70
Capítulo 16 RECEPTORES Y MECANISMOS
DE TRANSDUCCIÓN DE SEÑALES
Irene L. Ennis,
María Celeste Villa-Abrille
Palabras clave
Comunicación intercelular, receptores, ligandos, segundos mensajeros.
Abreviaturas utilizadas
AA: ácido araquidónico
AC: adenilato ciclasa
AMPc: AMP cíclico
CaM: calmodulina
DAG: diacilglicerol
ETA: receptor de tipo A de la endotelina
GDP: guanosin difosfato
GTP: guanosin trifosfato
IL: interleuquina
IP
3
: inositol trifosfato
MAPK: cinasas activadas por mitógenos
NO: óxido nítrico
PDE: fosfodiesterasas
PIP
3
: fosfatidilinositol trifosfato
PKA: proteína cinasa A
PKC: proteína cinasa C
PKG: proteína cinasa dependiente de GMPc
PLA
2
: fosfolipasa A
2
PLC: fosfolipasa C
TGF-β: factor de crecimiento tumoral β
Síntesis Inicial
Las células reconocen, integran y responden a múltiples señales procedentes de su entorno.
En los organismos pluricelulares existen sistemas de comunicación entre las células que componen un tejido, un
órgano o un sistema a n de facilitar un desarrollo armónico y coordinado.
La comunicación intercelular puede ser directa (exige el contacto físico entre las células) o indirecta (mediada por mo-
léculas de señalización o ligandos).
Los segundos mensajeros (producidos luego de la activación del receptor) desempeñan un papel crítico en la señalización
intracelular, brindando amplicación, especicidad y diversidad a la respuesta celular.
CONCEPTOS BÁSICOS
Los organismos unicelulares, como bacterias y protozoos, se
adaptan al medio que los rodea desarrollando diferentes me-
canismos para adquirir y aprovechar los nutrientes de este.
Sin embargo, la evolución de los organismos pluricelulares
exigió el desarrollo de sistemas de “comunicación” entre las
diferentes células y tejidos que los componen, de modo de
actuar en forma coordinada y armónica entre ellos. Todas
las células son capaces de recibir y procesar información. Es
posible distinguir distintos tipos de comunicación interce-
lular.

Receptores y mecanismos de transducción de señales 71
Comunicación intercelular directa
Tiene lugar únicamente entre células adyacentes cuyas mem-
branas plasmáticas están en contacto. Dentro de este tipo de
comunicación es posible distinguir diferentes formas:
1
Uniones tipo hendidura o gap
Están formadas por estructuras proteicas denominadas co-
nexones que se ensamblan y crean canales entre las membra-
nas de dos células en contacto, logrando que ambas se acoplen
metabólica y eléctricamente. Permiten el pasaje de iones inor-
gánicos y pequeñas moléculas (menores de ~1200 Da), como
Ca
+2
y AMPc, desde el citoplasma de una célula al citoplas-
ma de la célula adyacente. Desempeñan un papel crítico en
el músculo cardíaco y en el músculo liso. Los cardiomiocitos
auriculares y ventriculares se comunican directamente a través
de uniones de este tipo ubicadas en los discos intercalares y
permiten que los potenciales de acción generados en el siste-
ma de conducción se distribuyan rápidamente en el miocar-
dio para lograr una contracción coordinada.
Uniones adherentes
Están formadas por los dominios extracelulares de ciertas
proteínas transmembrana denominadas caderinas que inte-
raccionan en forma dependiente de Ca
+2
; se agrupan entre
ellas y con otras proteínas intracelulares, las cateninas. A su
vez, las cateninas están unidas a la actina del citoesqueleto.
Son importantes para mantener la morfología celular normal
como también la organización de grupos celulares dentro de
un tejido y participan en la señalización durante el desarrollo
o remodelado de los tejidos. Dentro de este tipo de uniones
también debemos considerar a las uniones estrechas, forma-
das por otras proteínas transmembrana, como las claudinas
y ocludinas. Trastornos en la permeabilidad al cloruro en
uniones estrechas del epitelio tubular renal podrían estar in-
volucrados en el desarrollo de hipertensión arterial.
Ligandos asociados a membrana
Otra forma de comunicación directa entre células adyacen-
tes es a través de la interacción de un receptor de la membra-
na plasmática de una célula con un ligando que es, a su vez,
parte de la membrana plasmática de la célula adyacente. Este
tipo de comunicación brinda, por ejemplo, información es-
pacial a células que migran, como en el caso de las células del
endotelio vascular.
Comunicación intercelular indirecta
Dos (o más) células pueden comunicarse entre sí sin estar en
contacto directo utilizando diferentes moléculas que funcio-
nan como señales químicas entre la célula que la produce y la
célula diana. Las moléculas señales (ligandos) pueden actuar
de tres formas diferentes (g.16-1):
• Autocrina: la molécula señal actúa sobre receptores ubi-
cados en la membrana de la célula que la produjo. Este
mecanismo permite la autorregulación de las funciones
celulares.
• Paracrina: la molécula señal es liberada al líquido intersticial
y difunde a células vecinas, modicando su función.
• Endocrina: la molécula señal, en este caso denominada
hormona, llega al torrente sanguíneo para así alcanzar cé-
lulas alejadas del sitio de producción. La selectividad está
dada por la presencia de receptores especícos para estas
moléculas en la célula diana. Si las células son neuronas
que vierten su neurotransmisor a la sangre se llama meca-
nismo neuroendócrino.
En la señalización autócrina y parácrina, para que la mo-
lécula señal tenga efecto local, su difusión debe estar restrin-
gida, ya sea porque es rápidamente endocitada, porque es
degradada por enzimas extracelulares o porque queda atra-
pada por la matriz extracelular. Un ejemplo típico de seña-
lización parácrina ocurre a nivel de la unión neuromuscular
al liberarse acetilcolina del terminal axónico que interactúa
con el receptor nicotínico de la bra muscular. La acetilcoli-
na libre de la hendidura sináptica es rápidamente degradada
por la acetilcolinesterasa.
Ligandos
Las moléculas que actúan como señales intercelulares pue-
den ser de naturaleza química variada: péptidos y proteínas,
como la angiotensina II, la endotelina y la insulina; este-
roides, como la aldosterona y los estrógenos; aminas, como
la adrenalina y la dopamina y pequeñas moléculas, como
Figura 16-1. Las tres formas de actuar de las moléculas señales
(ligandos). Autocrino, paracrino, endocrino.
AUTOCRINO
ENDÓCRINO
Hormonas
PARACRINO
Vaso
sanguíneo
Formas de acción de las moléculas seales (ligandos)

72 Cardiología
aminoácidos, nucleótidos, iones y gases (por ej.,óxido nítri-
co). Una vez liberada, la molécula señal debe interactuar con
receptores especícos localizados en la membrana plasmática
o en el interior de la célula diana.
Receptores
Son generalmente proteínas, aunque a veces pueden ser lipo-
proteínas. Si bien en algunos casos el receptor es directamen-
te el efector de la respuesta, la mayoría de las veces la interac-
ción del ligando con su receptor conduce a la activación de
moléculas efectoras del medio intracelular responsables de
iniciar la respuesta. En este caso. la transmisión de la infor-
mación generalmente consta los siguientes pasos:
• Paso 1: Reconocimiento de la molécula señal con su re-
ceptor. La misma molécula puede unirse a más de un tipo
de receptor. A cuál se una dependerá de la anidad del re-
ceptor por el ligando, de la abundancia de cada tipo de
receptor y de si existe algún otro agonista que compita por
el receptor. La unión ligando-receptor, desde el punto de
vista químico, puede ser: iónica, formada por 2 grupos de
cargas opuestas; interacción de tipo van der Waals, un di-
polo transitorio en un átomo genera un dipolo en el átomo
adyacente, lo que crea una interacción electroestática e in-
teracción hidrofóbica, que ocurre entre grupos no polares.
• Paso 2: Transducción del mensaje extracelular en una se-
ñal intracelular o un segundo mensajero. La unión del
ligando al receptor causa un cambio conformacional en
este último, que inicia la actividad catalítica intrínseca del
receptor o causa que el receptor interactúe con alguna en-
zima de la membrana o del citosol. La consecuencia nal
es la generación de un segundo mensajero o la activación
de una cascada catalítica de señalización intracelular.
• Paso 3: Transmisión de la señal del segundo mensajero al
efector apropiado, que puede ser una enzima, un factor
de transcripción, un canal iónico, etc.
• Paso 4: Modulación del efector: las cascadas de señaliza-
ción, en general, activan proteínas cinasas (enzimas que
transeren grupos fosfato a diferentes residuos de las pro-
teínas) y/o fosfatasas (enzimas que desfosforilan proteí-
nas), alterando la actividad de sus sustratos.
• Paso 5: Respuesta de la célula al estímulo original. La
respuesta nal dependerá de la integración y sumación de
múltiples señales intracelulares que están activas o inacti-
vas en el mismo momento.
• Paso 6: Terminación de la respuesta por mecanismos de
control en alguno de los niveles de la vía de señalización.
TIPOS DE RECEPTORES
Los receptores pueden dividirse en 4 categorías de acuerdo a
su ubicación y al mecanismo de transducción de señales que
tienen asociado (tabla 16-1).
3
Tabla 16-1 Tipos de receptores
Clase de Receptor Estructura del receptor Ligandos
Canales iónicos activados por
ligandos (receptores ionotrópicos)
Homo o Heteroméricas GABA
Glicina
ACh: músculo y nervio
5-HT
Glutamato
ATP (apertura)
Receptores acoplados a Proteínas
G, heterotrímeros (αβγ)
Una cadena polipeptídica con 7
segmentos transmembrana.
Pequeñas moléculas transmisoras
ACh
Norepinefrina
Péptidos
Oxitocina
Hormona paratiroidea
Neuropéptido Y
Gastrina
Colecistoquinina
Ciertas citoquinas, lípidos y moléculas relacionadas
Receptores Catalíticos Cadenas polipeptídicas simples
que cruzan la membrana 1 vez.
Pueden ser dímeros o dimerizarse
luego de la activación.
ANP, TGF-β
EGF, PDGF, FGF, insulina, IGF-1,IL-3,
IL-5, IL-6, EPO, LIF, CNTF, GH, IFN-α, IFN-β, IFN-γ, GM-CSF, CD45
Receptores intracelulares o
Nucleares
Homodímeros cada uno con
múltiples dominios funcionales
Heterodímeros cada uno con
múltiples dominios funcionales
Hormonas Esteroideas: Mineralocorticoides, Glucocorticoides,
Andrógenos, Estrógenos, Progestágenos
Otros:
Hormonas Tiroideas, Acido Retinoico, Vitamina D,
Prostaglandinas
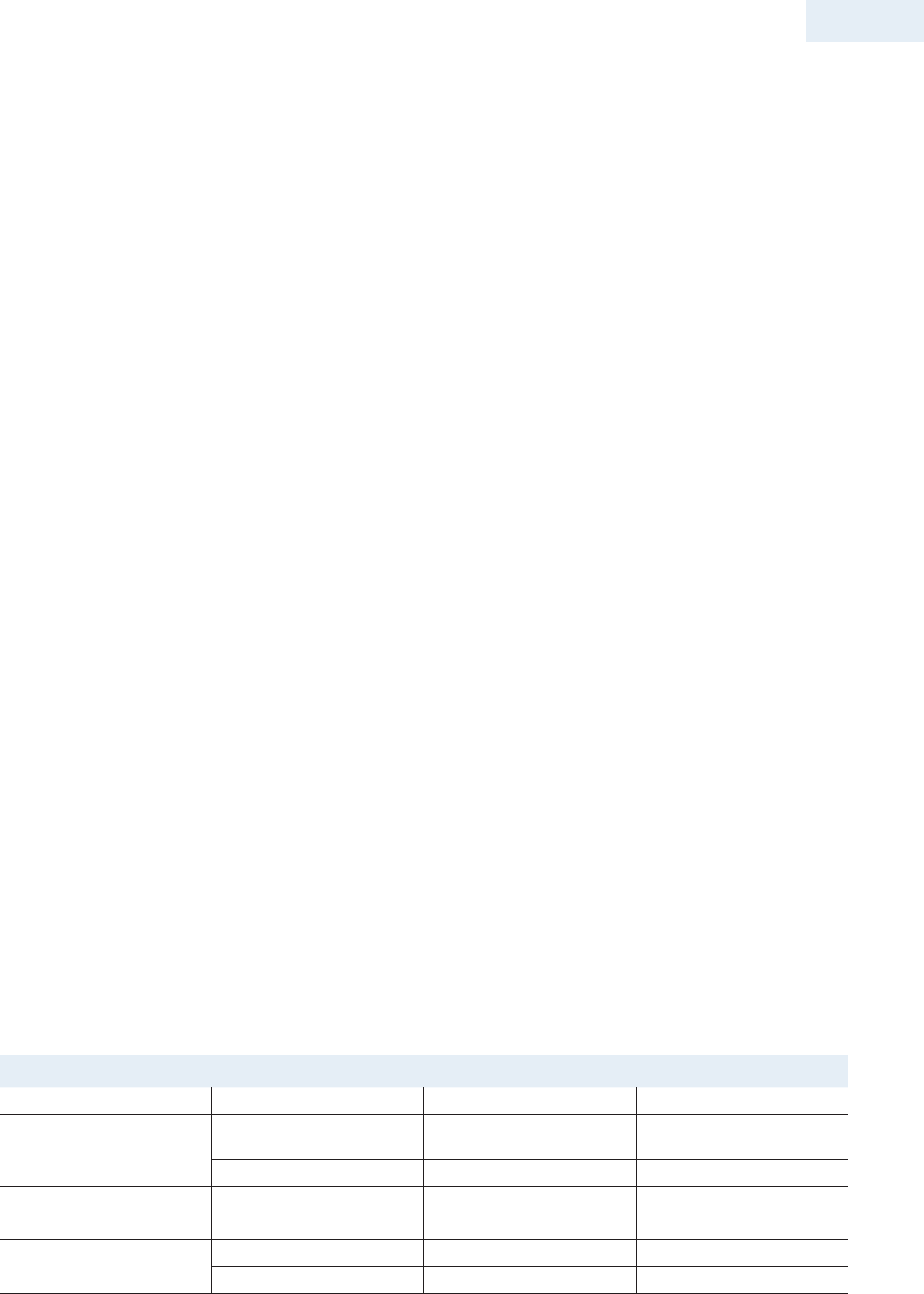
Receptores y mecanismos de transducción de señales 73
Receptores ionotrópicos
(canales iónicos activados por ligando)
Son proteínas integrales de membrana que participan en la
señalización de las células excitables. La unión de la molé-
cula de señalización a su receptor, que es parte de un canal
iónico, produce la apertura transitoria del canal, lo que al-
tera la permeabilidad de la membrana al ión, y se produce
la traducción de una señal química en eléctrica. El ejemplo
más conocido de esta clase de receptores ionotrópicos es el
receptor nicotínico de la placa neuromuscular, que ya fue
mencionado. Este receptor es un canal catiónico formado
por dos subunidades α, una subunidad β, una γ y una δ.
Cuando la acetilcolina se une al receptor nicotínico este
cambia de conformación y permite el ingreso de cationes
(fundamentalmente sodio) al interior celular y despolariza la
membrana. Su denominación se debe a que la nicotina del
tabaco actúa como ligando de este receptor, induciendo su
apertura y alterando el potencial de membrana.
Receptores asociados a proteína G
Constituyen la familia más numerosa de receptores de mem-
brana, con más de 1000 miembros descritos hasta la actualidad,
aunque para muchos de ellos aún se desconocen sus agonistas
(“receptores huérfanos”). Si bien se activan por una diversidad
de ligandos, como hormonas, neurotransmisores, mediadores
de inamación, péptidos vasoactivos, etc., la mayoría de estos
receptores comparten una estructura similar. Están compues-
tos por una única cadena polipeptídica que atraviesa siete veces
la membrana (siete dominios transmembrana), un dominio
amino-terminal extracelular y un dominio carboxilo-terminal
hidrofílico intracelular. No tienen actividad catalítica intrínseca
sino que funcionan indirectamente a través de un intermedia-
rio que activa o inactiva canales iónicos o enzimas asociados
a la membrana.
4
Este intermediario es una proteína con un
sitio de unión de alta anidad para GDP/GTP, denominada
proteína G. Las proteínas G se dividen en 2 grandes subfami-
lias: las pequeñas proteínas G (monoméricas) y las proteínas G
heterotriméricas.
5,7
Ambos tipos hidrolizan el GTP (es decir,
poseen actividad GTPasa). Su forma activa es la unida a GTP,
mientras que cuando esa molécula es hidrolizada, la proteína G
pasa al estado inactivo. Las proteínas G heterodiméricas son las
intermediarias entre los receptores acoplados a proteína G y sus
diversos efectores intracelulares, como la AC y la PLC. Están
compuestas por una subunidad α, una β y una γ. Existen al
menos 16 isoformas diferentes de subunidad α, 5 de la β y 11
de la γ en tejidos de mamíferos. Las diferentes isoformas adop-
tan una distinta distribución tisular y se unen a distintos tipos
de receptores y efectores. (tabla 16-2) La subunidad α es la que
une e hidroliza el GTP. Cuando el ligando se une al receptor se
produce un cambio conformacional que facilita el intercambio
de GDP por GTP, disociándose la subunidad α de las βγ. Tanto
la subunidad α activada (unida a GTP) como la subunidades
βγ libres pueden interactuar con uno o más efectores y generar
segundos mensajeros que participan de la cascada de señali-
zación intracelular. Cuando la subunidad α hidroliza el GTP
vuelve a asociarse a subunidades βγ. Las proteínas G pequeñas
están estructuralmente relacionadas con la subunidad α de las
G heterotriméricas; pueden estar asociadas a la membrana (por
ej., Ras) o desplazarse entre estas y el citosol (por ej., Rho).
5
Receptores catalíticos
Estas proteínas integrales de membrana son activadas por
la unión de su ligando y son enzimas o parte de complejos
enzimáticos. Muchas hormonas y factores de crecimiento se
unen a receptores de la membrana plasmática que tienen ac-
tividad enzimática en el lado citoplásmico. A continuación
se describen las cinco clases de receptores con actividad cata-
lítica conocidos hasta la actualidad.
Receptores con actividad de guanilato-ciclasa
Son receptores con un domino transmembrana con la capa-
cidad de generar GMPc a partir de GTP. Los receptores de
los péptidos natriuréticos (péptido natriurético atrial, cere-
bral y C) pertenecen a esta clase. Son receptores compuestos
de un único dominio transmembrana que poseen el sitio de
unión al ligando su porción extracelular y en su segmento
intracelular tiene los sitios catalíticos (guanilato ciclasa). La
unión del péptido natriurético induce un cambio de confor-
mación del receptor que provoca la dimerización y activa-
ción del receptor. Se produce entonces GMPc, que activa a
una PKG que fosforila proteínas en ciertos residuos de serina
y treonina. Dentro de este grupo de receptores también se
incluye al receptor del NO, que es una guanilato ciclasa so-
Tabla 16-2 Principales proteínas G
Subfamilia de proteína G
Subunidad α
Efector Receptor asociado
Gs
αs ↑ adenilato ciclasa Receptor β-adrenérgico;
Receptor Glucagón
αolf ↑ adenilato ciclasa
Receptor olfatorios
Gi
αi ↓ adenilato ciclasa M2, Receptor α
2
-adrenérgico
αt ↑ fosfodiesterasa de GMPc
Receptor de rodopsina
Gq
αq ↑ PLC β1, β2, β3 M1, Receptor α
1
-adrenérgico
α11- α16 ↑ PLC β1, β2, β3

74 Cardiología
luble (no asociada a membranas). Esta guanilato ciclasa se
activa en presencia de NO, un gas que difunde fácilmente
a través de la membrana celular y que, por ejemplo, es un
importante modulador del ujo y de la presión sanguínea al
favorecer la relajación del músculo liso vascular.
Receptores con actividad de tirosina quinasa intrínseca
Estos receptores, en general, están constituidos por una pro-
teína que atraviesa una sola vez la membrana y posee en su
fragmento citosólico la actividad catalítica. Cuando se une el li-
gando a la porción extracelular del receptor se produce un cam-
bio conformacional que facilita su dimerización y activación,
fosforilándose mutuamente ambos monómeros y permitiendo
la incorporación de fosfato a residuos de tirosina de otros efec-
tores citosólicos. Los residuos de tirosina fosforilados unen con
gran anidad otras moléculas intracelulares. Pertenecen a esta
clase los receptores de varios factores de crecimiento (factor de
crecimiento epidérmico, factor de crecimiento derivado de pla-
quetas, factor de crecimiento broblástico, etc.), el de insulina
y el del péptido similar a insulina. Estos dos últimos existen en
forma de dímero aun en ausencia del ligando.
Receptores asociados a tirosina cinasa
El receptor no posee actividad catálitica intrínseca, pero se
une en forma no covalente a diferentes tirosinas cinasas (Src
cinasa o la familia de Janus cinasas). Los receptores de varias
citoquinas (IL-2, IL-3, IL-4, IL-5, etc.), el de la hormona de
crecimiento, de la prolactina y otros pertenecen a esta clase.
Receptores con actividad de serina-treonina cinasa
La unión del ligando al receptor induce la fosforilación de
residuos serina o treonina del mismo receptor. Diferentes fac-
tores que controlan el crecimiento y la diferenciación celular
poseen este tipo de receptores, como por ejemplo, el receptor
del TGF-β. Este receptor es una glicoproteína con un único
dominio transmembrana y actividad catalítica intrínseca. El
ligando se une inicialmente al receptor de de tipo II, lo que
induce la formación de un complejo ternario (receptor de tipo
I, de tipo II y ligando) catalíticamente activo.
Receptores con actividad de tirosina fosfatasa
Desfosforilan residuos de tirosina de proteínas asociadas al
receptor (por ej., receptor CD-45 de la membrana de linfo-
citos T y B).
Receptores intracelulares
Son receptores proteicos localizados en el citosol o el núcleo,
capaces de relacionar señales extracelulares con la transcrip-
ción génica. Las hormonas tiroideas y esteroideas, como el
cortisol, las hormonas sexuales y la vitamina D atraviesan la
membrana plasmática e interactúan con sus receptores intra-
celulares que funcionan como factores de transcripción acti-
vados por ligando, y estimulan o reprimen la transcripción
de determinados genes.
Segundos mensajeros de los receptores acoplados a
proteínas G
Los segundos mensajeros son moléculas que permiten am-
plicar a nivel intracelular la señal recibida. La unión de un
ligando al receptor puede generar cientos de moléculas de
segundos mensajeros que, a su vez, pueden modicar a mi-
les de moléculas efectoras. Para que una molécula funcione
como segundo mensajero su concentración o ventana de ac-
tividad deben estar namente reguladas; deben producirse
muy rápidamente frente a la interacción ligando-receptor y
luego destruirse o inactivarse también de forma muy veloz.
Los segundos mensajeros permiten además especicidad y
diversidad de respuestas. Un mismo ligando produce efectos
diferentes, e incluso antagónicos, según el tipo de receptor
al que se una y al segundo mensajero que se produzca en
esa vía de señalización. Por ejemplo, la adrenalina produce
contracción de músculo liso vascular, lo que activa recep-
tores α
1
adrenérgicos (los segundos mensajeros de esta vía
son el IP3 y el DAG), favorece la relajación del músculo liso
bronquial a través de la activación de receptores β
2
adrenér-
gicos (el segundo mensajero es el AMP cíclico) y estimula
la degradación de triglicéridos en los hepatocitos por medio
de su unión a receptores β
1
adrenérgicos (segundo mensaje-
ro AMP cíclico). Además, distintos ligandos interactuando
con sus receptores especícos que estimulan una misma vía
de señalización van a producir un mismo efecto. Por ejem-
plo, tanto la angiotensina II como la adrenalina y la endo-
telina-1, al unirse a sus receptores (AT1, α
1
adrenérgicos y
ETA, respectivamente), estimulan la PLC, lo que aumenta la
producción de PIP3 y DAG y, de forma consecuente, la con-
tracción del músculo liso vascular. Existe además un amplio
grado de entrecruzamiento entre las distintas vías de señali-
zación intracelular que permite la generación de respuestas
integradas frente a distintos estímulos.
Entre los segundos mensajeros más relevantes desde el
punto de vista siopatológico describiremos los siguientes:
AMP cíclico
La activación de los receptores asociados a proteínas GS re-
sulta en la activación de la enzima AC, que sintetiza AMPc a
partir de ATP. El AMPc puede directamente inducir el efec-
to nal (como sucede en el caso de los receptores olfativos
en los que el AMPc directamente induce la apertura de un
canal catiónico) o activar a la PKA. La PKA cataliza la trans-
ferencia de un grupo fosfato del ATP a residuos especícos
de serina y treonina de determinadas proteínas, modicando
su localización y/o actividad.
La activación de los receptores acoplados a proteínas Gi,
como los receptores α
2
adrenérgicos, actúan inhibiendo a
la AC y, por lo tanto, disminuyen la producción de AMPc.

Receptores y mecanismos de transducción de señales 75
Metabolitos del ácido araquidónico
Los receptores asociados a proteínas G pertenecientes a la
familia Gi/Go, cuando son activados por la unión de su
agonista, estimulan la PLA2 a través de un mecanismo que
parece no involucrar a la subunidad α. El dímero βγ puede
activar directamente o indirectamente a la PLA2. El AA li-
berado por la hidrólisis de lípidos de membrana funciona
como segundo mensajero.
6
Se ha descrito una PLA2 citosó-
lica y especíca de la fosfatidilcolina (PLA2c) que origina al
AA de señalización y otra secretora (PLA2s) que da origen al
AA involucrado en la inamación.
Productos derivados de la ruptura de los fosfoinosítidos
de la membrana
La activación de los receptores asociados a proteínas Gq re-
sulta en la estimulación de la isoforma β de la PLC (PLCβ),
que hidroliza a un fosfolípido de la membrana (fosfatidilino-
sitol 4,5-bifosfato) y forma dos segundos mensajeros: el IP3
y el DAG. El IP3 es hidrosoluble y, al unirse a su receptor
(un canal de Ca
2+
activado por ligando) ubicado en la mem-
brana del retículo endoplásmico y posiblemente también de
la envoltura nuclear, facilita la liberación de Ca
2+
. El DAG es
lipídico, está anclado en la membrana y produce, junto con
el Ca
2+
, la activación de algunas isoformas de PKC.
Complejo Ca
2+
-Calmodulina (CaM)
El Ca
2+
actúa como segundo mensajero en distintas vías de
señalización intracelular. Muchos procesos celulares están
regulados por los niveles de Ca
2+
citosólicos. Uno de los me-
canismos a través del cual el Ca
2+
ejerce su control es unirse a
la CaM. El complejo Ca
2+
CaM se une a diferentes proteínas
e inuye en su actividad, entre ellas un grupo de cinasas de-
nominadas Ca
2+
CaM-cinasas, como la cinasa de la cadena
liviana de miosina del músculo liso, la Ca
2+
CaM cinasa II y
la fosforilasa cinasa.
Vía de las cinasas activadas por mitógenos
Las MAPK son una familia de serina-treonina cinasas impor-
tantes en la regulación de procesos celulares como el creci-
miento, la diferenciación, la expresión génica y la apoptosis.
Las MAPK son activadas por distintas vías de señalización
desencadenadas por factores de crecimiento, citoquinas,
neurotransmisores, hormonas o distintos agonistas cuyos
receptores están acoplados a proteína G o a tirosinas cina-
sas. En los mamíferos hay varias subfamilias de MAPK que
suelen agruparse en 2 clases: las convencionales (ERK1/2,
la JNK y la p38-MAPK) y las atípicas (ERK3/4, ERK5 (o
BMK), ERK7 y ERK8). Dentro de los blancos de las MAPK
se encuentran factores de transcripción importantes en la ex-
presión génica cardíaca, como el NFAT, GATA-4 y AP-1.
2
Regulación o nalización de las señales
Fosfodiesterasas
Las PDE son enzimas capaces de hidrolizar a los nucleóti-
dos cíclicos. Existen 11 isoenzimas de PDE (PDE1-11) de
acuerdo a la especicidad por el nucleótido que posean. Fi-
nalizan las señales del AMPc y el GMPc al convertirlos en
su metabolitos lineales, por ello regulan diversas respuestas
siológicas.
Fosfatasas
El balance entre la actividad de cinasas y fosfatasas es quien
determina la actividad nal de las proteínas reguladas.
Serina-treonina fosfatasas (PP)
Son una gran familia de fosfatasas estructuralmente relacio-
nadas, se clasican en tipo 1 (PP1) y 2 (PP2, existen PP2A,
PP2B o calcineurina y PP2C).
Fofotirosina fosfatasas (PTP)
Familia de enzimas que desfosforilan residuos tirosina y re-
gulan el estado de fosforilación de importantes moléculas
de señalización, como las MAPK. Están involucradas en la
regulación del crecimiento y la diferenciación celular.
Bibliografía sugerida
1. Cantley L. Signal Transduction. En: Boron WF, Boulpaep EL, eds.
Medical Physiology. 2º ed. Philadelphia: Saunders Elsevier 2009: 48-
74.
2. Chang L, Karin M. Mammalian map kinase signalling cascades. Na-
ture. 2001; 410: 37-40.
3. Harnett MM y Goodridge HS. Receptores de membrane y transduc-
ción de la Señal. En: Baynes J y Dominiczak M, eds. Bioquímica Mé-
dica. 3º ed. Elsevier España; 2011: 551-567.
4. Lefkowitz RJ. Seven transmembrane receptors: something old, some-
thing new. Acta Physiol (Oxford, England) 2007; 190: 9-19.
5. Rozengurt E. Mitogenic signaling pathways induced by G protein-
coupled receptors. J Cell Physiology 2007; 213: 589-602.
6. Jenkins CM, Cedars A, Gross RW. Eicosanoid signalling pathways in
the heart. Cardiovasc Res. 2009 May 1;82(2):240-9.
7. Takai Y, Sasaki T, Matozaki T. Small GTP-binding proteins. Physiol
Rev 2001; 81: 153-208.

capitulo resumen señalización celular.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.