SC 11.1. La instalación y el refinamiento de la organización
espacial de competencias de desarrollo en el área preplacodal.
V. Flores, M. Rapacioli
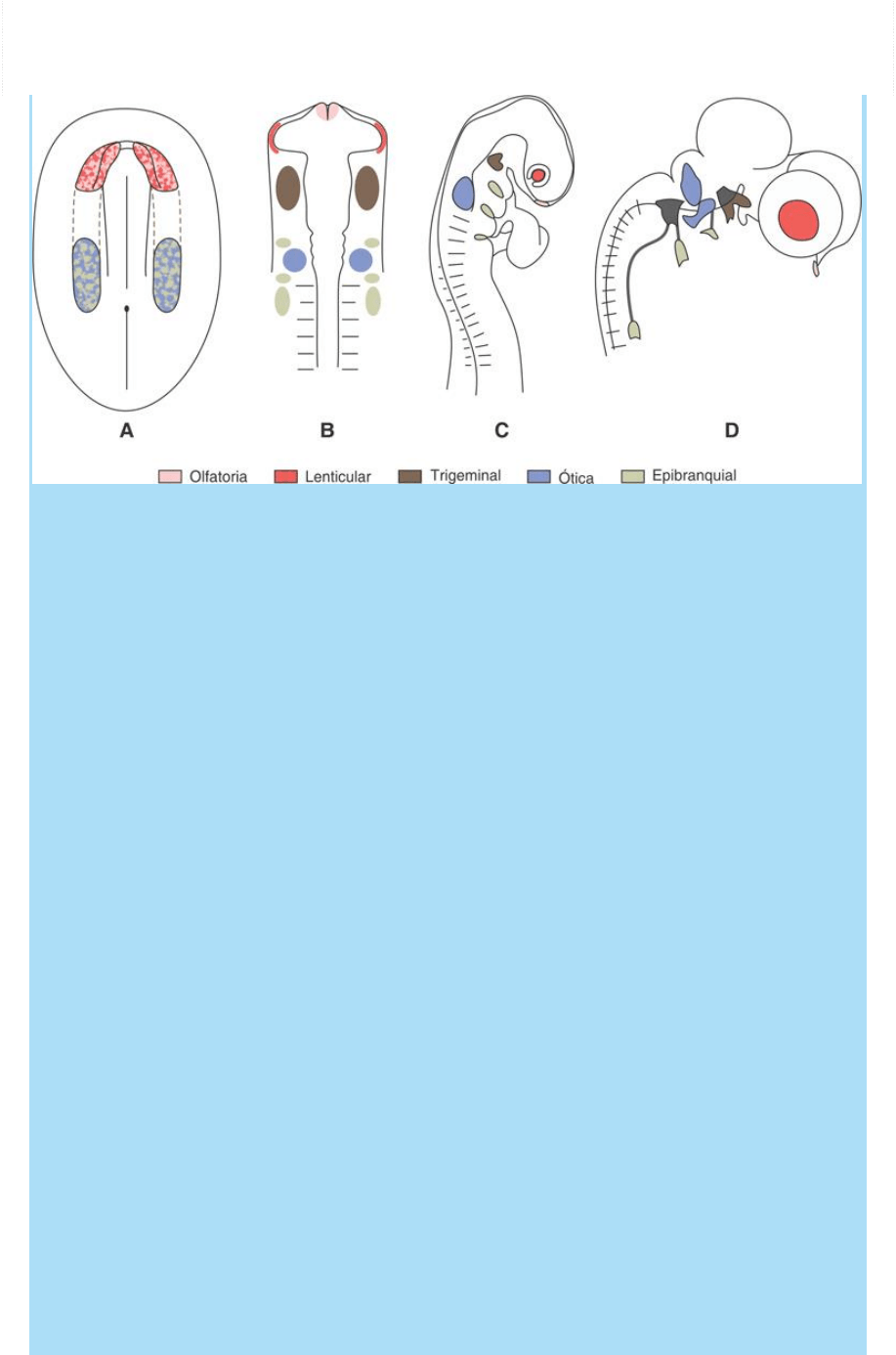
Fig. SC 11-1-1. La organización espacial de competencias del área
preplacodal (Modelo: Embrión de pollo). A. A fines de la gastrulación
se instalan zonas en las que coexisten células que pertenecerán a
distintas placodas. B-C. Estas zonas sufren luego un refinamiento en
la distribución espacial de las potencias de desarrollo
correspondientes a distintas placodas. D. A fines del período
somítico, las células derivadas de dichas placodas forman parte de
órganos sensoriales periféricos o ganglios sensoriales craneales.
Se denomina área preplacodal a la región ectodérmica, de fines de
la gastrulación o de principios del período somítico, que bordea, a
modo de herradura, el extremo cefálico de la placa neural. Se trata
de una región estrecha en la que coexisten competencias para
responder a diversas señales provenientes de tejidos adyacentes y
determinarse en diferentes tipos de poblaciones celulares
ectodérmicas. Entre estas poblaciones celulares se hallan las que
forman engrosamientos ectodérmicos (placodas) constituidas por
células con una potencia de desarrollo restringida a la formación de
algún órgano sensorial en particular. Algunas de estas placodas
también tienen una potencia neuronogénica, vale decir, una potencia
citogenética que incluye la generación de poblaciones de neuronas
sensoriales primarias que integran diversos ganglios sensoriales
craneales.
Los esquemas de la figura SC 11-1-1 A-D ilustran cómo, según
progresa el desarrollo, el área preplacodal sufre un proceso de
refinamiento en la distribución planar de potencias de desarrollo.
Estas áreas se hallan inicialmente superpuestas pero, gradualmente,
se van separando de modo que hacia el final del período somítico,
{kind=link}

las poblaciones celulares con diferente determinación se hallan en
zonas no superpuestas y bien delimitadas.
La figura SC 11-1-1 ilustra este proceso tal como ha sido descrito en
el embrión de pollo. En etapa temprana del desarrollo (A: día 1 del
embrión de pollo; fin de gastrulación; inicio de período somítico;
1.
er
par de somitas) existe una zona cefálica y medial que bordea la
placa neural que es común para varias futuras placodas. En la región
más cefálica y medial, cerca del extremo de la placa neural, se hallan
las células que luego formarán las futuras placodas olfatoria y lental
(área roja y rosa). Más caudalmente, en la zona adyacente al primer
par de somitas, aunque ocupando un área de mayor extensión, se
halla un territorio preplacodal común a la placoda ótica o auditiva y a
las placodas epibranquiales. Entre las dos zonas mencionadas se
halla la región en la que se constituirá, un poco más tarde, el territorio
precursor de la placoda trigeminal.
Hacia el estado de 10 pares de somitas (B), algunas áreas se han
segregado y ahora ocupan diferentes regiones. Las células
correspondientes a las placodas olfatorias se localizan en el
ectodermo del extremo cefálico del embrión mientras que las de las
placodas lentales pasaron a ubicarse en las regiones laterales del
futuro diencéfalo. Es ésta una región crítica ya que se hallan
suprayaciendo a las vesículas ópticas con las que ulteriormente
deben interactuar para un desarrollo sincrónico y armónico de la lente
y la retina. Por otro lado, en este momento ya se han formado las
placodas trigeminales en el ectodermo adyacente al posencéfalo.
Puede observarse que las placodas óticas quedan ubicadas a la
altura de las rombómeras 4/5 y rodeadas de varios dominios
ectodérmicos correspondientes a otras tantas placodas
neuronogénicas epibranquiales.
Un poco más tarde, a mediados del período somítico, cuando ya se
formado la curvatura cefálica y las prominencias cardíacas (C), las
placodas están localizadas y reconocibles como zonas ectodérmicas
engrosadas asociadas típicamente a otras poblaciones celulares con
las que evolucionan integradamente. Hacia estados más tardíos,
comparables a la 5.ª SD humano (D), las placodas olfatorias lentales
auditivas han dado sus derivados y las neuronogénicas
epibranquiales han originado masas ganglionares de nervios
craneales (trigeminal, estato-acústico o cócleo-vestibular,
glosofaríngea, facial y vagal).

SC 11.2. La transformación del área preplacodal (área
competente de formación de placodas) en línea wolffiana (área
de placodas) del sistema sensorial cefálico. V. Flores
El concepto de línea de Wolff (línea wolffiana) alude a la existencia
de una zona que, con simetría bilateral, recorre la región cefálica del
embrión. La línea de Wolff se inicia en la línea media del proceso
frontal y recorre, a modo de arco, las caras laterales de la
prominencia frontal y, luego, las regiones laterales de la región
branquial (Fig. SC 11-2-1). La línea de Wolff es la zona en la que se
distribuyen, desde el período somítico, las placodas que son
precursoras de los neuroepitelios receptores periféricos de los
órganos de los sentidos o estructuras asociadas a los órganos de los
sentidos, como por ejemplo la lente del ojo. En algunas especies
(peces y algunos anfibios), la línea de Wolff se continúa, desde la
región branquial, en sentido caudal, a lo largo de casi toda la región
lateral del cuerpo y da origen al órgano sensorial denominado línea
lateral. Los receptores ubicados a lo largo de la línea lateral de
diferentes especies han sufrido adaptaciones divergentes que
permiten detectar distintos tipos de perturbaciones del ambiente
(movimientos o vibraciones del agua, campos eléctricos, campos
magnéticos, etc.). A través de esos estímulos es posible registrar
características relevantes del medio, detectar presas o depredadores
o comunicarse con otros individuos de la especie.
La línea de Wolff, descrita clásicamente en el extremo cefálico del
embrión del período somítico, es el resultado de la reubicación del
área que, en la actualidad, se describe como área preplacodal que
se constituye hacia fines de la gastrulación.
La figura SC 11-2-1 A-D muestra las transformaciones y cambios de
posición que experimenta el área preplacodal desde el momento de
su aparición, al final de la gastrulación, hasta el final del período
somítico. La figura SC 11-2-1 A muestra la ubicación del área
preplacodal a fines de la gastrulación y la ubicación, dentro ella, de
tres subáreas, una cefálica y una caudal, que originarán diferentes
grupos de placodas; entre ambas (línea de puntos) se halla una
subárea intermedia en la que aparecerá más tarde otra placoda.
La figura SC 11-2-1 B ilustra el proceso de refinamiento o
delimitación de subáreas, correspondientes a algunas placodas, que
acontece durante el transcurso del período somítico. La figura ilustra
un esquema que corresponde a la ubicación de las células que
originarán diferentes placodas en un embrión del estado de 10 a 13
pares de somitas (corresponde a un estado de aproximadamente 23-
24 días del embrión humano). Las poblaciones celulares precursoras
{kind=link}

de diferentes placodas ya se han segregado en diferentes dominios
del ectodermo de la región cefálica. En la región o subárea cefálica
se han separado las células de las placodas olfatoria y lental. En la
región o subárea caudal, las placodas ótica y epibranquiales también
se han separado y estas últimas se han escindido en las tres áreas
neuronogénicas correspondientes a los ganglios craneales
geniculado (VII par craneal), petroso (IX par craneal) y nodoso (X par
craneal). Recuérdese que estos ganglios quedan integrados por
neuronas sensoriales primarias provenientes de dos orígenes
diferentes: por un lado de las crestas neurales y por otro de las
placodas neuronogénicas epibranquiales. En este momento, en la
región o subárea intermedia, entre los dos grupos de placodas
mencionados (las derivadas de las subárea cefálica y caudal), se han
especificado las células que corresponden a la placoda trigeminal
con sus divisiones oftálmica, maxilar y mandibular.
En estados más avanzados (Fig. SC 11-2-1 C), que corresponden a
un esquema de fines del período somítico, las células que integran
los derivados de las placodas señaladas en la figura anterior ya han
tomado posiciones definitivas y se relacionan con las poblaciones
celulares con las que se desarrollarán interactivamente y han iniciado
las etapas iniciales de su diferenciación:
a) La placoda olfatoria ha iniciado su transformación en fosa olfatoria
y se ubica en posición adyacente al futuro bulbo olfatorio del
prosencéfalo.
b) La placoda lental se ha transformado en vesícula lental y se halla
ubicada en la concavidad de la copa óptica diencefálica.
c) La placoda ótica se ha invaginado y forma el otocisto que se halla
adosado a la pared lateral o alar del posencéfalo.
d) Las placodas epibranquiales, que se ubican en los extremos
dorsales de los surcos branquiales, han originado precursores que se
invaginan en el mesénquima y se asocian a células de la cresta
neural con las que forman los ganglios de los nervios craneales
mencionados más arriba.
e) En otras especies, caudalmente a la región branquial, se
desarrollan los receptores que integran la línea lateral.
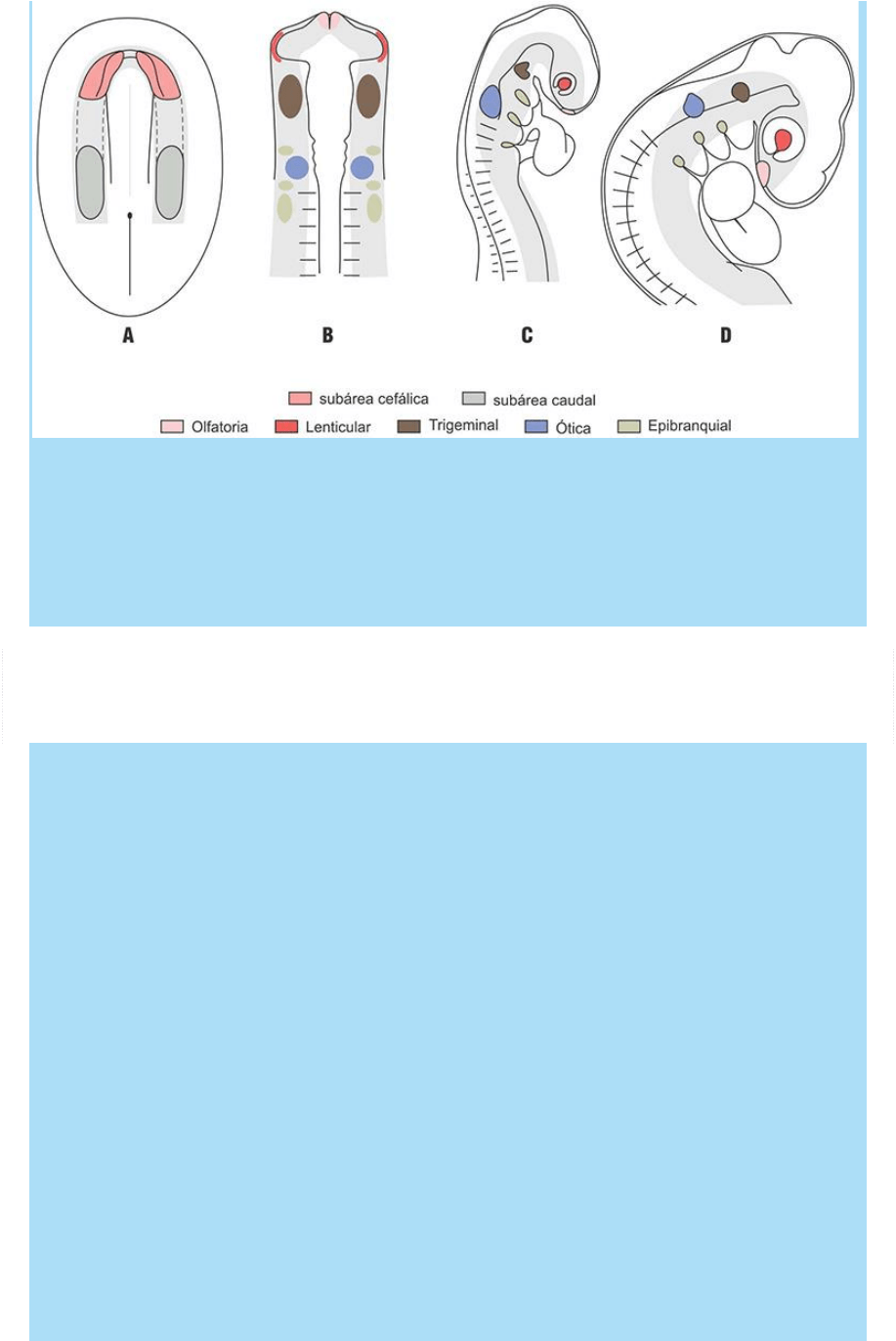
Fig. SC 11-2-1. Ubicación del área preplacodal, de sus subáreas
cefálica y caudal y de las placodas derivadas de cada una de ellas.
La secuencia de figuras A-D muestran: a) los cambios de posición
del área preplacodal y su transformación en la línea de Wolff y b) el
proceso de refinamiento (segregación de área placodales) que
acontece durante el período somítico. (Modelo: Embrión de pollo).
SC 11.3. La potencia citogenética del área preplacodal. Su
contribución a la citogénesis, histogénesis y organogénesis de
la región cefálica. V. Flores
El área preplacodal contribuye con más de diez categorías de células
a la histogénesis de diversas estructuras que integran el extremo
cefálico del embrión. Ello pone de relieve el papel central que ha
tenido en el proceso de cefalización (encefalización) que incluye,
entre otros aspectos, el desarrollo de a) el sistema sensorial
cefálico, b) el aparato de la contención neurosensorial y c) la
formación de estructuras que vinculan los sentidos con el sistema
nervioso central y la integración neuroendocrina necesaria para
elaborar respuestas integradas a los estímulos provenientes de un
ambiente permanentemente cambiante.
La figura SC 11-3-1 ilustra esquemáticamente las etapas tempranas
de la morfogénesis de diferentes tipos de placodas y la capacidad
citogenética de éstas. Algunas placodas precursoras de
neuroepitelios receptores y de células endocrinas sufren una
invaginación y luego se diferencian. Otras, además de invaginarse,
experimentan T e-m y aportan células migrantes al epitelio
mesenquimático. En estos casos, las células que quedan formando
parte del epitelio invaginado, en general, se diferencian en células
sensoriales (células sensoriales de laberinto membranoso) o
{kind=link}

neuronas sensoriales como las olfatorias; diferentemente, las células
que sufren T e-m, en algunos casos forman células mesenquimáticas
y en otros casos originan neuronas sensoriales primarias o células
gliales de ganglios craneales.
Con el objeto aportar mayor precisión a esta descripción,
analizaremos los diferentes tipos celulares generados a partir de
cada uno de los diferentes tipos de placodas.
1) Placodas sensoriales y neuronogénicas como las olfatorias y
óticas. Se invaginan totalmente (ótica) o parcialmente (olfatoria) y
originan neuroepitelios sensoriales y neuronas asociadas (células
sensoriales, sus células de sostén, neuronas sensoriales primarias y
células gliales de los ganglios asociados y nervios aferentes).
2) Placodas neuronogénicas. Poseen función específicamente
neuronogénica. Existen dos grupos: a) el grupo dorsal o trigeminal,
que aporta neuronas sensoriales primarias al ganglio del trigémino
(Gasser) en sus porciones oftálmica, maxilar y mandibular y b) el
grupo epibranquial, que origina las neuronas sensoriales primarias
de las porciones distales de los ganglios de los nervios craneales
(geniculado, nodoso, petroso).
3) Placoda endocrina o hipofisaria. Tiene una función de
integración neuroendocrina. Origina los diversos tipos de células
endocrinas de la adenohipófisis que se hallan reguladas por las
neuronas hipotalámicas y que, a su vez, regulan a las glándulas
endocrinas o los órganos blancos periféricos.
4) Placoda lental. Tiene como función generar células que se
diferencian, forman las fibras del cristalino y durante dicho proceso
mueren.
5) Placodas formadoras de mesénquima. En algunas especies
existen placodas cuya función específica es realizar T e-m y
amplificar la masa de mesénquima que forma el aparato de la
contención neurosensorial. En vertebrados superiores, con
excepción de la placoda lental, todas las demás tienen capacidad de
aportar mesénquima que contribuye a generar el tejido conectivo que
forma las cubiertas de los órganos de los sentidos o glándulas
mencionadas.
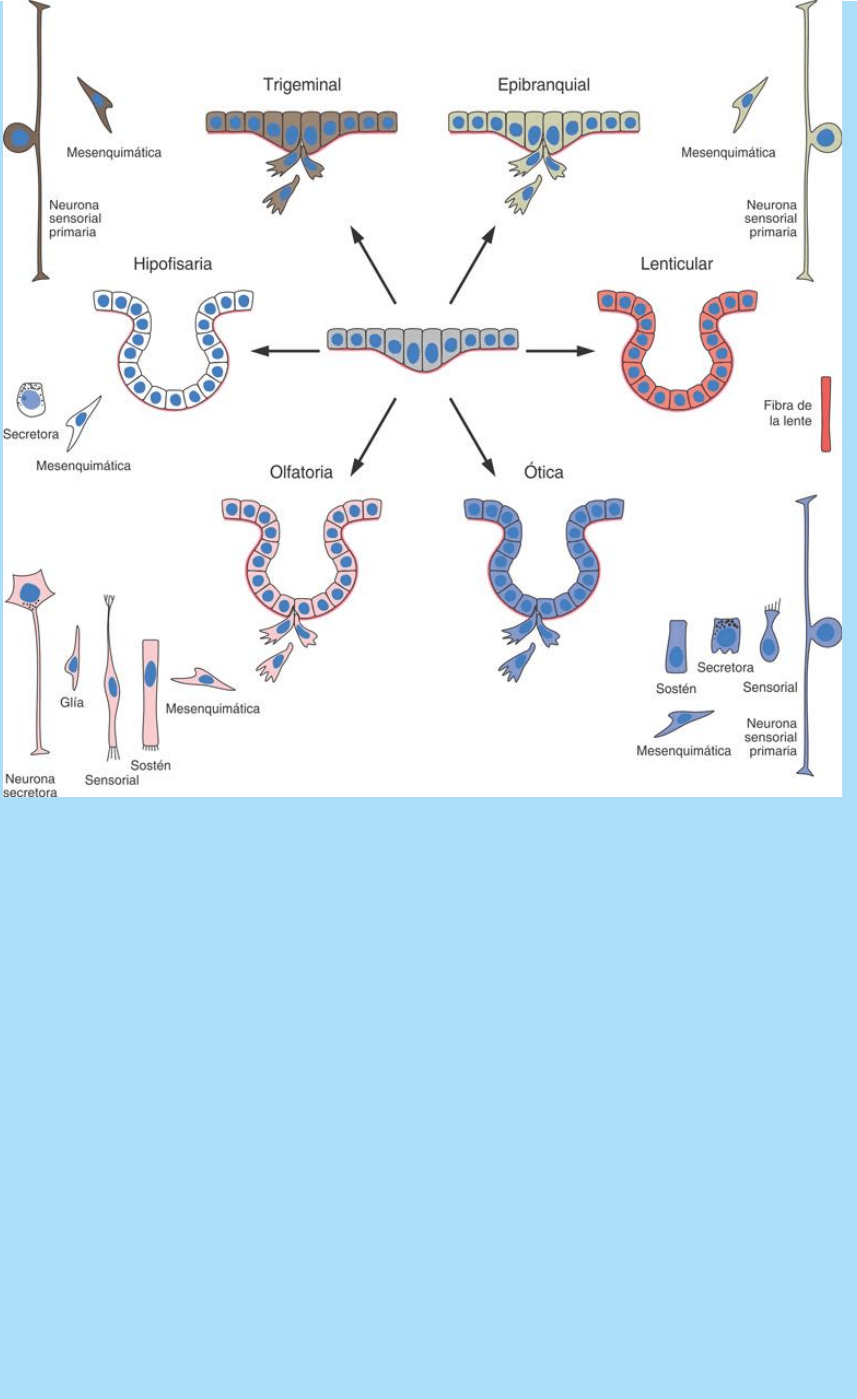
Fig. SC 11-3-1. El gráfico ilustra esquemáticamente las etapas
tempranas de la morfogénesis de diferentes tipos de placodas y su
amplia capacidad citogenética. Las placodas han sido ordenadas de
modo que la complejidad morfo-histogenética del proceso y la
potencia citogenética de las placodas aumentan de arriba abajo del
esquema. Algunas placodas de órganos sensoriales o glándulas
endocrinas sólo sufren una invaginación y luego se diferencian como
la placoda lental y la hipofisaria. Otras se invaginan pero algunas de
sus células sufren T e-m e ingresan al compartimento
mesenquimático. Las células que quedan formando parte del epitelio
invaginado en general se diferencian en células sensoriales (células
sensoriales de laberinto membranoso) o neuronas sensoriales como
las olfatorias. Las células que sufren T e-m, en algunos casos, forman
células mesenquimáticas y, en otros casos, originan neuronas
sensoriales primarias o células gliales de ganglios craneales.
La figura muestra distintos tipos de evolución que realizan las
diferentes regiones ectodérmicas de la zona preplacodal.
Algunas a) evolucionan sólo hasta el estado de placoda

(engrosamiento ectodérmico) y luego liberan células que migran y
forman células mesenquimáticas de la región branquial u otros tipos
celulares especializados tales como células endocrinas, neuronas,
células gliales, etc. Otras b), luego del estado de placoda, sufren una
invaginación, se convierten en una fosa que luego se suelta del
ectodermo superficial y se introduce en el mesénquima. Allí forman
una vesícula que luego se diferencia en un órgano definido.
Finalmente, c) otras tienen un comportamiento mezcla de los dos
anteriores: además de formar fosas y vesículas, liberan células que
adquieren diferentes vías evolutivas dependiendo de la placoda que
las originó. El diferente comportamiento morfogenético y citogenético
de las distintas placodas se debe a que las poblaciones celulares que
las constituyen ya están determinadas hacia una evolución definida.
En general, la determinación de las células que ocupan las diferentes
regiones precede a las manifestaciones estructurales mencionadas
(formación de placoda, invaginación, formación de vesícula, etc.).
El fenómeno de formación del área preplacodal, sus subáreas y las
placodas que dentro de ellas se desarrollan se encuentra dentro del
marco general de determinación progresiva que experimenta el
ectodermo de la región cefálica del embrión, asociado a la
cefalización. Si bien existen particularidades para cada una de las
placodas, en general, tempranamente, en un área definida pueden
coexistir células que luego forman parte de diferentes placodas y
diferentes órganos definitivos. La expresión de combinatorias
específicas de factores de transcripción como por ejemplo los
factores Dlx3, Dlx5, Pax6 y la proteína marcador panneuronal Hu,
que es considerado un marcador temprano de determinación en
sentido neuronal, han sido utilizados para analizar cómo
gradualmente se determina la potencia citogenética de diversas
placodas en función del tiempo. El estudio de la expresión de estos
marcadores aplicados a situaciones experimentales de trasplante de
las áreas preplacodales a distintas regiones del embrión permite ver
que tempranamente las áreas poseen una potencia amplia y que
gradualmente se va restringiendo a un destino cada vez más
estrecho y mejor definido.
El proceso de determinación progresiva en el ectodermo de la
región cefálica implica las siguientes etapas:
a) Una determinación y restricción de destinos correspondientes a
ectodermo neural y ectodermo no-neural.
b) En la zona de transición entre ambas se determina a continuación
la zona correspondiente a la cresta neural.
c) A continuación, en la zona lateral del ectodermo no neural, se
genera el área preplacodal. Ésta posee dos subáreas celulares, una
cefálica y otra caudal, de potencias superpuestas correspondientes
a conjuntos de placodas.
d) A continuación, en las áreas cefálica y caudal se determinan y
delimitan topográficamente cada una de las placodas que de ellas
derivan. Además, en la zona comprendida entre las dos subáreas
mencionadas, se constituyen nuevas placodas.
e) El resto del ectodermo no nerual de dicha región, que no sufrió las
determinaciones mencionadas, queda como el ectodermo
epidérmico que originará la dermis y las glándulas anexas de la piel
de dicha región.
SC 11.4. Modelo de inducción de la región preplacodal. V.
Flores
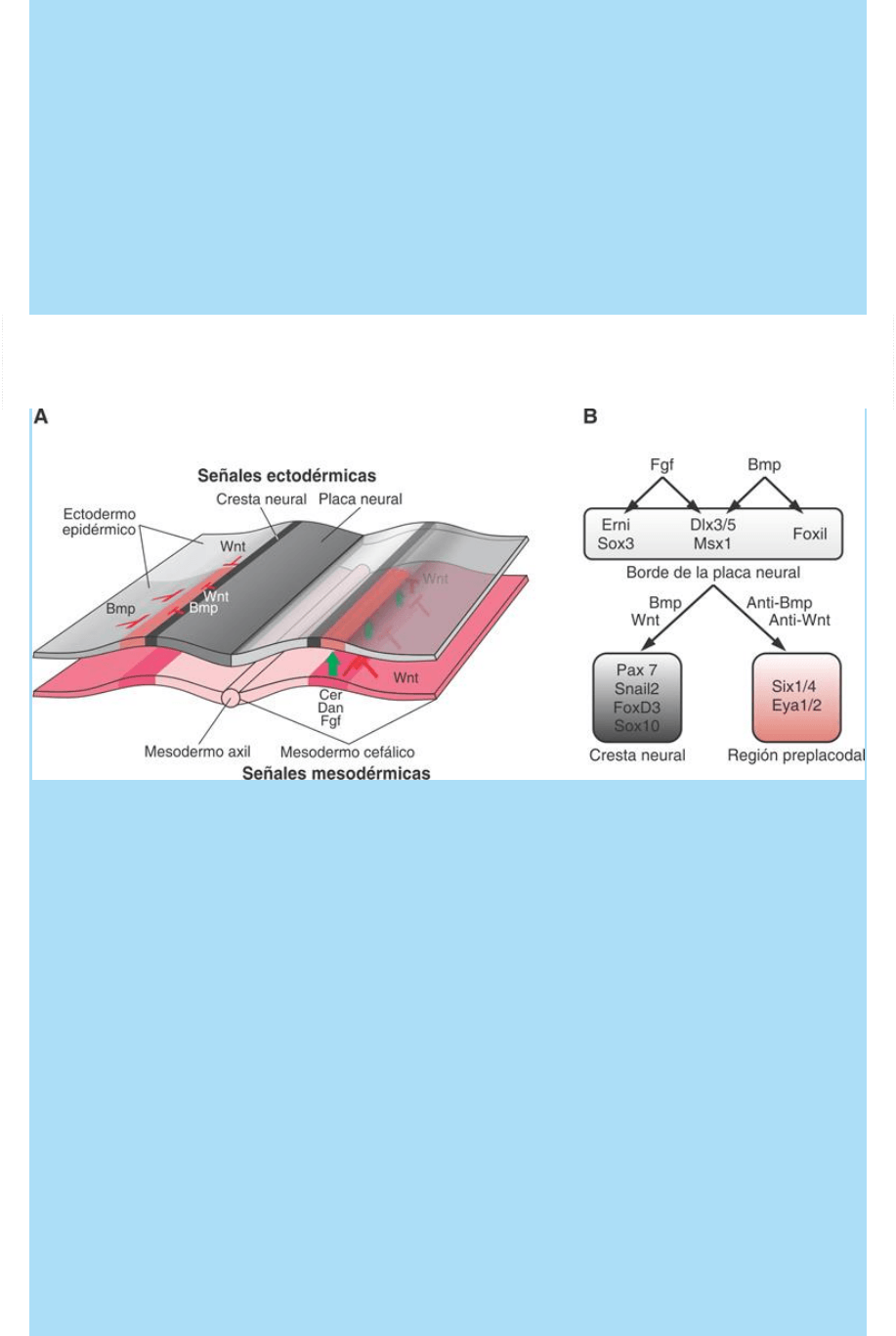
Fig. SC 11-4-1. A. Representación esquemática de una sección
transversal de la región preplacodal del embrión (Estado de 2-4
somitas). B. Panel superior: señales y factores de transcripción que
operan en el ectodermo del borde de la placa neural. Panel inferior:
reprogramación de la expresión y direccionamiento en sentido de
cresta neural y de región preplacodal.
Varios estudios realizados en el embrión de pollo sobre la distribución
de las áreas de expresión de diferentes proteínas señal, sus
receptores, proteínas de señalización intracelular y de factores de
transcripción han llevado a proponer un modelo provisorio acerca de
cómo se determina y localiza el área generadora de placodas (área
preplacodal) en el ectodermo del extremo cefálico del embrión.
El modelo propone que el área preplacodal se determina entre el final
de la gastrulación y el período somítico temprano (Estados de 2-4
somitas; Embrión de pollo). Según el modelo, un conjunto de señales
positivas (estimulantes del desarrollo) y negativas (inhibitorias del
desarrollo) contribuyen a determinar el área preplacodal y a
{kind=link}

localizarlo entre el territorio correspondiente a la crestas neural
craneal y el resto del ectodermo epidérmico.
En el lado derecho de la figura SC 11-4-1A se muestra que las
proteínas señal Cer, Dan, Fgf y otras, generadas en el mesodermo
subyacente en el área preplacodal participan positivamente en la
determinación y el desarrollo del área preplacodal. Por el contrario,
el mesodermo de las regiones lateral y caudal generan la proteína
señal Wnt que inhibe la evolución en sentido preplacodal. En el lado
izquierdo de la figura SC 11-4-1A se ilustra que el ectodermo de las
regiones lateral y caudal y los pliegues neurales también generan
señales inhibitorias (Wnt y Bmp) que contribuyen a limitar y localizar
la extensión del área preplacodal. Por el contrario, el factor Fgf y otras
proteínas inhibitorias de Wnt y BMP protegen al área preplacodal de
la acción inhibitoria de estas últimas y permiten la formación de
poblaciones celulares precursoras de placodas.
En el esquema B de la figura SC 11-4-1 se ilustra parcialmente el
proceso de determinación progresiva y refinamiento de la región. En
el panel superior se indica que tanto las señales Fgf como Bmp
actúan conjuntamente promoviendo la expresión de una
combinatoria de factores de transcripción que caracteriza a la región
borde de la placa neural. A partir de esta combinatoria de factores de
transcripción, las células pueden experimentar reprogramaciones
que las guían a determinarse en sentido de cresta neural o, por el
contrario, a determinarse en área preplacodal. Como se ilustra en el
panel inferior, esta dicotomía depende de si las células se hallan en
una zona con niveles locales altos de actividad de las señales Bmp y
Wnt o, por el contrario, en una zona con niveles locales altos de
actividad de sus inhibidores (proteínas anti-Bmp y anti-Wnt). De las
relaciones entre estas actividades contrapuestas depende que las
células de la zona de transición placa neural-ectodermo epidérmico
se determinen en zona precursora de placa neural o área
preplacodal.
SC 11.5. Papel del posencéfalo y tejidos adyacentes en el
patterning del otocisto. V. Flores
La combinación de métodos experimentales dirigidos a la elaboración
de mapas de destino y al análisis de la organización espacial de la
expresión de factores de transcripción ha permitido acumular
información suficiente para proponer un modelo del modo como se
organiza el patterning del otocisto, vale decir, cómo se organiza en el
espacio el proceso de determinación de las diferentes regiones que
lo integran. Las figuras SC 11-5-1 a SC 11-5-3 ilustran modelos
{kind=link}
{kind=link}
sobre la posible influencia del posencéfalo y otros tejidos adyacentes,
en la determinación de compartimentos con diferente potencia
evolutiva en el campo ótico. Este proceso de determinación se
produciría en forma progresiva.
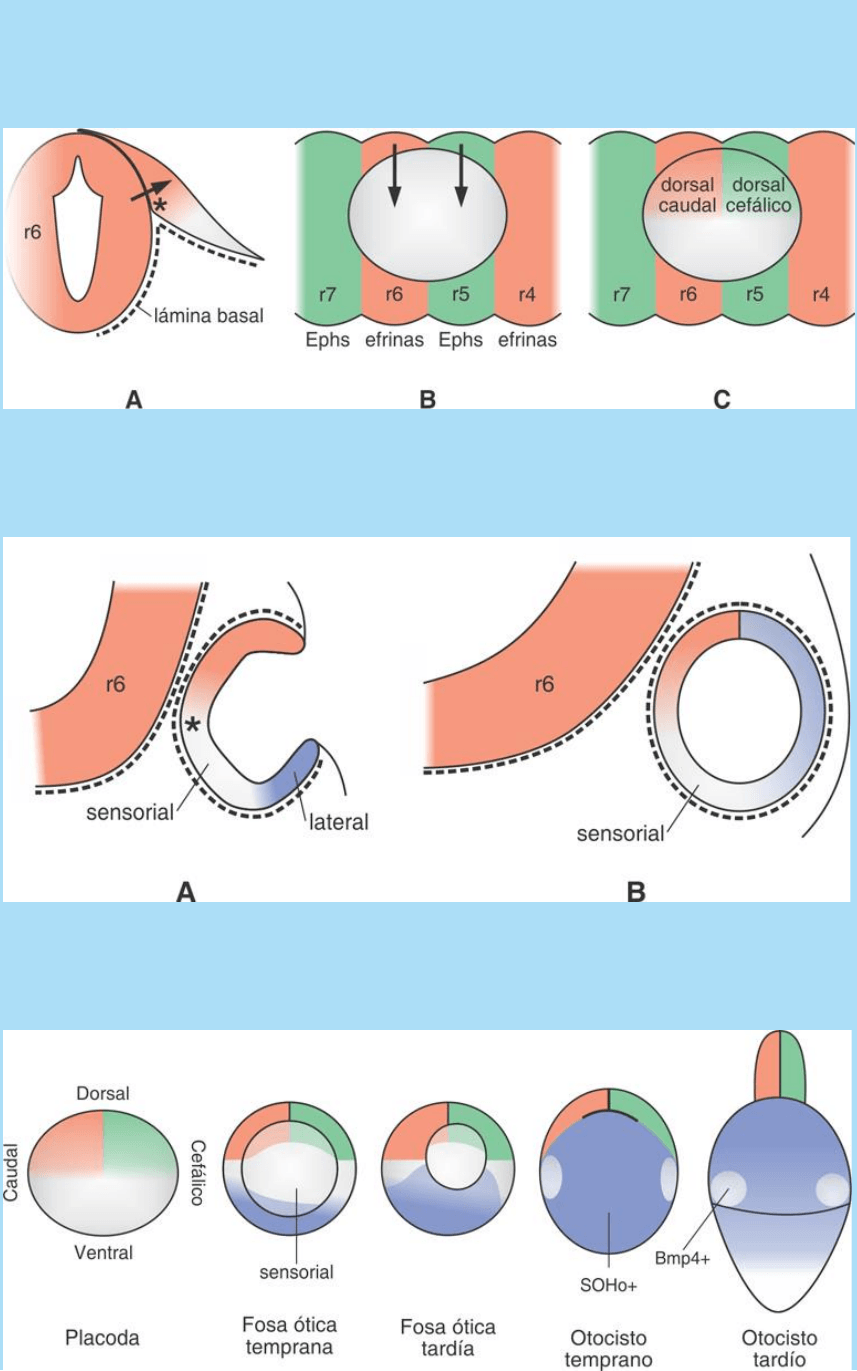
Fig. SC 11-5-1. A-C. Ilustra la fase de contacto entre placoda ótica y
posencéfalo y la determinación de los compartimentos dorsal, dorsal-
cefálico y dorsal-caudal en relación con las rombómeras 5 y 6
(flechas).
Fig. SC 11-5-2. A-B. Ilustra la determinación de los compartimentos
medial y lateral durante la invaginación de la placoda ótica y
formación del otocisto. La zona ventral aún indeterminada será la
futura región sensorial.
{kind=link}

Fig. SC 11-5-3. La serie de esquemas ilustra el proceso de cierre de
la fosa ótica, en el polo dorsal del otocisto, y la redistribución espacial
de sus compartimentos diferentemente determinados. Éstos
expresan diferentes patrones de expresión de señales y factores de
transcripción.
Establecimiento de los compartimentos cefálico-medial y caudal-
medial. La figura SC 11-5-1 A muestra que las superficies basales de
la porción medial-dorsal de la placoda y del posencéfalo se hallan en
íntimo contacto pues carecen de una membrana basal bien
desarrollada (línea de puntos). Se considera que este hecho permite
o facilita las interacciones entre ambas poblaciones celulares.
Durante esta etapa temprana se imprimiría en la placoda, por la
acción de señales provenientes del posencéfalo (flechas), el carácter
medial o propiedades de lugar mediales. Durante dicho contacto se
instalaría también un borde de determinación diferente entre las
propiedades cefálica y caudal en la zona medial de la placoda. En
las figuras SC 11-5-1 B y C se ilustra que los límites entre sucesivas
rombómeras están instalados por la expresión regional alternante y
nítidamente delimitada de las proteínas receptores Ephs y sus
proteínas ligandos ephrinas. Obsérvese que la placoda ótica está en
contacto directo con las rombómeras 5 y 6 y el límite entre ambas
coincide con el límite entre las porciones cefálica y caudal de la
placoda de la región dorsal de la placoda. Debido a ello se propone
que este segundo proceso de compartimentalización de potencias
podría deberse a señales determinantes (flechas) provenientes de
las rombómeras 5 y 6 (flechas).
Establecimiento del compartimento lateral y las zonas con
competencia sensorial. La figura SC 11-5-2 muestra que durante la
invaginación de la placoda ótica se genera una región longitudinal
que opera como bisagra de giro entre sus mitades dorsal y ventral.
Durante dicho proceso, la mitad inferior del campo ótico se
aproximaría a tejidos ventrales generadores de otras señales
determinantes (SC El patterning del otocisto. Organización espacial
de las regiones precursoras sensoriales en la vesícula ótica). Recién
entonces, como resultado de la acción de dichas señales, la porción
lateral del campo ótico adquiriría la identidad correspondiente al
compartimento lateral. Esto ocurriría mientras la placoda se invagina
y adquiere forma de copa. La zona profunda de la copa, ubicada entre
los compartimentos medial y lateral, se determinaría luego en sentido
sensorial. Si bien se desconoce con exactitud la naturaleza y el origen
de estas señales, se sabe que existen conjuntos se señales de
diferente origen que se expresan durante estos procesos y

promueven la expresión de diferentes combinatorias de factores de
transcripción en distintas regiones del otocisto en formación (SC
11.6. Señales difusibles y factores de transcripción involucrados en
la determinación de la placoda ótica).
Cierre de la fosa ótica y redistribución de compartimentos. Durante el
cierre de la fosa ótica, las regiones con diferente identidad arriba
mencionadas (compartimento o cuadrantes cefálico-medial y caudal-
medial, la mitad ventral y la zona sensorial) sufren cambios de
posición relativa y terminan ordenándose como ilustra la figura SC
11-5-3. En la zona de cierre, que pasa a ocupar una posición dorsal,
confluyen y entran en contacto los tres primeros compartimentos
señalados. Las regiones sensoriales pasan a ocupar entonces las
regiones ecuatorial y ventral del otocisto. Es probable que estas
últimas sean las que generan las células que se liberan del otocisto
y pasan a formar parte de las neuronas sensoriales primarias de los
ganglios acústico y vestibular.
SC 11.6. Señales difusibles y factores de transcripción
involucrados en la determinación de la placoda ótica. V. Flores
La inducción y ulterior desarrollo de la placoda ótica a partir del
ectodermo preplacodal depende de una sucesión de procesos de
señalización mediados por señales generadas en varios tejidos
adyacentes al campo ótico.
La inducción y localización temprana del campo ótico dentro del área
preplacodal depende, al menos en parte, de la secreción de la
proteína señal Fgf3 a partir del posencéfalo (SC La extensión del
campo ótico y la determinación y localización de la placoda ótica.
Papel del factor de crecimiento fibroblástico 3). En relación con esta
señalización, las células del campo ótico, a su vez, se caracterizan
por la expresión de una combinatoria típica de factores de
transcripción de los tipos Dlx y Fox (Dlx3, Dlx4, Sox9, Foxi1).
El Fgf3 no es la única señal de esta familia que se secreta en la región
y que influye en el patterning del otocisto; por ejemplo, el
mesénquima periótico, ubicado entre el posencéfalo y el área
preplacodal también, cumple un papel importante en esta
señalización ya que secreta las proteínas Fgf 10, 15 y 19.
Se considera que la expresión combinada de todos los Fgf (3, 10,15
y 19) secretados por el posencéfalo y el mesénquima preótico
constituye un código de señales Fgf que estimula el desarrollo de
la placoda ótica y, además, contribuye a generar una expresión
diferencial espacialmente organizada de factores de transcripción
que establece varios compartimentos diferentemente determinados
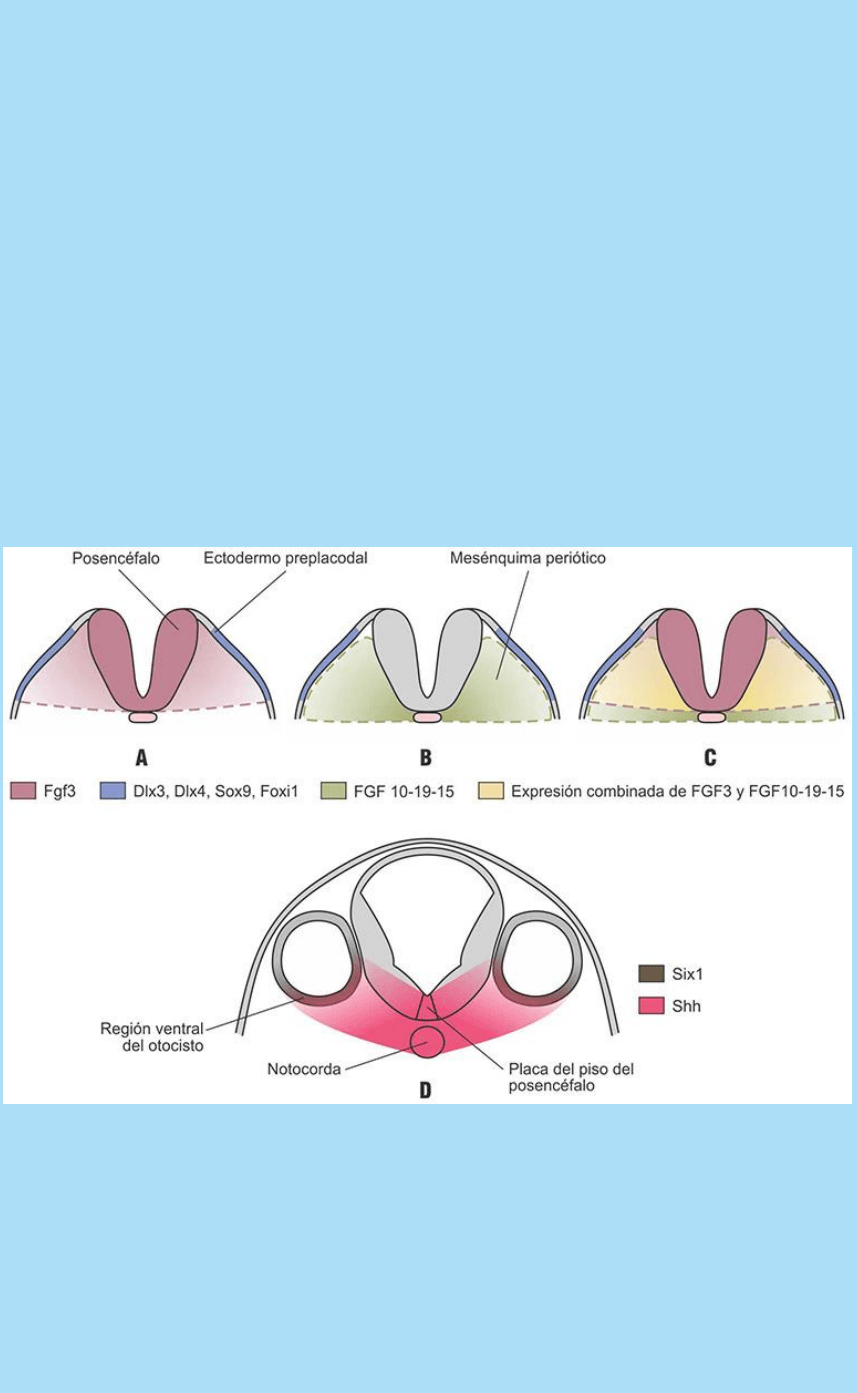
(SC Expresión combinatoria de factores de transcripción y
especificación de compartimentos en el otocisto).
A continuación se agregan otros fenómenos de señalización que
agregan mayor especificidad regional al otocisto. Por ejemplo, la
secreción de la proteína señal Shh, por parte de la notocorda y la
placa del piso del tubo neural, tiene un rango de alcance que llega
hasta el otocisto en formación. En relación con esta estimulación, que
alcanza a la región ventral del otocisto, esta región inicia la expresión
de ciertos factores como, por ejemplo, el factor de transcripción
Six1 que, al parecer, participa en determinar la identidad de dicha
región o compartimento.
Existen resultados experimentales que muestran que una variedad
de otros factores de transcripción se expresan en regiones definidas
del otocisto y contribuyen a especificar combinatoriamente los
diferentes compartimientos del otocisto (SC Expresión combinatoria
de factores de transcripción y especificación de compartimentos en
el otocisto).
Fig. SC 11-6-1. Modelo del patterning del otocisto mediado por una
combinatoria de proteínas señal secretadas por distintas poblaciones
organizadoras cercanas al campo ótico y de expresión diferencial de
factores de transcripción. La organización espacial de las señales
genera una organización espacial de diferentes compartimentos en
el otocisto. Código: A. Posencéfalo: FGF3, ectodermo preplacodal:
expresa Dlx3, Dlx4, Sox9, Foxi1. B. El mesénquima periódico:
produce FGF 10-19-15. C. La expresión combinada de FGF3 y
FGF10-19-15 genera un código que induce el desarrollo placodal en
{kind=link}

una región específica del ectodermo. D. La región ventral se
especifica por la expresión combinada de Six1 en el otocisto y por la
expresión a distancia de Shh proveniente de la placa del piso y
notocorda.
SC 11.7. Un modelo de compartimentos y fronteras (bordes)
para explicar el patterning y la morfogénesis del oído interno.
V. Flores, M. Rapacioli
El proceso de morfogénesis que sufre el epitelio del otocisto, que lo
lleva a transformarse en el revestimiento epitelial del laberinto
membranoso, está entre los más difíciles de explicar. Basta
considerar la a) complejidad estructural del laberinto membranoso, b)
la diversidad de tejidos y células que lo componen, c) la disposición
espacial de cada una de ellas y d) la armonía estructural requerida
para una correcta integración estructura-función para notar cuántos
y cuán complejos deberían ser los cambios a fin de que una pequeña
esfera epitelial origine todos esos componentes adecuadamente
ensamblados.
Los CCD que cumplen las células del otocisto se hallan
temporoespacialmente organizados por medio de la acción de varias
poblaciones celulares señalizadoras localizadas en las adyacencias
del otocisto y que imprimen polaridades (ejes) y determinan
compartimentos en la estructura del otocisto (SC 11.5. Papel del
postencéfalo y tejidos adyacentes en el patterning del otocisto; SC
11.6. Señales difusibles y factores de transcripción involucrados en
la determinación de la placoda ótica). Estas polaridades derivan de
las propias polaridades céfalo-caudal, medio-lateral y dorso-ventral
global del embrión.
Las polaridades mencionadas se traducen, a continuación, en las
diversas combinatorias de factores de transcripción que definen
diversos dominios o compartimentos del otocisto (SC Expresión
combinatoria de factores de transcripción y especificación de
compartimentos en el otocisto).
Con el objeto de analizar el patterning del otocisto, desde el punto de
vista teórico, se pueden considerar, como referencia espacial, los tres
planos perpendiculares a cada uno de los ejes o polaridades arriba
especificados: cf-cd, d-v y md-lt. Estos tres planos son
perpendiculares entre sí y cada uno de ellos divide al otocisto en dos
hemisferios. Vale decir, los planos son concebidos como fronteras
entre hemisferios homónimos a los ejes mencionados. Así, un plano
perpendicular ‒el eje céfalo-caudal (o anteroposterior)‒ corresponde
a la frontera entre los hemisferios cefálico (anterior) y caudal
(posterior). Otros dos planos, perpendiculares a los ejes md-lt y d-v
Este documento contiene más páginas...
Descargar Completo
Cap 16 SC Flores.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.