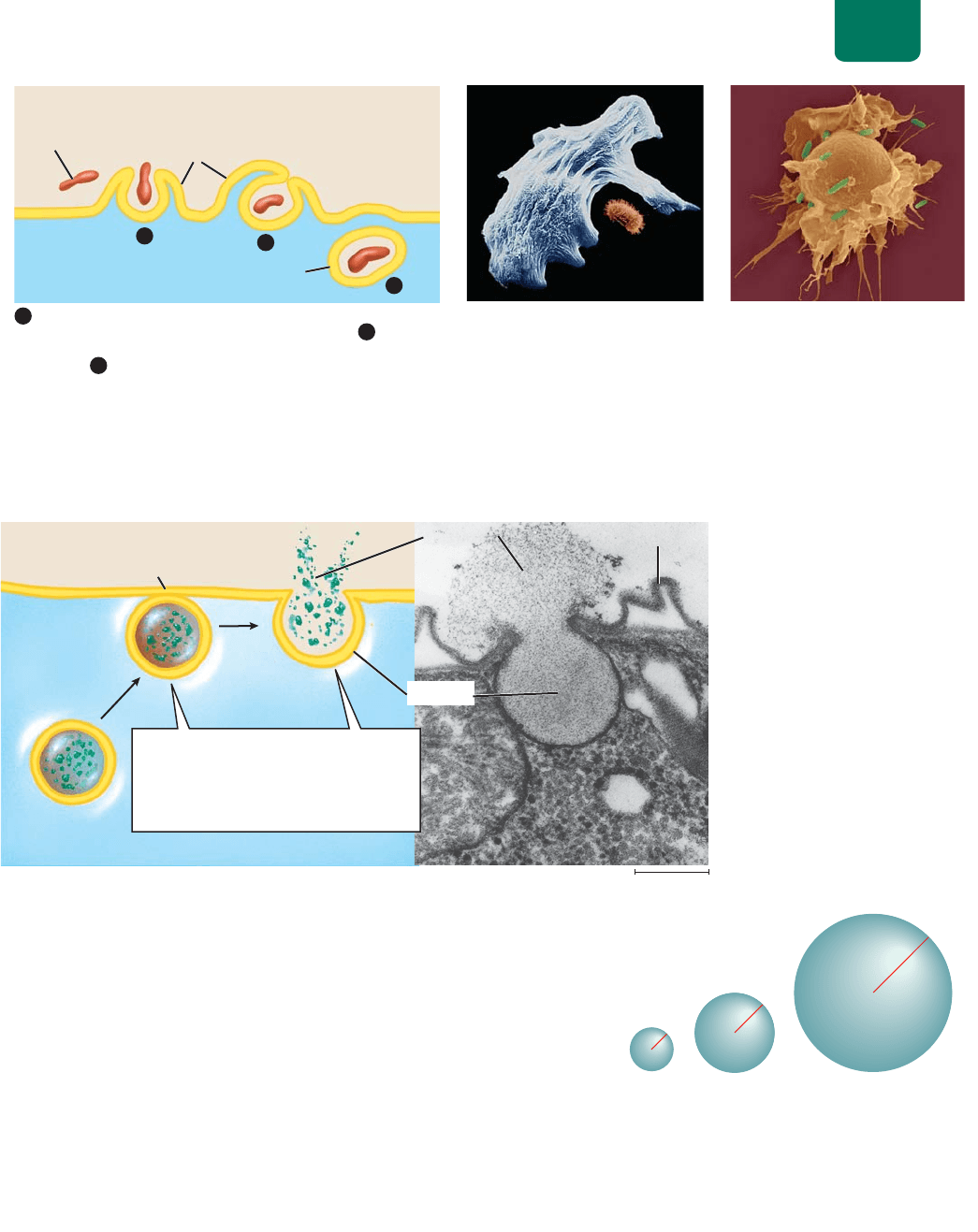
FIGURA 5-13 Endocitosis mediada por receptor Los números encerrados en círculos corresponden
a (a) el diagrama y (b) la micrografía de un microscopio electrónico de transmisión.
tracelular contenido en la depresión de la membrana. Por tanto,
la célula adquiere materiales en la misma concentración que en
el fluido extracelular.
La endocitosis mediada por receptores introduce moléculas específi-
cas al interior de la célula. Con el fin de concentrar selectivamente
materiales que no se mueven por los canales, las células toman
moléculas o complejos de moléculas específicas (por ejemplo, pa-
quetes que contengan lipoproteínas y colesterol) por medio de un
proceso llamado endocitosis mediada por receptor (
FIGURA
5-13
). Este proceso depende de las proteínas receptoras especiali-
zadas situadas en la membrana plasmática en depresiones hondas
llamadas fosas recubiertas. Cuando las moléculas correspondientes
se unen a estos receptores, la fosa recubierta se desprende en la
forma de una vesícula que lleva las moléculas al citoplasma.
La fagocitosis lleva partículas grandes al interior de la célula. Por
fagocitosis (que significa “acción de comer de la célula”) la célu-
la toma partículas grandes, incluyendo microorganismos (
FIGU-
RA 5-14
). Por ejemplo, cuando el protista de agua dulce Amoeba
detecta un suculento paramecio, extiende parte de su membrana
exterior. Estas extensiones se llaman seudópodos (término latino
que significa “pies falsos”). Los seudópodos se fusionan alrede-
dor de la presa, con lo que la engloban dentro de una vesícula
llamada vacuola alimentaria, para ser digerida (
FIGURA 5-14b).
Como la Amoeba, los leucocitos fagocitan y someten a digestión
intracelular a bacterias invasoras que engloban y destruyen, en un
drama que ocurre dentro de tu cuerpo (
FIGURA 5-14c).
La exocitosis expulsa material de la célula
Las células también usan energía para desechar las partículas que
no se digirieron o para secretar sustancias, como hormonas, al
fluido extracelular por medio de exocitosis (término griego que
significa “fuera de la célula”;
FIGURA 5-15). En la exocitosis, una
vesícula rodeada de membrana y que lleva el material que se va a
expulsar pasa a la superficie de la célula, donde la membrana de
la vesícula se fusiona con la membrana plasmática. La vesícula se
abre al líquido extracelular para que su contenido se difunda por el
fluido fuera de la célula.
El intercambio de materiales por las membranas
influye en el tamaño y la forma de la célula
Como vimos en el capítulo 4, casi todas las células son demasiado
pequeñas para ser detectadas a simple vista. Tienen un diámetro
Estructura y función de la membrana celular Capítulo 5 91
aproximado de 1 a 100 micras (millonésimas de metro; véase la
figura 4-1). ¿Por qué? Cuando una célula más o menos esférica cre-
ce, sus regiones internas van quedando más alejadas de la membra-
na plasmática, que es la que adquiere los nutrimentos de la célula
y elimina los productos de desecho. Buena parte del intercambio
ocurre por difusión lenta. En una hipotética célula gigante de 20
centímetros de diámetro, las moléculas de oxígeno tardarían 200
días en difundirse al centro de la célula, pero para entonces ésta ya
se habría muerto por falta de oxígeno. Además, conforme la esfera
crece, su volumen aumenta con mayor rapidez que su área superfi-
cial. Entonces, una célula grande y aproximadamente esférica (que
necesitaría más nutrimentos y produciría más desechos) tendría
una membrana de área relativamente menor para realizar este in-
tercambio que si fuera una célula esférica pequeña (
FIGURA 5-16).
En una célula muy grande y aproximadamente esférica, el
área superficial de la membrana plasmática sería demasiado pe-
queña y las distancias de la difusión excesivamente grandes para
cubrir las necesidades metabólicas de la célula. Estas restricciones
limitan el tamaño de las células; sin embargo, algunas se vuel-
membrana plasmática
membrana plasmática
(citoplasma)
vesícula
0.2 micra
material
excretado
(fluido extracelular)
El material se encierra en una vesícula
que se fusiona con la membrana
plasmática, se expulsa para que su
contenido se difunda en el fluido
extracelular
FIGURA 5-15 Exocitosis La
exocitosis es, por su función, lo
contrario de la endocitosis.
PREGUNTA ¿En qué difiere
la exocitosis de la difusión de
materiales al exterior de la célula?
SDUWķFXODGHDOLPHQWR
VHXGʼnSRGR
YDFXROD
DOLPHQWDULD
H-HNVJP[VZPZ
I<UH(TVLIHMHNVJP[H
\UWHYHTLJPV
J<USL\JVJP[VPUNPLYLIHJ[LYPHZ
MS\PKVL_[YHJLS\SHY
JP[VWSHZTH
/DPHPEUDQDSODVPÀWLFDSUR\HFWDVHXGʼnSRGRVKDFLD
XQDSDUWķFXODH[WUDFHOXODUSRUHMHPSORFRPLGD/RV
H[WUHPRVGHORVVHXGʼnSRGRVVHIXVLRQDQ\HQJOREDQOD
SDUWķFXOD6HIRUPDXQDYHVķFXODOODPDGDYDFXROD
DOLPHQWDULDTXHFRQWLHQHODSDUWķFXODHQJOREDGD
FIGURA 5-14 Fagocitosis (a) Mecanismo de la fagocitosis. Las fotografías de (b) una Amoeba y (c) un
leucocito fueron tomadas con un microscopio electrónico de barrido, que da imágenes tridimensionales. El
color es generado por computadora para distinguir las células.
distancia al
centro (r)
área superficial
(4 r
2
)
volumen
(4/3 r
3
)
π
π
área/volumen
1.0
2.0
4.0
12.6 50.3
4.2 33.5
3.0
1.5
201.1
268.1
0.75
r
r
r
FIGURA 5-16 Relaciones entre el área superficial y el
volumen A medida que aumenta el tamaño de la esfera, su volumen
se incrementa mucho más que su área superficial. Así, una célula
esférica grande tiene un área superficial relativamente menor por la
cual obtener nutrimentos y eliminar los desechos. Como resultado,
las células aproximadamente esféricas son muy pequeñas.
Este documento contiene más páginas...
Descargar Completo
Transmision sináptica215.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.