SC 14.1. Vías de señalización y factores de transcripción
involucrados en la compartimentalización del somita y su
ulterior evolución. V. Flores
Los somitas se hallan sometidos a redes de señalización
espacialmente organizadas que influyen diferentemente sobre las
células de sus distintas regiones. Ello hace que un somita posea
compartimentos o dominios celulares diferentemente especificados
que cumplen distintas funciones de desarrollo (esclerotomo,
miotomo, dermatomo, sindetomo, etc.).
Una vez que el somita madura, las células de sus diferentes regiones
realizan diferentes tipos de comportamientos y migran a diferentes
regiones corporales. Este proceso de migración está precedido de
una T e-m que las regiones del somita sufren en distintos momentos.
La compartimentalización del somita depende de la acción de
señales provenientes de centros organizadores de posición dorsal,
ventral, medial y lateral que habitualmente poseen efectos
contrapuestos. La figura SC 14-1-1. ilustra la complejidad y la
organización espacial de la red de señalización involucrada en la
compartimentación del somita.
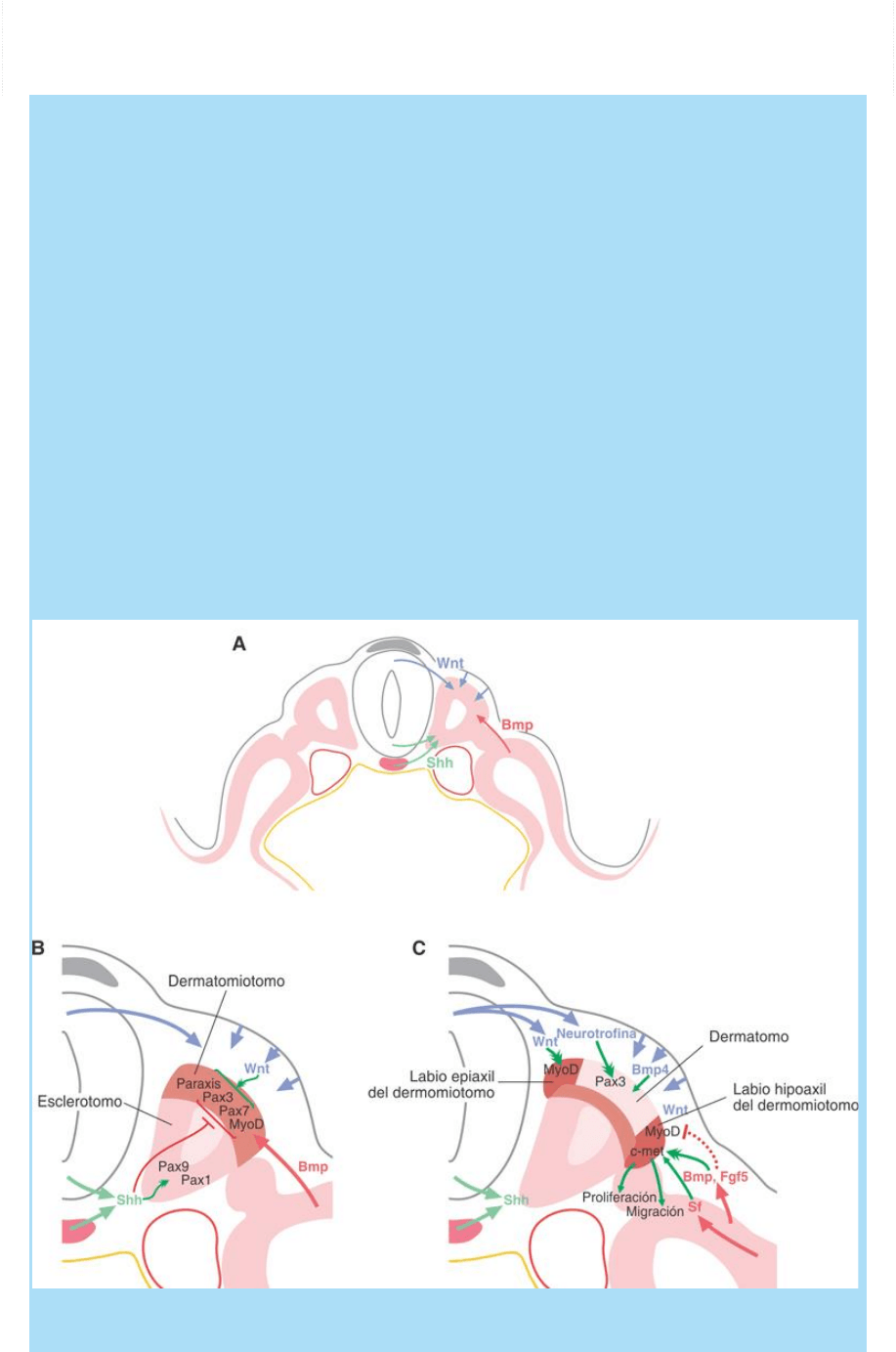
Fig. SC 14-1-1. Ilustra las principales poblaciones celulares que
conforman el “entorno” del mesodermo paraxil y que actúan como las
{kind=link}

poblaciones celulares organizadoras que generan la red de
señalización celular que instalan el patterning de los somitas. Las
flechas indican las diversas influencias locales recibidas por las
células de las diversas regiones del somita. Véase descripción en el
texto.
El patrón morfogenético D-V es instalado interactivamente entre
la señalización vía Wnt (desde el ectodermo superficial dorsal y el
tubo neural dorsal) y la señalización vía Shh y Noggin (desde el
mesodermo axil ventral).
El patrón morfogenético M-L es instalado interactivamente entre
la señalización vía Shh proveniente de la notocorda y placa basal
del tubo neural y la señalización vía Bmp proveniente del
mesodermo somatopleural.
Estas distintas influencias hacen que en el somita aparezcan
regiones diferentemente especificadas.
La especificación del esclerotomo. El esclerotomo es especificado
por varias señales mediales y ventrales provenientes de la notocorda
y placa del piso del tubo neural. Depende principalmente de la
activación de la vía Shh que promueve, o al menos mantiene, la
expresión del factor de transcripción Pax1. Otro blanco de la vía
Shh es el factor de transcripción Pax9que también se expresa en
el esclerotomo. La activación de la vía Shh también contribuye
al patterning D-V inhibiendo la expresión de factores de
transcripción dorsales como Pax3, Pax7 y MyoD que se expresan
en el dorso (dermatomiotomo).
La especificación del dermatomiotomo. La especificación del
dermatomiotomo requiere la proteína señal Wnt proveniente del
ectodermo y del tubo neural dorsales que estimulan la expresión de
los factores de transcripción Pax3, Pax7 y Paraxis. El
dermatomiotomo luego se segrega, debido a influencias adicionales,
en miotomo y dermatomo.
La especificación del dermatomo. La especificación del
dermatomo dependería, por un lado, de la proteína señal
neurotrofina que es secretada por la placa del techo del tubo neural
y que estimula la expresión del factor de transcripción Pax3 y, por
otro, de la señal Bmp4 secretada por el ectodermo adyacente. Una
vez especificado por la acción de dichas señales se inicia la
migración de las células del dermatomo que recubre la superficie
basal del ectodermo de la región dorsal. Estas células ulteriormente
se diferenciarán en el tejido conectivo de la dermis dorsal.
La especificación del miotomo. La especificación del miotomo
involucra, por un lado, a) su especificación en sentido muscular, por

otro, b) su especificación como epímero o hipómero y, además,
sufren luego una c) T e-m que permite su segregación o
desprendimiento desde el dermomiotomo y los convierte en
mioblastos proliferantes migratorios.
La región del labio dorsal-medial o epiaxial del dermomiotomo,
debido a la acción de las señales dorsales mencionadas, expresa
tempranamente factores de transcripción miogénicos de la
familia MyoD de cuya expresión depende, al parecer, su
especificación como epímero (subpoblación precursora de
mioblastos que originan músculos dorsales o extensores del raquis).
Por otro lado, la región del labio ventral-lateral o hipoaxial del
dermomiotomo, con el concurso de la activación de lavía de
señalización del Bmp, secretado a partir del mesodermo
somatopleural, se especifica en hipómero (subpoblación precursora
de mioblastos que forman los músculos anterolaterales [flexores del
raquis, los de los miembros, el diafragma y la lengua]). El mesodermo
somatopleural también secreta el factor de crecimiento
fibroblástico 5 o Fgf5.
El efecto de la estimulación del miotomo por el mesodermo
somatopleural, por un lado, retarda la expresión de los factores de
transcripción miogénicos del tipo MyoD (que se expresan
tempranamente en el dorso) y, por otro, estimula la expresión de
la proteína receptor c-met. Este receptor habilita a las células que
lo expresan a responder al factor de crecimiento de hepatocito
(Hgf/Sf) que estimula su migración. Esta proteína señal es emitida
por el mesénquima del esbozo de miembro. Así, estas señales serían
las responsables de estimular la expansión proliferativa y migratoria
de las células del hipómero hacia la somatopleura y hacia los
esbozos de los miembros.
El proceso de migración dirigida de los mioblastos embrionarios,
antes de iniciar su diferenciación, requiere una T e-m previa y una
amplificación y expansión proliferativa de la población. En la
estimulación de esta expansión proliferativa participan también
las proteínas señal factores de crecimiento fibroblástico y
transformante tipo beta (Fgf y Tgf-β).
Cuando los mioblastos cesan su proliferación y se transforman en
mioblastos posmitóticos, inician la síntesis de los factores de
transcripción miogénicos del tipo MyoD. Estos factores de
transcripción inician una cascada de expresión de diversas proteínas
que permiten el avance a través de las diferentes fases de la
citodiferenciación del miocito esquelético. Los factores de
transcripción MyoD tienen como función modificar el patrón de
actividad génica de modo que ésta sea, en sucesivos pasos, dirigida

a la activación de los genes necesarios para la síntesis de las
diversas proteínas específicas del miocito. Los factores de
transcripción miogénicos son a su vez modulados por otras proteínas
que exacerban o disminuyen su actividad.
Además de estos hechos, vinculados a la citodiferenciación, también
se pone en juego la expresión de otros genes del tipo Hox,
involucrados en la organización espacial de la diferenciación de los
diversos grupos musculares del cuerpo. Los factores de
transcripción Hoxa-11 y 13 se hallan involucrados en la represión
transitoria de MyoD durante el desarrollo temprano de los músculos
de las extremidades.
SC 14.2. El esbozo de miembro como modelo de campo
morfogenético I. Asignación de información posicional y
programación temporoespacial de la morfohistogénesis. V.
Flores, M. Rapacioli
El esbozo de miembro aparece hacia el final del período somítico (en
la especie humana, durante la 5ª SD) como una pequeña
prominencia en la región lateral del embrión. Dicha prominencia está
formada por un acúmulo de células mesodérmicas revestidas por el
ectodermo epidérmico. En el ectodermo del extremo distal del esbozo
se forma tempranamente un engrosamiento lineal,
denominado cresta apical ectodérmica (Cae), a lo largo del límite
entre las superficies dorsal y ventral del esbozo. Luego se produce la
elongación del esbozo, demarcándose en él dos zonas que, en el
caso del miembro superior, corresponden a la mano (la distal) y al
brazo (la proximal).
En el mesénquima del esbozo se distinguen una zona externa,
compacta y pobremente vascularizada, y una zona central laxa.
Cuando el esbozo crece en longitud, la parte central se transforma
en una condensación mesenquimática que indica el inicio de la
formación de cartílago. La formación de condensaciones
mesenquimáticas progresa en sentido distal hasta la aparición de los
precursores cartilaginosos de los dedos de la mano. El desarrollo
subsiguiente incluye la formación de los huesos, el crecimiento de
éstos en longitud (por adición epifisaria), el crecimiento de los dedos,
la pérdida de tejido interdigital, etc., hasta la diferenciación de todas
las regiones del miembro. Durante este proceso, el esbozo de
miembro es invadido por otros tejidos (mioblastos, nervios, vasos,
etc.).
El esbozo de miembro, un modelo de campo morfogenético. El
esbozo de miembro, en el momento de su formación, posee

propiedades de campo morfogenético (SC El patrón como sistema
de referencia que asigna información posicional dentro de un campo.
El modelo de la bandera francesa; SC 3.4. Concepto de campo
morfogenético). Entre estas propiedades se cuentan las siguientes:
a) Es autodiferenciante, es decir, posee la capacidad de elaborar su
patrón de organización estructural básico en forma independiente de
las otras estructuras. El esbozo de miembro trasplantado a otra
región del embrión o a un medio de cultivo, se desarrolla y forma la
estructura completa. Ello indica que en él radican los elementos
informativos y estructurales necesarios para desarrollar,
interactivamente, la estructura completa.
b) Posee capacidad regulativa, vale decir, capacidad para formar
una estructura completa ante excesos o déficits en las poblaciones
celulares que lo componen. El esbozo de miembro se comporta como
un sistema de regulación hasta un estado del desarrollo
relativamente tardío. Algunos experimentos realizados en el embrión
de pollo (estados 19-20) en los que se elimina 30- 80% del
mesénquima del centro del esbozo miembro, dan como resultado
miembros normales. Otros experimentos similares realizados desde
el estado 22 en adelante dan como resultado miembros con diversos
déficits. A medida que avanza el desarrollo, la capacidad regulativa
disminuye (para la lectura detallada de estos experimentos véanse
referencias:
- Zwilling E. (1961). Limb morphogenesis. Advan
Morphogenesis 1:301-30. - Bodemer C. (1968); Inductive Interactions
and Progressive Determination. In: Modern Embryology.
Holt,Rinehart & Winston of Canada Ltd. - Hamburger V. (1938).
Morphogenetic and axial self-differentiation of transplanted limb
primordia of 2-day chick embryos. J Exp Zool 77:379-99. - Hampé A.
(1959). Contribution to the study of the development and the
regulation of deficiencies and excesses in the feet of chick
embryos. Arch Anat Microsc Morphol Exp 48:345-478. - Amprino R,
Camosso M. (1959). On the role of the "apical ridge" in the
development of the chick embryo limb bud. Acta Anat (Basel) 38:280-
8. - Saunders JW Jr. (1948). The proximo-distal sequence of origin of
the parts of the chick wing and the role of the ectoderm. J Exp
Zool 108(3):363-403.
La capacidad regulativa tiende a la formación de una estructura
completa y armónica. Si experimentalmente se divide un esbozo de
miembro inferior en tres porciones (proximal, media y distal), la
porción media, cultivada en la membrana corioalantoidea, origina una
articulación femorotibial. Si las porciones proximal y distal se
fusionan, ambas forman un miembro con articulación femorotibial,

aunque sin peroné. Estos resultados se obtienen sólo durante un
período de tiempo.
c) El esbozo de miembro experimenta un proceso de determinación
progresiva, vale decir, sus distintas regiones se van determinando y
diferenciando gradualmente en el tiempo.
La Cae situada en el extremo distal del miembro es esencial para su
desarrollo. Si se extirpa la Cae, el desarrollo del miembro se detiene.
El resultado de la eliminación de la Cae depende del momento en
que se realiza la extirpación. Cuanto más temprano se realiza la
extirpación mayor es la cantidad de regiones faltantes. Vale decir,
cuanto más tarde se elimina la Cae, más segmentos posee el
miembro terminalmente desarrollado. Esto sugiere que las
estructuras se determinan y desarrollan según una secuencia
temporal y próximo-distal.
Los miembros superiores e inferiores son entidades diferentes. A su
vez, cada uno de ellos está compuesto por varias regiones
anatómicamente diferentes. Sin embargo, los tejidos y los tipos
celulares que los constituyen son los mismos. Ello implica que los
procesos de determinación celular y citodiferenciación involucrados
en el desarrollo son básicamente similares. Así, una explicación de
las diferencias estructurales entre miembros superiores e inferiores o
de las diferencias entre sus regiones no se funda en diferencias en la
citodiferenciación sino en el modo como los procesos de
determinación y diferenciación se organizan en el espacio. Dicho
fenómeno se denomina “patterning” y depende del modo como los
CCD se organizan temporal y espacialmente. Ello implica que el
proceso de patterning se ejecuta diferentemente en cada miembro y
en cada una de sus diferentes regiones.
La existencia de un patterning remite a la existencia de procesos de
control epigenético, tanto temporales como espaciales, de los CCD.
El proceso de patterning, en consecuencia, requiere sistemas de
referencia espacial y temporal que regulan la operación organizada
de las combinatorias de CCD involucrados en el desarrollo de una
cierta estructura.
Se ha propuesto que el patterning es un proceso multipaso que
implicaría varias fases:
a) Constitución del esbozo o campo morfogenético de la estructura
global, en este caso, el campo del miembro superior o inferior. En
esta fase se asocian las poblaciones mesenquimáticas y
ectodérmicas que constituyen el campo y, como consecuencia de
procesos de señalización instalados por poblaciones organizadoras
exteriores al campo, éste se determina. La determinación se refiere

al nivel de organización correspondiente al “miembro”, pero no a sus
niveles subalternos (regiones y subregiones del miembro).
b) La formación del esbozo de miembro implica también la
constitución de 1) subpoblaciones celulares con función informativa
(integran centros señalizadores internos del esbozo de miembro,
como la zona de actividad polarizante [zAP]) y 2)la subpoblación con
función estructural (células sensibles a las señales generadas dentro
del campo y que elaboran su estructura).
c) Instalación del pattern o gradiente de morfógeno y asignación
de información de posición. Este es un proceso de señalización
celular espacialmente organizado por medio del cual, por un lado, las
células informativas (de los centros señalizadores del campo)
generan gradientes de morfógenos y, por otro, las células con función
estructural, que ocupan diferentes posiciones dentro del campo, vale
decir, sometidas a diferentes concentraciones de morfógeno,
adquieren la propiedad denominada información posicional.
d) La adquisición de información posicional implica experimentar
diferentes tipos de reprogramaciones epigenéticas (especificaciones
lábiles) que habilitan, a las células que ocupan diferentes posiciones
del campo, a ejecutar diferentes modos de organización de los CCD
que participan de la morfogénesis y la histogénesis. La adquisición
de diferente información posicional (en distintas regiones del esbozo)
implica que las células diferentemente posicionadas, pese a retener
transitoriamente similar potencia citogenética, son no
equivalentes en el sentido de que ejecutarán diferentemente los
procesos morfogenéticos e histogenéticos y, en consecuencia,
generarán diferencias estructurales regionales en el miembro.
e) Finalmente, las subpoblaciones celulares no equivalentes,
diferentemente posicionadas en el campo, sufren un proceso de
determinación. Vale decir, las reprogramaciones epigenéticas
sufridas son fijadas irreversiblemente. En dicho momento, el campo
deja de existir como tal y pasa a constituirse como un sistema
mosaico.
f) El paso final consiste en ejecutar los procesos de morfogénesis e
histogénesis acordes con las programaciones del desarrollo
determinadas para cada región y, en consecuencia, la elaboración de
los diferentes niveles de organización del fenotipo del miembro.
Durante esta fase, el pattern informativo inicial y las diferencias en la
información posicional del campo, se “traducen” en el patrón de
organización estructural típico de cada región.
Desde el punto de vista filogenético, una de las implicaciones teóricas
del modelo de establecimiento de patrones basado en asignación de
información de posición por medio de gradientes de morfógenos, es

que esta estrategia parece hallarse bastante difundida en la
naturaleza. Así, no sólo las estrategias sino también las señales que
participan en el establecimiento del patrón de esbozos de miembro
en general podrían ser eficaces y compartidas entre diferentes
especies, aun relativamente poco emparentadas. Las diferencias
anatómicas entre miembros de distintas especies se deberían a
diferentes modos de interpretar una misma señal simple con función
morfogenética. Algunos experimentos en los que se han realizado
trasplantes de zAP entre distintas especies de vertebrados han
demostrado que la zAP de una especie puede ser biológicamente
eficaz en otra especie. En los vertebrados, la señal parece ser la
misma y lo que ha cambiado durante la evolución es, al parecer, el
modo de ejecutar la respuesta a la señal.
SC 14.3. El esbozo de miembro como modelo de campo
morfogenético II. La asignación de información posicional a lo
largo del eje próximo-distal. V. Flores
Existen resultados experimentales que pueden ser interpretados en
el sentido de que la organización espacial de los procesos
involucrados en la morfogénesis e histogénesis del esbozo de
miembro se realiza sobre la base de al menos tres sistemas de
referencia espacial. Cada uno de ellos se vincularía a los tres ejes
del espacio que habitualmente se utilizan como referencia para
describir los miembros: a) un eje próximo distal (de hombro a punta
de los dedos), b) un eje céfalo-caudal o anteroposterior (de dedo
pulgar [primer dedo] a dedo meñique [quinto dedo]) y c) un eje dorso-
ventral (de dorso a palma de la mano).
De acuerdo con esto, se podría suponer que la morfohistogénesis en
el esbozo de la extremidad se organiza sobre la base de CCD que se
hallan sometidos a un sistema de referencia espacial que tendría las
características de un sistema tridimensional de coordenadas
ortogonales. La información disponible sugiere que esto no es
necesariamente así sino que la información posicional es asignada
de un modo más simple, como si la información referida a los tres
ejes mencionados fueran asignados por tres sistemas de referencia
lineales o unidimensionales, con modalidades diferentes de
asignación de información posicional.
El crecimiento en longitud del miembro en gran medida depende de
la actividad proliferativa de las poblaciones celulares
mesenquimáticas y ectodérmicas que ocupan la región distal del
miembro. La actividad proliferativa en dicha región va generando
subpoblaciones de células que se van agregando al extremo distal
del miembro en crecimiento en función del tiempo. Por tal motivo a
dicha región se la denomina zona de crecimiento. Esa zona está
ocupada por una población celular mesenquimática autorrenovante,
denominada zona de progreso (zP), que se halla envuelta por una
placoda o engrosamiento ectodérmico denominado cresta apical
ectodérmica (Cae). El mantenimiento de ambas poblaciones se
realiza interactivamente por medio de procesos de señalización
celular recíproca entre ambas. Por un lado, la formación de la Cae
depende de señales generadas en la zP y, por otro, la Cae mantiene
a la zP. La zP ocupa una región espacial discoidal, de sólo unos 300
micrómetros, restringida la zona subectodérmica de la Cae. Su
extensión depende del alcance (zona de influencia o rango de acción)
de las señales generadas en la Cae.
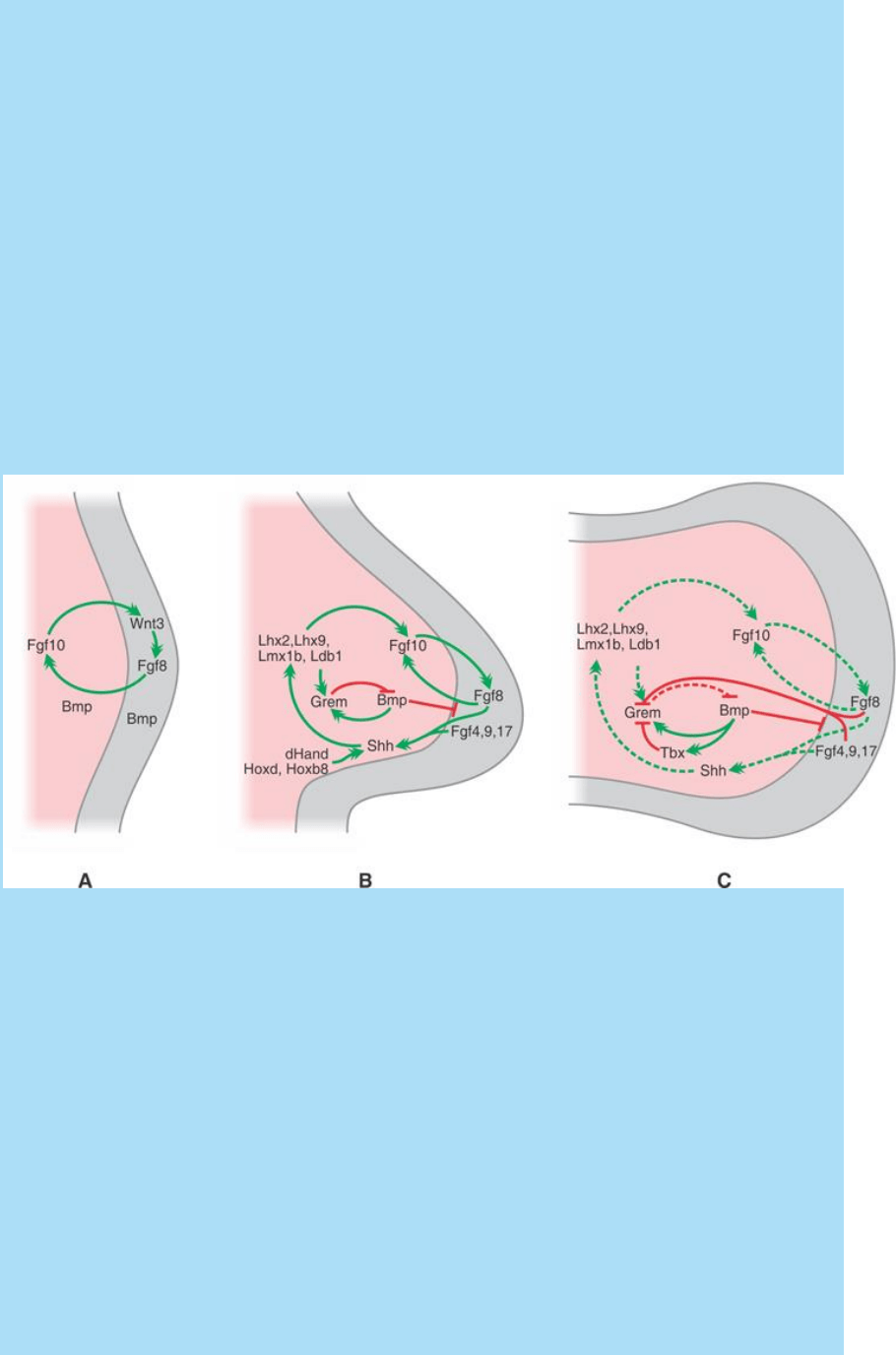
Se postula la siguiente sucesión de interacciones (Fig. SC 14-3-1):
Fig. SC 14-3-1. A. Inducción de la Cae en la región ventral del anillo
ectodérmico. B. Aparición de la zAP y fase de crecimiento del esbozo.
C. Fase de finalización del mantenimiento de la Cae y la zAP. En
todos los casos se indican las principales vías de señalización y
algunos de los principales factores de transcripción que son
potenciales efectores de dichas vías. Descripción en el texto.
Inducción de la Cae en la región ventral del anillo ectodérmico.
El Fgf10 secretado por el mesénquima promueve en el ectodermo la
expresión de Wnt3. La activación de la vía Wnt3/Beta catenina
estimula la expresión del Fgf8 en el ectodermo. El Fgf8 promueve la
expresión de Fgf10 en el mesénquima y así se constituye un circuito
retroalimentación positiva (estimulación recíproca). La formación de
la Cae requiere, además, la acción de la Bmp secretada por el
mesénquima y el ectodermo ventral.
{kind=link}

Aparición de la zona de actividad polarizante (zAP) y fase de
crecimiento del esbozo. En la constitución de la zAP (véase SC
14.4. El esbozo de miembro como modelo de campo morfogenético
III. La asignación de información posicional a lo largo del eje cefálo-
caudal (anteroposterior)) participan Fgfs secretados por la Cae y
factores de transcripción dHand, Hoxd y Hox8b. La zAP secreta Shh
que estimula la expresión de factores de transcripción que mantienen
la expresión del Fgf10 y de la proteína gremlina 1 (Grem1). Así,
durante la fase de crecimiento del esbozo, por un lado, se mantiene
el ciclo de estimulación recíproca entre Fgf10 y Fgf8 y, por otro, se
crea un ciclo de retroalimentación positiva (estimulación recíproca)
entre la Cae y la zAP que se mantienen mutuamente. Además se
bloquea la acción de la Bmp, que inhibiría a los Fgf de la Cae.
Fase de finalización del mantenimiento de la Cae y la zAP. Hacia
el final del desarrollo del esbozo, la alta actividad de los Fgf
sobrepasaría un umbral e inhibiría, directa o indirectamente, la
actividad de la gremlina. Así, las Bmp inhibirían la acción de los Fgf
de la Cae. Ésta involucionaría y ya no mantendría a la zAP. Otra
hipótesis postula que las Bmp estimularían la expresión del factor de
transcripción Tbx y que éste inhibiría la expresión de la proteína
gremlina.
Algunos datos experimentales indican que el proceso de asignación
de información posicional a lo largo del eje próximo-distal está
asociado a la generación, en forma sucesiva, de las subpoblaciones
celulares que integrarán las diferentes regiones del miembro desde
la zona proximal (hombro) a la distal (extremo digital). Esta
asignación se establecería en forma dependiente del tiempo de
permanencia de las células en la zP.
En la zona de crecimiento del miembro existen señales
correspondientes a vías de señalización que posibilitan que las
células de dicha región, por un lado, mantengan una intensa actividad
proliferativa asimétrica y, por otro, experimenten sucesivas
reprogramaciones epigenéticas.
El mecanismo que se propone es el siguiente (Fig. SC 14-3-2):
{kind=link}
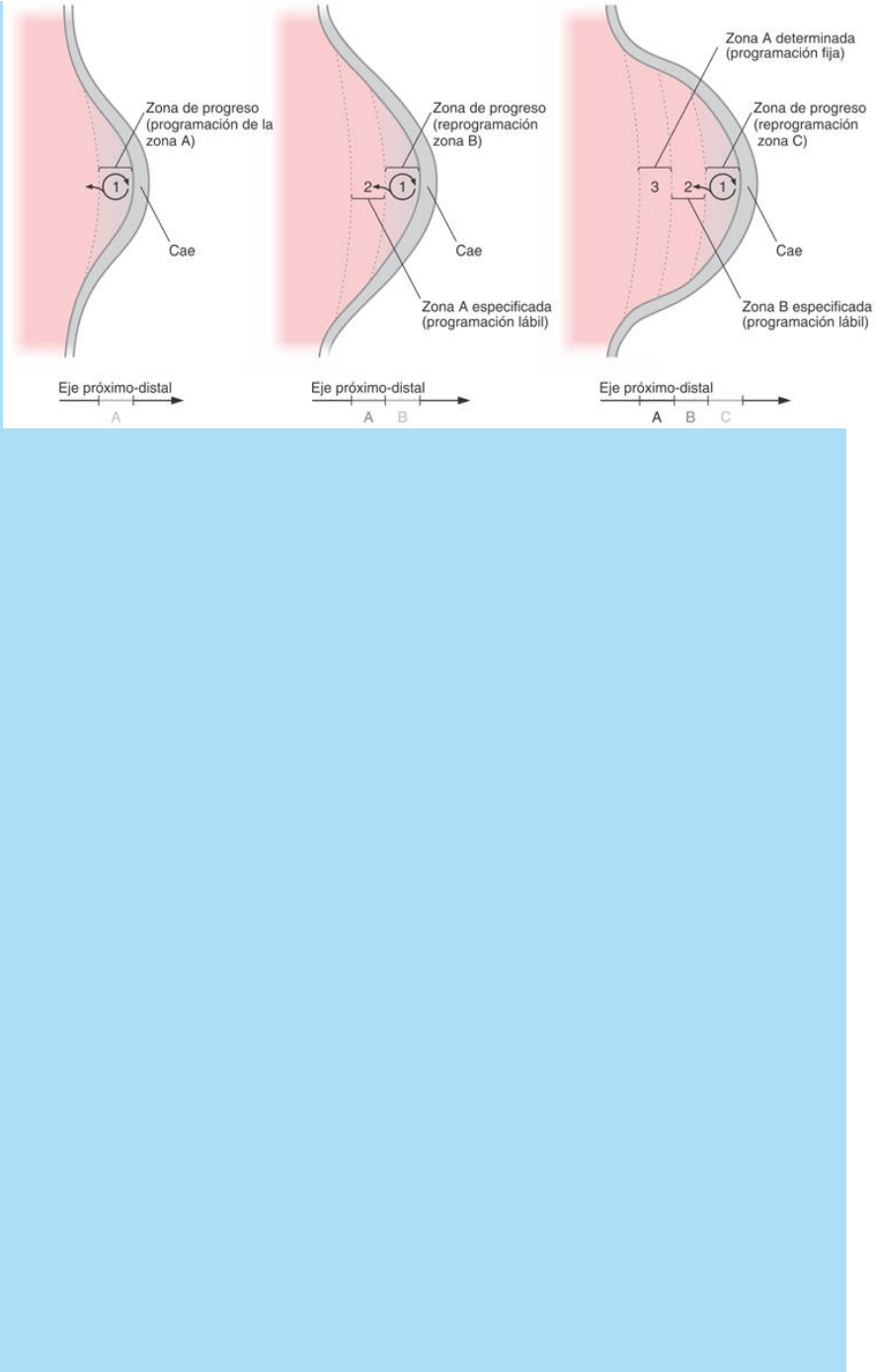
Fig. SC 14-3-2. Modelo de asignación de información posicional a lo
largo del eje próximo-distal. Las diferentes zonas del eje próximo-
distal (A, B y C) van pasando por diferentes estados: 1) de
programación/reprogramación, 2) de especificación o programación
lábil y 3) de determinación o programación fija. Descripción en el
texto.
a) Dada la intensa actividad proliferativa en la zP, esta población se
halla en expansión permanente y ocupa un espacio cada vez mayor.
b) Dado que las características que definen la zP dependen de
fenómenos de señalización que operan exclusivamente en la zona
de interacción con la Cae, las células que adquieren posiciones que
van más allá de dicho rango de acción pueden ser consideradas
como células que abandonan la zP.
c) De esta forma, diversas subpoblaciones celulares abandonan la
zP en función del tiempo o en función del número de ciclos
proliferativos. Naturalmente, las células que abandonan la zP
tempranamente generan regiones del miembro que son proximales
con respecto a las regiones generadas por células que abandonan la
zP en períodos más tardíos.
d) Las células que sucesivamente, en función del tiempo, abandonan
la zP quedan fuera del rango de acción de las señales que regulan
sus funciones de desarrollo, en consecuencia: modifican su dinámica
proliferativa y quedan con la programación epigenética
correspondiente al momento en que abandonaron la zP.
e) De las diferentes programaciones epigenéticas con la que las
células abandonan la zP dependen los diferentes procesos
morfohistogenéticos de las regiones correspondientes a diferentes
posiciones del eje próximo-distal. Cuando las células abandonan la
zP, dicha programación es lábil, vale decir, las células pueden ser

reprogramables, pero pasado cierto tiempo, o llegado a una cierta
distancia respecto de la zP, el programa queda fijo en forma
irreversible. De esta forma se determinan en forma sucesiva en el
tiempo las subpoblaciones celulares que generan las diversas
regiones correspondientes al eje próximo-distal.
Debido al modo como ocurren los procesos descritos en los puntos
“a” a “e”, se propone que la determinación de regiones a lo largo
del eje próximo distal depende de un proceso de asignación de
información posicional dependiente del tiempo de
permanencia de las células en la zP. Debido a ello se establece
que el proceso de determinación es progresivo, vale decir, no se
determinan todas las regiones del eje simultáneamente, sino que
progresa en función del tiempo siguiendo la asimetría próximo
distal.
Así, la asignación de información de posición referida al eje próximo-
distal tendría lugar sólo dentro de la zP y las células que la
abandonan, en condiciones normales, ya no se reprograman. De esta
forma, las células mesenquimáticas que inicialmente ocupan el
esbozo, luego de los primeros ciclos proliferativos quedan fuera de la
zP y forman la región que articula el miembro con el tronco (hombro
o cintura escapulohumeral). Las que a continuación abandonan la zP
forman el brazo, luego el antebrazo, y así sucesivamente hasta
completarse las programaciones correspondientes a cada una de las
regiones anatómicamente distinguibles a lo largo del eje próximo-
distal.
El modo como se estructuran los eventos de desarrollo descritos más
arriba explicaría cómo la eliminación de la Cae, y la consiguiente
desaparición de la zP, deriva en una interrupción del desarrollo y de
la formación de las sucesivas regiones del miembro. Al eliminarse las
poblaciones mencionadas disminuye la proliferación y cesan las
sucesivas reprogramaciones que sólo ocurren en la zP.
El comportamiento arriba descrito también explica cómo, cuando se
extirpa la zP de un esbozo “joven” (en el que sólo se ha determinado
el segmento proximal 1 y en su lugar se trasplanta la zP de un esbozo
viejo en el que ya se han determinado los segmentos 1, 2 y 3, el
desarrollo del esbozo del miembro continúa con la formación de las
células correspondientes a los segmentos 4 y los más distales. Vale
decir, el resultado es la formación de un miembro con una
“discontinuidad anatómica” resultante del hecho de que faltan los
segmentos que hubieran sido especificados por la zP del esbozo

“joven”. Tal discontinuidad anatómica resulta de la “discontinuidad en
los valores de información de posición” instalada por el trasplante.
El comportamiento descrito en los puntos “a” a “e” explica también el
resultado obtenido en el experimento inverso: eliminación de la zP de
un esbozo de miembro “viejo” (en el que ya se ha asignado la
información de posición correspondiente a los segmentos 1, 2, y 3 y
su reemplazo por la zP de un esbozo “joven”, en el que recién se ha
asignado la información de posición correspondiente al segmento
proximal 1. En este caso, el resultado del intercambio de las zP lleva
a la formación de un miembro con una duplicación de algunas
regiones del eje próximo-distal: los segmentos 2 y 3 se hallan
repetidos debido a que la información de posición necesaria para su
formación se ha asignado dos veces.
Estos resultados confirman que la asignación de información de
posición a lo largo del eje pd depende de interacciones de corto
rango de alcance y reprogramaciones que se producen
autónomamente en las células de la zP. No se detecta acción alguna
de las células mesenquimáticas que ya han abandonado la zP sobre
las células de esta última. Vale decir, la zP realiza sucesivas
reprogramaciones epigenéticas, en función del tiempo, en forma
autónoma, independientemente de los tejidos que ya se han
determinado o de los que falta determinar.
Existen algunos resultados experimentales clásicos de irradiación de
esbozos de miembros seguidos de trasplantes que también darían
sustento al modelo planteado. La irradiación de esbozos de
miembros, con dosis subletales de raxos X, seguidos de su trasplante
a embriones no irradiados, da lugar a miembros en los que faltan
segmentos proximales y desarrollo normal de los distales. En estos
experimentos se constata que la extensión de la región proximal
faltante se relaciona con la intensidad de la dosis de radiación: cuanto
mayor es la dosis de radiación mayor es el número de segmentos
proximales faltantes. Una interpretación de estos resultados propone
que la radiación eliminaría un cierto porcentaje de células de la zP y
que a mayor dosis de radiación se produciría porcentaje mayor de
muerte celular. En estas circunstancias, las células sobrevivientes
deberían, primero, a) repoblar la zP hasta alcanzar el tamaño normal
y luego b) abandonar la zP con una información de posición “más
vieja” que la que hubieran recibido en condiciones normales. Así,
cuanto mayor es la dosis de radiación, menor es el número de células
sobrevivientes y mayor el número de ciclos proliferativos y el tiempo
de permanencia de las células en la zP. De esta forma, faltarían los
segmentos proximales cuyas reprogramaciones ocurren durante el

período de tiempo en el que ninguna célula abandona la zP y, las que
salen de la zP ya lo hacen con información correspondiente a los más
distales.
El modelo descrito proporciona algunas pistas sobre la posible
patogenia de ciertas malformaciones congénitas de las extremidades
como por ejemplo la focomelia. La droga talidomida administrada a
una mujer gestante causa focomelia, entre otras anormalidades. Se
desconoce su modo de acción, pero estos resultados sugieren que
de alguna manera podría actuar de manera similar a la radiación
destruyendo las células del esbozo temprano de la extremidad.
SC 14.4. El esbozo de miembro como modelo de campo
morfogenético III. La asignación de información posicional a lo
largo del eje cefálo-caudal (anteroposterior). V. Flores
Los esbozos de los miembros, tanto anteriores como posteriores, se
constituyen por medio de la asociación del mesénquima
somatopleural con el ectodermo. Varios datos indican que, de los dos
componentes del esbozo, el que primero adquiere la especificidad de
esbozo de miembro es el mesénquima somatopleural. Tal
especificidad es adquirida como consecuencia de señales originadas
en poblaciones celulares circundantes a las regiones en las que los
esbozos se forman.
Los sitios de formación de los esbozos superiores e inferiores están
definidos con bastante precisión: a) ocupan una zona definida del eje
céfalo-caudal del embrión y b) a lo largo de la zona frontera entre las
regiones dorsal y ventral del embrión. El mesénquima del esbozo,
una vez asignada la identidad de componente del miembro, prolifera
activamente y el esbozo sobresale, como una pequeña prominencia,
a lo largo del borde mencionado.
La adquisición del carácter de “mesénquima del esbozo de miembro”
produce, como manifestaciones más tempranas, a) un cambio en la
actividad proliferativa y b) la adquisición de la capacidad de “inducir”,
en el ectodermo suprayacente la formación de una placoda
ectodérmica. Dicha placoda ocupa, durante todo el desarrollo del
miembro, el extremo distal o apical del esbozo. Debido a ello, recibe
el nombre de cresta apical ectodérmica (Cae). La Cae se ubica
como un engrosamiento lineal del extremo del esbozo, a lo largo de
la frontera entre las superficies dorsal y ventral de éste.
La adquisición de identidad, por parte del mesénquima y la inducción
de la Cae, en el ectodermo, constituyen las primeras manifestaciones
del inicio de las interacciones necesarias para el desarrollo del
miembro. Dichas interacciones se mantienen hasta que quedan
constituidas todas las poblaciones celulares con la información de

posición necesaria para formar todas las regiones anatómicas del
miembro. Si tales interacciones son interrumpidas
experimentalmente, por eliminación de alguno de los dos
componentes (por ejemplo: extirpación de la Cae), el crecimiento del
miembro y la formación de sus diferentes regiones se detiene.
La asignación de información de posición referida al desarrollo del
miembro es un fenómeno que requiere la instalación de al menos tres
polaridades: las correspondientes a los tres ejes anatómicos sobre la
base de los cuales se organizan los tejidos (SC 14.2. El esbozo de
miembro como modelo de campo morfogenético I. Asignación de
información posicional y programación temporoespacial de la
morfohistogénesis; SC 14.3. El esbozo de miembro como modelo de
campo morfogenético II. La asignación de información posicional a lo
largo del eje próximo-distal). En esta síntesis nos ocuparemos
específicamente de algunos resultados experimentales que aclaran
algunos aspectos sobre cómo se instala la polaridad cf-cd y cómo se
asigna información de posición a lo largo de dicho eje espacial.
Un modelo muy difundido para el análisis de este fenómeno es el
esbozo del ala (miembro superior) del embrión de pollo y la formación
de los dedos que se distribuyen, precisamente a lo largo del eje cf-
cd. Tal eje, en el caso de la mano humana, se extiende desde el
pulgar al meñique.
En la región caudal del borde apical del esbozo ‒a lo largo del cual
se halla la Cae‒ del ala del pollo se ha detectado la existencia de una
pequeña población celular que, al ser extirpada, o ser trasplantada a
una región alejada del extremo del esbozo, determina que no se
formen los dedos. Varios resultados experimentales indican que
dicha región, denominada zona de actividad polarizante
(zAP) debido a que instala la polaridad cfcd, es un centro
señalizador que genera gradiente(s) de morfógeno(s).
El gradiente de morfógeno instalado por la zAP sería la entidad
informativa que asigna la información de posición referida a la
especificación de los diferentes dedos a lo largo del eje cf-cd. Así, tal
asignación dependería de la distancia, o la posición, que ocupan las
células del campo respecto de la zAP o, mejor, de la concentración
de morfógeno a la cual se hallan sometidas las células que ocupan
diferentes posiciones a lo largo del eje mencionado. Se postula que
la zAP produce y libera un morfógeno que, distribuyéndose en forma
de gradiente, proporciona el mecanismo para el establecimiento de
valores de información a lo largo del eje cf-cd. Según este modelo,
los distintos dedos estarían especificados por distintos umbrales al
factor morfógeno.
Este documento contiene más páginas...
Descargar Completo
Cap 16 SC Flores.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.