
SC 9.1. El cierre del tubo neural. V. Flores
Luego de su formación (determinación y diferenciación parcial) y
modelación, la placa neural, debido a interacciones con tejidos
adyacentes con los cuales contacta, se transforma en tubo neural. El
proceso global consta de varios fenómenos: a)generación de un
surco medial, b) sobreelevación de los bordes de la placa y
formación de los pliegues o labios del surco neural, c) acercamiento
de los pliegues neurales a la línea media, d) fusión de los pliegues
en la línea media, e) segregación de las tres poblaciones celulares
que integran los labios del surco neural (ectodermo neural, cresta
neural y ectodermo epidérmico), f) fusión de los lados derecho e
izquierdo de la placa neural y del ectodermo epidérmico, g) migración
de las células de la cresta neural y h) recomposición y estabilización
de los epitelios.
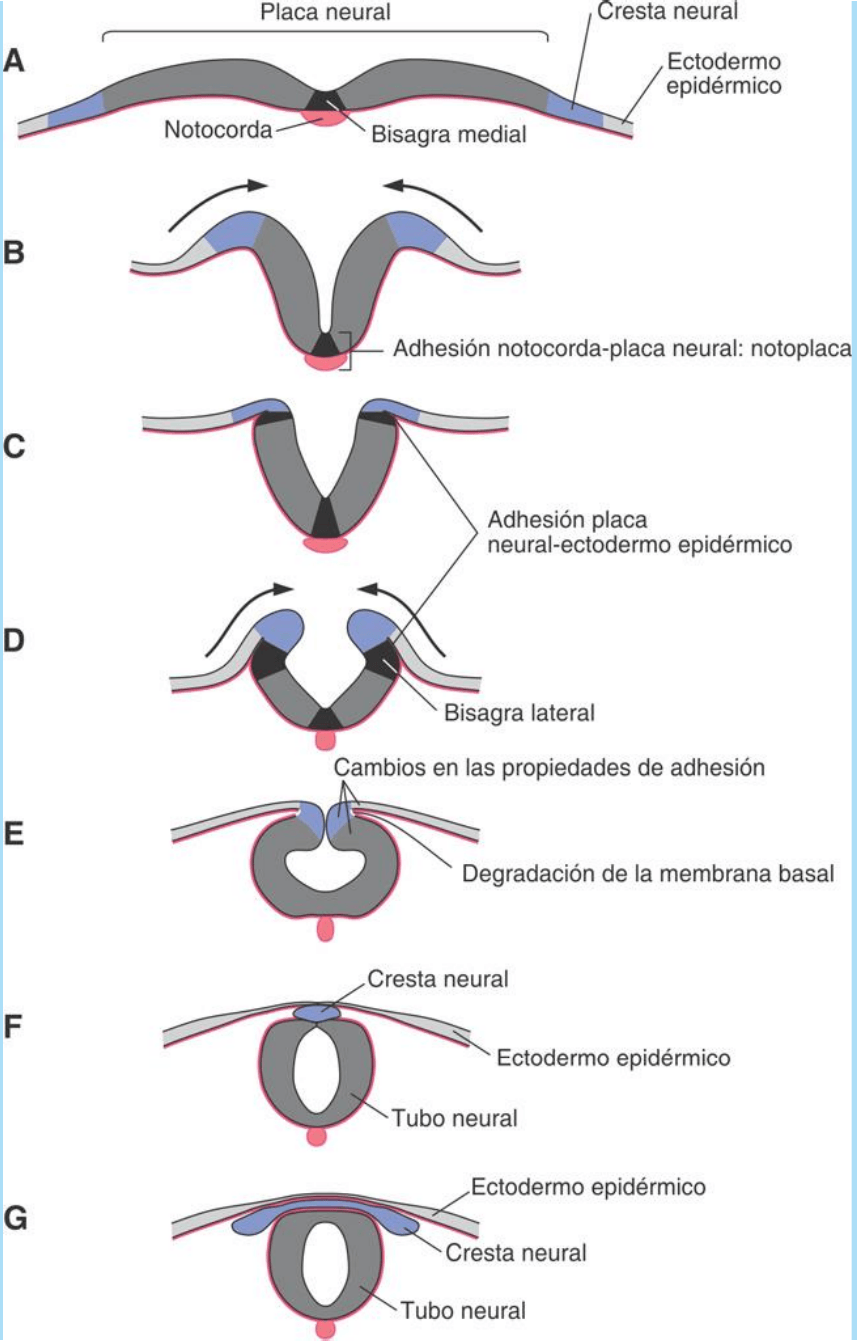
- Al principio, la placa neural es plana (Fig. SC 9-1-1 A). La formación
del tubo neural se inicia con la formación del surco medial en la placa
neural. Este fenómeno depende principalmente del comportamiento
de células que ocupan la línea media de la placa neural. En la región
cefálica de la placa neural –la zona que interactúa con el mesodermo
precordal y que dará origen al prosencéfalo–, las células mediales se
originan directamente del ectodermo neural. En la región caudal –la
que interactúa con la notocorda y que originará al cerebro posterior y
la médula– las células mediales provienen del nódulo de Hensen.
Éste genera tanto las células de la notocorda como también las de la
placa del piso. Las células mediales, independientemente de su
origen, sufren cambios de forma promovidos por el mesodermo
subyacente, de cilíndricas pasan a ser cónicas o piramidales truncas
y, como consecuencia, la placa se pliega generándose un surco
medial, el surco neural. El sitio en que se genera el surco neural
opera como eje de giro de las mitades derecha e izquierda de la placa
neural; debido a ello se denomina “bisagra medial” (Fig. SC 9-1-1 A-
B).
- Los bordes de la placa neural –la zona de transición entre ésta y el
ectodermo epidérmico– se encuentran ocupados por una población
celular determinada a formar la cresta neural. Tres conjuntos de
fuerzas, actuando simultáneamente, podrían producir la elevación
hacia el dorso de los bordes del surco neural (Fig. SC 9-1.1 B): 1) el
efecto de la bisagra medial eleva los bordes de la placa neural hacia
el dorso, 2) la notocorda y la placa del piso se encuentran
fuertemente adheridas y al conjunto se lo denomina “notoplaca” (Fig.
SC 9-1-1 B). Ésta, en el momento en que empiezan a sobreelevarse
los pliegues neurales, sufre un proceso de rápida elongación.
Durante dicho proceso, la notoplaca es la estructura más rígida del

embrión. Las regiones no mediales de la placa neural, por el
contrario, exhiben un comportamiento elástico. La notoplaca realiza
una fuerza de tracción que genera una línea de tensión generalizada
a lo largo de la línea media y las regiones laterales de la placa, debido
a su comportamiento elástico, descomponen dicha fuerza a lo largo
de arcos o curvas orientadas perpendicularmente a la línea de
tensión medial. La placa neural adopta la forma de dichos arcos y se
pliega. Este efecto puede ser apreciado por una experiencia simple:
se sugiere al lector que tense una lámina elástica generando una
tracción sobre ella. A lo largo de la línea de tensión se generará un
surco (corresponde al surco medial de la placa); lateralmente a éste
se formarán dos pliegues (corresponden a los pliegues neurales); 3)
debido a la acción de estas dos fuerzas, los bordes de la placa se
elevan, dejan de estar en contacto con el mesodermo paraxil y ello
permite que la superficie basal de la placa neural realice
interacciones de adhesión directa con la superficie basal del
ectodermo epidérmico (Fig. SC 9-1-1 C-D). Estas interacciones
generan fuerzas de adhesión intersuperficiales que contribuyen a
acentuar los pliegues neurales que entonces sobresalen hacia el
dorso. Si se cultivan pequeños trozos del borde de la placa neural
que contienen la zona de transición y el ectodermo epidérmico,
ambos se adosan fuertemente y forman pliegues similares a los
pliegues neurales.
- El acercamiento de los pliegues neurales hacia la línea media podría
ser consecuencia de la operación de tres conjuntos de fuerza: 1) el
incremento en la adhesión entre las dos hojas que forman cada
pliegue neural hace que aumente la superficie de contacto entre
ambas y que los bordes libres de cada pliegue se curven hacia la
línea media (Fig. SC 9-1-1 D); 2) en las regiones laterales de la placa,
promovidas por el contacto con el ectodermo epidérmico, se generan,
a cada lado, cambios de forma celular que producen el mismo efecto
descrito en la bisagra medial. Estas dos regiones se denominan
“bisagras laterales” (Fig. SC 9-1-1 D); ellas generan fuerzas que
curvan el borde libre del pliegue hacia la línea media; 3) los dos
efectos descritos sólo son efectivos porque el surco medial de la
placa se mantiene fuertemente unido en profundidad a la notocorda
que sigue operando como elemento rígido (Fig. SC 9-1-1 D-G). La
interacción con la notocorda hace que la línea media de la placa
quede adherida profundamente en la línea media ventral;
simultáneamente las fuerzas de adhesión entre las superficies
basales de la placa neural y del ectodermo epidérmico y las bisagras
laterales hacen que los pliegues se aproximen a la línea media dorsal
(Fig. SC 9-1-1 D-E). Por otro lado, estos efectos son posibles debido

que el resto del ectodermo epidérmico en este momento se comporta
como un material viscoso, se acomoda a las tracciones generadas
sobre toda la superficie corporal, se distiende fácilmente y las
tracciones desaparecen.
- Una vez que los labios del surco neural contactan en la línea media,
ellos se adosan fuertemente (Fig. SC 9-1-1 E-F). En la zona de
contacto, el epitelio se desestabiliza debido a cambios en las
propiedades de adhesión de las tres poblaciones celulares que se
localizan en los bordes que contactan (células de la placa neural, de
la cresta neural y del ectodermo epidérmico). Mientras las células de
la placa neural expresan E-cadherinas, su continuidad con el
ectodermo epidérmico se mantiene estabilizada. A medida que los
bordes de los pliegues neurales se aproximan hacia la línea media,
las células de la placa neural cambian sus propiedades de adhesión.
Cesa la síntesis de E-cadherinas e inician la expresión de N-
cadherinas y N-CAM. Esto desestabiliza la zona de continuidad de
los epitelios y se produce un fenómeno de segregación y
reagregación por adhesividad diferencial. De esa forma, las células
de los bordes derecho e izquierdo de la placa neural se desprenden
del ectodermo epidérmico y se fusionan entre sí (Fig. SC 9-1-1 F-G).
- Este proceso de desagregación y reagregación de los epitelios está
acompañado de una degradación de la membrana basal. Durante
dicho proceso se pierde transitoriamente, en sitios localizados, la
separación neta entre epitelio y mesénquima; las células de la cresta
neural abandonan entonces el epitelio y se introducen en el
compartimento mesenquimático (Fig. SC 9-1-1 F). Una vez que
migran las células de la cresta neural se reconstruye, en la línea
media dorsal, la continuidad entre los bordes derecho e izquierdo del
ectodermo epidérmico (Fig. SC 9-1-1 F-G). De esta forma, el
ectodermo epidérmico cubre la superficie dorsal (este ectodermo
genera señales que tienen importante influencia en el desarrollo
ulterior de la cresta neural y el tubo neural) y se constituye el tubo
neural que queda cubierto por el ectodermo. Entre ambos quedan las
células de la cresta neural. En la región cefálica las células de la
cresta neural abandonan el epitelio antes que se cierre el tubo neural,
en la región medular lo hacen recién cuando los labios derecho e
izquierdo se fusionan.
{kind=link}

Fig. SC 9-1-1. Representación esquemática de los diversos cambios
que llevan a la formación del tubo neural a partir de la invaginación
de la placa neural. Durante dicho proceso se forman también la cresta
neural y los segmentos bilaterales de cresta que llevan a la formación
de los ganglios (espinales y craneales) de neuronas sensoriales
primarias.
- Finalmente, las células de la cresta neural migran lateralmente y se
ubican a ambos lados del tubo neural. Mientras tanto, en los somitas
del mesodermo paraxil, que quedan ubicados a ambos lados del tubo
neural, se constituye el dermatomo. Una parte de las células del
dermatomo migran medialmente entre el ectodermo epidérmico y el
tubo neural y generan el mesénquima que contribuye a separarlos.
Al mismo tiempo se refuerzan la membrana basal y la matriz
extracelular subyacente a dichos epitelios. En dicho mesénquima,
más tarde se introducen poblaciones de células del esclerotomo;
éstas contribuyen a generar las cubiertas cartilaginosas y luego
óseas que forman los huesos que delimitan el conducto raquídeo. En
la región craneal dichos tejidos provienen del mesénquima cefálico
que generan las propias células de la cresta neural.
Puede apreciarse que el proceso descrito, aunque morfológicamente
simple, resulta de muchos CCD que actúan en forma integrada en el
tiempo y en el espacio. Los CCD más directamente implicados son el
cambio de forma celular, la adhesividad intercelular diferencial, la
proliferación diferencial y otros. Todos estos CCD dependen, a su
vez, de procesos biomoleculares sincronizados en tiempo y espacio
y que se regulan interactivamente entre las poblaciones celulares
participantes. En teoría, las alteraciones en cualquiera de las
moléculas involucradas en la ejecución o control de los CCD
señalados pueden ocasionar alteraciones de la línea media dorsal
denominadas genéricamente disrafias (véase Capítulo 9, Desarrollo
del sistema nervioso).
SC 9.2. Poblaciones celulares organizadoras (pcO) y la
regionalización y determinación progresiva del tubo neural. V.
Flores
El inicio del desarrollo del sistema nervioso central está marcado por
el efecto determinante ejercido por un conjunto de señales
provenientes del organizador clásicamente denominado primario o
nódulo de Hensen. Este efecto consiste en la determinación en
sentido neural o “neuralización” y no implica, al parecer, ninguna
especificación particular adicional (como por ejemplo regiones o
categorías o tipos celulares básicos). Este fenómeno se produce
antes de la gastrulación; cuando las células de la futura notocorda

aún se encuentran en la hoja dorsal del embrión. Vale decir, es un
efecto de señalización por medio de señales que difunden en el plano
del epitelio dorsal del embrión o epiblasto. Se ha estimado que en
algunas especies alrededor del 50% de las células epiblásticas
experimentan neuralización. Las restantes células corresponderían
al ectodermo epidérmico. Sin embargo, una parte de las células que
ocupan la zona de transición entre los ectodermos neural y
epidérmico se determina en sentido de cresta neural.
Con respecto a la organización longitudinal o céfalo-caudal del
tubo neural, el siguiente paso en la determinación progresiva de
regiones corresponde a la especificación de los cerebros anterior, por
un lado, y de cerebro posterior y médula espinal, por otro. En el
principio de la gastrulación, el mesodermo precordal (prolongación
cefálica) migra en sentido cefálico y se ubica por debajo del epiblasto
neuralizado. Dicha región ectodérmica, que ya en el epiblasto
pregastrular se encuentra cefálicamente al nódulo de Hensen,
originará la región cefálica ensanchada de la placa neural. Ella
formará el cerebro anterior o prosencéfalo y deriva en su totalidad del
epiblasto neuralizado. La gastrulación continúa con la regresión del
surco primitivo y, a medida que el nódulo de Hensen se desplaza en
sentido caudal, va dejando una población de células delante de él.
Algunas de ellas quedan en la superficie y forman la línea media de
la placa neural (futura placa del piso). Las otras se invaginan debajo
de las primeras y van formando la notocorda. Esta región de la placa
se determina en cerebro posterior y médula por efecto de señales
provenientes de la notocorda. Durante la gastrulación esta región de
la placa, junto con la notocorda, crecen en sentido caudal. La región
caudal de la placa tiene, en consecuencia, dos orígenes: a) la zona
medial que contacta con la notocorda, junto con ésta, deriva de
células del nódulo de Hensen y b) las zonas laterales de la placa
derivan del epiblasto superficial.
Las especificaciones que siguen, que agregan más detalles a la
organización longitudinal del tubo neural, depende de la aparición de
nuevas pCO.
Las pcO mejor conocidas son la ANR (Anterior Neural Ridge:
engrosamiento neural anterior), la ZLI (Zona
LimitansIntratalámica: zona limitante intratalámica) y el IsO
(Isthmic Organizer: organizador ístmico). Con el tiempo aparecen
nuevas pcO involucradas en la subregionalización de las diferentes
regiones. Aparte de estas existen pcO a lo largo de las zonas
mediales dorsal y ventral del tubo neural que instalan
el patterning dorsoventral del tubo neural.
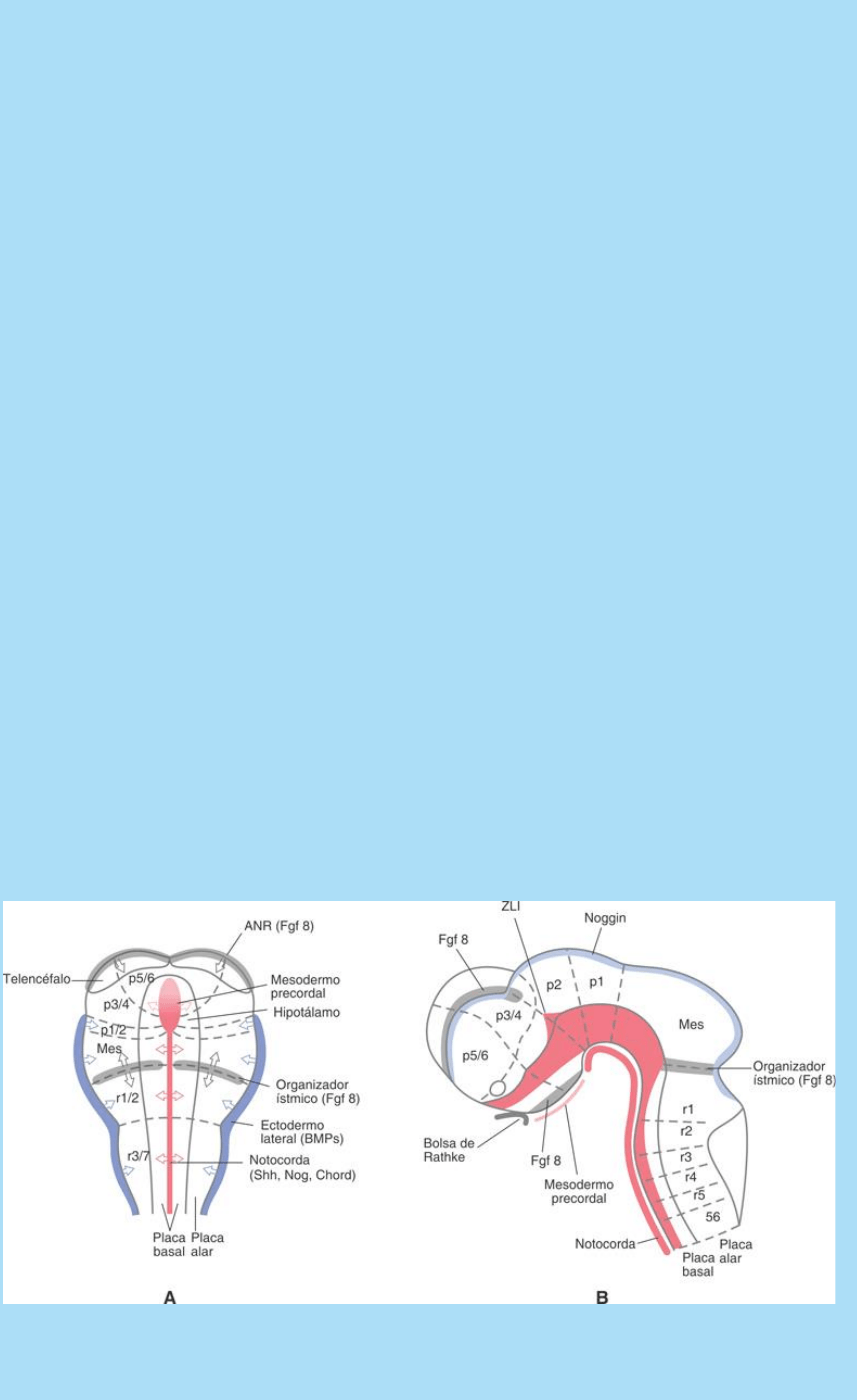
La ANR es una región semilunar engrosada que bordea
cefálicamente a la placa neural, en el límite con el ectodermo
epidérmico. La ANR genera señales que especifican los segmentos
más cefálicos del prosencéfalo (p4-6). La ANR secreta laproteína
señal Fgf-8 en respuesta a la cual las células de la zona más cefálica
de la región prosencefálica expresan laproteína factor de
transcripción Brain Factor 1 (BF1), que participa en al regulación
de los CCD en dicha región.
Hay indicios de que en la región prosencefálica de la placa neural
existen dos zonas con diferente competencia. Las zonas cefálica y
caudal de la región prosencefálica ofrecen diferentes respuestas a
las proteínas señal Fgf-8 y Shh. En el límite entre ambas zonas se
forma la ZLI, que corresponde a la frontera entre los prosómeros 2 y
3 (frontera p2/3). Con respecto a los derivados de estos segmentos
prosencefálicos véase SC La metamerización del SNC.
Más caudalmente se forma otra pcO, el IsO, en el límite entre el
mesencéfalo y el posencéfalo. Esta pcO instala el patterningdel
mesencéfalo y de los dos primeros segmentos posencefálicos (r1 y
r2). El IsO instala dos gradientes decrecientes de morfógeno, uno en
sentido cefálico y otro en sentido caudal. Ambos tienen su máximo
en el IsO. El mesencefálico tiene su mínimo en el límite con el
diencéfalo y organiza los CCD que ejecutan las células del
mesencéfalo. El segundo gradiente posee un rango de alcance que
llega sólo hasta el 2º segmento posencefálico o r2 y organiza los CCD
de las células de dichos segmentos. El patterning de los restantes
segmentos del posencéfalo y de los de la médula depende de la
expresión de un patrón típico de genes Hox que asignan identidad de
segmento.
Fig. SC 9-2-1. El patterning céfalo-caudal del SNC. A. Esquema de
vista dorsal de la placa neural. Se representan las poblaciones
celulares organizadoras que instalan el patterning de varias
{kind=link}

regiones del encéfalo y las moléculas señal que operan como
morfógenos en este proceso. Se ilustran también las posibles
regiones precursoras de diferentes grupos de
neurómeros. B. Esquema de vista lateral de la región encefálica del
embrión. Se representan las regiones del encéfalo, los centros
organizadores, la distribución espacial de señales involucradas en
el patterning y la ubicación de los neurómeros. Este esquema se ha
construido a partir de datos de distribución de morfógenos en el SNC
de embrión de ratón.
Con respecto a la organización dorsoventral del tubo neural,
también existen poblaciones celulares y señales organizadoras que
los especifican y determinan. La organización típica de cada
segmento del sistema nervioso comprende una población de
neuronas periféricas (neuronas sensitivas o sensoriales primarias o
aferentes) y dos poblaciones de neuronas centrales, una de
neuronas de asociación y una de neuronas eferentes (motoras). Esta
organización también se especifica tempranamente durante el inicio
del desarrollo. Mientras se cierra el tubo neural, se genera la
población de células de la cresta neural que originarán, entre otros
tipos celulares, a las neuronas sensoriales primarias de los ganglios
sensitivos de los nervios craneales y raquídeos. Con respecto a las
neuronas intrínsecas del tubo neural, ellas adquieren carácter de
neuronas de asociación o eferentes dependiendo de la posición que
ocupan entre las líneas medias dorsal y ventral del tubo.
El ectodermo epidérmico de la zona medial dorsal y la notocorda, en
la zona medial ventral del tubo, generan señales difusibles que
especifican el carácter asociativo (placa alar) o eferente (placa basal)
de las neuronas del tubo neural. Estasproteínas señal (BMP4,
BMP7, Dorsalin, Sonic hedgehog y otras) operan paracrinamente.
Se distribuyen en forma de gradientes con un máximo en el sitio en
el que son secretadas y un mínimo en la zona opuesta. Poseen
acciones antagónicas y el efecto final sobre las neuronas depende
de la concentración relativa de ambas señales en el sitio en el que
cada neurona se encuentre. Así, el destino de cada neurona, como
alar (asociativa) o basal (eferente), depende de su posición dentro
de un par de gradientes cruzados de señales con efectos
opuestos (Fig. SC 9-2-1 A).
Estas señales proveen un grado de especificidad aún mayor que el
correspondiente a la organización de las neuronas en las placas
alares y basales. Ambas placas tienen organización columnar con
diferentes subtipos de neuronas. Las diferentes combinaciones de
valores de concentraciones relativas de ambas señales, actuando a
través de sus vías de señalización, generarían la expresión de
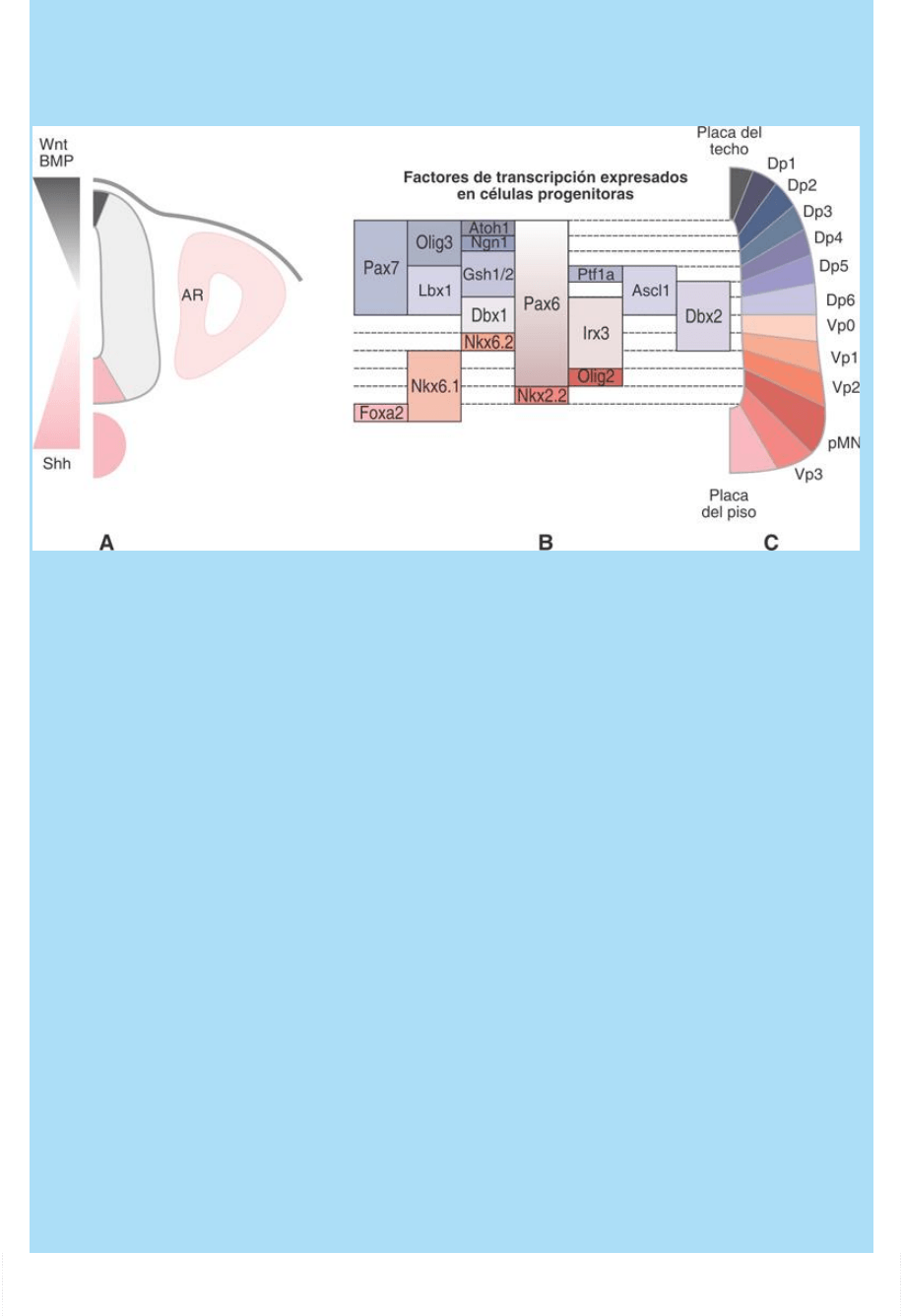
diferentes combinaciones de factores de transcripción. De estas
diferentes combinaciones de factores de transcripción dependería
también la especificación de cada uno de los subtipos de neuronas
de ambas placas (Fig. SC 9-2-2 A-C).
Fig. SC 9-2-2. Representación esquemática
del patterning transversal (dorsoventral) de la médula espinal. A. El
tubo neural recibe señales de poblaciones celulares vecinas
(ectodermo, somitas, notocorda). Un gradiente D
V de señales
originadas en el ectodermo y la placa del techo (Wnt, Bmp y otros)
determina a las células troncales neurales pluripotentes en sentido
alar e instala un proceso de determinación de tipo neuronal
dependiente de la posición en la placa alar. Un gradiente V
D de
señales originadas en la notocorda y la placa del piso (Shh)
determina a las células troncales neurales pluripotentes en sentido
basal e instala un proceso de determinación de tipo neuronal
dependiente de la posición en la placa basal. B. La diferente posición
de las células troncales neurales pluripontes dentro de los gradientes
indicados hace que éstas se hallen sometidas a diferentes
concentraciones de morfógenos que promueven la expresión de
diferentes combinatorias de factores de transcripción a lo largo del
eje dorso-ventral del tubo neural. C. Cada combinatoria de factores
de transcripción especifica un tipo neuronal definido, con una
posición característica dentro de ambas placas. De este modo se
produce un proceso de determinación de tipo neuronal
espacialmente organizado. Dp: cTNP dorsales, Vp: cTNP ventrales,
pMN: cNT precursora de neurona motora, RA: ácido retinoico.
SC 9.3. La metamerización del SNC. V. Flores
{kind=link}

A la regionalización y subregionalización del tubo neural le sigue el
proceso de metamerización. Este fenómeno agrega un mayor grado
de detalle y especialización en la organización céfalo-caudal del
SNC. En el SNC se forma una metámera para cada segmento
corporal. Cada metámera es capaz de formar las categorías
neuronales básicas que inervan una metámera o segmento corporal.
La metamerización consiste en la definición de bloques o poblaciones
celulares con organización simétrica bilateral que se reiteran a lo
largo del eje céfalo-caudal. Aunque son estructuralmente similares,
cada segmento es único y posee identidad. Ésta le es asignada por
la expresión de una combinatoria particular de factores de
transcripción que operan durante el desarrollo embrionario y que
tienen específicamente dicha función. La metamerización acontece
en cada una de las regiones y subregiones definidas a lo largo del
eje céfalo-caudal. La designación genérica para los segmentos del
tubo neural es el de “neurómeros”. La designación genérica
correspondiente a cada región es la de prosómero (segmento
prosencefálico), mesómero (segmento mesencefálico), rombómero
(segmento posencefálico) y medulómero o mielómero (segmento
medular). Sin embargo, dado que cada segmento posee identidad
(es único), cada uno de ellos posee una denominación propia que
está integrada por tres elementos: a) un prefijo que alude a la región
a la que pertenece, b) la raíz “mero” (que alude a metámera) y c) un
número ordinal que alude a su orden de aparición en la región o, lo
que es lo mismo, la ubicación espacial que le corresponde, en su
región, a lo largo del eje céfalo-caudal. El modo como se ordenan
estos tres elementos en la designación de la metámera depende del
idioma que se utilice, pero en cualquier idioma está compuesto por
los tres elementos. Así, por ejemplo, el tercer segmento que aparece
en el prosencéfalo se denomina “prosómero 3º” o “tercer prosómero”.
El concepto de organización segmentaria del sistema nervioso
central es clásico. Tal idea permitió aclarar la existencia de
correspondencias entre: (a) “sitios de origen de las neuronas
motoras” y “grupos musculares que inervan”, y (b)“regiones de
inervación sensorial” y “segmentos medulares”.Tales
correspondencias anatómicas y funcionales resultan del hecho de
que a) durante el desarrollo embrionario, somitas (que forman
músculos esqueléticos y dermis), segmentos del SNC y segmentos
de células de la cresta neural poseen correspondencia espacial muy
precisa y b) ellas se conservan, aunque a veces en forma
enmascarada, en el adulto. Clásicamente, la definición de segmentos
en el tubo neural en desarrollo tuvo bases estructurales; se basó en

que, al igual que los somitas, los rombómeros y los medulómeros son
identificables morfológicamente como poblaciones celulares con
simetría bilateral, repetidas a la lo largo del eje céfalo-caudal. Esta
definición estructural es aplicable sin dificultad en el cerebro posterior
y los segmentos medulares. En las regiones más cefálicas las
manifestaciones morfológicas de organización segmentaria son
menos claras.
Las regiones cefálicas del SNC recibieron clásicamente el nombre de
estructuras suprasegmentarias. Una denominación eminentemente
fisiológica, relacionada con el concepto de nivel de organización
funcional. Alude a que, si bien los segmentos medulares pueden
funcionar autónomamente, respondiendo en forma refleja a los
estímulos provenientes del propio segmento corporal, en general no
funcionan de esa forma sino en forma coordinada e integrada. En
efecto, la mayor parte de los actos voluntarios de la vida son
respuestas complejas que requieren el procesamiento e integración
de información que ingresa en el sistema por múltiples vías. Tal
capacidad de procesamiento, integración y elaboración de
respuestas complejas no radica en los segmentos medulares sino en
las regiones “superiores” del SNC. A dichas regiones se las
denominaba suprasegmentarias debido a que, en el esquema de
organización funcional del sistema, corresponden a un nivel de
organización superior al de segmento.
La identidad de cada segmento y, en consecuencia, el modo de
desarrollo particular de cada uno de ellos está asociado a la
expresión de una combinatoria particular de homeoproteínas factores
de transcripción Hox. Diversos estudios de biología molecular sobre
la distribución espacial de la expresión de estos factores de
transcripción permiten una identificación de los segmentos aun antes
de que la metamerización pueda ser apreciada morfológicamente. La
información proveniente de estos estudios, integrada a estudios
estructurales y de expresión de diversos marcadores, permite la
identificación de 6 prosómeros, el cerebro medio (corresponde a
un mesómero), 7-8 rombómeros y un número de
mielómeros igual al número de segmentos corporales.
Los prosómeros 1 a 3 forman parte del diencéfalo; los prosómeros
4 a 6 forman, en la región ventral, la porción anterior del piso del
hipotálamo y, en la región lateral, originan el telencéfalo.
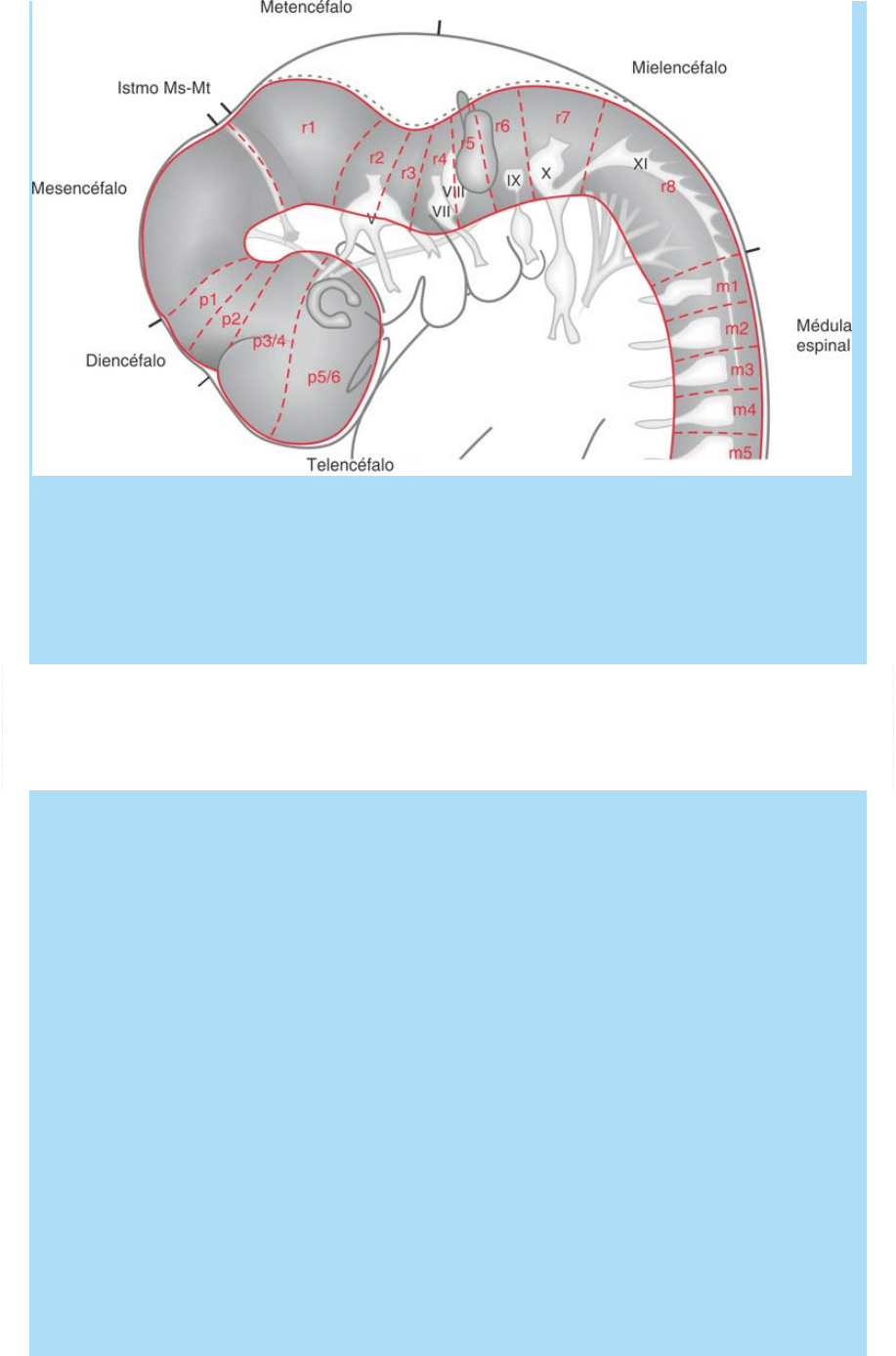
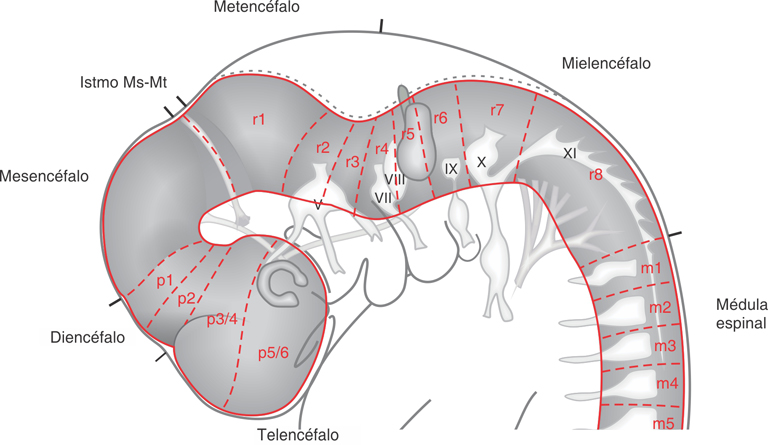
Fig. SC 9-3-1. El patterning del SNC. La regionalización y la
metamerización del tubo neural. Esquema de vista lateral de las
regiones del tubo neural. Se representan las regiones del encéfalo,
sus subregiones (proencéfalo, mesencéfalo y posencéfalo), la
médula espinal y la ubicación de sus neurómeros. p: prosómero; r:
rombómero; m: mielómero.
SC 9.4. Las placas alares y basales. Su función de desarrollo.
Su evolución diferencial en función del espacio (eje céfalo-
caudal). V. Flores
En los mamíferos existen tres categorías principales de neuronas
tanto para el sistema nervioso de la vida de relación como para el de
la vida vegetativa. En el caso del SN de la vida de relación dichas
categorías de neuronas corresponden a a) las neuronas
aferentes, b) las de asociación y c) las eferentes.
a) Las neuronas aferentes o sensoriales primarias corresponden al
SNP y se localizan en su mayor parte en los ganglios de los nervios
craneales y raquídeos. Estas neuronas poseen un soma ubicado, en
general, en un ganglio, una dendrita que conecta con un receptor
periférico y un axón que ingresa en el SNC y que conecta con una
neurona eferente y una o más neuronas de asociación (conexión
monosináptica entre input y output) o conecta sólo con una o más
neuronas de asociación (conexión polisináptica entre input y output).
Esta categoría de neuronas deriva en su mayor parte de la cresta
neural. Un grupo de estas neuronas se genera a partir de una
población celular proliferativa de la línea media dorsal del tubo neural.
b) las neuronas de asociación son intrínsecas del SNC y se ubican
entre la neurona aferente y la eferente. Sus dendritas y axones se
{kind=link}

hallan dentro del SNC y forman todos los circuitos tanto de
proyección como locales que existen en el SNC. Todos los circuitos
neurales intrínsecos del SNC constituyen una gran red de
interconexiones entre neuronas de asociación que se encargan de
recibir información de las neuronas aferentes, procesar e integrar
dicha información y elaborar respuestas que se transmiten a las
neuronas eferentes. Esta categoría de neuronas se genera a partir
de las células neuroepiteliales placas alares del tubo neural.
c) Las neuronas eferentes son neuronas que poseen árbol dendrítico
y soma en el SNC y un axón largo que emerge del SNC y, a través
de un nervio periférico inerva un efector periférico, en general,
muscular. Esta categoría de neuronas se genera a partir de las
células neuroepiteliales de las placas basales del tubo neural.
El número de neuronas y la complejidad estructural de los circuitos
que se hallan a lo largo del eje céfalo-caudal varía en relación con la
riqueza de la información que ingresa en el SNC y con la amplitud del
campo periférico para inervar en cada región. Así, en el caso de la
médula espinal, los engrosamientos cervical y lumbar de donde
nacen los plexos braquial y crural son un ejemplo de mayor riqueza
de inputs y de campos de inervación periféricos más amplios.
Si bien los segmentos medulares tienen cierto grado de
independencia funcional, aparte de la organización segmentaria
(segmentos distribuidos a lo largo del eje céfalo-caudal), el SNC
dispone también de estructuras denominadas clásicamente
suprasegmentarias, formadas por neuronas de asociación o alares,
que se encargan de organizar un comportamiento integrado de los
segmentos (SC 9.2. Poblaciones celulares organizadoras (pcO) y la
regionalización y determinación progresiva del tubo neural).
A lo largo de la evolución filogenética de los cordados se produjo una
concentración de funciones de recepción de estímulos ambientales y
de procesamiento de dicha información en la región cefálica del
embrión y del SNC. Dicho proceso implicó cambios significativos en
los CCD del SNC, especialmente en la intensidad de la actividad
proliferativa de las placas alares y basales. En efecto, el desarrollo
de las neuronas de asociación generadas a partir de las placas alares
se hizo mucho más intenso en las regiones cefálicas que en las
caudales. Este fenómeno posibilitó el desarrollo de grandes
poblaciones de neuronas que forman las estructuras de asociación
de los hemisferios cerebelosos (a partir del posencéfalo) y los
tubérculos cuadrigéminos (a partir del mesencéfalo), y también
posibilitó el desarrollo, desde cefalocordados en adelante, de una
gran estructura exclusivamente alar o asociativa, el telencéfalo. El

desarrollo de la corteza cerebral y de los ganglios de la base
contenidos en los hemisferios cerebrales depende exclusivamente
del desarrollo de neuronas alares. El componente basal del tubo
neural no se extiende en los cordados superiores hasta el extremo
cefálico del tubo neural. Se extiende sólo hasta el mesencéfalo. En
efecto, las neuronas eferentes (derivadas de placas basales) más
cefálicas son las que corresponden al núcleo motor del III par craneal
o motor ocular común. Desde dicho nivel, en sentido cefálico, no
existen poblaciones neuronales eferentes.
Este hecho se explica, desde el punto de vista ontogenético, por el
hecho de que la región más cefálica de la placa neural, la que
corresponde al prosencéfalo o cerebro anterior, se origina
exclusivamente del epiblasto y que no posee en la línea media una
placa del piso originada a partir del nódulo de Hensen.
El nódulo de Hensen origina la notocorda y la placa del piso de la
placa neural caudal al cerebro anterior. Dado que ambas estructuras
–notocorda y placa del piso– están implicadas secuencialmente en la
especificación de las placas basales, su ausencia en la región
prosencefálica se acompaña de ausencia de placas basales y en
consecuencia de neuronas eferentes, en la región del tubo neural
derivada del prosencéfalo (Fig. SC 9-4-1 A-E).
{kind=link}
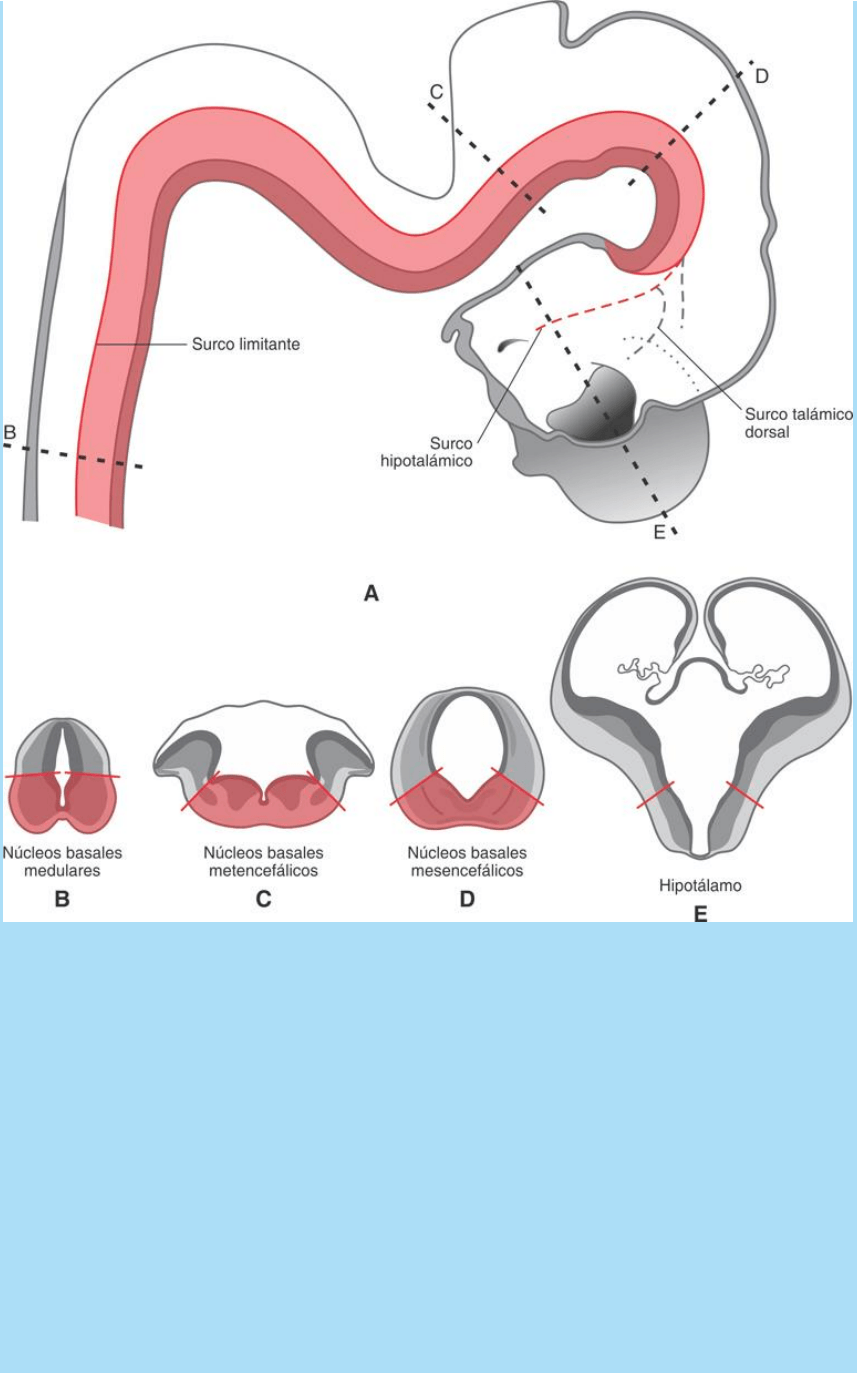
Fig. SC 9-4-1. Representación esquemática de proporciones
relativas de poblaciones neuronales ventrales (basales) y dorsales
(alares) en diversas regiones del neuroeje de un embrión humano de
la 7ª SD. A. Vista lateral derecha del encéfalo y médula cervical. Las
líneas de puntos B a E corresponden a los sitios en los que se
realizaron las secciones ilustradas abajo. B-E. Secciones
transversales del tubo neural. La línea roja corresponde al nivel del
surco limitante que separa las regiones alar y basal en las secciones
B a D. En el caso E la línea roja corresponde al nivel del surco
hipotalámico que representa la continuación cefálica del surco
limitante del mesencéfalo. B. Médula espinal
cervical. C.Prosencéfalo. Dorsalmente a la línea roja se observan los
labios rómbicos que originarán a los hemisferios
cerebelosos. D. Mesencéfalo. La región alar originará los tubérculos
Este documento contiene más páginas...
Descargar Completo
Cap 16 SC Flores.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.