
Resumen Endocrino Pablo C Página 1
El sistema endocrino
Es el conjunto de órganos y tejidos del organismo, que segregan un tipo de sustancias llamadas hormonas, que son
liberadas al torrente sanguíneo y regulan algunas de las funciones del cuerpo.
Es un sistema de señales similar al del sistema nervioso, pero en este caso, en lugar de utilizar impulsos eléctricos a
distancia, funciona exclusivamente por medio de sustancias (señales químicas).
Actúa como una red de comunicación celular que responde a los estímulos liberando hormonas y es el encargado de
diversas funciones metabólicas del organismo. Los órganos endocrinos también se denominan glándulas sin
conducto o glándulas endocrinas, debido a que sus secreciones se liberan directamente en el torrente sanguíneo,
mientras que las glándulas exocrinas liberan sus secreciones sobre la superficie interna o externa de los tejidos
cutáneos, la mucosa del estómago o el revestimiento de los conductos pancreáticos.
Las hormonas secretadas por las glándulas endocrinas regulan el crecimiento, el desarrollo y las funciones de
muchos tejidos, y coordinan los procesos metabólicos del organismo.
Las glándulas más representativas del sistema endocrino son la hipófisis, la glándula tiroides y la suprarrenal. Las
glándulas endocrinas en general comparten características comunes como la carencia de conductos, alta irrigación
sanguínea y la presencia de vacuolas intracelulares que almacenan las hormonas. Esto contrasta con las glándulas
exocrinas como las salivales y las del tracto gastrointestinal que tienen escasa irrigación y poseen un conducto o
liberan las sustancias a una cavidad.
Hormonas
Las hormonas son sustancias químicas localizadas en las glándulas endocrinas. Básicamente funcionan como
mensajeros químicos que transportan información de una célula a otra. Por lo general son liberadas directamente
dentro del torrente sanguíneo, solas (biodisponibles) o asociadas a ciertas proteínas (que extienden su vida media) y
hacen su efecto en determinados órganos o tejidos a distancia de donde se sintetizaron.
Dependiendo de su secreción
Hormonas endocrinas, producidas por glándulas o por células especializadas que las secretan a la sangre
circulante y que influyen en la función de células diana situadas en otros lugares del organismo.
Hormonas neuroendocrinas, secretadas por las neuronas hacia la sangre y que influyen en las funciones de
células diana de otras partes del cuerpo.
Hormonas paracrinas, las células liberan sustancias químicas que se extienden a través del líquido
extracelular hasta otras células que se encuentran cerca.
Hormonas autocrinas, producidas por células y que pasan al líquido extracelular desde el que actúan sobre
las mismas células que las fabrican.
Dependiendo de su composición química
PROTEICAS Y
POLIPEPTIDICAS
ESTEROIDEAS
DERIVADOS DE AA TIROSINA
TIROIDEAS
CATECOLAMINAS
ORGANO
Adeno y
neurohipófisis,
Páncreas,
Paratiroides
Corteza
suprarrenal,
ovarios y
testículos, placenta
Glándula tiroides
Medula suprarrenal
SINTESIS
A partir de
aminoácidos en REG
A partir de
colesterol
A partir de tirosinas
combinadas y yodo
A partir de tirosinas
ALMACENAMIENTO
Vesículas en
citoplasma
No se almacenan
Prolongado en los
folículos de la glándula
Vesículas
preformadas
TRANSPORTE
Hidrosolubles, libres
en sangre
Liposolubles,
difunden por MB y
circulan unidas a
proteínas
transportadoras
Combinadas con
proteínas plasmáticas
Hidrosolubles, libres
en sangre
VIDA MEDIA
Corta
Prolongada
T4: 6 a 7 días
T3: 1 día
Corto (1 min aprox.)
RECEPTORES
En MB
Citoplasma
Núcleo celular
En MB
Este archivo fue descargado de https://filadd.com
FILADD.COM

Resumen Endocrino Pablo C Página 2
Las hormonas que se encuentran unidas a las proteínas plasmáticas se eliminan de la sangre con una velocidad
mucho menor y a veces permanecen en la circulación durante varias horas o incluso días.
Vía metabólica
Hormona
1-Sintesis. 2- Transporte. 3- Concentración. 4- Control de secreción. 5- Duración.
6- Receptores hormonales
Célula diana o célula blanco
Las células en donde las hormonas ejercen su efecto son capaces de reaccionar a éstas porque contienen receptores
específicos con los que éstas pueden unirse.
La retroalimentación negativa
Todas las hormonas están, en principio, muy controladas. En la mayoría de los casos, este control se ejerce a través
de mecanismos de retroalimentación negativa. Cuando un estímulo induce la liberación de una hormona, los estados
o los productos derivados de la acción de esta tienden a detener dicha liberación.
La retroalimentación positiva
Un ejemplo de esta retroalimentación positiva es el gran aumento de la síntesis de hormona luteinizante (lh) que se
produce como consecuencia del efecto estimulador ejercido por los estrógenos sobre la adenohipófisis antes de la
ovulación. La LH secretada actúa en los ovarios, donde estimula la síntesis de más estrógenos que, a su vez,
favorecen la secreción de LH.
Receptores
La acción de una hormona comienza con su unión a un receptor específico de la célula efectora. Las células que
carecen de receptores para una hormona no responden a ella.
Estos receptores son macromoléculas o asociaciones macromoleculares a las cuales la hormona se fija
selectivamente en virtud de una estrecha adaptación conformacional o complementariedad estructural. Al unirse la
hormona correspondiente, induce en ellos un cambio conformacional iniciándose los eventos determinantes del
efecto final. Los receptores hormonales se encuentran situados en la superficie de las células o en el interior de las
mismas. Todos o casi todos los receptores hormonales son proteínas, además, cada receptor suele ser específico
para una única hormona; ello determina qué hormona actuará sobre un tejido particular.
Los distintos tipos de receptores hormonales se encuentran
1. En o sobre la superficie de la membrana celular. Los receptores de membrana son específicos sobre todo de
las hormonas proteicas y peptídicas y de las catecolaminas.
2. En el citoplasma celular. Los receptores principales de las distintas hormonas esteroideas se encuentran
fundamentalmente en el citoplasma.
3. En el núcleo celular. Los receptores de las hormonas tiroideas se encuentran en el núcleo.
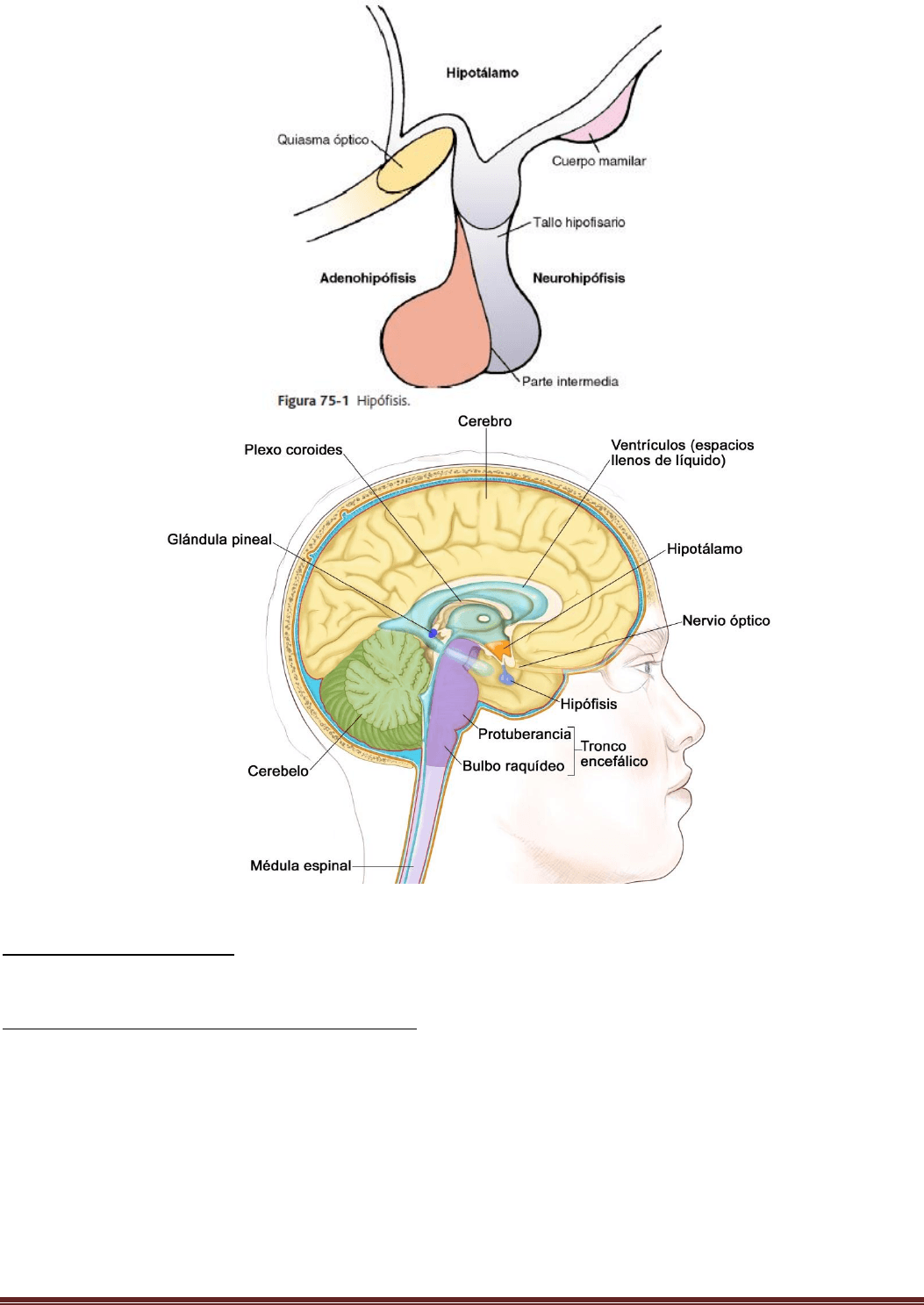
La hipófisis y su relación con el hipotálamo
La hipófisis (figura), denominada también glándula pituitaria, es una pequeña glándula de alrededor de 1 cm de
diámetro y 0,5-1 g de peso, situada en la silla turca (una cavidad ósea de la base del cráneo) y unida al hipotálamo
mediante el tallo hipofisario.
La hipófisis se divide en dos partes bien diferenciadas: el lóbulo anterior o adenohipófisis y el lóbulo posterior o
neurohipófisis. Entre estas dos partes existe una pequeña zona poco vascularizada y denominada parte intermedia,
mucho menos desarrollada en la especie humana y mucho más grande y funcional en algunos animales.
Este archivo fue descargado de https://filadd.com
FILADD.COM
Resumen Endocrino Pablo C Página 3
La adenohipófisis secreta seis hormonas peptídicas necesarias y otras de menor importancia:
• La hormona del crecimiento estimula el crecimiento de todo el cuerpo mediante su acción sobre la formación de
proteínas y sobre la multiplicación y diferenciación celulares.
Funciones fisiológicas de la hormona del crecimiento
Todas las hormonas adenohipofisarias más importantes, salvo la hormona del crecimiento, ejercen sus efectos
principalmente mediante la estimulación de las glándulas efectoras, sin embargo, la hormona del crecimiento no
actúa a través de ninguna glándula efectora específica, sino que ejerce un efecto directo sobre todos o casi todos los
tejidos del organismo.
Induce el crecimiento de casi todos los tejidos del organismo que conservan esa capacidad.
Favorece el aumento de tamaño de las células y estimula la mitosis, dando lugar a un número creciente de
células y a la diferenciación de determinados tipos celulares.
Favorece la movilización de los ácidos grasos libres en la sangre y potencia el uso de los ácidos grasos como
fuente de energía.
Disminuye la cantidad de glucosa utilizada en todo el organismo
Este archivo fue descargado de https://filadd.com
FILADD.COM
Resumen Endocrino Pablo C Página 4
Aumenta la traducción de ARN para facilitar la síntesis proteica en los ribosomas
Facilita el trasporte de AA a través de las membranas
Descenso del catabolismo de las proteínas y los AA.
Aumento de la trascripción nuclear del ADN para formar ARN (en períodos prolongados 24 a 48
horas)intensificará la síntesis de proteínas y el crecimiento (siempre que se disponga de una cantidad
suficiente de energía , AA, vitaminas)
Induce la liberación de los ácidos grasos del tejido adiposo aumenta su concentración en los líquidos
corporales.
Estimula la secreción de insulina
Estimula el crecimiento óseo y cartilaginoso
Aumentan longitudinalmente. Aumentan el grosor.
Aumenta la producción y la actividad de células osteogénicas y condriocitricas
• La corticotropina controla la secreción de algunas hormonas corticosuprarrenales, que, a su vez, afectan al
metabolismo de la glucosa, las proteínas y los lípidos.
Estructura
Proviene de la célula corticótropas
Cadena sencilla de 39 AA.
Estímulos:
Estrés físico o mental Provienen del hipotálamo
Dolores.
Funciones
Controla la secreción de algunas hormonas corticosuprarenales que a su vez afectan al metabolismo de la
glucosa las proteínas y los lípidos. Ejemplo cortisol, corticoesteroides, andrógenos y estrógenos.
• La tirotropina (hormona estimulante del tiroides) controla la secreción de tiroxina y triyodotironina por la glándula
tiroides; a su vez, estas hormonas regulan casi todas las reacciones químicas intracelulares que tienen lugar en el
organismo.
Estructura:
Provienen de la célula tirotropas
Glucoproteína: formada por dos subunidades α (89 AA) y β (112 AA)
Estímulos:
Exposición al frío
Reacciones emocionales
Funciones
Controla la secreción de tiroxina y triyodotrironina por la glándula tiroides, a su vez estas hormonas regulan casi
todas las reacciones químicas intracelulares que tienen lugar en el organismo.
• La prolactina estimula el desarrollo de las glándulas mamarias y la producción de leche.
Estructura:
Provienen de la célula Lactótropas y de la Mamótropas
Cadena única de 198 AA.
Estímulos:
Succión de la mama
Caricia de madre a hijo
Escuchar el llanto del bebé
Señales emocionales
Funciones:
Induce el crecimiento de la glándula mamaria
Estimula la secreción de leche
Este archivo fue descargado de https://filadd.com
FILADD.COM

Resumen Endocrino Pablo C Página 5
• Dos hormonas gonadótropas distintas, la hormona estimulante de los folículos y la hormona luteinizante, controlan
el crecimiento de los ovarios y los testículos, así como su actividad hormonal y reproductora. Las dos hormonas
secretadas por la neurohipófisis desempeñan otras funciones.
Estructura:
Provienen de la célula gonadótropas
LH: glucoproteína formada por dos subunidades α (89 AA) y β (115 AA)
FSH: glucoproteína formada por dos subunidades α (89 AA) y β (112 AA)
Estímulos:
Responden a la secreción clínica de GnRH.
Funciones:
LH:
1. Estimula la síntesis de testosterona (en células de Leydig)
2. Estimula la ovulación y al cuerpo lúteo.
3. Estimula la síntesis de estrógenos y progesterona
FSH
1. Estimula la maduración de espermatozoides (en células sertoli)
2. Induce el crecimiento de los folículos ováricos
• La hormona antidiurética (denominada también vasopresina) controla la excreción de agua en la orina, con lo que
ayuda a regular la concentración hídrica en los líquidos corporales.
• La oxitocina contribuye a la secreción de leche desde las glándulas mamarias hasta los pezones durante
posiblemente, interviene también en el parto, al final de la gestación.
La adenohipófisis contiene diversos tipos celulares que sintetizan y secretan hormonas
Existe un tipo celular por cada hormona principal formada en la adenohipófisis.
1. Somatótropas: hormona del crecimiento humana (GH).
2. Corticótropas: corticotropina (ACTH).
3. Tirótropas: tirotropina (TSH).
4. Gonadótropas: hormonas gonadótropas, es decir, la hormona luteinizante (LH) y la hormona foliculoestimulante
(FSH).
5. Lactótropas: prolactina (PRL).
El hipotálamo controla la secreción hipofisaria
El hipotálamo es un área del cerebro que se encuentra por debajo del Tálamo. Recoge la información del bienestar
interno y controla secreción hormonal hipofisaria.
Casi toda la secreción de la hipófisis está controlada por señales hormonales o nerviosas procedentes del
hipotálamo.
La secreción de la neurohipófisis está controlada por las señales nerviosas que se originan en el hipotálamo y
terminan en la neurohipófisis. Por el contrario, la secreción de la adenohipófisis está controlada por hormonas
llamadas hormonas (o factores) de liberación y de inhibición hipotalámicas; estas se sintetizan en el propio
hipotálamo y pasan a la adenohipófisis a través de minúsculos vasos sanguíneos denominados vasos porta
hipotalámico-hipofisarios. Estas hormonas liberadoras e inhibidoras actúan sobre las células glandulares de la
adenohipófisis y rigen su secreción.
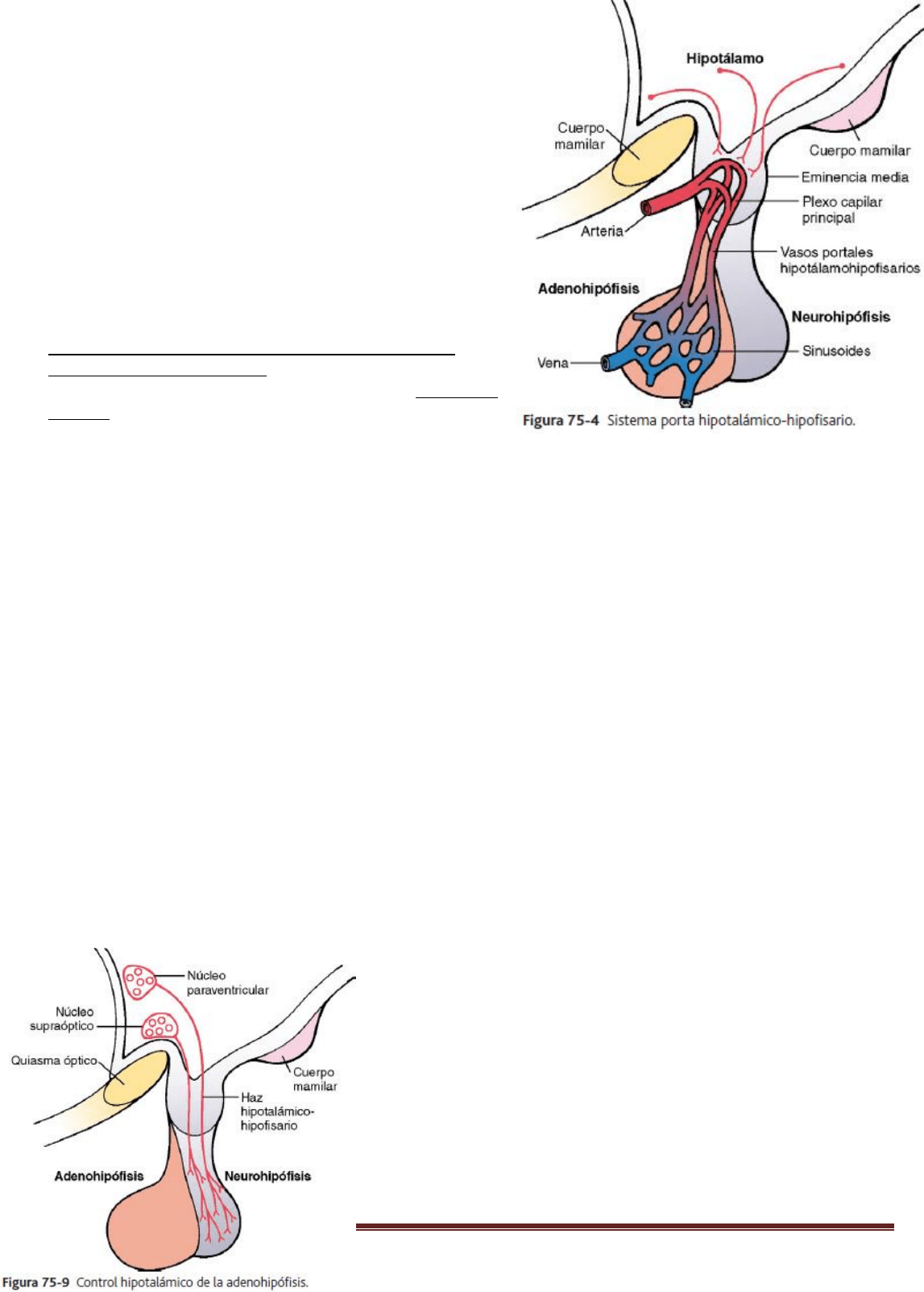
Sistema porta hipotalámico-Hipofisario:
La adenohipófisis es una glándula muy vascularizada que dispone de amplios senos capilares entre las células
glandulares. Casi toda la sangre penetra en estos senos atraviesa en primer lugar otro lecho capilar del hipotálamo
inferior (llamada eminencia media unida al tallo hipofisario). Luego la sangre fluye a través de unos diminutos
vasos porta hipotalámico-Hipofisario y accede a los senos adenohipofisiarios.
Este archivo fue descargado de https://filadd.com
FILADD.COM
Resumen Endocrino Pablo C Página 6
Es decir a la eminencia media le penetran una pequeñas
arterias y otros vasos de pequeño calibre regresan a la
superficie donde se unen formando el sistema porta. Estos
vasos descienden a lo largo del tallo y riegan los senos
adenohipofisiarios.
El sistema porta se define como un sistema común para los
organismos, en que un conducto o vía, comúnmente de
irrigación o transporte, se divide ramificándose en pequeños
conductos hasta un punto en el que estos conductillos vuelven
a unirse des-ramificándose para volver a formar la vía principal
sin cambiar de función.
Las hormonas liberadoras e inhibidoras hipotalámicas se
secretan a la eminencia media
El hipotálamo dispone de neuronas especiales que sintetizan y
secretan las hormonas liberadoras e inhibidoras hipotalámicas
encargadas de controlar la secreción de las hormonas
adenohipofisarias. Estas neuronas se originan en diversas partes del hipotálamo y envían sus fibras nerviosas a la
eminencia media.
Las terminaciones de estas fibras difieren de casi todas las demás encontradas en el sistema nervioso central, ya que
su función no consiste en transmitir señales de una neurona a otra, sino en secretar las hormonas liberadoras e
inhibidoras hipotalámicas hacia los líquidos tisulares. Estas hormonas pasan de inmediato al sistema porta
hipotalámico-hipofisario y viajan directamente a los senos de la glándula adenohipofisaria.
Las principales hormonas liberadoras e inhibidoras hipotalámicas son:
1. Tiroliberina u hormona liberadora de tirotropina (TRH), que induce la liberación de tirotropina.
2. Corticoliberina u hormona liberadora de corticotropina (CRH), que produce la liberación de corticotropina.
3. Somatoliberina u hormona liberadora de la hormona del crecimiento (GHRH), que produce la liberación de
hormona del crecimiento, y hormona inhibidora de la hormona del crecimiento (GHIH), denominada
también somatostatina, que inhibe la liberación de la hormona del crecimiento.
4. Gonadoliberina u hormona liberadora de las gonadotropinas (GnRH), que produce la liberación de dos
hormonas gonadótropas: las hormonas luteinizante (LH) y foliculoestimulante (FSH).
5. Hormona inhibidora de la prolactina (PIH), que inhibe la secreción de prolactina.
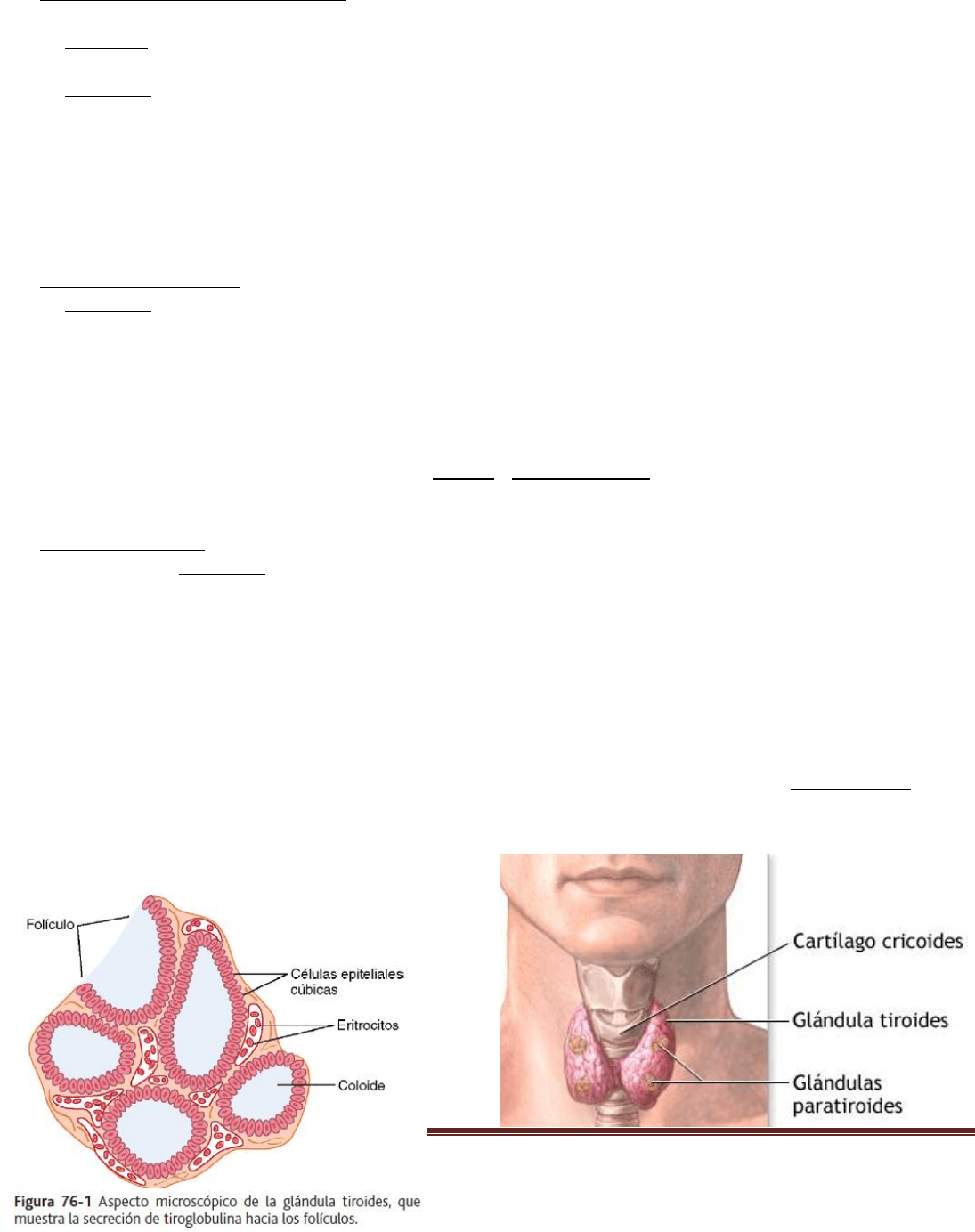
La neurohipófisis y su relación con el hipotálamo
La neurohipófisis, conocida también como hipófisis posterior se compone sobre todo de células similares a las
gliales, denominadas pituicitos. Estas células no secretan hormonas, sino que constituyen meras estructuras de
sostén para un gran número de fibras nerviosas terminales y de terminaciones nerviosas de las vías procedentes de
los núcleos supraóptico y paraventricular del hipotálamo, como se muestra en la figura.
Estas vías acceden a la neurohipófisis a través del tallo hipofisario. Las
terminaciones nerviosas son nódulos bulbosos provistos de numerosos
gránulos secretores. Estas terminaciones reposan sobre la superficie de
los capilares, hacia los que secretan dos hormonas neurohipofisarias:
1) Hormona antidiurética (ADH), llamada también vasopresina, y 2)
oxitocina.
Las hormonas se sintetizan inicialmente en los cuerpos celulares de los
núcleos supraóptico y paraventricular y después se transportan en
combinación con proteínas «transportadoras», denominadas
neurofisinas, a las terminaciones nerviosas de la neurohipófisis, a las
que tardan varios días en llegar.
Este archivo fue descargado de https://filadd.com
FILADD.COM
Resumen Endocrino Pablo C Página 7
La ADH se forma principalmente en el núcleo supraóptico, mientras que la oxitocina se forma sobre todo en el
paraventricular. Cada uno de estos núcleos puede sintetizar, además de su hormona correspondiente, hasta una
sexta parte de la otra. Cuando se transmiten los impulsos nerviosos a lo largo de las fibras desde los núcleos
supraóptico o paraventricular, los gránulos secretores de las terminaciones nerviosas liberan de inmediato la
hormona, mediante un proceso de secreción habitual conocido como exocitosis, y aquella penetra en los capilares
adyacentes.
Funciones de la hormona antidiurética
Estímulos:
- Aumento de la concentración en el LEC, en contacto con la osmorreceptores ubicados en el hipotálamo.
Funciones:
- Reduce la excreción renal de H
2
O
- Aumenta en gran medida la permeabilidad de los conductos y túbulos colectores (CC y TCP).
- Produce la apertura de las acuaporinas.
- Produce cambios en la presión arterial debido al aumento del LEC corporal.
En pocas palabras, si no existe ADH, los túbulos y conductos colectores serán prácticamente impermeables al agua,
lo que evitará su reabsorción e inducirá una pérdida extrema de líquido en la orina, que estará muy diluida.
Funciones de la oxitocina
Funciones:
- Estimula con fuerza la contracción del útero en el embarazado (en especial al final de la gestación)
- Estimula la expulsión de leche por las mamas
La glándula tiroides
Situada justo por debajo de la laringe y a ambos lados y por delante de la tráquea, es una de las glándulas endocrinas
más grandes, con un peso que oscila entre 15 y 20 g.
La tiroides secreta dos hormonas importantes, la tiroxina y la triyodotironina, conocidas a menudo como T4 y T3,
respectivamente. Ambas inducen un notable aumento del metabolismo del organismo.
La secreción tiroidea está controlada por la tirotropina (TSH), secretada por la adenohipófisis. La glándula tiroides
secreta, además, calcitonina, una hormona importante para el metabolismo del calcio.
Alrededor del 93% de las hormonas con actividad metabólica secretadas por la glándula tiroides corresponde a
tiroxina y el 7% restante, a triyodotironina. No obstante, con el tiempo, casi toda la tiroxina se convierte en
triyodotironina en los tejidos, por lo que ambas desempeñan funciones importantes.
La triyodotironina es unas cuatro veces más potente que la tiroxina, si bien se detecta una cantidad mucho menor en
la sangre y su duración es más breve.
La glándula tiroides se compone de un elevado número de folículos cerrados (100 a 300 μm de diámetro), que están
repletos de una sustancia secretora denominada coloide y revestidos por células epiteliales cúbicas que secretan a la
luz de los folículos. El componente principal del coloide es una glucoproteína de gran tamaño, la tiroglobulina, cuya
molécula contiene las hormonas tiroideas.
La glándula tiroides contiene también células C que secretan calcitonina, una hormona que contribuye a la
regulación de la concentración plasmática de iones
calcio,
Este archivo fue descargado de https://filadd.com
FILADD.COM
Resumen Endocrino Pablo C Página 8
El yoduro es necesario para la formación de tiroxina
Para formar una cantidad normal de tiroxina se precisan al año unos 50 mg de yodo (ingerido en forma de yoduros)
o el equivalente a 1 mg/semana.
La primera etapa de la formación de las hormonas tiroideas, consiste en el transporte de los yoduros desde la sangre
hasta las células y los folículos de la glándula tiroides. La membrana basal de estas células posee la capacidad
específica de bombear de forma activa el yoduro al interior celular. Este bombeo se consigue mediante la acción de
un simportador del yoduro de sodio, que cotransporta el ion yoduro a lo largo de dos iones sodio a través de la
membrana basolateral (plasma) a la célula. (El proceso de concentración de yoduro en la célula se denomina
atrapamiento de yoduro.)
Tiroglobulina y química de la formación de tiroxina y triyodotironina
El componente principal del coloide es una glucoproteína de gran tamaño, la tiroglobulina, cuya molécula contiene
las hormonas tiroideas.
Cada molécula de tiroglobulina contiene unas 70 moléculas del aminoácido tirosina, que es el sustrato principal que
se combina con el yodo para dar lugar a las hormonas tiroideas. Así pues, las hormonas tiroideas se forman dentro
de la molécula de tiroglobulina. Es decir, la tiroxina y la triyodotironina formadas a partir de los aminoácidos tirosina
constituyen una parte de la molécula de tiroglobulina durante la síntesis de las hormonas tiroideas y también
después, cuando se almacenan en el coloide de los folículos.
El primer paso crítico para la formación de las hormonas tiroideas consiste en la conversión de los iones yoduro en
una forma oxidada del yodo, que luego puede combinarse directamente con el aminoácido tirosina. La oxidación del
yodo depende de la enzima peroxidasa y su peróxido de hidrógeno acompañante, que constituyen un potente
sistema capaz de oxidar los yoduros.
El yodo oxidado se une directamente, aunque con lentitud, al aminoácido tirosina. No obstante, en las células
tiroideas el yodo oxidado se asocia a la enzima tiroidea peroxidasa que hace que el proceso tenga lugar en segundos
o minutos.
La tirosina se yoda primero a monoyodotirosina y después a diyodotirosina. A continuación, en los siguientes
minutos, horas o incluso días, números crecientes de residuos de yodotirosina se acoplan entre sí.
El principal producto hormonal de la reacción de acoplamiento es la molécula tiroxina (T 4 ), que se forma cuando se
unen dos moléculas de diyodotirosina; la tirosina forma parte aún de la molécula de tiroglobulina. En otras
ocasiones, una molécula de monoyodotirosina se une con una de diyodotirosina para formar triyodotironina (T3),
que representa alrededor de la quinceava parte del total final de hormonas.
Almacenamiento de la tiroglobulina
La glándula tiroides es la única glándula endocrina que posee la capacidad de almacenar grandes cantidades de
hormona. Una vez finalizada la síntesis de las hormonas tiroideas, cada molécula de tiroglobulina contiene hasta 30
moléculas de tiroxina y algunas de triyodotironina. De esta forma, los folículos pueden almacenar una cantidad de
hormona tiroidea suficiente para cubrir las necesidades normales del organismo durante 2 o 3 meses.
Liberación y transporte de tiroxina y triyodotironina del tiroides
La mayor parte de la tiroglobulina no se libera a la sangre circulante, sino que es preciso que la tiroxina y
triyodotironina se escindan de la molécula de tiroglobulina; a continuación, ambas se secretan en forma libre.
Varias enzimas proteinasas digieren las moléculas de tiroglobulina, y liberan la tiroxina y la triyodotironina, que se
difunden entonces a través de la base de la célula tiroidea, hacia los capilares circundantes, y de este modo pasan a
la sangre.
En condiciones normales, alrededor del 93% de la hormona tiroidea liberada por la glándula tiroides corresponde a
tiroxina y solo el 7% es triyodotironina. No obstante, en los días siguientes, la mitad de la tiroxina se desyoda con
lentitud y forma más triyodotironina.
Cuando acceden a la sangre, más del 99% de la tiroxina y la triyodotironina se combina de inmediato con diversas
proteínas plasmáticas, todas ellas sintetizadas por el hígado.
Al entrar en las células, la tiroxina y la triyodotironina se unen de nuevo a las proteínas intracelulares, aunque la
tiroxina lo hace con mayor fuerza que la triyodotironina. Por consiguiente, vuelven a almacenarse, aunque esta vez
en las propias células diana, y se utilizan con lentitud a lo largo de períodos de días o semanas.
Este archivo fue descargado de https://filadd.com
FILADD.COM

Resumen Endocrino Pablo C Página 9
Funciones de las hormonas tiroideas
El efecto general de las hormonas tiroideas consiste en la activación de la transcripción nuclear de un gran número
de genes. Por consiguiente, en casi todas las células del organismo se sintetiza una elevada proporción de enzimas
proteicas, proteínas estructurales, proteínas transportadoras y otras sustancias. El resultado neto es un aumento
generalizado de la actividad funcional de todo el organismo.
Antes de actuar sobre los genes e incrementar la transcripción genética, gran parte de la tiroxina liberada pierde un
yoduro y se forma triyodotironina. Alrededor del 90% de las moléculas de hormona tiroidea que se unen a los
receptores es triyodotironina.
Las hormonas tiroideas activan receptores nucleares
Los receptores de hormona tiroidea se encuentran unidos a las cadenas genéticas de ADN o junto a ellas. El receptor
suele formar un heterodímero con el receptor retinoide X (RXR) en los elementos específicos de respuesta a la hormona
tiroidea del ADN. Después de unirse a esta hormona, los receptores se activan e inician el proceso de transcripción. A
continuación, se forma una cantidad elevada de ARN mensajero de distintos tipos, seguido en unos minutos u horas de
la traducción del ARN en los ribosomas citoplásmicos, para formar cientos de proteínas intracelulares nuevas.
Las hormonas tiroideas incrementan las actividades metabólicas de casi todos los tejidos del organismo.
La velocidad de utilización de los alimentos como fuente de energía se encuentra muy acelerada. Aunque la síntesis de
proteínas aumenta, también lo hace el catabolismo proteico. La velocidad de crecimiento de las personas jóvenes
experimenta una gran aceleración. Los procesos mentales se estimulan y las actividades de las demás glándulas
endocrinas se potencian.
Las hormonas tiroideas incrementan el número y la actividad de las mitocondrias
Las hormonas tiroideas facilitan el transporte activo de iones a través de la membrana celular
Una de las enzimas que aumentan en respuesta a la hormona tiroidea es la Na/K ATPasa; a su vez, este aumento de la
actividad potencia el transporte de los iones sodio y potasio a través de la membrana celular de determinados tejidos.
Este proceso requiere energía e incrementa la cantidad de calor producida por el organismo, por lo que se ha propuesto
que quizá constituya uno de los mecanismos mediante los cuales la hormona tiroidea eleva el metabolismo.
Efecto en el crecimiento
En los niños hipotiroideos, la velocidad de crecimiento es mucho más lenta, mientras que los hipertiroideos a menudo
experimentan un crecimiento esquelético excesivo, por lo que son bastante más altos de lo que les correspondería
según su edad. No obstante, los huesos también maduran con mayor rapidez y las epífisis se cierran a una edad
temprana, por lo que el crecimiento resulta más breve y la estatura final en la edad adulta puede ser, en realidad,
menor.
Estimulación del metabolismo de los H de Carbono en casi todas las fases entre ellos:
La rápida captación de glucosa por las células.
El aumento de la glucólisis.
El incremento de la gluconeogenia.
Una mayor absorción en el tubo digestivo.
Una mayor secreción de insulina.
Estimulación del metabolismo de los lípidos:
a. Potencia su metabolismo
El incremento de la Hormona tiroidea induce:
a. Un descenso de la concentración plasmática de colesterol, fosfolípidos y triglicéridos
b. Eleva la concentración plasmática de ácidos grasos libres.
Aumenta las necesidades de vitaminas
Aumenta el apetito
Este archivo fue descargado de https://filadd.com
FILADD.COM
Resumen Endocrino Pablo C Página 10
Aumenta el consumo de alimentos
Sobre el aparto cardiovascular:
a. Aumenta el flujo sanguíneo
b. Aumenta el gasto cardíaco
c. Aumenta la frecuencia cardíaca
d. Aumenta la fuerza cardíaca
e. Aumento de la respiración
Aumenta la respiraciónel aumento del metabolismo aumenta el consumo de O
2
y la formación de CO
2
.
Aumento de la motilidad digestiva favorece la secreción de jugo gástrico
Tienen efectos excitadores sobre el sistema nervioso central
a. Acelera la función cerebral
Tienen efecto en la función muscular
Tienen efecto sobre el sueño ya que las personas con hipertiroidismo suelen sentirse siempre cansadas y le es
difícil conciliar el sueño
Tienen efecto sobre otras glándulas endocrina
Tienen efecto sobre la función sexual:
a. En las mujeres la falta de H. tiroidea produce: menorragia y polimenorrea (hemorragia menstrual excesiva y
frecuente).
Regulación de la secreción de hormonas tiroideas
Para mantener una actividad metabólica normal en el organismo, es necesario que se secrete una adecuada
cantidad de hormona tiroidea.
La TSH adenohipofisaria incrementa la secreción tiroidea. Los efectos que ejerce sobre esta glándula son:
1) Eleva la proteólisis de la tiroglobulina
2) Incrementa la activad de la bomba de yoduro
3) Intensifica la yodación de la tirosina
4) Aumenta el tamaño y la activad secretora de la células tiroideas
5) Incrementa el número de las células tiroideas
Efecto de retroalimentación de las hormonas tiroideas:
El ascenso de la concentración de la hormona en los líquidos corporales reduce la secreción de TSH.
Enfermedades del tiroides:
Hipertiroidismo:
Causas:
Aumento del tamaño de la glándula tiroides que llega a duplicarse o triplicarse producción de hormonas
Hiperplasia celular y un plegamiento del revestimiento de las células foliculares
Aumento de la secreción de cada célula (5 a 15 veces de su valor normal)
Se puede deber por ejemplo a la enfermedad de Graves o un Adenoma tiroideo
Síntomas:
Estado de gran excitabilidad
Intolerancia al calor
Aumento de la sudoración
Adelgazamiento leve o extremo
Diarrea de diversa magnitud
Este archivo fue descargado de https://filadd.com
FILADD.COM
Resumen Endocrino Pablo C Página 11
Debilidad muscular
Nerviosismo u otro trastornos psíquicos
Fatiga extrema e incapacidad para conciliar el sueño
Temblor de las manos
Exoftalmos: cierto grado de prominencia de los globos oculares.
Hipotiroidismo
Causas:
Se inicia por autoinmunidad contra la glándula tiroides (enfermedad de Hashimoto)
o Puede en algunos casossufre una tiroiditis (inflamación del tiroides)
o Puede en algunos casos destruir la glándula por fibrosis
Bocio (significa =glándula tiroides de gran tamaño) coloide endémico asociado a la deferencia alimenticia de
yodo
Bocio coloide idiopático no tóxico tienen tiroiditis en partes de la glándula
Estos casos de bocio coloide se puede deber a:
o Una deficiencia del mecanismo de atrapamiento del yoduro
o Sistema de peroxidasas defectuoso
o Acoplamiento defectuoso de la tirosina yodada
Síntomas
Fatiga y somnolencia extrema
Lentitud muscular desmesurada
Disminución de la F. Cardíaca
Menos gasto cardíaco
Reducción del volumen sanguíneo
En ocasiones aumento del peso corporal
Estreñimiento
Lentitud metal
Reducción del crecimiento del cabello
Descamación cutánea
Voz ronca y carraspea
Mixedema (rasgo característico) grandes bolsas bajo los ojos y tumefacción facial.
Elevación del colesterol sanguíneo incremento de la arteriosclerosis
Este archivo fue descargado de https://filadd.com
FILADD.COM
Resumen Endocrino Pablo C Página 12
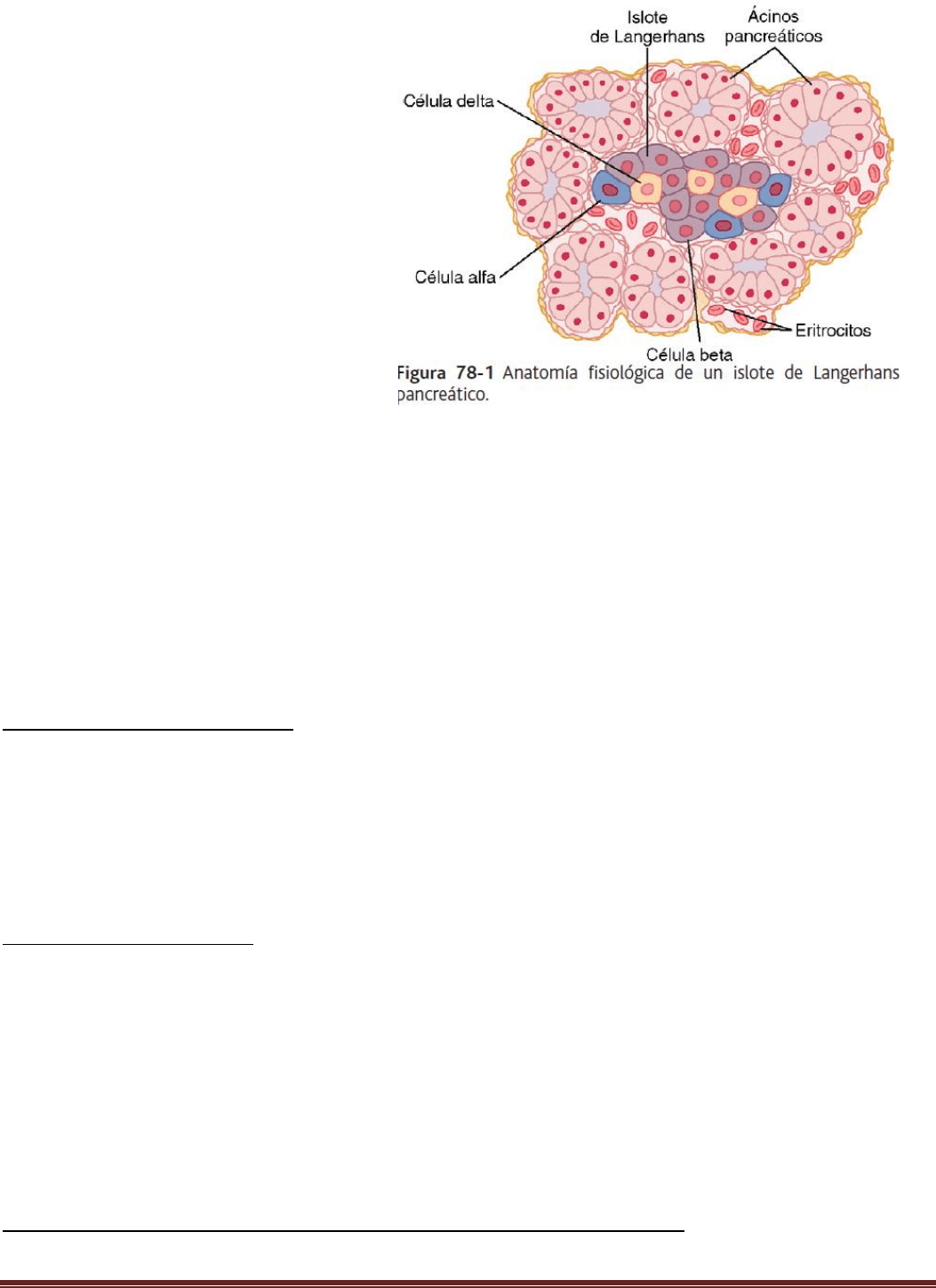
Páncreas
El páncreas, además de poseer funciones
digestivas, secreta dos hormonas, la insulina y
el glucagón, que son esenciales para la
regulación del metabolismo de la glucosa, los
lípidos y las proteínas.
Se compone de dos grandes tipos de tejidos,
como ilustra:
1) Los ácinos, que secretan jugos digestivos al
duodeno, y 2) los islotes de Langerhans, que
secretan insulina y glucagón de forma directa a
la sangre.
El páncreas humano cuenta con 1 a 2 millones
de islotes de Langerhans; los islotes se
organizan en torno a pequeños capilares, hacia
los que vierten sus hormonas, y contienen tres
tipos fundamentales de células, alfa (α), beta
(β) y delta (δ), que se diferencian entre sí por
sus características morfológicas y de tinción.
Las células β representan casi el 60% de la totalidad de las células de los islotes y se encuentran sobre todo en el
centro de cada uno y secretan insulina y amilina, hormona que suele liberarse en paralelo con la insulina, pese a que
no se conoce bien su función. Las células α, que componen casi el 25% del total, secretan glucagón, y las células δ,
que representan el 10%, somatostatina. Además, existe por lo menos otro tipo de célula, la célula PP, en menor
cantidad y que produce una hormona de función incierta denominada polipéptido pancreático. Las relaciones
íntimas entre estos tipos celulares de los islotes de Langerhans facilitan la comunicación intercelular y el control
directo de la secreción de algunas de las hormonas. Por ejemplo, la insulina inhibe la secreción de glucagón; la
amilina inhibe la secreción de insulina y la somatostatina, la de insulina y glucagón.
Insulina
La insulina y sus efectos metabólicos
Si hay un exceso de alimentos energéticos en la dieta y, sobre todo, de hidratos de carbono, aumenta la secreción de
insulina. A su vez, la insulina desempeña una función primordial en el almacenamiento de la energía sobrante. Si se
consumen hidratos de carbono en exceso, estos se depositarán principalmente como glucógeno en el hígado y en los
músculos. Al mismo tiempo, y también por efecto de la insulina, el exceso de hidratos de carbono que no puede
almacenarse como glucógeno se convierte en grasa y se conserva en el tejido adiposo. En cuanto a las proteínas, la
insulina ejerce un efecto directo para que las células absorban más aminoácidos y los transformen en proteínas. Por
último, esta hormona inhibe la degradación de las proteínas intracelulares.
Química y síntesis de la insulina
Esta preproinsulina inicial luego se desdobla en el retículo endoplásmico para formar la proinsulina. La mayor parte
de la proinsulina sigue escindiéndose en el aparato de Golgi para formar insulina.
La mayor parte de la insulina liberada hacia la sangre circula de forma no ligada. Dado que su semivida plasmática es
de unos 6 min por término medio, desaparece de la circulación en unos 10 a 15 min. Con excepción de la parte de
insulina que La mayor parte de la insulina liberada hacia la sangre circula de forma no ligada. Dado que su semivida
plasmática es de unos 6 min por término medio, desaparece de la circulación en unos 10 a 15 min. Con excepción de
la parte de insulina que se une a los receptores de las células efectoras, el resto se degrada por efecto de la enzima
insulinasa, sobre todo en el hígado y, en menor medida, en los riñones y en los músculos, y de forma muy ligera en
casi todos los demás tejidos. Su desaparición inmediata del plasma tiene interés porque, a veces, es tan importante
desactivar con rapidez el efecto de la insulina como activar sus funciones reguladoras.
Es el receptor activado, y no la insulina, es el que desencadena los efectos posteriores
Este archivo fue descargado de https://filadd.com
FILADD.COM

Resumen Endocrino Pablo C Página 13
La insulina se une a la subunidad α del receptor, lo que determina la autofosforilación de la subunidad β del mismo.
A su vez, esto induce una actividad tirosina cinasa. La actividad tirosina cinasa del receptor desencadena una cascada
de fosforilación celular que aumenta o reduce la actividad de diversas enzimas, incluidos los sustratos del receptor
de insulina, que intervienen en los efectos en el metabolismo de la glucosa, lípidos y proteínas.
El efecto neto es la activación de algunas de estas enzimas y la inactivación de otras. Por este mecanismo, la insulina
dirige la maquinaria metabólica intracelular para provocar los efectos deseados sobre el metabolismo de los hidratos
de carbono, los lípidos y las proteínas. Los principales efectos finales de la estimulación insulínica son los siguientes:
1. Pocos segundos después de la unión de la insulina a sus receptores de membrana, se produce un notable
incremento de la captación de glucosa por las membranas de casi el 80% de las células, sobre todo de las células
musculares y adiposas.
La glucosa, que se transporta en mayor cantidad a la célula, se fosforila de inmediato y sirve de sustrato para todas
las funciones metabólicas habituales de los hidratos de carbono.
2. La membrana celular se hace más permeable para muchos aminoácidos y para los iones potasio y fosfato, cuyo
transporte al interior de la célula se incrementa.
3. En los 10 a 15 min siguientes se observan efectos más lentos que cambian la actividad de muchas más enzimas
metabólicas intracelulares. Estos efectos se deben, sobre todo, a una variación de la fosforilación enzimática.
4. Durante algunas horas e incluso días tienen lugar otros efectos, mucho más lentos, que se deben a cambios de la
velocidad de traducción de los ARN mensajeros dentro de los ribosomas para dar lugar a nuevas proteínas e incluso
(los efectos más tardíos) a variaciones de las velocidades de transcripción del ADN del núcleo celular.
Insulina y los hidratos de carbono
Favorece la captación y el metabolismo musculares de la glucosa
Durante gran parte del día, la energía utilizada por el tejido muscular no depende de la glucosa, sino de los ácidos
grasos. La razón principal de esta dependencia de los ácidos grasos es que la membrana muscular en reposo es muy
poco permeable a la glucosa, salvo que la fibra muscular reciba el estímulo de la insulina; la cantidad de insulina
secretada entre las comidas es demasiado escasa para propiciar una entrada importante de glucosa dentro de las
células musculares. Sin embargo, existen dos situaciones en las que el músculo consume mucha glucosa. Una de ellas
es el ejercicio moderado e intenso. Para esta utilización de la glucosa no se necesitan grandes cantidades de insulina,
porque la contracción facilita la difusión de la glucosa en la célula. El segundo estado en el que el músculo consume
mucha glucosa son las horas siguientes a las comidas. En esta fase, la concentración sanguínea de glucosa se eleva y
el páncreas secreta mucha insulina. La insulina «extra» induce un transporte rápido de la glucosa al miocito. Por
tanto, este utiliza glucosa en lugar de ácidos grasos durante ese período.
Depósito de glucógeno en el músculo
Si el músculo no se ejercita después de una comida, pero la glucosa se transporta en abundancia a su interior, la
mayor parte de ella se depositará como glucógeno muscular y no se empleará como sustrato energético, hasta un
límite del 2 al 3% de su concentración. Este glucógeno se aprovechará más tarde para fines energéticos. El glucógeno
es útil especialmente para los períodos cortos de utilización intensa de energía por el músculo o incluso para los
instantes de máxima energía anaerobia, es decir, para los escasos minutos de degradación glucolítica del glucógeno
a ácido láctico, que suceden incluso en ausencia de oxígeno.
La insulina para facilitar el transporte de glucosa por la membrana de la célula muscular
La insulina facilita la captación, el almacenamiento y la utilización de glucosa por el hígado
Uno de los efectos más importantes de la insulina es el depósito rápido de glucógeno en el hígado a partir de casi
toda la glucosa absorbida después de una comida. Más tarde, entre las comidas, cuando ya no se dispone de
alimento y la glucemia empieza a descender, la secreción de insulina disminuye con rapidez y el glucógeno hepático
se transforma de nuevo en glucosa, que se libera otra vez a la sangre para evitar que la glucemia descienda
demasiado.
La insulina favorece la conversión del exceso de glucosa en ácidos grasos e inhibe la gluconeogenia hepática
Este archivo fue descargado de https://filadd.com
FILADD.COM

Resumen Endocrino Pablo C Página 14
Cuando la cantidad de glucosa que entra en el hepatocito es superior a la que se puede depositar como glucógeno o
utilizar para su metabolismo local en el hígado, la insulina favorece la conversión de todo este exceso de glucosa en
ácidos grasos. Luego estos ácidos grasos se empaquetan como triglicéridos dentro de lipoproteínas de muy baja
densidad, que son transportadas por la sangre al tejido adiposo para depositarse como grasa.
Insulina y los lípidos
La insulina favorece la síntesis y el depósito de lípidos
En primer lugar, aumenta la utilización de la glucosa en casi todos los tejidos orgánicos y reduce automáticamente la
utilización de la grasa, es decir, ahorra lípidos. No obstante, la insulina también fomenta la síntesis de ácidos grasos,
en mayor medida cuantos más hidratos de carbono se ingieran, dado que estos no se emplean de inmediato para
producir energía y aportan el sustrato para la síntesis de grasas. Gran parte de esta síntesis tiene lugar en los
hepatocitos; luego los ácidos grasos son transportados desde el hígado por las lipoproteínas de la sangre a las células
adiposas, donde se almacenan.
La deficiencia de insulina aumenta el uso de la grasa con fines energéticos
Todos los fenómenos relacionados con la degradación de los lípidos y su uso con fines energéticos se estimulan en
gran medida cuando falta insulina. Esta potenciación tiene lugar incluso en condiciones normales entre las comidas,
porque la secreción de insulina es mínima, pero puede agravarse en personas con diabetes mellitus, dado que en ella
la secreción de insulina es casi nula.
El déficit de energía provoca lipolisis de la grasa almacenada con liberación de ácidos grasos a la sangre aumentando
el colesterol provocando arteriosclerosis.
La enzima lipasa sensible a la insulina de las células adiposas experimenta una gran activación. Con ello, se hidrolizan
los triglicéridos almacenados y se liberan enormes cantidades de ácidos grasos y de glicerol a la sangre circulante. La
consecuencia es que las concentraciones plasmáticas de ácidos grasos libres empiezan a ascender a los pocos
minutos. Estos ácidos grasos libres se transforman luego en el sustrato energético principal de casi todos los tejidos
orgánicos, salvo el encéfalo.
El exceso de ácidos grasos del plasma, junto con la falta de insulina, favorece también la conversión hepática de
algunos de los ácidos grasos en fosfolípidos y colesterol. Este incremento de los lípidos, sobre todo del colesterol,
acelera el desarrollo de ateroesclerosis en los enfermos con diabetes grave.
Insulina y las proteínas
La insulina facilita la síntesis y el depósito de proteínas
En las horas que siguen a una comida, si la sangre circulante contiene un exceso de nutrientes, se depositarán en los
tejidos hidratos de carbono, grasas y proteínas; para ello se precisa la insulina.
1. La insulina estimula el transporte de muchos aminoácidos al interior de las células.
2. La insulina aumenta la traducción del ARN mensajero, es decir, la síntesis de nuevas proteínas. Por vías
desconocidas, la insulina «activa» la maquinaria ribosómica. Cuando falta insulina, los ribosomas dejan de trabajar;
la insulina actúa casi como un mecanismo de «encendido-apagado».
3. Durante un período aún más largo, la insulina acelera, además, la transcripción de determinadas secuencias
genéticas del ADN de los núcleos celulares, haciendo que se formen mayores cantidades de ARN y prosiga la síntesis
de proteínas;
4. La insulina inhibe el catabolismo de las proteínas.
5. Dentro del hígado, la insulina disminuye el ritmo de la gluconeogenia. En consecuencia, reduce la actividad de las
enzimas neoglucogénicas. Como los sustratos más utilizados para la síntesis de glucosa mediante gluconeogenia son
los aminoácidos del plasma, la supresión de la gluconeogenia hace que estos se conserven para su depósito corporal
en forma de proteínas.
La deficiencia de insulina provoca el descenso de las proteínas y el incremento de los aminoácidos en el plasma
Cuando falta insulina, el depósito de proteínas se interrumpe casi por completo. El catabolismo de las proteínas
aumenta, la síntesis de proteínas cesa y se vierten enormes cantidades de aminoácidos al plasma. La concentración
plasmática de aminoácidos aumenta mucho y casi todo el exceso se emplea bien directamente, como fuente
Este archivo fue descargado de https://filadd.com
FILADD.COM

Resumen Endocrino Pablo C Página 15
energética, bien como sustrato de la gluconeogenia. Esta degradación de los aminoácidos provoca, en última
instancia, una mayor eliminación de urea en la orina. La pérdida resultante de las proteínas es una de las secuelas
más graves de la diabetes mellitus intensa. Puede causar una debilidad extrema, así como la alteración de
numerosas funciones orgánicas.
Tejido insulinodependiente: musculo, hígado, tejido adiposo.
Tejido no insulinodependiente: encéfalo, retina, epitelio germinal de las gónadas
Glucagón
El glucagón es una hormona secretada por las células α de los islotes de Langerhans cuando disminuye la glucemia y
cumple varias funciones opuestas a las de la insulina.
Estímulos: hipoglucemia, estimulo simpáticos y catecolaminas, aminoácidos, ejercicio intenso (posiblemente por el
aumento de aminoácidos circulante o por estimulación nerviosa autónoma de los islotes).
Inhibidores: hiperglucemia, insulina, somatostatina, Ac grasos libres
Funciones:
Hidratos de carbono: el efecto más importante es aumentar la glucogenólisis hepática, aumentando la glucemia.
Esto se da a través de un sistema de cascada que utiliza AMPc como segundo mensajero en el que cada producto
sucesivo se fabrica en cantidad mayor al precursor representando un mecanismo de amplificación muy poderoso.
Basta con unos microgramos de glucagón para aumentar la glucemia 2 o 3 veces en pocos minutos.
Aumenta la gluconeogensis hepática. Activando numerosas enzimas necesarias para el transporte de aminoácidos,
estimulando la velocidad de absorción de aminoácidos por los hepatocitos y la conversación posterior de muchos de
ellos en glucosa.
Otros efectos: otros efectos que se manifiestan solo cuando su concentración aumenta muy por encima del máximo
habitual: activación de la lipasa de células adiposas que aumenta la disponibilidad de aminoácidos, inhibe el depósito
de triglicéridos en el hígado aumentando la disponibilidad para los demás tejidos.
Estimula la contracción cardiaca. Aumenta el flujo sanguíneo de algunos tejidos sobre todo riñones. Estimula la
secreción biliar. Inhibe la secreción de HCl en el estómago.
GLUCEMIA
Valores normales
En ayuno 80-90mg/100ml
Postprandial 120-140mg/100ml
Regulación hormonal y nerviosa
1. el hígado funciona como un importante sistema amortiguador de la glucemia. Cuando la glucemia se eleva
después de una comida se secreta insulina la cual hace que 2/3 de la glucosa absorbida se almacenen en el
hígado en forma de glucógeno. Cuando la glucemia y la insulina disminuyen, le hígado devuelve la glucosa a
la sangre.
2. La insulina y el glucagón operan como sistemas retroactivos esenciales para mantener la glucemia dentro de
límites normales. Aumenta la glucemia --> se secreta insulina --> se reduce la glucemia. Descenso de la
glucemia --> se secreta glucagón --> aumenta la glucemia. El mecanismo de la insulina tiene mucha más
importancia que el glucagón
3. En las hipoglucemias graves, el descenso de la glucemia sobre el hipotálamo estimula el Simpático. La
adrenalina secretada por las glándulas suprarrenales propicia la liberación de glucosa por el hígado.
4. Tanto la hormona de crecimiento como el cortisol se liberan cuando existe una hipoglucemia prolongada.
Reducen la tasa de utilización de glucosa por casi todas las células del organismo que empiezan a consumir
más lípidos.
Importancia de la regulación de la glucemia
No puede descender demasiado porque:
1. La glucosa es el único nutriente que utilizan en forma habitual el encéfalo, la retina y el epitelio germinal de
las gónadas.
No puede ascender demasiado porque:
2. Puede ejercer una presión osmótica intensa en el LEC, provocando deshidratación celular
3. Concentración exagerada de glucosa en sangre induce una pérdida de la misma por la orina, causando una
diuresis osmótica renal que disminuye los líquidos y electrolitos orgánicos.
Este archivo fue descargado de https://filadd.com
FILADD.COM
Este documento contiene más páginas...
Descargar Completo
ENFERMEDAD PERIODONTAL.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.