
SC 5.1. El origen de las células que forman la placa
cardiogénica. V. Flores
Las células que forman el corazón han sido detectadas y seguidas
desde la gastrulación en adelante. Inicialmente se hallan en el
epiblasto, durante la gastrulación migran hacia la línea media y se
invaginan a través del surco primitivo. Se postula que la posición de
las células, a lo largo del eje céfalo-caudal, cuado se invaginan en el
surco primitivo se relaciona con su futura ubicación en la placa
cardiogénica y con la región del corazón que originarán.
El modelo presentado en la figura SC 5-1-1 A y B ilustran dicha
relación. En A se observa que las células que se invaginan
medialmente, en el extremo del surco primitivo, forman la notocorda
y, adyacentes a ellas, las células que formarán zonas mediales del
endodermo. Por el modo como se producen los desplazamientos
gastrulares ‒indicado por las flechas de lafigura SC 5-1-1 A ‒ Las
células que ocupan posiciones cefálicas del surco originarán
regiones cefálicas de la placa y las que ocupan posición caudal
originarán partes caudales de la placa. Así, las células ubicadas
cefálicamente en el surco primitivo originarán la parte medial y
cefálica de la placa cardiogénica; esta región originará el bulbo
cardíaco (ventrículo derecho, cono y tronco de salida de los
ventrículos y, probablemente, también la aurícula izquierda). Un poco
más caudalmente se ubican las células que formarán el ventrículo
primitivo (originará el ventrículo izquierdo) y, a continuacion, las que
formarán aurículas (aurícula derecha) y, probablemente, senos
venosos.
Así, debido al modo como se producen los desplazamientos
grastrulares, por la posición que ocupan las células en el surco
primitivo puede deducirse qué regiones del corazón originarán.
La figura SC 5-1-1 C ilustra este hecho y muestra cómo quedarán
ubicadas dichas regiones cardíacas durante la torsión del corazón
tubular primitivo.
{kind=link}
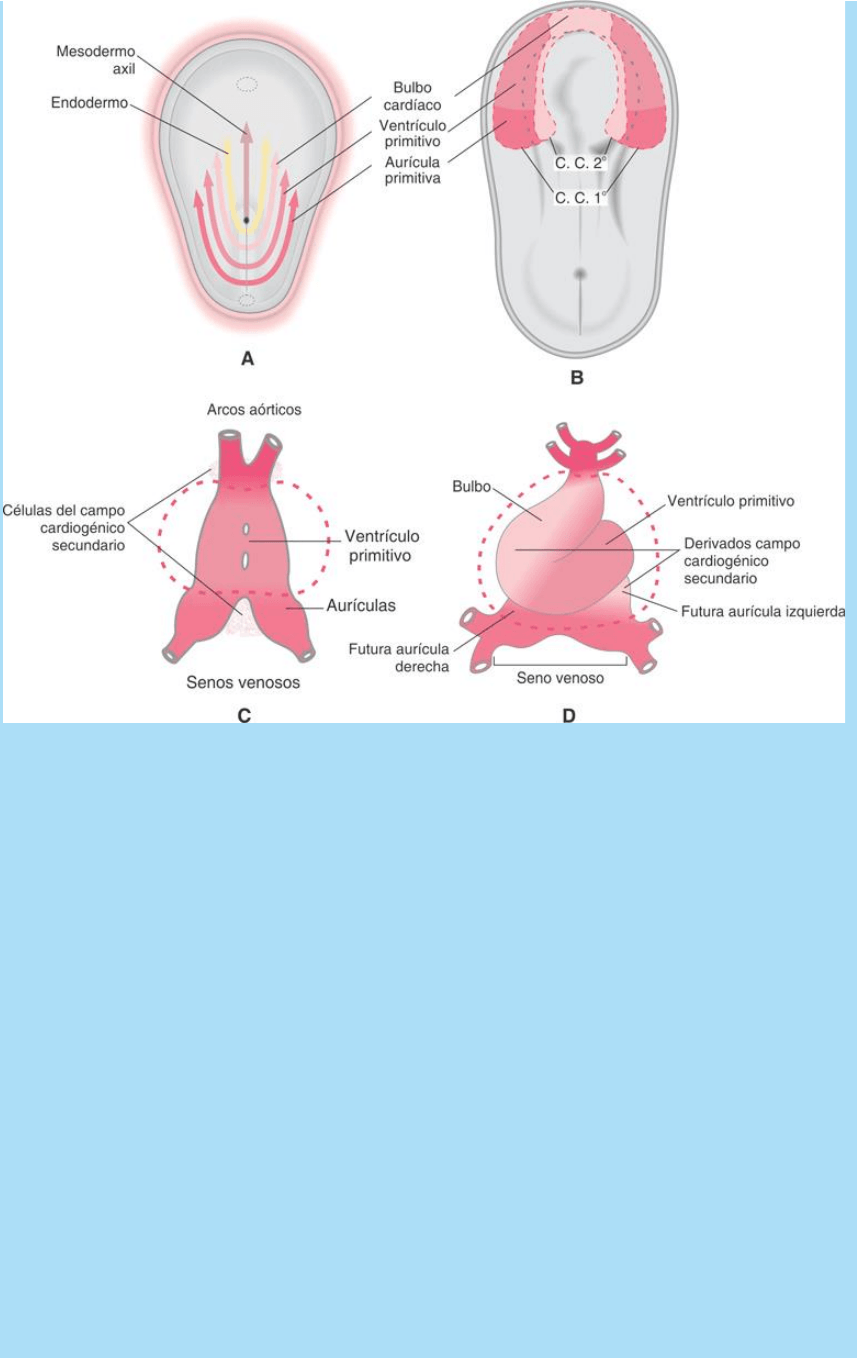
Fig. SC 5.1.1. A-B. Origen epibástico de las células de la placa
cardiogénica. La posición de estas células en el epiblasto se
relaciona con la posición que ocupan en la placa y la región cardíaca
que originan. C. El ordenamiento céfalo-caudal observable a lo largo
del surco primitivo se repite en la placa cardiogénica y se traduce en
el ordenamiento céfalo-caudal de las cavidades del corazón tubular
primitivo recto medial (CTP). D. Durante la torsión del CTP, las
cavidades modifican sus posiciones relativas.
Ello no implica, sin embargo, que las subpoblaciones celulares
ubicadas a lo largo del surco primitivo ya estén determinadas a formar
las distintas regiones mencionadas. Esta especificación ocurre más
tarde (SC 5.2. La determinación de la placa o campo cardiogénico.
Comportamientos celulares y moleculares involucrados). Al parecer,
la especificación regional de las células cardiogénicas no se produce
simultáneamente en todo el campo cardiogénico. Al menos las
células ventriculares son especificadas antes que las auriculares
(SC La determinación progresiva de las células procardiogénicas, las
de los campos cardiogénicos primario y secundario y restantes
estirpes celulares que forman el corazón; SC Determinación y

diferenciación de las células del área cefálica del campo cardiogénico
secundario durante la formación del bulbo cardíaco).
Las células de la futura placa cardiogénica parecen poseer cierto
grado de determinación genérica en sentido cardiogénico, a juzgar
por ciertos marcadores que expresan muy tempranamente, aún
antes de la gastrulación.
Por otro lado, existen estudios de linaje celular con marcadores
retrovirales que sugieren que ya antes de la gastrulación es posible
distinguir clones de células con diferente capacidad histogenética:
endocardiogénicas y miocardiocitogénicas.
Así, se ha propuesto que ya pregrastrularmente existirían células
determinadas en sentido cardiogénico. Como puede apreciarse,
estos diferentes estudios aluden a fenómenos de especificación y
determinación de diferente naturaleza: a) por un lado, una
determinación genérica en sentido cardiogénico, b) una
especificación que alude a la definición de regiones y c) una
especificación en el nivel tisular o histogenético.
SC 5.2. La determinación de la placa o campo cardiogénico.
Comportamientos celulares y moleculares involucrados. V.
Flores
El campo cardiogénico se constituye con células provenientes del
epiblasto que durante la gastrulación forman el mesodermo lateral.
Estas células migran cranealmente, convergen por delante de la
placa precordal y forman la región semilunar denominada placa
cardiogénica. En sentido estricto, es la hoja esplácnica resultante de
la deslaminación de esa zona del mesodermo lateral la que debe
recibir el nombre de campo cardiogénico. La hoja somática de dicha
región no participa en el desarrollo cardíaco.
La especificación y posición del campo cardiogénico depende de la
posicón espacial de varias fuentes de señales positivas (activadoras)
y negativas (inhibidoras) de la cardiogénesis.
El modelo ilustrado en la figura SC 5-2-1, basado en el embrión de
pollo, propone que la ubicación y forma del campo cardiogénico
depende de los siguientes hechos: a) las células mesodérmicas de la
región 1 no reciben señales, b) las células de la región 2 reciben
ambos tipos de señales y c) las celulas de la región 3 reciben señales
positivas y se hallan fuera del rango de acción de las negativas.
{kind=link}
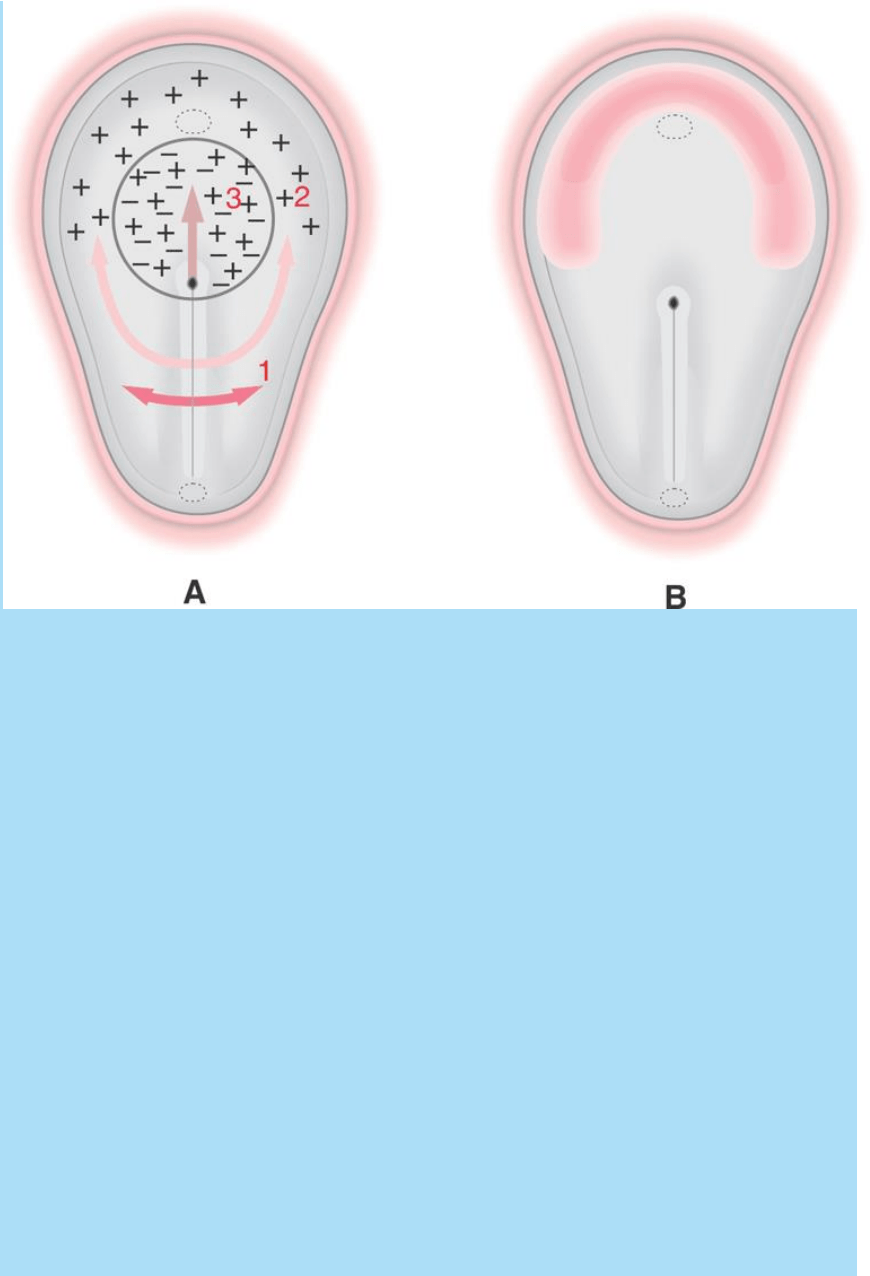
Fig.SC 5-2-1. Modelo de determinacion y localización especificacion
del campo cardiogénico por medio de la acción de senáles
activadoras (+) e inhibidoras (-) de la cardiogénesis. A. Distribución
espacial de señales + y -. En la region 1 no existen señales
cardiogénicas; en la 3 sí existen, pero son contrarrestadas por
señales inhibidoras. En la 2 el mesodermo sólo recibe señales
cargiogénicas. B. Ilustra la localización y la forma semilunar del
campo cardiogénico resultante de la distribución espacial de señales
+ y -.
Las células del campo cardiogénico están determinadas a formar
sólo algunos tipos celulares del corazón definitivo. La capacidad
citogénica del campo cardiogénico se ilustra en el cuadro SC 5-2-1.
Las restantes células cardíacas son de origen extracampo
cardiogénico, como las provienentes de la población proepicárdica y
de la cresta neural cardiogénica, y dependen de fenómenos de
determinación diferentes (SC 5.4. El origen múltiple de las
poblaciones celulares que intervienen en la cardiogénesis; SC 5.3. La
determinación progresiva de las células procardiogénicas, las de los
campos cardiogénicos primario y secundario y restantes estirpes
celulares que forman el corazón).
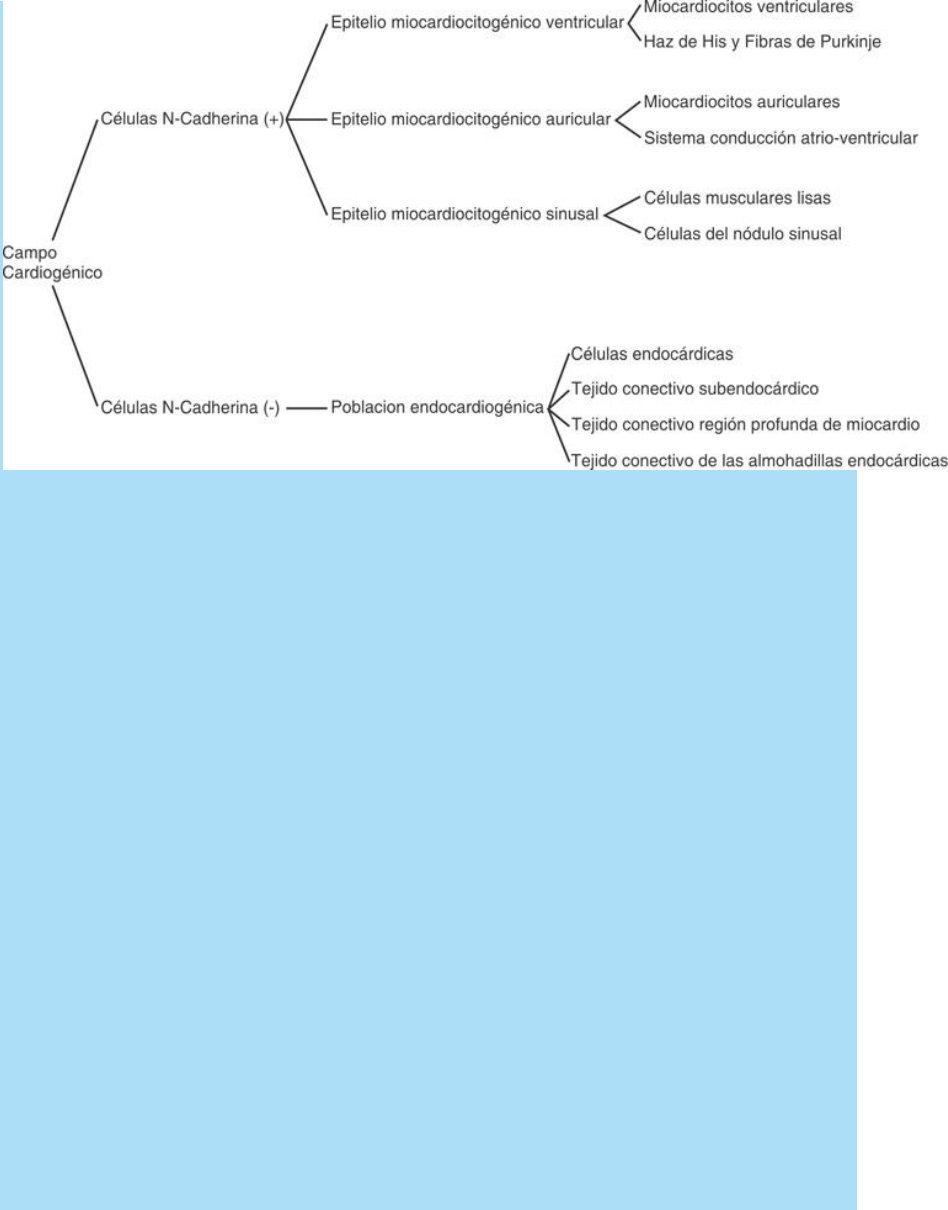
Cuadro SC 5-2-1. Ilustra la capacidad citogénica del campo
cardiogénico originado durante la gastrulación a partir de
precursores hipoblásticos N-Cadherina (+) y (-). (SC El origen
múltiple de las poblaciones celulares que intervienen en la
cardiogénesis).
AT.: cambiar atrio-ventricular => atrioventricular; Población =>
Población
Las señales involucradas y sus fuentes de origen. La figura SC
5-2-3 ilustra un modelo sobre la especificación y el patterningdel
campo cardiogénico en el ratón basado en la distribución espacial de
señales cardiogénicas y anticardiogénicas.
Las señales positivas (cardiogénicas) provienen fundamentalmente
del endodermo e incluyen a la proteína morfogenética ósea 2
(BMP-2), el factor de crecimiento fibroblástico 8 (FGF-8) y las
proteínas Crescent y Cerberus, que son proteínas inhibitorias de la
acción de las proteínas Wnt actuando sobre sus receptores. El
ectodermo epidérmico también es fuente de BMP.
Las señales negativas (anticardiogénicas) provienen
fundamentalmente de a) el tubo neural, fuente de las proteínas
Wnt3a y Wnt8 que promueven la formación de vasos sanguíneos e
inhiben la del corazón y de b) el mesodermo axil cordal que secreta
las proteínas señal Nogina y Cordina que inhiben a la señal BMP
procedentes del endodermo.
{kind=link}
{kind=link}
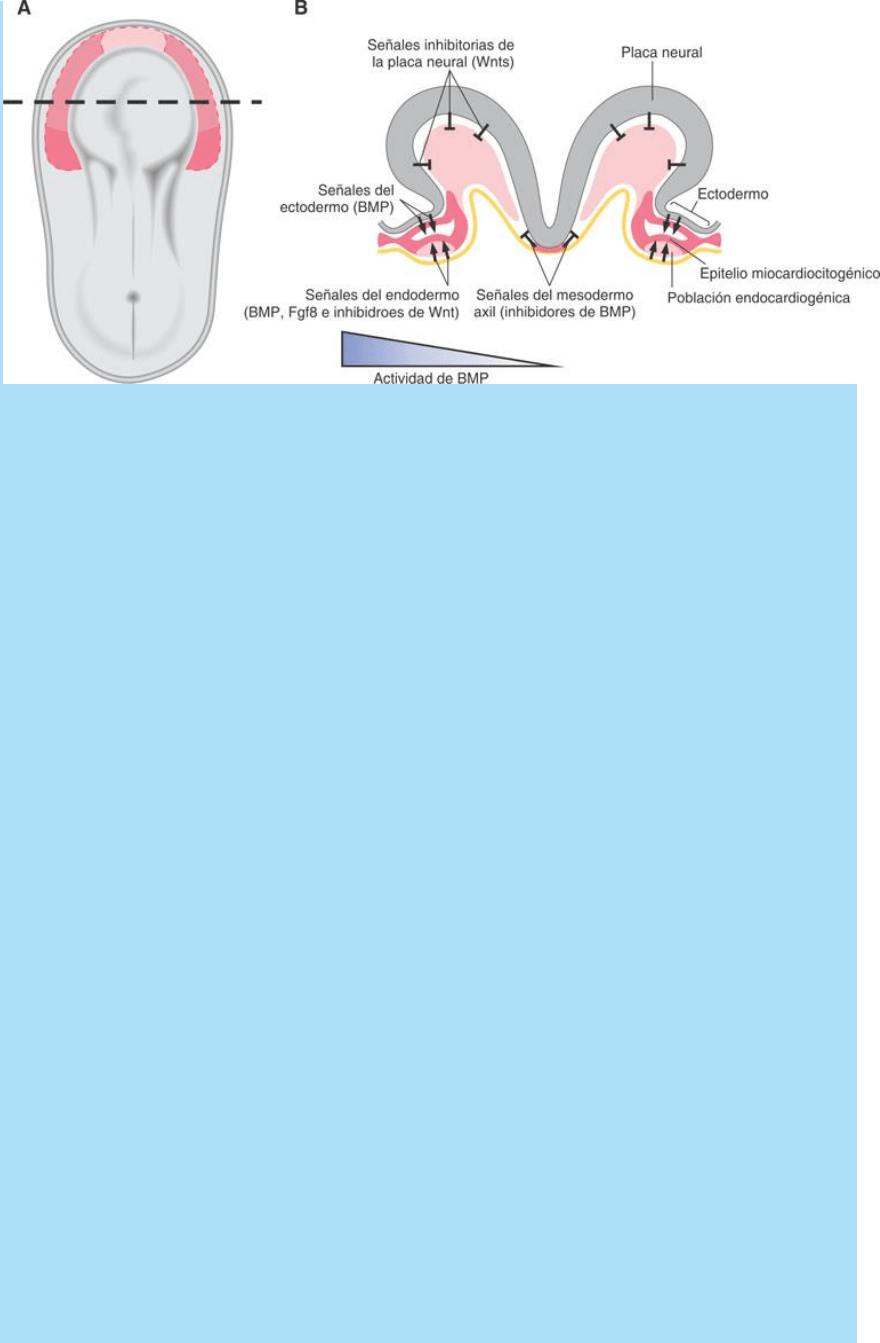
AT. arriba: cambiar inhibidroes => inhibidores
Fig. SC 5-2-3. Modelo de determinación, localización y modelación
del campo cardiogénico. A. llustra esquemáticamente la posición y
orientación del corte (línea de puntos) mostrado en B. B. Corte
transversal del embrión. Se indican las posiciones de las poblaciones
celulares señalizadoras (ectodermo, endodermo faríngeo, placa
neural y mesodermo axil) y sus correspondientes señales
estimulantes (flechas) e inhibitorias (barras T) de la cardiogénesis. El
triángulo azul y la gradación de su color representan la variación en
la intensidad del efecto cardiogénico resultante de la acción de dichas
señales. Este fenómeno ubica y determina la extensión del campo
cardiogénico a lo largo del corte (eje medio-lateral). Se observan el
mesodermo deslaminado, el celoma pericárdico, la hoja esplácnica o
campo cardiogénico que ya posee una población endocardiogénica
adyacente al endodermo faríngeo y un epitelio miocardiocitogénico
limitando ventralmente el celoma pericárdico.
Las señales cardiogénicas se distribuyen en forma de gradiente
látero medial. La señal BMP es originada tanto en el ectodermo
como en el endodermo del intestino anterior (faríngeo). Éste también
secreta Fgf8 e inhibidores de Wnt.
Puede apreciarse que el factor determinante del patterning del
campo cardiogénico es la distribución de los tejidos fuente de las
señales positivas y negativas: el papel de las señales positivas es
estimular la cardiogénesis en el mesodermo esplácnico de la región
cefálica y el papel de las señales negativas es delimitar la extensión
del campo y definir el área apropiada.
Las proteínas Bmp inducen la expresión de Fgf-8 en el endodermo y
éste participa estimulando la expresión de las proteínas factores de

transcripción Tal 1, Tboxs 2,3 y 5, Nkx 2.5 y cGATA que participan
en la programación de la evolución en sentido cardíaco.
Se ha comprobado que esta combinatoria “cardiogénica” de factores
de transcripción inicia una cascada de activación génica que incluye
la expresión de varios genes que codifican proteínas específicas de
músculo cardíaco (actina cardíaca, factor natriurético atrial y las
cadenas de alfa-miosina y otras).
SC 5.3. La determinación progresiva de las células
cardiogénicas: la placa cardiogénica, los campos
cardiogénicos primario y secundario y otras estirpes celulares.
V. Flores
Varios estudios clásicos de mapeo de destino de células de la placa
cardiogénica hicieron suponer que el corazón tubular primitivo (CTP)
recto posee las poblaciones celulares precursoras de todos los
tejidos y todas las regiones cardíacas. Algunos estudios más
recientes indican que el CTP recto no cumple ninguna de ambas
propiedades. El CTP recto representa sólo a la región denominada
ventrículo primitivo que origina al ventrículo izquierdo definitivo. La
región denominada a) tracto de entrada, que comprende los
conductos auriculoventricular, auricular y sinusal (caudal al
ventrículo primitivo) y la región denominada b) tracto de salida, que
comprende el bulbo cardíaco (cefálico al ventrículo primitivo) se
agregan secundariamente al CTP recto durante la torsión cardíaca.
Se considera que el agregado de dichas regiones al CTP recto es,
precisamente, el cambio morfológico denominado torsión
cardíaca. El tracto de entrada al CTP recto se agrega a éste antes
que el tracto de salida o bulbo cardíaco. Las regiones que se
agregan tardíamente al CTP recto, y que provocan su torsión, se
generan en el campo cardiogénico secundario (SC 5.4. El origen
múltiple de las poblaciones celulares que intervienen en la
cardiogénesis; SC Determinación y diferenciación de las células del
área cefálica del campo cardiogénico secundario durante la
formación del bulbo cardíaco). Esta población celular fue identificada
por medio de un marcador que no comparte con el resto de las
células de la placa cardiogénica. Con el objeto de distinguir a esta
población celular del resto de las células de placa cardiogénica se la
ha denominado campo cardiogénico secundario. Así, las
restantes células de la placa cardiogénica constituyen un campo
cardiogénico primario (SC 5.5. El área cefálica del campo
cardiogénico secundario y sus derivados anatómicos e histológicos).
Las porciones de entrada al CTP recto (aurícula primitiva y senos

venosos) se forman y agregan a él a partir de la porción caudal,
bilateral, de la placa cardiogénica.
La cardiogénesis incluye varios fenómenos determinantes y
morfogenéticos que, como en otros casos, van de lo general a lo
particular de acuerdo con criterios de ordenamiento temporoespacial.
a) El proceso de especificación más temprano ocurre
pregastrularmente: la aparición de células genéricamente
procardiogénicas en el epiblasto. Esta población tempranamente
se segrega en dos subpoblaciones: una endocardiogénicay
otra miocardiocitogénica. Este fenómeno implica una
especificación de tipos tisulares pero no instala una especificación
regional. Este segundo aspecto empezaría a instalarse durante la
gastrulación, dependiendo de la posición que ocupan las células
migrantes a lo largo del eje céfalo-caudal del surco primitivo. Sin
embargo, tampoco este fenómeno implica un proceso de
especificación sino más bien de relación probabilística entre posición
inicial en el surco primitivo y posición final en la placa cardiogénica.
b) La segunda etapa es la especificación y patterning de la placa
cardiogénica. Se trata de un conjunto de procesos de señalización
espacialmente estructurados mediado por la distribución
de señales cardiogénicas y anticardiogénicas en el que
participan varios centros señalizadores, varias proteínas señal,
sus respectivos receptores e inhibidores (SC 5.2. La determinación
de la placa o campo cardiogénico. Comportamientos celulares y
moleculares involucrados). Con respecto a la especificación de
regiones en la placa cardiogénica, se ha propuesto un modelo según
el cual la especificación se produce en sentido céfalo-caudal: primero
la región del ventrículo primitivo y luego aurícula primitiva. También
existe un modelo que propone que la parte final o senos venosos no
sufren un proceso de especificación de tipo cardiogénico. Lo descrito
hasta aquí corresponde al campo cardiogénico primario de la
placa cardiogénica.
c) Poco después se produce la determinación de las células
del campo cardiogénico secundario. A medida que estas células
se incorporan al CTP recto se determinarían en bulbo, cono y tronco
arterioso. La determinación temprana de las células del campo
secundario parece ser similar a la determinación de las del campo
primario. Sin embargo, durante su incorporación al CTP recto, como
tracto de salida de este último, sufren una reprogramación adicional
que probablemente esté relacionada con su determinación y
diferenciación en bulbo cardíaco (SC Determinación y diferenciación

de las células del área cefálica del campo cardiogénico secundario
durante la formación del bulbo cardíaco).
d) Poco más tarde, mientras se van formando los arcos aórticos y sus
correspondientes arcos branquiales, el mesénquima regional es
invadido también por células provenientes de los 3 o 4 últimos
segmentos de la cresta neural vagal. Estas células migran siguiendo
el trayecto de los arcos aórticos, llegan al saco aórtico e invaden,
desde el extremo cefálico al caudal, las paredes del tronco y cono,
llegando profundamente hasta la región de los conductos
auriculoventriculares. Las células de la región de cresta neural
mencionada, denominada por dicho motivo cresta neural
cardiogénica, participan aportando tejidos conectivo y muscular liso
a los derivados del tronco-cono (porción proximal de grandes vasos).
e) Finalmente, las células que forman el pericardio visceral o
epicardio, el tejido conectivo perivascular de los vasos coronarios y,
probablemetne, también su musculatura lisa vascular provienen de
una población celular que ingresa en el corazón a través de su polo
caudal. Estas células tienen su origen en una región diferenciada de
mesénquima, denominadapoblación proepicárdica, que rodea a
los senos venosos y la superficie del septum transversum.
SC 5.4. El origen múltiple de las poblaciones celulares que
intervienen en la cardiogénesis. V. Flores
Existen ejemplos de poblaciones celulares embrionarias que poseen
una amplia potencia citogenética y que, en consecuencia, en forma
divergente, originan varios tipos celulares que forman parte de
distintos órganos. Existen, por otro lado, órganos que se forman
como consecuencia de la confluencia, combinación e integración de
poblaciones celulares que poseen su origen en diferentes
poblaciones celulares.
El desarrollo del corazón es un ejemplo que reúne ambas situaciones
ya que, por un lado, a) su desarrollo requiere la participación varios
tipos celulares de distinto origen embrionario que confluyen y se
integran en la elaboración de los tejidos cardíacos y, por otro lado, b)
varias poblaciones celulares embrionarias precursoras de células y
tejidos cardíacos aportan también células que participan en el
desarrollo de otros órganos.
Entre las varias poblaciones que participan en el desarrollo del
corazón están las siguientes.
1) Las células epiblásticas que forman la placa o campo
cardiogénico. Estas células tienen la capacidad de formar en principio
dos subpoblaciones: a) endocardiogénica o N-cadherina (-
) y b) miocardiocitogénica o N-cadherina (+). Las primeras, N-
cadherina (-), originan las células endocárdicas de todo el corazón,
el tejido conectivo subendocárdico y el tejido conectivo de la porción
profunda del miocardio (SC 5.3. La determinación progresiva de las
células procardiogénicas, las de los campos cardiogénicos primario
y secundario y restantes estirpes celulares que forman el corazón).
Las segundas, N-cadherina (+), originan los miocardiocitos
auriculares y ventriculares y las células del sistema de conducción
auriculoventricular, el haz de His y las fibras de Purkinje de los
ventrículos (Fig. SC 5-4-1).
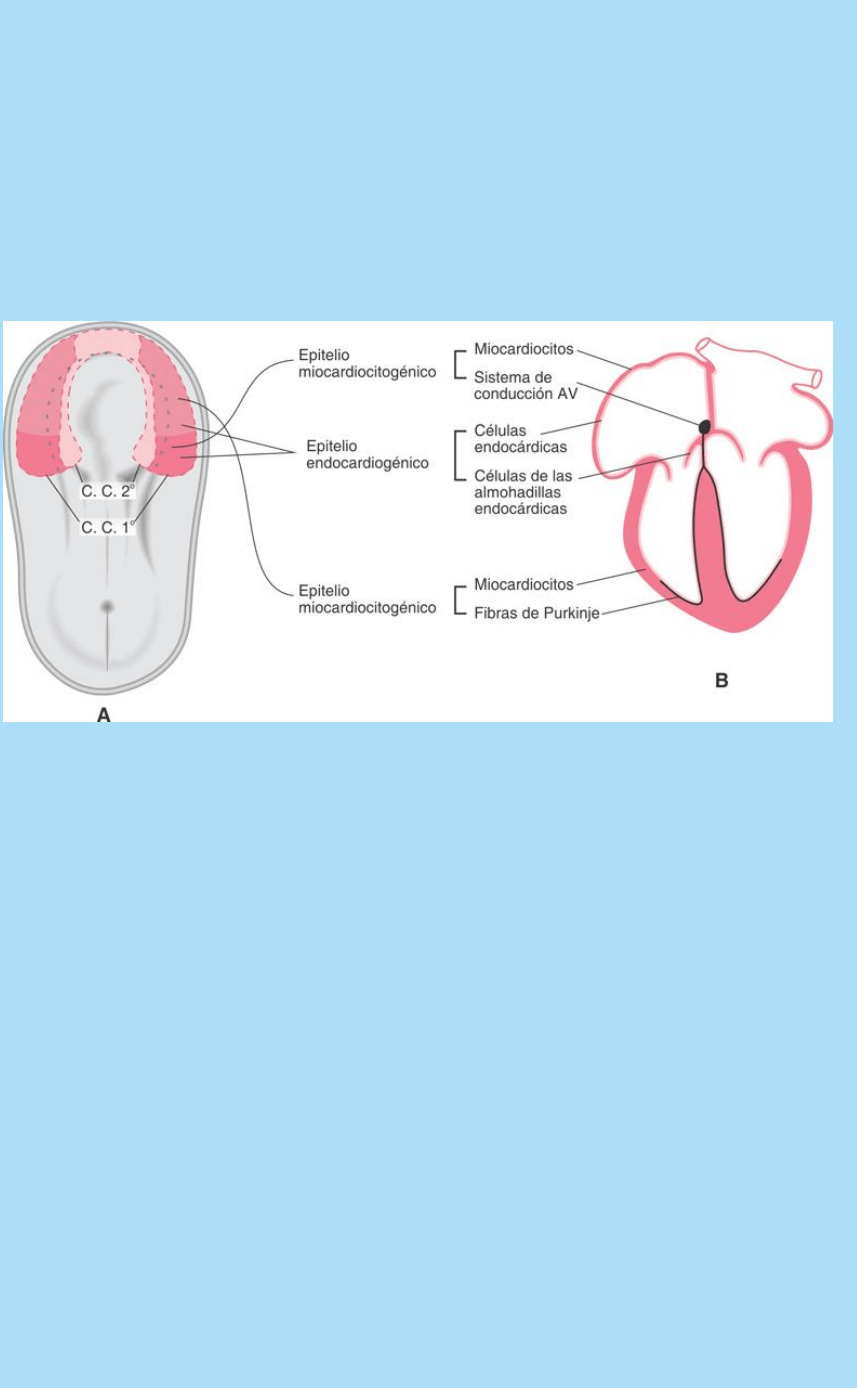
Fig. SC 5-4-1. Esquema de la relación entre posición dentro del
campo cardiogénico primario (C.C.1º), región cardíaca y tipo celular.
Distintas regiones de la placa cardiogénica aportan diferentes tipos
celulares a las diferentes regiones del corazón.
2) A estas células se les agregan luego las que se segregaron
como campo cardiogénico secundario que forman algunos de
los tipos celulares ya mencionados pero, en el ventrículo
derecho y el cono y, además, células musculares lisas
vascularesde los infundíbulos y porciones proximales de las
arterias aorta y pulmonar (SC Determinación y diferenciación de
las células del área cefálica del campo cardiogénico secundario
durante la formación del bulbo cardíaco). El campo cardiogénico
secundario es una adquisición filogenética reciente, está asociado a
la evolución del circuito pulmonar en relación con los cambios
evolutivos sufridos en la transición de la forma de vida acuática a la
terrestre. Desde esta perspectiva, se considera que también forman
parte del campo cardiogénico secundario las células del
mesoesófago ventral en la que se desarrolla la vasculatura pulmonar,
que se continúan anatómicamente con el corazón y que se
incorporan al mismo formando la mayor parte de la pared de la
{kind=link}
aurícula izquierda. Así, el campo cardiogénico secundario se ubica
durante el período somítico temprano a lo largo del mesocardio
dorsal y tiene dos regiones importantes: a) el área superior o
cefálica o del tracto de salida origina albulbo cardíaco del que
luego derivan el tracto de salida del corazón y el ventrículo
derecho, y b) el área inferior o caudal odel tracto de entrada y
pulmonar, asociada al esbozo pulmonar, originaría parte importante
de la aurícula izquierda. Vale decir, las dos cámaras cardíacas
correspondientes al circuito pulmonar. Algunos investigadores
sostienen, con argumentos valederos, que el área caudal del
campo cardiogénico secundario incluye también la población
celular proepicárdica asociada al seno venoso ubicado en el septum
transversum (SC La formación de la población pcPE. Su relación con
el campo cardiogénico secundario. Mecanismo de transferencia
celular seno-venoso CTP en mamíferos) (Fig. SC 5-4-2).
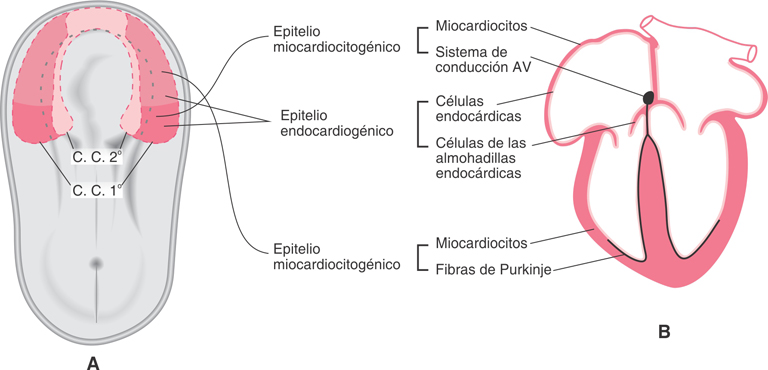
Fig. SC 5-4-2. Representación esquemática de la capacidad
citogenética de las células del campo cardiogénico secundario y su
relación con las cavidades cardíacas. La población proepicárdica
integra el campo cardiogénico secundario. Junto a estas células
ingresan células endoteliales de los sinusoides hepáticos.
3) Una tercera población celular proviene de la cresta neural
cardiogénica. Esta cresta está compuesta por los tres primeros
segmentos occipitales que se encuentran incluidos en la cresta vagal.
Ésta está integrada por los 7 primeros segmentos occipitales. Las
células de la cresta neural cardiogénica migran ventralmente y,
siguiendo el trayecto de los arcos aórticos 3º, 4º y 6º ingresan en el
{kind=link}
tronco-cono del corazón, migran a través de él por debajo del
subendocardio y se introducen profundamente en el corazón (Fig. SC
5-4-2). Aportan células que forman transitoriamente parte de
las crestas troncoconalesy llegan hasta la porción membranosa
del tabique interventricular. Estas células acompañan la migración
de las fibras vagales que ingresan en el corazón y, además, forman
algunas de las neuronas parasimpáticas del corazón. En la región
ascendente de las grandes arterias, aorta y pulmonar, estas
células se diferencian en células musculares lisas vascularesde la
túnica media (SC 5.7. Las células de la cresta neural y de la población
celular proepicárdica (pcPE) en el desarrollo del corazón. Su
contribución al desarrollo de la vasculatura coronaria) (Fig. SC 5-4-
3).
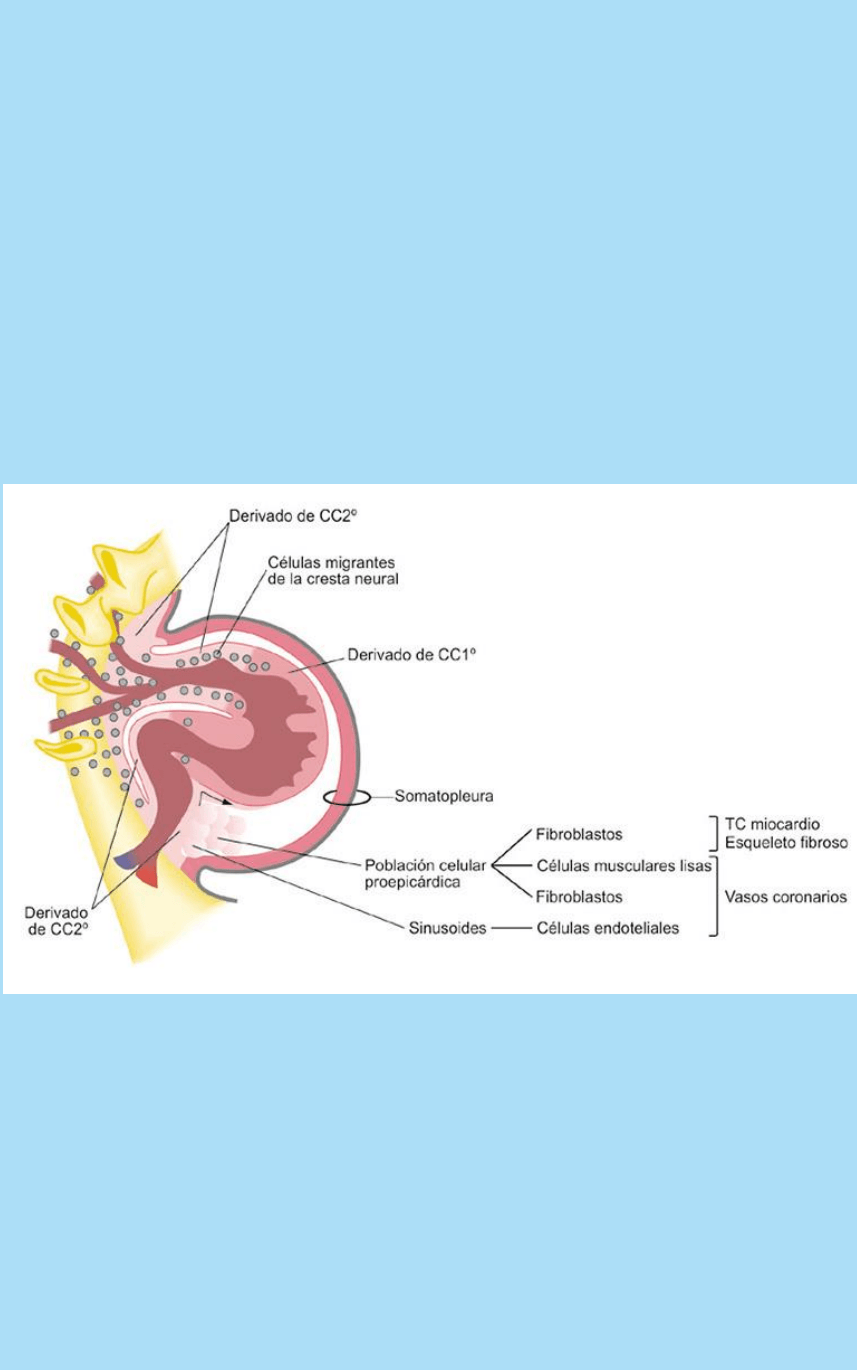
Fig. SC 5-4-3. Ilustra la relación entre posición de las crestas
cardiogénicas, los arcos aórticos a través de los cuales ingresan en
el corazón, las regiones cardíacas y los tipos celulares que originan.
4) La población celular proepicárdica es una población de células
mesenquimáticas aplanadas que forman parte del mesotelio,
el septum transversum, en el polo venoso del corazón (en el límite
con el seno venoso) (Fig. SC 5-4-2). Estas células invaden la
superficie del corazón a modo de una monocapa de células planas y
forman el revestimiento epicárdico del corazón. Junto con estas
células ingresan también células endoteliales que forman el
revestimiento endotelial de los vasos coronarios y el tejido conectivo
perivascular. Muchas de estas células se transforman en las células
musculares lisas de los vasos coronarios, fibroblastos perivasculares
o miofibroblastos. Además aportan una parte importante de
mesénquima que luego se diferencia en el tejido conectivo de la
mayor parte del miocardio (SC 5.5 El área cefálica del campo
{kind=link}

cardiogénico secundario y sus derivados anatómicos e histológicos).
Numerosas células que cumplen una función similar a éstas ingresan
también en el corazón a partir del mesénquima que rodea el polo
arterial del corazón
SC 5.5. El área cefálica del campo cardiogénico secundario y
sus derivados anatómicos e histológicos. V. Flores
La placa cardiogénica posee una porción amplia,
denominada campo cardiogénico primario, y una porción más
pequeña, de ubicación medial, aunque bastante variable dependiente
de las especies, denominada campo cardiogénico secundario.
Varios estudios de seguimiento de células marcadas supravitalmente
permiten mostrar que el campo primario origina el ventrículo primitivo
(ventrículo izquierdo), el canal auriculoventricular, la aurícula
primitiva (forma parte de las aurículas derecha e izquierda) y los
senos venosos (parte de la aurícula derecha). En el campo
cardiogénico secundario se han descrito dos o tres regiones
diferentes, según distintos autores. En principio, este campo posee
dos regiones: una superior oárea cefálica y una inferior o área
caudal. Algunos autores incluyen en esta última la población
celular proepicárdica (SC La formación de la población pcPE. Su
relación con el campo cardiogénico secundario. Mecanismo de
transferencia celular seno-venoso CTP en mamíferos).
Con respecto al área cefálica del campo cardiogénico secundario,
diversos estudios indican que el bulbo cardíaco del corazón tubular
primitivo (CTP), incluyendo sus tres porciones (ventrículo derecho,
los conos de salida de los ventrículos y el tronco arterioso), no
derivan del campo primario sino del área cefálica del campo
cardiogénico secundario. Durante la formación del CTP recto las
células del campo secundario de la placa cardiogénica se distribuyen
a lo largo de la zona media del piso de la faringe desde el 1º al 2º
arco branquial (hasta la región caudal del 2º arco), un poco por detrás
del extremo cefálico (tronco de salida) del CTP. Desde esta posición
se incorporan luego al CTP durante el fenómeno
denominado torsión en “C”.
En la primera fase de la torsión cardíaca o formación del asa
cardíaca (torsión en “C”), las células del campo cardiogénico
secundario, que formarán el bulbo, se despegan del mesodermo
ventral de la faringe y se agregan al extremo cefálico del CTP o
ventrículo primitivo. El bulbo ingresa así en el celoma pericárdico y
desvía la unión bulboventricular hacia la derecha y adelante
formando el asa bulboventricular que constituye precisamente el
fenómeno denominado “torsión en “C” (Fig. SC 5-5-1 A). La segunda
{kind=link}
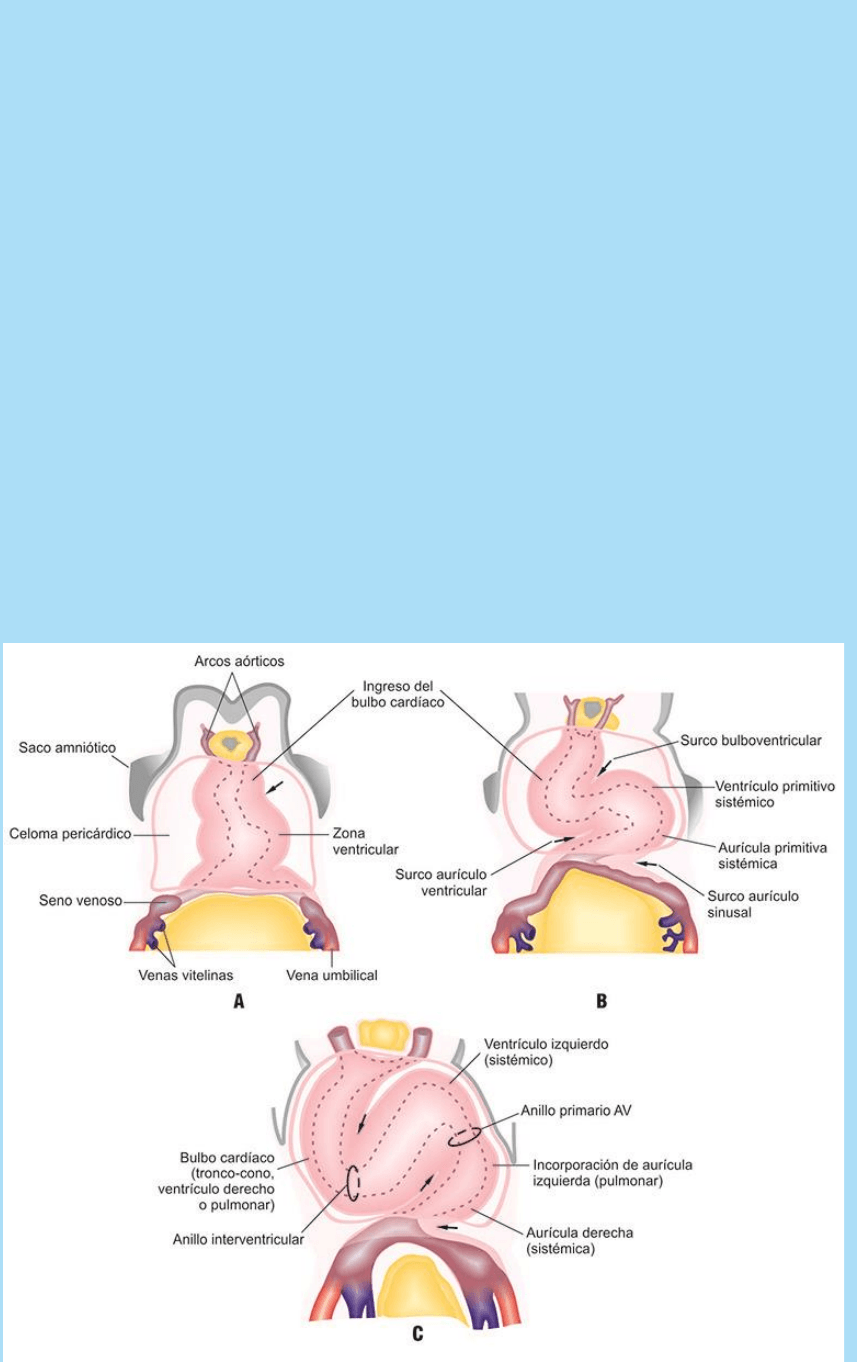
fase de la torsión, o torsión en “S”, ocurre más tarde, cuando al
extremo caudal del ventrículo primitivo se agrega la región auricular
izquierda (Fig. SC 5-5-1 B). Ésta ingresa en el celoma pericárdico
desplazando hacia la izquierda, hacia arriba y hacia el dorso la zona
de unión auriculoventricular. La formación de la torsión en “S”
conduce a la formación de otros dos surcos en el asa cardíaca: el
surco ventriculoauricular, entre el ventrículo y la futura aurícula
derecha, y el surco auriculosinusal, entre la aurícula izquierda y el
seno venoso izquierdo. La figura SC 5-5-1 muestra cómo la
formación de los tres surcos mencionados confieren la forma de “S”
que didácticamente se atribuye al asa cardíaca. Más tarde, toda la
región auricular asciende y se ubican dorsalmente a los ventrículos.
Con dichos cambios concluye la torsión del corazón tubular primitivo.
La formación del tracto de salida del corazón implica el agregado, en
forma sucesiva, de las regiones del bulbo cardíaco, el cono y el tronco
al extremo cefálico del CTP. Estas regiones se agregan al CTP antes
de la llegada de una oleada de células migrantes provenientes de
la cresta neural cardiogénica que también ingresan en el corazón
a través de su extremo cefálico y participan de la histogénesis
cardíaca y de los grandes vasos (aorta y pulmonar).
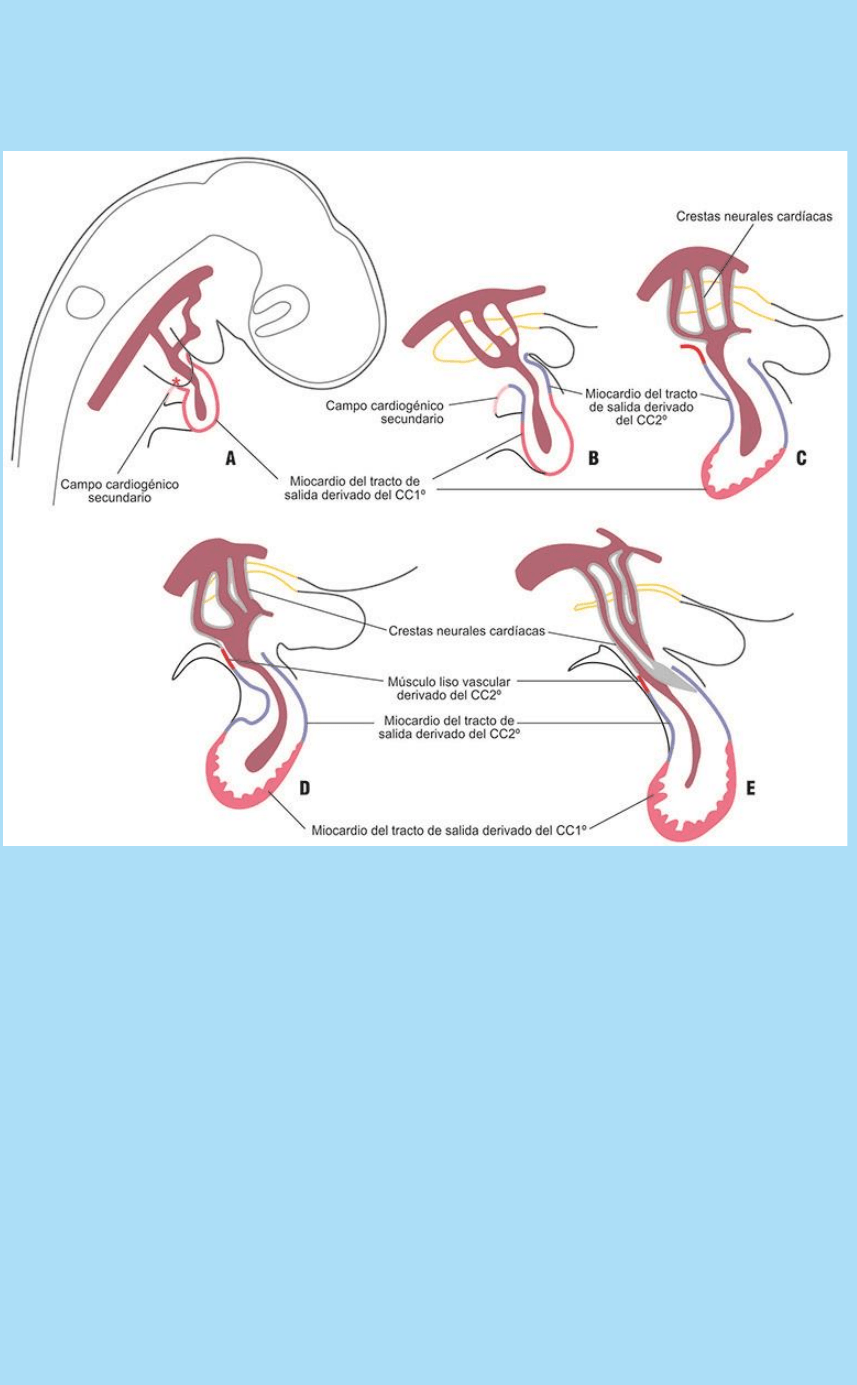
Fig. SC 5-5-1. A. Esquema de la torsión cardíaca en vista frontal. A.
La incorporación del campo cardiogénico secundario al CTP produce
la torsión en “C”. B-C. El agregado de la porción correspondiente a la
aurícula izquierda produce la torsión en “S”.
Fig. SC 5-5-2. El miocardio del tracto de salida (bulbo y cono) deriva
del mesodermo esplácnico adyacente al extremo cefálico del CTP
recto (Modelo; Embrión de pollo). A. Diagrama del estado 14.
Muestra la ubicación del campo del tracto de salida secundario
(asterisco rojo) que genera el bulbo, el cono y el tronco. El campo se
halla en relación con el segundo arco branquial. En este estado se
inyecta un marcador celular supravital en la región del campo del
tracto de salida secundario. B-F. Se ilustra cómo las células del
campo secundario se van incorporando, como bulbo, cono y tronco,
al extremo cefálico del CTP. Durante dicho proceso el sitio de unión
a los arcos aórticos se corre desde el 1er arco hasta el 4º o 6º arco
aórtico. Se indican los tejidos derivados del CTP recto (CC1º) y los
que derivan del campo cardiogénico secundario (CC2º).
Se considera que, en la especie humana, la zona en la que las
paredes ventriculares se continúan con las arterias aorta y pulmonar
constituye una zona de mezcla de poblaciones celulares diferentes
{kind=link}
Este documento contiene más páginas...
Descargar Completo
Cap 16 SC Flores.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.