
SC 8.1. Criterios que definen el sexo de un individuo. V. Flores
El genotipo se expresa en todos los niveles de organización en los
que se estructura el fenotipo. El dimorfismo sexual se manifiesta en
prácticamente todos esos niveles y, en consecuencia, en todos ellos,
desde el molecular al anatómico, se pueden definir criterios sobre la
base de los cuales se diagnostica el sexo de un individuo.
La vida en sociedad implica permanentes diagnósticos del sexo de
los individuos con los que interactuamos y decisiones, aun
inconscientes, de la conducta social. Los seres humanos
diagnosticamos el sexo de los demás en forma instantánea, sin
realizar ningún análisis profundo, tomando en cuenta, aunque no lo
hagamos consciente, varios criterios. Entre ellos los más importantes
son: a) las características anatómicas externas, forma corporal
(hombros, cintura, caderas, senos, relieves musculares y
óseos), b) la distribución pilosa (barba, bigotes, etc.), c) el tono de la
voz, d) las posturas corporales, las actitudes y la conducta en
general, entre otros. Muy accesoriamente tomamos en cuenta la
vestimenta. Con estos criterios, en general, realizamos
correctamente diagnósticos de sexo y reconocemos a un varón,
aunque esté vestido como mujer o a una mujer aunque esté vestida
como varón.
Todas las cualidades mencionadas, sin embargo, son características
sexuales secundarias y, desde el punto de vista médico, cuando se
consideran aisladamente pueden conducir a error, aun cuando se
tomen en cuenta los órganos genitales externos.
En el momento del nacimiento, el médico define el sexo al que
pertenece una persona y, en ese momento, no dispone de ninguna
de las características sexuales secundarias que permiten
diagnosticar el sexo. Pero el médico dispone de la inspección de los
órganos genitales externos. En el caso de individuos normales, la
decisión médica es correcta. Sin embargo, existen recién nacidos
que presentan “ambigüedad” en los órganos genitales externos y
resulta difícil, o imposible, definir el sexo sin el auxilio de otros
criterios. También existen recién nacidos que no presentan
ambigüedad, no generan duda, y el diagnóstico es incorrecto.
Por todos estos motivos, cuando se alude al sexo de una persona,
en el área de la medicina se recomienda especificar el o los criterios
tomados en cuenta para realizar el diagnóstico. Así, se habla del sexo
cromosómico o gonadal, por ejemplo. Algunos autores describen
esta situación diciendo que hay varios sexos (cromosómico, gonadal,
etc.). Tal conceptualización es incorrecta, la especie humana posee

dimorfismo sexual y los seres humanos corresponden a una de dos
categorías: el sexo masculino o el femenino. No existen varios sexos.
Existen varios criterios que definen el sexo del individuo.
Entre los criterios para considerar se pueden mencionar los
siguientes:
- El criterio genético. Sólo es posible aplicarlo por medio de un
análisis genético. En la actualidad se recurre a enunciar la presencia
o ausencia del gen Sry identificado como el encargado de la
determinación en sentido masculino (presencia de Sry= masculino;
ausencia de Sry = femenino). Sin embargo, con el avance del
conocimiento sobre cómo se regula la expresión de factores de
transcripción involucrados en la determinación sexual es de suponer
que, con el tiempo, se defina un conjunto de genes responsables de
la diferenciación para cada sexo. En la actualidad se considera que
el gen Dax es crucial en cuanto a la diferenciación en sentido
femenino.
- Criterios de regulación de la expresión génica. Cada vez más es
necesario tomar en cuenta este criterio pues alude, como se indica
en el punto anterior, a las combinatorias de factores de
transcripción que instalan redes de regulación de la expresión
génica responsables de determinación y diferenciación de tipo
masculino o femenino. Estas redes de regulación de la expresión
génica al modo masculino o femenino se instalan recién desde la 7ª
SD en adelante cuando cesa el período bipotencial del desarrollo
gonadal.
- Criterio cromosómico. Se realiza por medio del análisis del
cariotipo. Los individuos con cariotipo XY o XX son respectivamente
masculinos o femeninos desde este criterio.
- Criterio gonadal. Alude al modo como se produjo la histogénesis
gonadal. El sexo masculino se caracteriza por la diferenciación
gonadal en sentido testicular y el femenino por la diferenciación en
sentido ovárico.
- Criterio genital interno. En condiciones normales depende de, y
es coherente con, la diferenciación gonadal. Alude al modo como se
selecciona el tipo desarrollo de los conductos genitales internos: el
modo de desarrollo de tipo femenino (atrofia de conductos de Wolff y
desarrollo de conductos de Müller) o el desarrollo de tipo masculino
(desarrollo de conductos de Wolff e involución de conductos de
Müller. De acuerdo con estas modalidades de desarrollo el sexo
masculino se define por la presencia del epidídimo, conducto
deferente, etc. y el femenino por la presencia de trompas, útero,
vagina.

- Criterio genital externo. En condiciones normales depende de, y
es coherente con, la diferenciación gonadal. Alude a la modalidad
según la cual se diferencian el tubérculo genital y las eminencias
uretrales y genitales. En la modalidad masculina se desarrollan el
pene, la uretra peneana y las bolsas escrotales; en la modalidad
femenina: el clítoris, los labios menores y mayores. En condiciones
normales es el criterio que aplica el médico para definir el sexo de un
individuo recién nacido. Ante dudas en esta instancia se recurre a
evaluar otros criterios.
En la especie humana se aplican también los términos sexo legal,
sexo de crianza, sexo social, sexo psicológico y otros que no
siempre tienen definición precisa en el campo de la biología humana.
Los términos sexo legal y sexo de crianza no aluden a criterios
biológicos que definen el sexo. Se denomina sexo legal o civilal que
se asigna a la persona en el momento del nacimiento y que figura en
su documento de identidad. El sexo de crianza, a su vez, deriva del
sexo legal. Alude al, y resulta del, modo como se educa al individuo
en la familia, la escuela, etc. Los términos sexo psicológico y sexo
social están muy relacionados y aluden a la modalidad y/o rol
masculino o femenino con que el individuo maneja su conducta
general en su vida privada y/o se presenta y desempeña en su vida
en sociedad.
En consonancia con modificaciones legales introducidas
recientemente en nuestro país, en la actualidad existe también el
concepto de “identidad de género”. Este concepto alude al derecho
que asiste a las personas, en nuestra sociedad, de elegir una
“condición social” de tipo femenino o masculino, independientemente
de su sexo biológico, de acuerdo con las características psicológicas,
sociales, preferencias sexuales, etc., que el individuo elija como más
apropiada a su persona. Vale decir, como una elección de una forma
de vida que posibilita satisfacer sus expectativas de realización como
ser humano.
- Criterio conductual o de diferenciación del sistema nervioso
central. En los animales cuya conducta sexual implica la realización
de un cortejo integrado por un conjunto típico de actos estereotipados
que conducen a la cópula, se define también un criterio denominado
conductual. Este criterio pone de manifiesto principalmente el tipo de
determinación y diferenciación sexual que experimentó el sistema
nervioso (SC La diferenciación sexual del sistema nervioso central).
El sistema nervioso central sufre un proceso de determinación sexual
que, al igual que otros órganos que exhiben dimorfismo sexual,
depende de la presencia o ausencia de hormonas masculinizantes
durante un período crítico breve.

Algunos autores incluyen otros dos criterios: el criterio
cromatínico y el criterio hormonal. El criterio cromatínico alude al
hecho de que el examen de la cromatina de células interfásicas
descamadas permite ver con facilidad el corpúsculo de Barr (un
cromosoma X condensado). El número de corpúsculos de Barr
informa cuántos cromosomas X posee la célula. El criterio hormonal
alude a que tanto en varones como en mujeres, si bien ambos
producen hormonas masculinas y femeninas, existe un patrón (o
combinación) típico de concentraciones de hormonas de tipo
masculino y otro de tipo femenino.
Tomando en consideración todos estos criterios resulta claro que la
definición del sexo de cualquier individuo depende de la
coherencia con la que se expresan. Desde el punto de vista
biológico, la coherencia en la modalidad de expresión de dichos
criterios define la normalidad y el individuo es de sexo masculino o
femenino: a) es de sexo masculino aquel individuo en el que todos
los criterios se expresan al modo masculino y b) es de sexo femenino
aquel individuo en el que todos los criterios se expresan al modo
femenino. Los individuos en los que tales criterios se expresan
incoherentemente padecen de alguna de las alteraciones de la
determinación o la diferenciación sexual analizados en el capítulo 8,
o alguna otra que por motivos de espacio o relevancia no han sido
incluidas.
En los animales, la incoherencia en la expresión entre criterios que
permiten definir el sexo se denomina genéricamente estado
intersexual. Éste incluye básicamente todo el conjunto de patologías
congénitas de la diferenciación sexual presentes en la especie
humana.
SC 8.2. Combinatorias de factores de transcripción
involucrados en la regulación de la determinación y
diferenciación sexual de las gónadas. R. Rey, V. Flores.
Clásicamente se considera que existe un sexo básico, el femenino, y
que la diferenciación en sentido masculino es consecuencia de un
redireccionamiento del sexo básico. El concepto clásico es que la
diferenciación en sentido masculino requiere algún factor
determinante testicular (TDF) en tanto que la omisión de éste
produce diferenciación en sentido ovárico.
La existencia de un TDF fue un requisito de la concepción clásica.
Más de un “factor” ha ocupado, en los últimos tiempos, el lugar del
TDF. Así, en la literatura clásica en general se menciona el término
determinación sólo para referirse al desarrollo en sentido testicular.

Últimamente, sin embargo, han aparecido trabajos en los que se
habla también de determinación ovárica.
La determinación del sexo en sentido masculino no es consecuencia
de la expresión de alguna molécula en particular, sino consecuencia
de la expresión de una combinatoria de factores de transcripción,
entre los cuales se halla la proteína Sry. Los genes codificantes de
dichos factores de transcripción están localizados tanto en
cromosomas sexuales como en autosomas. De modo similar, la
determinación del sexo en sentido femenino también es
consecuencia de la expresión de otracombinatoria de factores de
transcripción, entre los cuales se halla la proteína Dax. También en
este caso los genes codificantes de dichas proteínas se hallan en
cromosomas sexuales y autosomas.
El período bipotente de las gónadas. Antes de la determinación del
sexo existe un período de tiempo durante el cual las gónadas inician
su diferenciación pero aún no están determinadas. Dicho período se
denomina bipotente ya que la gónada retiene potencia para
determinarse y diferenciarse en cualquiera de ambos tipo de gónadas
(Fig. SC 8-2-1). Durante el período bipotente, las gónadas de
individuos XX o XY expresan la misma combinatoria de proteínas
factores de transcripción Wt1 y Gata4.
{kind=link}
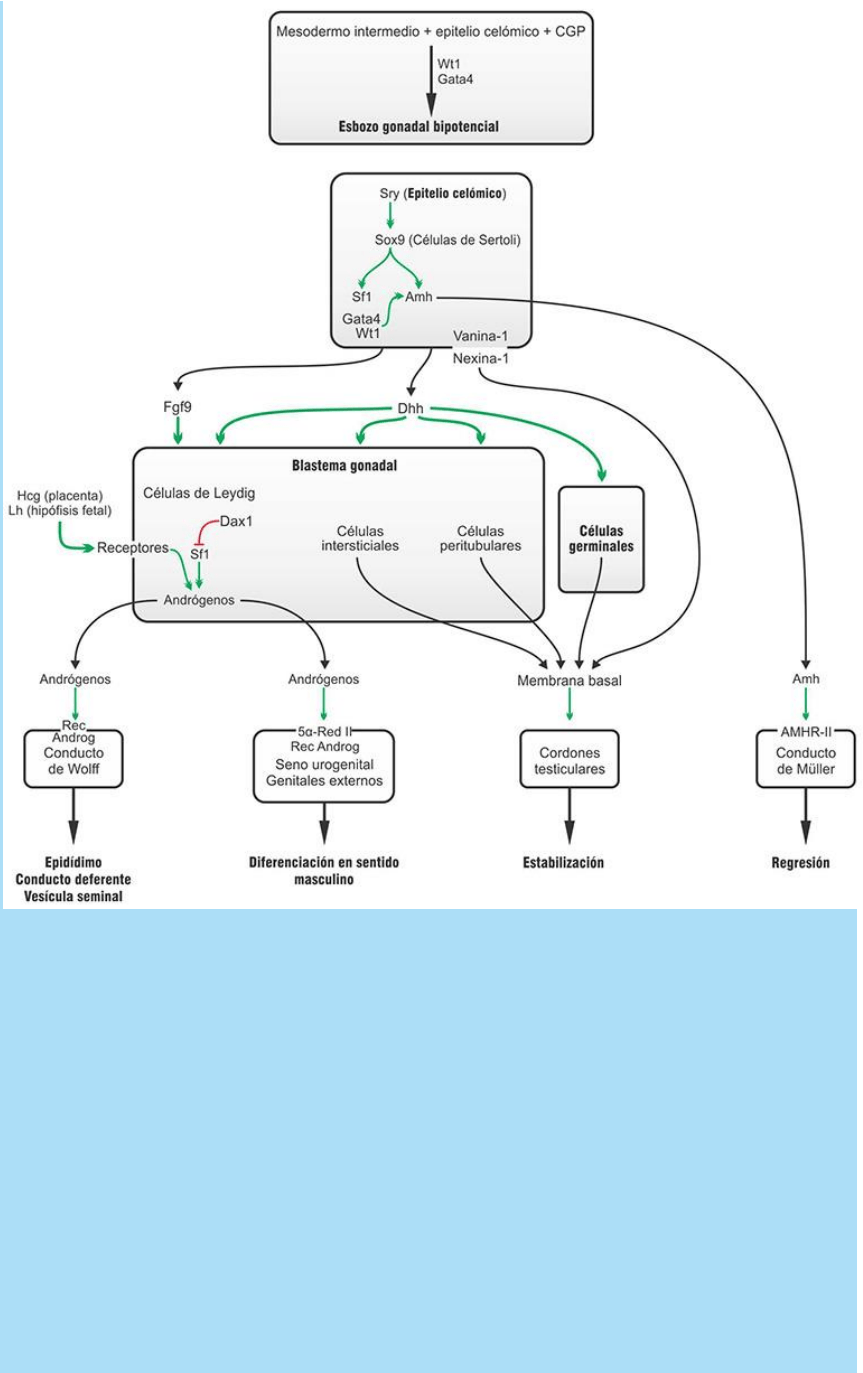
Fig. SC 8-2-1. Esquema de los comportamientos moleculares
involucrados en la determinación y diferenciación sexual en sentido
masculino.
La determinación sexual de las gónadas en sentido masculino.
También se denomina determinación primaria (Fig. SC 8-2-1)
- Se inicia con la expresión del factor de transcripción Sry en las
células del epitelio celómico que son precursoras de las células de
Sertoli.
- El factor Sry estimula la expresión de la proteína factor de
transcripción Sox9 en las células del epitelio celómico y estimula su
diferenciación en células de Sertoli.
- La expresión de la proteína Sry es transitoria, pero inicia una
cascada génica de diferenciación en sentido masculino en la que se

expresan varias otras proteínas que participan en la diferenciación de
todos los tejidos testiculares.
- Las células del epitelio celómico inician la expresión de la proteína
señal factor de crecimiento fibroblástico 9 (Fgf9), que atraería
quimiotácticamente a las células del mesodermo intermedio que
forman el blastema gonadal.
- La proteína señal Dhh, secretada por las células de Sertoli, y la
proteína receptor Patched2 (receptor de Dhh), expresada por las
células germinales, las peritubulares y las intersticiales, participarían
en las interacciones celulares por medio de las cuales se sintetizan y
depositan los componentes de la membrana basal necesarios para
el armado y mantenimiento de los cordones testiculares.
- Las células de Sertoli en diferenciación también expresan las
proteínas de membrana vanina-1 y nexina-1 que también
participarían en el mantenimiento de la membrana basal que rodea
los cordones testiculares y en la adhesión a las células peritubulares.
- El factor de transcripción Sox9 expresado por las células de Sertoli
estimula la síntesis y secreción de la proteína señal hormona
antimülleriana (AMH) por parte de estas células. La proteína Sox9
también estimula la expresión de la proteína factor esteroidogénico 1
(Sf1).
- La proteína Sf1, a su vez, en las células de Leydig en diferenciación,
estimula la expresión de las enzimas responsables de la producción
de hormonas esteroideas, como la testosterona.
- Los factores de transcripción Gata4 y Wt1 y también Sf1 modulan
positivamente y refuerzan la expresión de AMH inducida por Sox9.
- Las señales Fgf9 y Dhh generadas por las células de Sertoli
estimulan la diferenciación de las células de Leydig, secretantes de
andrógenos, a partir del mesénquima del blastema gonadal. La
señalización vía Hedgehog (Hh) es responsable de estimular la
diferenciación de las células progenitoras Sf1+ del blastema gonadal
en células de Leydig fetales. Sin embargo, no todas las células
progenitoras Sf1+ se diferencian en células de Leydig fetales.
Algunas de ellas permanecen indiferenciadas durante todo el
desarrollo. Se ha postulado que estas últimas originan a las células
de Leydig adultas y que la señalización vía Hh participaría
dinámicamente en modular las proporciones en la generación de
ambos tipos de células a partir de las progenitoras Sf1+.

- La expresión del factor de transcripción Sry antagoniza la acción del
factor de transcripción Dax1 que, indirectamente, instala una
inhibición a la expresión del Sf1.
- Así, la expresión de Sf1 actúa sin freno en las células de Leydig y
aumenta la síntesis de testosterona necesaria para la diferenciación
testicular y de los restantes órganos sexuales masculinos internos y
externos.
- La diferenciación de las células de Leydig incluye la expresión de la
proteína receptor de las hormonas luteinizante/gonadotrofina
coriónica (HCG). Estas hormonas aumentan la proliferación de las
células de Leydig y estimulan una alta secreción de andrógenos. La
HCG, de origen placentario, activa la síntesis de andrógenos por
parte de las células de Leydig durante los dos primeros trimestres de
la gestación. La LH producida por la hipófisis fetal lo hace durante el
último trimestre.
Así, dos hechos principales en la diferenciación testicular son a) la
constitución de los cordones de células de Sertoli a partir del epitelio
celómico y b) la diferenciación de las células de Leydig a partir del
mesénquima. Del normal funcionamiento de estas dos poblaciones
celulares depende la correcta diferenciación de los restantes órganos
del aparato reproductor masculino y la coherencia en la
diferenciación sexual de éstos (véase SC 8.1. Criterios que definen
el sexo de un individuo).
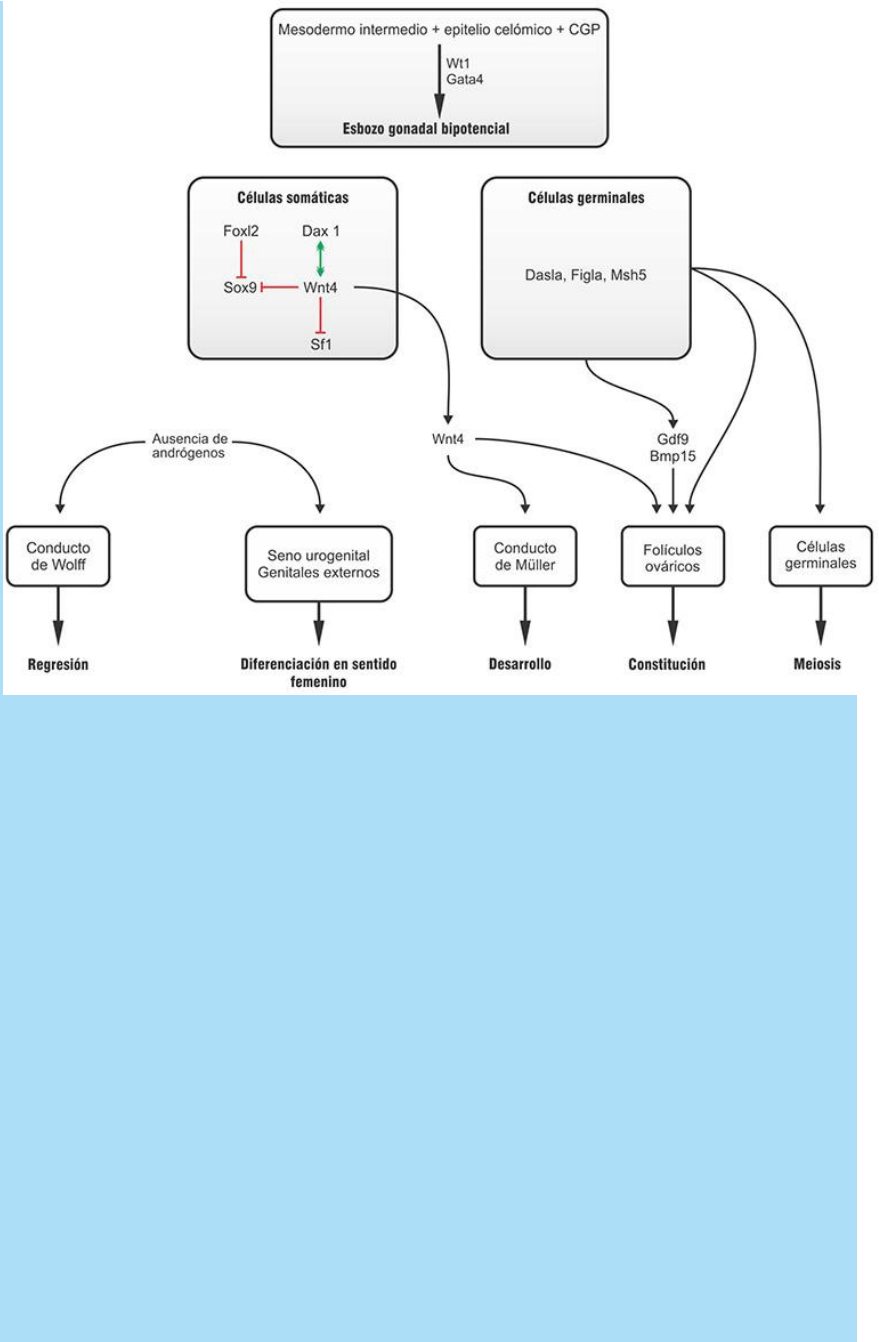
Fig. SC 8-2-2. Esquema de los comportamientos moleculares
involucrados en la determinación y diferenciación sexual en sentido
femenino.
La determinación sexual de las gónadas en sentido femenino.
También depende de la expresión de una combinatoria de factores
de transcripción que antagonizan el efecto de los que estimulan la
determinación en sentido masculino (Fig. SC 8-2-2).
- Incluida en dicha combinatoria de factores de transcripción se halla
la proteína Dax1 que algunos han propuesto como el factor
determinante ovárico por su papel antagonista de los efectos de Sry.
- La acción del factor Sry es antagonizada por los factores de
transcripción Dax1, Wnt4, Foxl2 y Sox3.
- La proteína Dax1 estimula la expresión del factor de transcripción
Wnt4.
- La proteína Wnt4, a su vez, inhibe la expresión de Sf1 que de este
modo se ve impedido de estimular la diferenciación de células o de
estimular en las células del mesénquima la expresión de las enzimas
responsables de la producción de testosterona.
{kind=link}

- La histogénesis ovárica, específicamente la constitución de folículos
integrados por células germinales rodeadas por células del epitelio
celómico, es dependiente de la presencia de células germinales
primitivas al esbozo de gónada.
- Por ello, todas las señales quimiotácticas e interacciones
necesarias para la migración y proliferación de las células germinales
primitivas son indispensables para la diferenciación ovárica.
- Además, ciertos factores como las proteínas Dazla, Figla y
Msh5 que participan de la regulación de la meiosis y los factores de
crecimiento como Bmp15 y Gdf9 son importantes reguladores de la
foliculogénesis. La falta de cualquiera de ellos ocasiona anomalías
del desarrollo ovárico.
SC 8.3. Regulación del desarrollo del aparato reproductor
durante la fase bipotencial. V. Flores, R. Rey
La población celular integrante del aparato reproductor que más
temprano aparece es la de células geminales (2ª SD). Durante la 3ª
SD ya se generan poblaciones celulares somáticas que integrarán el
aparato reproductor y en la 4ª SD empiezan los primeros procesos
de determinación. La más temprana probablemente sea el
mesodermo intermedio. Estos procesos de determinación no aluden
al sexo sino sólo a su carácter de integrantes del aparato reproductor.
El carácter sexual (masculino o femenino) se determina más tarde.
Ello se debe a que, con excepción de los conductos genitales
internos, los órganos de los aparatos reproductor masculino y
femenino no son distintos órganos sino los mismos pero
diferentemente diferenciados. Debido a ello, los esbozos de los
órganos del aparato reproductor, desde el punto de vista de su
carácter sexual, pasan por dos etapas: una primera fase bipotencial
en la que los esbozos son bipotentes (poseen capacidad para
determinarse y diferenciarse en cualquiera de ambos sexos) y una
segunda fase de determinación y diferenciación sexual. Ingresados
en esta segunda fase, los esbozos sólo pueden diferenciarse
normalmente en uno de ambos sexos.
La fase bipotencial. Se extiende hasta la 7ª SD o un poco más
dependiendo de qué órgano se trate. El órgano que más temprano
se determina sexualmente y del cual depende la diferenciación de los
demás es el testículo. En la regulación de los procesos de desarrollo
que ocurren durante la etapa bipotencial participan factores
morfogenéticos generales que participan en el desarrollo de otras
regiones corporales u otros aparatos o sistemas. Vale decir, no están

específicamente vinculadas a la diferenciación sexual. Las
alteraciones de la diferenciación sexual ocurren después de dicho
período. La fase bipotencial incluye la formación de varios esbozos
bipotentes.
Constitución del esbozo gonadal. Requiere la asociación de tres
poblaciones celulares: a) el mesénquima del mesodermo intermedio
(blastema gonadal), b) la hoja visceral del mesodermo lateral que lo
cubre superficialmente (epitelio celómico) yc) las células germinales
primitivas.
a) Determinación del mesodermo intermedio. Ocurre durante la
transición del período presomítico al somítico. Las células que van a
formar el mesodermo intermedio, al ingresar a través del surco
primitivo ya toman una posición intermedia entre el mesodermo
paraxil y el lateral: las células del mesodermo paraxil y del
mesodermo lateral se ubican en posición cefálica y caudal,
respectivamente, con respecto al mesodermo intermedio. Ya en el
período somítico, el mesodermo intermedio se extiende desde los
segmentos cervicales hacia el extremo caudal. En su determinación
parecen participar tanto señales mediales, de origen axial o paraxial,
como laterales, originadas en el mesodermo lateral caudal. La
expresión de los factores de transcripción Pax2, o Lim1,
considerados marcadores del mesodermo intermedio depende de
señales mediales. Cuando el mesodermo intermedio es separado,
por una incisión quirúrgica, del mesodermo paraxil no expresan
dichos marcadores.
b) Determinación del mesodermo lateral. Una vez formado el
mesodermo, a lo largo del eje medio-lateral se instala un gradiente L
m de expresión de la proteína señal
Bmp4 (Bone Morphogenetic Protein: proteína morfogenética del
hueso). Algunos experimentos indican que los elementos
mesodérmicos distribuidos a lo largo de dicho eje (región medial y
lateral del somita, mesodermo intermedio y mesodermo lateral) se
determinan y localizan en forma dependiente de las diferencias en la
actividad de la vía de señalización del Bmp. La determinación de
los mesodermos lateral, intermedio y paraxil requeriría
concentraciones altas, intermedias y bajas de Bmp4,
respectivamente. Aumentando la concentración de Bmp4 a lo largo
de dicho eje se puede disminuir la extensión de los elementos
mediales o, incluso, convertirlos en los más laterales. En presencia
de alta concentración de Bmp4, todo el mesodermo se transforma en
mesodermo lateral.
El proceso de determinación del mesodermo lateral y de sus
subregiones procede de lo general a lo particular. Tempranamente,

debido a la expresión diferencial, espacialmente organizada, de
factores de transcripción se segregan las regiones correspondientes
a la placa cardiogénica (corresponde a la región torácica) y el
mesodermo lateral posterior o caudal (corresponde a la región
abdominal). A continuación, esta última se deslamina en las hojas
somática y visceral del mesodermo lateral caudal. De ambas, la
visceral se fusiona con el blastema gonadal y pasa a formar
transitoriamente el esbozo gonadal.
c) Formación de las células germinales primordiales (CGP). Su
migración a la cresta gonadal. Las CGP se generan a partir de una
subpoblación de células epiblásticas pluripotentes localizada cerca
del extremo caudal del epiblasto pregastrular. Su aparición en el
epiblasto está promovida por señales del ectodermo
extraembrionario, como las proteínas señal Bmp2, 4 y 8, que
estimulan la expresión de la proteína transmembrana inducible
por interferón Fragilis Al parecer esta proteína marca el inicio de la
competencia, o de la determinación, en sentido de germinal. A
continuación, estas células, por medio deinteracciones
homotípicas, se segregan de las células somáticas vecinas. Sólo
aquellas que expresan alta concentración de Fragilis empiezan la
expresión de la proteína Stella, considerada como marcador de
determinación de la vía germinal. Algunos autores niegan esta
posibilidad. Las células Stella+, a continuación, sufren una represión
irreversible de los genes con caja homeótica. Algunos consideran
este hecho como indicio de represión de la vía somática y elección
de la germinal. Sólo unas 40 células del epiblasto realizan este
proceso. Varias investigaciones recientes indican que la expresión de
la proteína Stella no es esencial en la determinación de células
germinales. Otras proteínas, quizá más importantes, participarían en
este proceso. Durante la gastrulación, las CGP del epiblasto migran
al mesénquima extraembrionario y se ubican cerca de la base del
alantoides. Según algunos autores, recién en este lugar se produce
la determinación definitiva en sentido germinal ya que, al parecer,
algunas de estas células aún pueden evolucionar como mesénquima
extraembrionario.
Con el plegamiento del embrión, la región ocupada por las CGP es
llevada al interior del embrión. Desde allí migran a través de la hoja
visceral y llegan hasta la región del epitelio celómico de la cresta
gonadal.
Durante su migración las CGP proliferan, pero no se diferencian.
La proteína señal factor de células madre (SCF), expresada en el
mesénquima visceral y en el epitelio celómico del esbozo de gónada,

estimula la proliferación y migración de las CGP. Éstas, a su vez,
expresan en su membrana el receptor de SCF, la proteína receptor
c-kit. La migración dirigida de las CGP también depende de
interacciones con las proteínas de la matriz extracelular laminina
y fibronectina.
El número de CGP que llega y se incorpora a la cresta gonadal
depende del balance entre la proliferación y la apoptosis durante su
migración. En este balance interviene un conjunto de proteínas factor
de crecimiento y otras; como ejemplos pueden citarse Fgfb, Egf, Il4,
Tnf, Gdnf, Bcl/Bax, etcétera.
Cuando las CGP llegan a la cresta gonadal, junto con los otros
tejidos, integran el esbozo gonadal y se inicia la fase bipotencial del
esbozo de gónada. Con respecto a la fase de diferenciación sexual,
se considera que las CGP son indispensables para la histogénesis
del ovario pero prescindibles para la del testículo (SC
8.2. Combinatorias de factores de transcripción involucrados en la
regulación de la determinación y diferenciación sexual de las
gónadas; SC Regulación hormonal de la diferenciación sexual.
Claves para relacionar alteraciones hormonales y alteraciones de la
diferenciación sexual).
Constitución de los esbozos de gonaductos.
La formación de los conductos mesonéfricos (Wolff) está mediada
por T m-e que se producen en los extremos de crecimiento de los
conductos pronéfricos mientras éstos invaden el mesénquima
mesonéfrico. El crecimiento y formación de cada conducto implica un
proceso de migración celular dirigido seguido de una T m-e.
Los factores transcripción Emx2 y Pax2 y otras varias proteínas
expresadas en el mesénquima participan de dicha T m-e (SC
0.19 Transiciones reversibles mesenquimático-epitelial y epitelio-
mesenquimática. CMD involucrados en su regulación). Las células
mesenquimáticas que han iniciado la T m-e inician la diferenciación
de un dominio basal de la membrana plasmática acompañada de la
formación de una lámina basal en las zonas periféricas que quedan
en contacto con el mesénquima. En este proceso de generación y
estabilización de una interfaz de interacción entre epitelio y
mesénquima desempeñan un papel importante las proteínas de
matriz extracelular laminina, fibronectina, nidógeno 1
(entactina), proteoglucanos, colágeno tipo IV y otras. La
expresión de las proteínas homeóticas Hox10 y Hoxd13 es
también necesaria para una normal morfogénesis de los conductos
de Wolff.
Del tubo epitelial resultante de esta T m-e deriva el epitelio de
revestimiento de las estructuras glandulares de la mayor parte de las

vías espermáticas. La región del mesénquima peritubular, que no se
epiteliza, originará las capas de tejido conectivo y de músculo liso de
las vías espermáticas. Estas células tienen una importante función
en la respuesta a los andrógenos de origen testicular que estimulan
el crecimiento y diferenciación de los conductos de Wolff.
Los conductos paramesonéfricos (Müller) se forman como resultado
de invaginaciones longitudinales del epitelio celómico de disposición
paralela a los conductos mesonéfricos de cada lado del cuerpo.
Dicha disposición parece deberse a que éstos estimulan el desarrollo
de aquéllos. El mesénquima del mesodermo intermedio también
estimula, a través de la liberación de la proteína señal Wnt4, la
formación de los conductos paramesonéfricos. La expresión
de proteínas homeóticas Hoxa13, y otras, es necesaria para el
desarrollo temprano de los conductos de Müller.
Una vez constituido el conducto de Müller, su epitelio de
revestimiento secreta la proteína señal Wnt7a que estimula en el
mesénquima circundante la expresión de la proteína receptor de la
AMH (hormona antimülleriana). En el caso del sexo masculino, la
expresión de este receptor conduce a muerte celular y desaparición
del conducto paramesonéfrico por acción de la proteína
AMH secretada por las células de Sertoli. Este efecto se produce a
lo largo de un período sensible de sólo una semana o un poco más.
En el caso del sexo femenino, en ausencia de la proteína AMH, los
conductos paramesonéfricos siguen su desarrollo en tanto que la
ausencia de andrógenos llevan a la atrofia de los conductos
mesonéfricos.
Constitución de los esbozos de órganos genitales externos
El tubérculo genital y los pliegues uretrales y labioescrotales están
formados por condensaciones mesenquimáticas recubiertas por
epitelio ectodérmico. Durante la fase bipotencial del desarrollo del
tubérculo genital participan varias proteínas. Entre ellas figuran
la proteína homeótica factor de transcripción Hox4 y la proteína
señal factor de crecimiento fibroblástico 8 (Fgf8). Este factor de
crecimiento es sintetizado por el epitelio y estimula, en el
mesénquima subyacente, la expresión de otros factores de
crecimiento como Fgf10, Bmp4, y de las homeoproteínas
factores de transcripción Msx1 y Hoxd13. Éstas participan en la
morfogénesis del tubérculo genital y también de los esbozos de los
otros órganos genitales externos.

SC 8.4. Regulación hormonal de la diferenciación sexual.
Claves para relacionar alteraciones hormonales y alteraciones
de la diferenciación sexual. R. Rey
Diferenciación sexual masculina. El testículo fetal secreta dos
hormonas principales. Las células de Sertoli de los cordones
seminíferos producen la proteína señal hormona antimüllerriana
(AMH) y las células de Leydig del intersticio sintetizan lahormona
esteroidea masculina testosterona. La AMH se une a su receptor
presente en los conductos de Müller, provocando su regresión. La
testosterona, al unirse a sus receptores en los conductos de Wolff,
permite su diferenciación en epidídimos, conductos deferentes,
vesículas seminales y conductos eyaculadores. En el seno urogenital
y en los esbozos de los genitales externos, la testosterona es
transformada, por la enzima 5-alfa-reductasa tipo 2 en otra
hormona esteroidea masculina dihidrotestosterona (Dht), un
andrógeno más potente que se une al receptor de andrógenos con
más afinidad. La Dht estimula a) la diferenciación de la próstata, b) el
crecimiento del tubérculo genital (que forma el pene), c) el
crecimiento de la lámina uretral endodérmica y d) la fusión completa
de los pliegues uretrales que forman así la uretra peneana y la fusión
completa de los pliegues labioescrotales que se diferencian en
escroto. También interviene en f) el descenso testicular a través del
conducto inguinal en el 3
er
trimestre de la gestación. Durante los dos
primeros trimestres de la gestación, la producción de andrógenos
depende de la estimulación de las células de Leydig por la proteína
hormona gonadótrofina coriónica (HCG). En el último trimestre
depende de la secreción de la proteína hormona luteinizante
(LH) secretada por la hipófisis fetal.
Diferenciación sexual femenina. Los ovarios secretan muy
pequeñas cantidades de andrógenos y de AMH a partir de la 36ª SD.
Los ovarios secretan estrógenos pero éstos no determinan el sexo
en sentido femenino. El ovario fetal no produce sustancias que
influyan en la diferenciación sexual. La diferenciación en sentido
femenino de los órganos genitales se produce normalmente en
ausencia de ovarios. Lo mismo ocurre en embriones XY castrados
antes de que se produzca la determinación sexual.
La ausencia de las hormonas de origen testicular es suficiente para
la diferenciación en sentido femenino: a) la ausencia de AMH permite
la estabilización, el desarrollo y diferenciación de los conductos
paramesonéfricos que forman útero, trompas y parte superior de la
vagina y b) la ausencia de andrógenos produce la regresión del
conducto mesonéfrico. Por otro lado, c)el seno urogenital origina la
porción inferior de la vagina. Por su parte, d) el tubérculo genital
Este documento contiene más páginas...
Descargar Completo
Cap 16 SC Flores.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.