La eliminación de
los intrones del pre
RNAm deja un mensajero usual. -2.2 kb mediante un
denominado corte y empalme del RNA.
La eliminación de los intrones constituye una parte
importante de la producción del RNA en todas las
eucariotas.
RNA nuclear. Su tamaño promedio es mucho mayor
que el de RNAm, es muy inestable y la complejidad de
su secuencia es mucho mayor. Dada su amplia
distribución, se le denominó RNA nuclear
heterogéneo (RNAhn).
La forma física del RNAhn es de
ribonucleoprotcína (RNPhn), donde el RNAhn está
unido a proteínas.
El corte
y empalme ocurre en el núcleo, cunado
las otras modificaciones que experimentan los RNA
estén sintetizados. El producto de la transcripción tiene
un capuchón en el extremo 5’ (motivo de la
eliminación de intrones) y está poliadenilado en el
extremo 3’
Tipos de corte y empalme:
Se pueden identificar varios tipos de sistemas de
corte y empalme:
• Los intrones se retiran del preRNAm de las
eucariotas superiores por un sistema que reconoce
solo las secuencias consenso conservadas en los
limites intrón-exón y el interior del intrón. Esa
reacción requiere un gran aparato de corte y
empalme que asume la forma de un arreglo de
proteínas y ribonucleoproteinas.
El mecanismo de corte y empalme requiere
transesterificación, además de que el centro
catalítico incluye el RNA y proteínas.
• Ciertos RNA tienen la capacidad de excluir sus
intrones de manera autónoma, los cuales forman 2
grupos según su estructura secundaria o terciaria,
ambos usan reacciones de transesterificación en el
que el RNA es el agente catalítico.
• La eliminación de intrones de los precursores
RNAt nucleares de levaduras implican actividades
enzimáticas. Estas reacciones de corte y empalme
son por enzimas que dan lugar a la escisión y
ligadura.

El corte y empalme de los intrones nucleares se debe
considerar la naturaleza de los sitios de corte y
e
mpalme, los dos límites exón-intrón que incluyen los
sitios de escisión y nueva unión.
Es posible asignar un
extremo específico
a cada intrón dependiendo de la
conservación de las uniones exón-intrón, que pueden
ser alineadas
para construir la secuencia de consenso.
Los submdices indican el porcentaje de aparición de la
base específica en cada posición de consenso. La
conservación es elevada sólo inmediatamente dentro
del
! intrón en las supuestas uniones, de modo que la
secuencia de un intrón genérico se identifica como:
GU .
.... .. AG
Se nombran de izquierda a derecha, a lo largo del
intrón, como sitio de corte y empalme 5' (a veces
llamado sitio izquierdo o donador) y sitio de corte y
empalme 3' (también llamado sitio derecho o aceptor).
Los extremos de un intrón no son homólogos ni
ampliamente complementarios, las uniones, sin
embargo, tienen secuencias de consenso bien
conservadas, aunque bastante cortas.
Cuando sólo se ha perdido intrón, esto ocurre
prácticamente siempre en el 5 o 6, cualquiera puede ser
el que se pierda primero. El intrón 3 nunca (o cuando
muy rara vez) se pierde en uno de los primeros
pasos del corte y empalme.
Las reacciones de corte y empalme como tales
son independientes de la transcripción o modificación
del RNA, sin embargo. ocurren normalmente
en forma coordinada y su eficacia puede ser influida
por otros sucesos de procesamiento.
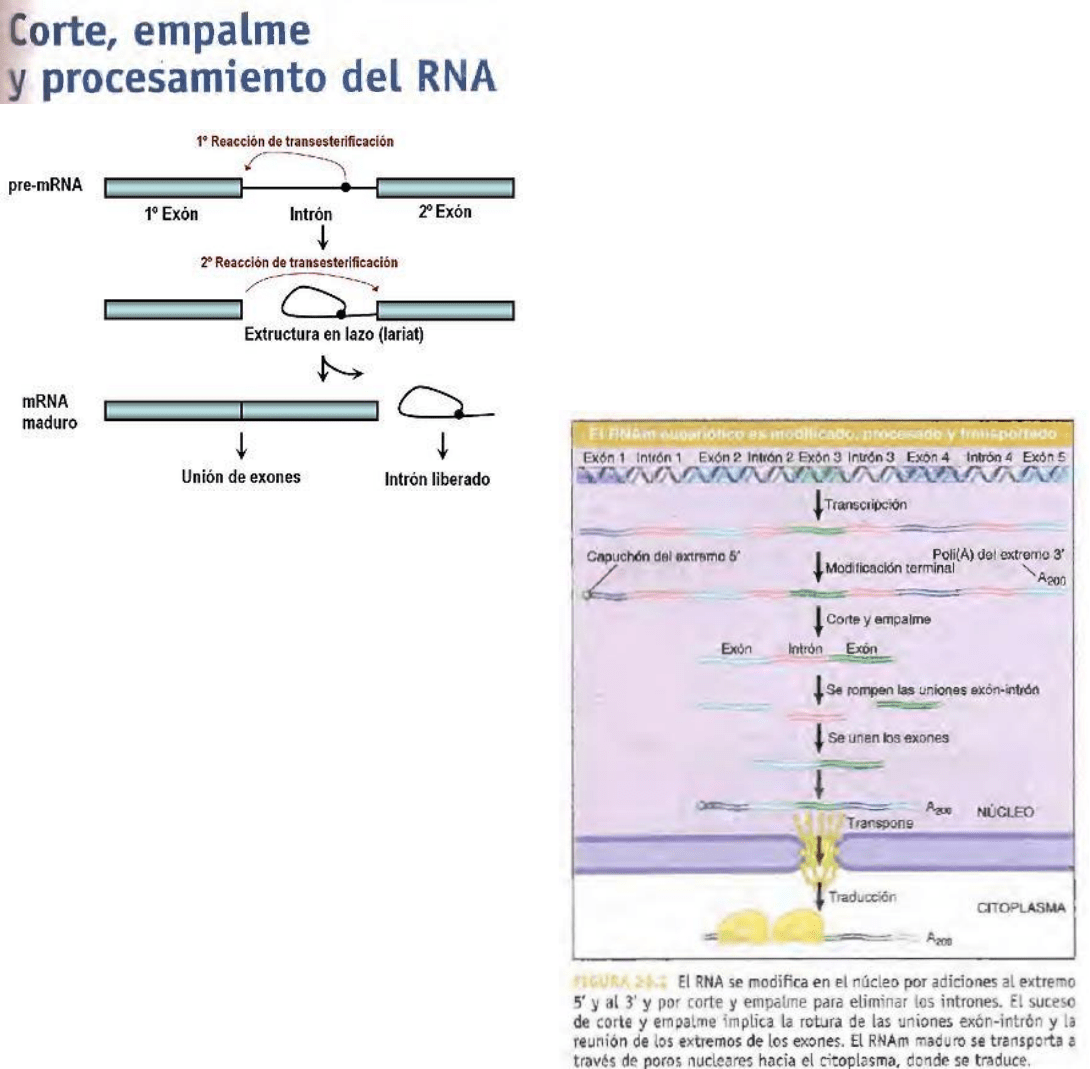
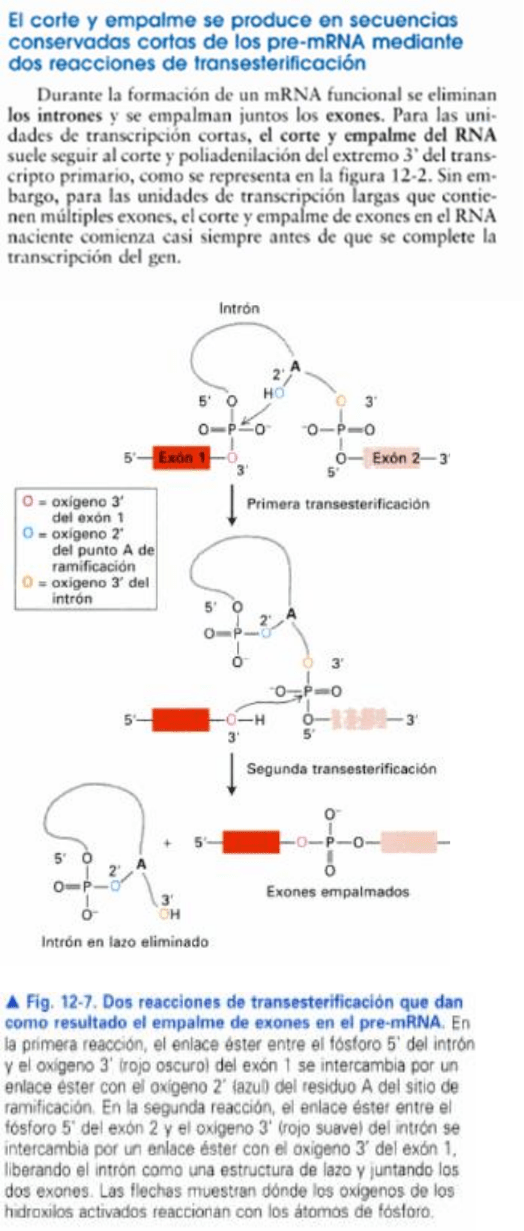
El primer paso es escindir en e l sitio de corte
y empalme 5', separando el exón izquierdo de la
molécula de intrón-exón derecha; el exón izquierdo
asume la forma de una molécula lineal y la molécula
intrón-exón derecha forma un lazo, en el cual el
extremo 5' generado en el final del intrón se une
por enlace 5'- 2' con una base, dentro del intrón. La
base diana es una A en una secuencia llamada sitio
de ramificación.
La escisión en el sitio 3' de corte y empalme libera
al intrón libre con forma de lazo; el exón derecho se
liga (corta y empalma) con el exón izquierdo.
El sitio de ramificación yace a una distancia de18a 40
nucleótidos en flujo <ascendente respecto
del sitio 3' de corte y empalme, en el cual las
mutaciones.
El enlace constituido por el lazo va, de la posición
5' de la G invariable que estaba en el extremo
5' del intrón a la posición 2' de la A invariable
sitio de ramificación, que corresponde a la
molécula A de la caja UACUAAC de las levaduras.
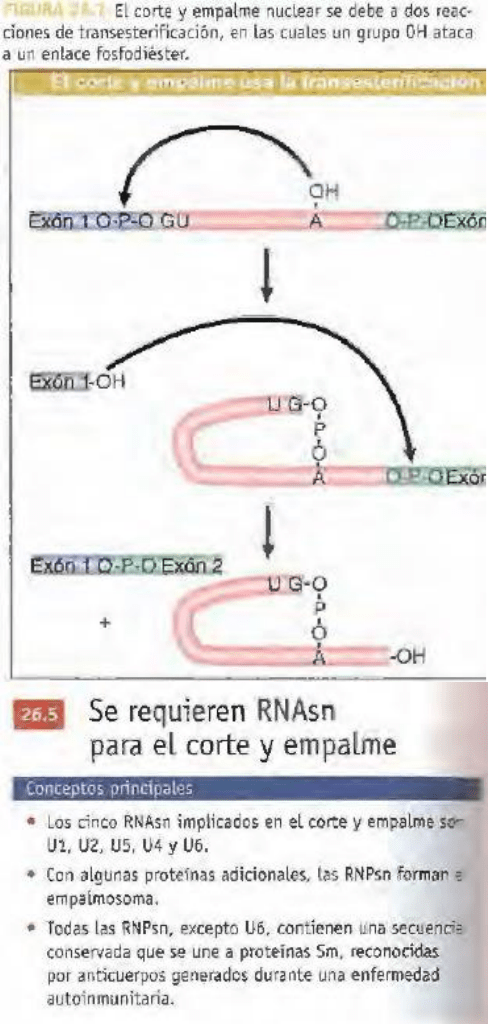
el primer paso es un ataque nucleótido por el
extremo 2'-0H de la A invariable de la secuencia
UACUAAC en el sitio 5' de corte y empalme. El
segundo paso, el extremo 3'-0H libre del exón
librado por la primera reacción, ataca ahora el del sitio
3' de corte y empalme.
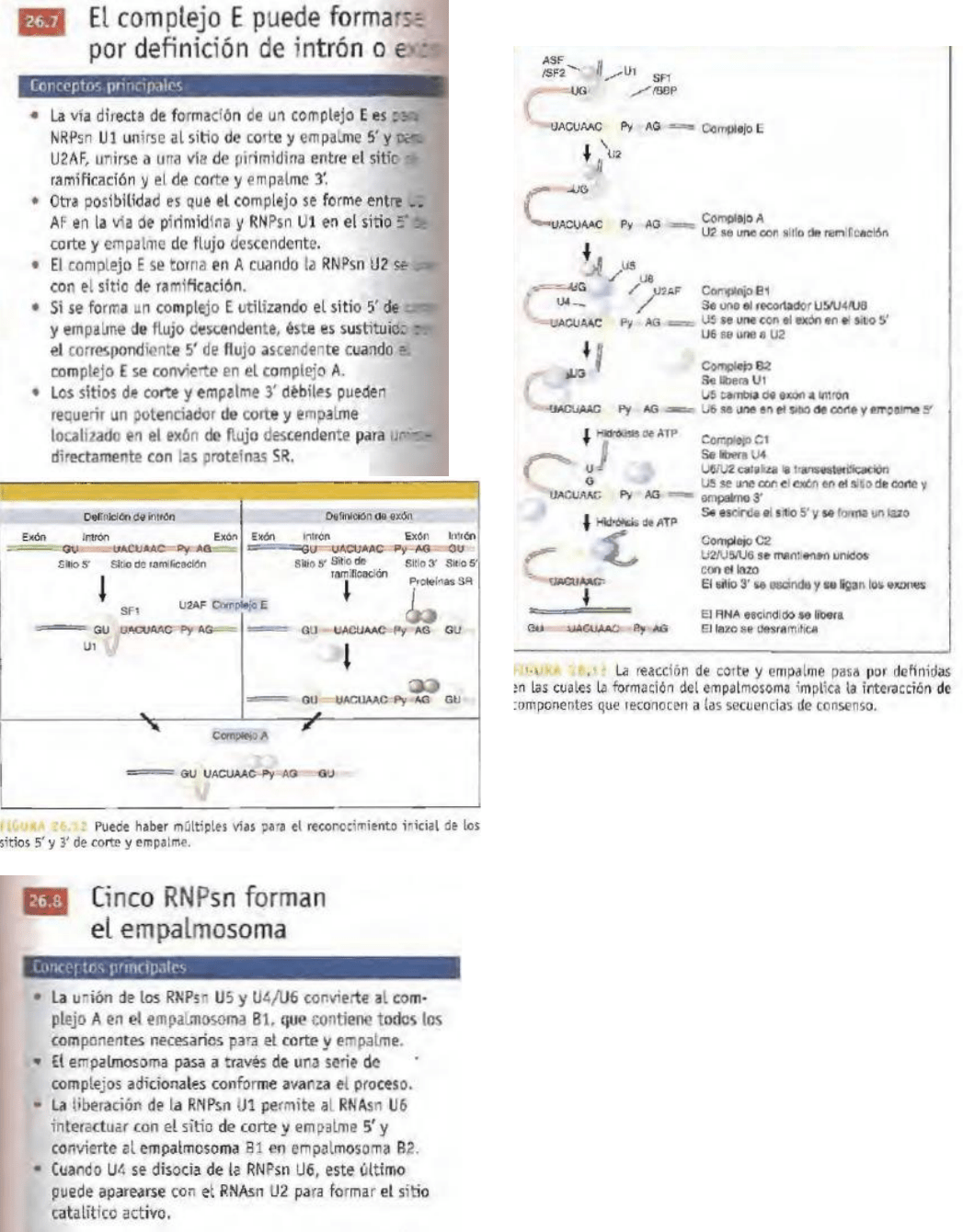
Los sitios de corte y empalme 5' y 3', y la secuencias
de ramificación, son reconocidos por componentes
del aparato de corte y empalme que se ensamblan para
formar un gran complejo, el cual une los sitios
5’y 3' de corte y empalme antes de que ocurra
cualquier reacción, lo que explica por qué una
diferencia en cualquiera de esos sitios puede impedir el
inicio de la reacción. El complejo se ensambla de
manera secuencial en el pre-RNAm y los complejos
fraccionadores de diferentes tamaños pueden
reconocer diversos intermediarios. El corte y empalme
tiene lugar sólo después de que todos los componentes
se han ensamblado.
Los restringidos al núcleo se llaman RNA nucleares
pequeños (RNAsn ), y los del citoplasma,
RNA citoplasmáticos pequeños (RNAsc).
Las RNPsn involucradas en el corte y empalme
además de muchas otras proteínas, forman un
gran complejo específico llamado empalmosoma.
El empalmosoma se forma en el RNA precursor
intacto y pasa a través de un estado intermedio en el
cual contiene la molécula lineal individual del exón
5' y el lazo-intrón-exón derecho. El complejo tiene
muy poco producto de corte y empalme, lo cual
sugiere que suele liberarse de inmediato, después de la
escisión del sitio 3' y la ligadura de los exones.
Las RNPsn implicadas en el corte y empalme
son U l, U2, US, U4 y U6, nombrados de acuerdo
con Jos .RNAsn presentes. Cada RN'Psn contiene un
sólo RNAsn y varias proteínas.
RNPsn consta de un grupo de ocho proteínas.
Algunas de las proteínas de la RNPsn pueden
participar directamente en el corte y empalme,
mientras que otras tal vez sean necesarias para
actividades estructurales o sólo para el ensamblaje o
las interacciones entre panículas de RNPsn.
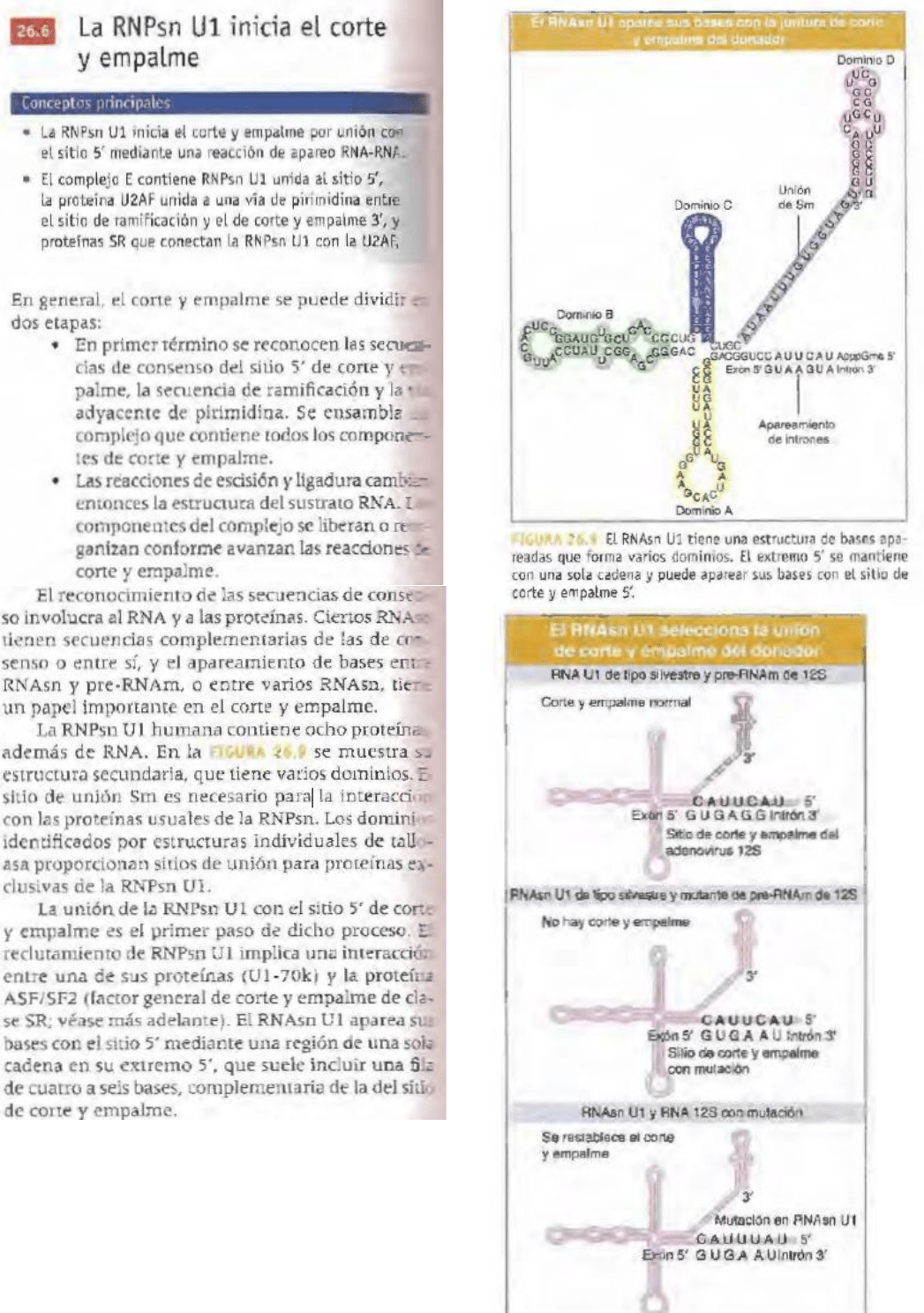
Una secuencia cercana al extremo 5' del RNAsn aparea
sus bases a la secuencia de ramificación en el intrón.

corte y empalme.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.