
SC 10.1. La influencia de la asimetría sobre la organización
espacial de los CCD: el eje superficie-profundidad de la
vesícula lental. V. Flores, M. Rapacioli
El desarrollo de la vesícula lental ofrece un ejemplo acerca de cómo
las características fisicoquímicas del ambiente de las células en
desarrollo influyen sobre sus CCD. Las células de la vesícula lental
exhiben desde temprano una diferencia en sus CCD según ocupen
la porción superficial o profunda de ella. Las del hemisferio superficial
se mantienen durante mucho tiempo con capacidad proliferativa y sin
capacidad de diferenciarse, en tanto que las células del hemisferio
profundo cesan rápidamente la proliferación y se diferencian en fibras
del cristalino. En otros términos, la lente exhibe un gradiente de
desarrollo tal que se encuentra más avanzado en el polo profundo.
Dicho cambio de comportamiento se produce en la zona del ecuador
entre ambos casquetes (superficial y profundo). Como las células de
la superficie anterior proliferan, se van desplazando hacia el ecuador,
las que llegan al ecuador cambian bruscamente de comportamiento:
dejan de comportarse como pertenecientes a la población superficial
y adquieren la conducta típica de las de la región profunda.
Es como si la superficie anterior sólo proveyera las células que se
diferencian sólo en la región profunda. Las células que llegan al polo
profundo se diferencian, el citoesqueleto va adquiriendo una
estructura cristalina integrada por un conjunto típico de proteínas
específicas de la lente (cristalinas), pierde vitalidad el núcleo y los
organotes se disgregan y se transforman en estructuras con
propiedades de fibras ópticas. En sentido estricto, las fibras del
cristalino no son células sino el resto que queda de las células una
vez que mueren.
El epitelio anterior de la vesícula lental sigue proliferando y
produciendo nuevas células por mucho más tiempo. Como resultado
de la intercalación de las nuevas células, el epitelio anterior se
expande y las células más viejas van corriéndose hacia el borde o
ecuador del cristalino. Una vez allí inician la diferenciación. Así, a lo
largo del desarrollo se van agregando nuevas cohortes de células a
lo largo de toda la banda ecuatorial como nuevas capas concéntricas
de nuevas fibras ópticas. ¿Están determinadas a comportarse
diferentemente las células lentales de las regiones superficial y
profunda? Algunas experiencias de rotación de la lente durante el
desarrollo indican que no.
La microcirugía permite extraer la lente y reubicarla en una posición
diferente. Si durante el desarrollo se extrae la lente, que ya ha

iniciado su desarrollo, y exhibe claras diferencias entre células
superficiales (no diferenciadas y proliferantes) y fibras profundas (ya
diferenciadas) y se la rota 180º de modo que el hemisferio profundo
pase a ser superficial y viceversa, ocurre el siguiente
fenómeno: a) las células que se encontraban en la cara superficial
pasan a estar ubicadas en el interior de la copa óptica; allí cesan su
proliferación y rápidamente se diferencian; b) las células que se
encontraban en el hemisferio profundo y que ya se habían convertido
en fibras del cristalino (elementos inertes sin vitalidad) quedan como
están, pero ahora en la superficie anterior; c) un fenómeno
significativo ocurre en la población de células que ocupan el ecuador
de la lente en desarrollo.
Las células de la banda del ecuador siguen en el ecuador pero con
una polaridad espacial superficie profundidad invertida. Las células
que estaban pasando de la cara anterior a la posterior estaban
cesando su proliferación e iniciando la diferenciación. Al producirse
la inversión del eje superficieprofundidad de la lente, las células que
estaban de la región superficial del ecuador, que aún proliferaban,
pasan a iniciar la diferenciación, y las células que estaban en la
región profunda del borde, que estaban iniciando la diferenciación,
reasumen la proliferación. Así se genera una nueva población de
células lentales en desarrollo con una polaridad invertida con
respecto a las que habían iniciado el desarrollo de la lente. Las
nuevas células profundas inician la formación de un nuevo núcleo de
fibras primarias a las cuales se agregan luego nuevas fibras
secundarias. Las nuevas células superficiales regeneran un nuevo
epitelio anterior que se extiende sobre la superficie de la exporción
profunda ya diferenciada. Estas dos nuevas poblaciones celulares
continúan el desarrollo de una lente que se construye sobre la que
se había formado inicialmente y que consiste en una lente mixta con
un doble carácter, ya que las fibras ópticas formadas antes de la
inversión tienen una orientación espacial y las que se formaron luego
de la inversión tienen la orientación opuesta.
Este resultado experimental ilustra con claridad la importancia de la
organización espacial de los CCD y la dependencia de dicha
organización respecto de las vías de señalización espacialmente
estructuradas que “imprimen” una asimetría en el entorno de las
células en desarrollo (Fig. SC 10-1-1).
{kind=link}
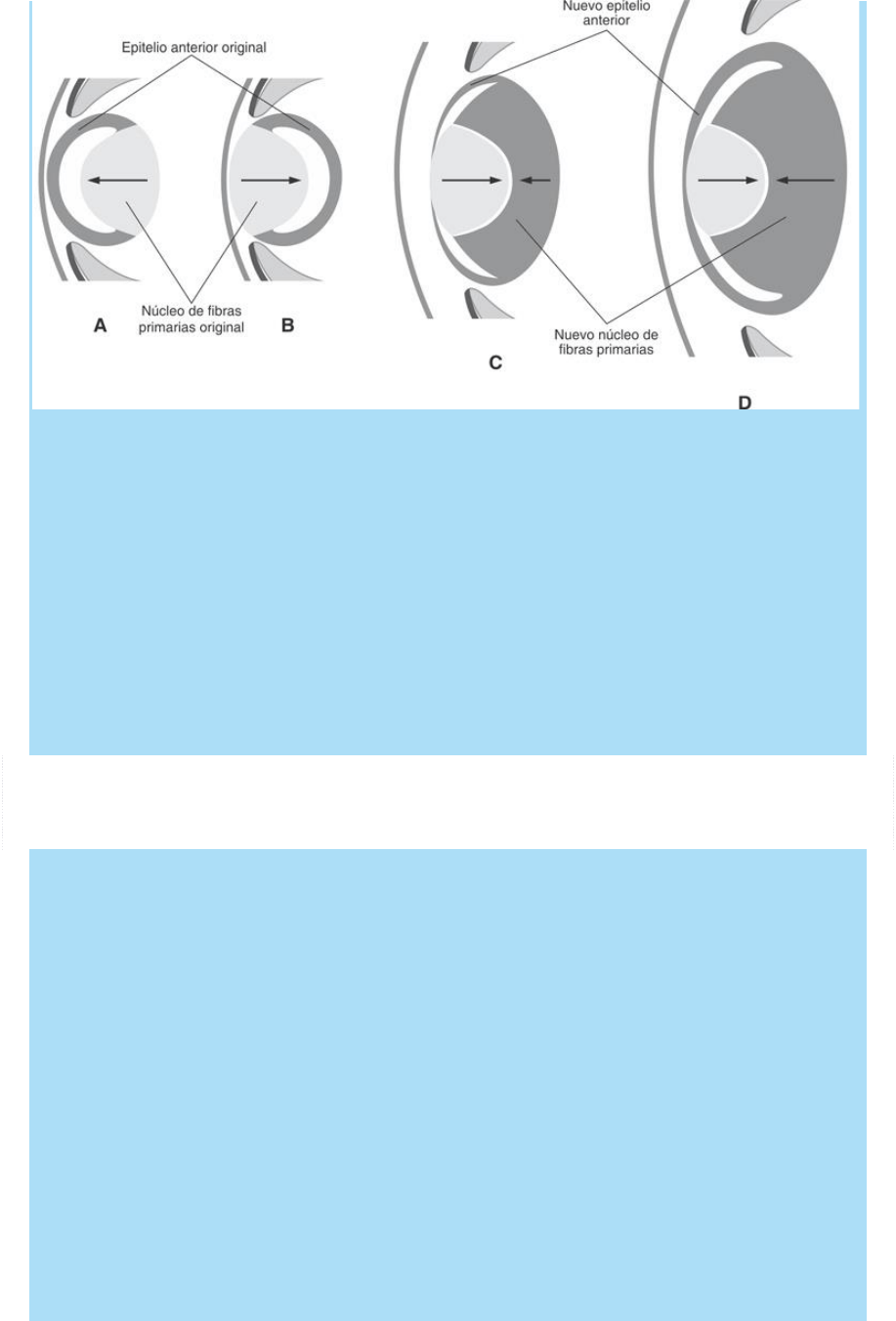
Fig. SC 10-1-1. Representación esquemática de la experiencia de
rotación de la lente. A. Estado inicial: lente en su posición
normal. B. Estado inmediato posterior a la cirugía: lente rotada
180º. C. Seis días después de la cirugía: las células que inicialmente
formaban el epitelio anterior (proliferativo) se diferencian en “nuevas
células profundas” e inician la formación de un “nuevo” núcleo de
fibras primarias y las “nuevas células superficiales del ecuador”
reasumen la proliferación y regeneran un nuevo epitelio
anterior. D. Diez días después de la cirugía.
SC 10.2. El campo lental del extremo cefálico del embrión.
Interacciones celulares y vías de señalización involucradas en
su constitución y evolución. V. Flores
El ectodermo epidérmico de las regiones laterales del prosencéfalo,
la zona que cubre las vesículas ópticas adquiere, desde muy
temprano, potencia para formar la lente. Dicha región ectodérmica
integra la zona denominada línea de Wolff que origina placodas
ectodérmicas. Las placodas ectodérmicas generan células que
luego se diferencian en a) neuroepitelios receptores periféricos
(olfatorio, auditivo, del equilibrio, etc.), b) células que se invaginan e
incorporan a ganglios sensoriales craneales (placodas óticas,
placodas epibranquiales) y, además, generan células
que c) contribuyen a amplificar las poblaciones mesenquimáticas
cefálica y branquial.
El área de ectodermo que durante el período somítico forma la línea
de Wolff deriva de la zona ectodérmica, ya definida durante la
gastrulación, denominada área preplacodal. Esta área, en estados
más tempranos, ocupa la región del ectodermo epidérmico que

bordea, a modo de herradura, el extremo cefálico de la placa neural
(SC 11.4. Modelo de inducción de la región preplacodal).
La zona llamada línea de Wolff está integrada por el ectodermo
epidérmico y una delgada capa de mesénquima subyacente con la
que interactúa. Este mesénquima está formado mayoritariamente por
células provenientes de la cresta neural preótica o craneal. En la
región encefálica, las células de la cresta neural se segregan del
compartimento epitelial incluso antes de que se cierre el tubo neural.
Parte de estas células se distribuyen en superficie por debajo del
ectodermo y junto con él forman parte de la línea de Wolff.
Algunos experimentos clásicos muestran que, cuando dicha región
ectodérmica es trasplantada muy tempranamente (antes de la
interacción con su correspondiente mesénquima), no se transforma
en placodas sino simplemente en epidermis de la región a la que
fueron trasplantadas. Por el contrario, cuando se explantan o
trasplantan, en estados más avanzados, porciones definidas de la
línea de Wolff, estos tejidos originan las placodas que hubieran
formado en sus lugares de origen.
Este segundo resultado implica que se hallaban determinadas antes
del trasplante. Se sabe que para que la determinación ocurra el
ectodermo del área preplacodal debe recibir previamente diferentes
tipos de estímulos de tejidos mesodérmicos y ectodérmicos
adyacentes. Entre estas señales existen algunas positivas (+) que
son determinantes y existen algunas que son inhibitorias o negativas
(-). Las primeras posibilitan que las células expresen la combinatoria
de factores de transcripción correspondiente al área placodal. Las
segundas sirven para localizar la acción de modo que el área no se
extienda más de lo normal.
En el caso concreto de la placoda lental, se considera que uno de los
factores transcripción incluidos en la combinatoria de factores que
participa en su determinación es el factor Pax6. Casi
simultáneamente, las células de la región ocular de la placa neural,
región precursora de la vesícula óptica, expresan Pax6. No se sabe
si la expresión de este factor indica determinación en sentido lental o
adquisición de competencia para responder a la acción determinante
del mesénquima cefálico.
De todos modos, se sabe que si el ectodermo de dicha región es
trasplantado a la región de la placoda auditiva, el ectodermo se
diferencia en placoda (aun sin interacción con la vesícula óptica)
reteniendo su carácter lental. Este hecho muestra que la región ya
está determinada antes de su interacción con la vesícula óptica.

Estos experimentos también indican que la población celular
determinante que restringe la potencia del ectodermo epidérmico y lo
determina en sentido lental es el mesénquima cefálico regional.
También existen experimentos que indican que la región posee
propiedades de campo, vale decir, antes de la constitución de la
placoda lental ya está determinada pero con propiedades regulativas.
Acciones permisivas y diferenciación de la placoda lental. Una
vez determinada la evolución en sentido lental, la etapa siguiente es
la diferenciación en placoda lental. A este fenómeno le siguen la
invaginación de la placoda, la formación de la vesícula lental y la
diferenciación de ésta en lente. En este proceso participa, al parecer
permisivamente, el neuroepitelio de la vesícula óptica. El carácter
permisivo de tal acción es supuesto a partir del hecho que: a) no
asigna carácter lental pues ya lo posee desde antes y b) en
condiciones experimentales, el efecto del neuroepitelio de la vesícula
óptica puede ser reemplazado exitosamente por señales generadas
por una diversidad de otros tejidos. Se considera que durante la
diferenciación las células del campo lental reciben señales de la
vesícula óptica ya que ésta secreta varias proteínas señal (p.
ej., proteína señal Bmp4) que estimulan la expresión de
nuevos factores de transcripción como Sox2, Pax, la proteína
factor de transcripción KLF6 (Krüppel-like factor-6) y otros.
La formación de la fosa y la vesícula lental. Los procesos que
conducen a la formación de la fosa y vesícula lental, si bien pueden
producirse en otros lugares en los que la placoda es trasplantada, no
se cumplen de la misma forma. Ello se debe a que la precisión de
dichos procesos morfogenéticos está regulada por medio de
interacciones de adhesión, fuerzas interfaciales de intensidad
regulada que sólo se logran a través de interacciones con la vesícula
óptica y el ectodermo regional (SC 10.5. El papel morfogenético del
contacto placoda lental-vesícula óptica).
SC 10.3. El papel del mesénquima cefálico periocular como
integrador de la morfogénesis y la histogénesis ocular. V.
Flores
El mesénquima cefálico es una adquisición evolutiva relativamente
reciente. Se ha desarrollado a lo largo del proceso de cefalización
que ocurrió desde la aparición de los cefalocordados en adelante.
Cumple un papel primordial en la morfogénesis y la histogénesis del
aparato de la contención neurosensorial y la de los propios órganos
de los sentidos.

En el caso del desarrollo del ojo, que posee varias cubiertas
conectivas con importantes diferencias interespecíficas, el
mesénquima cefálico ha sufrido varias adaptaciones vinculadas a los
cambios evolutivos del sistema visual. Varias son las instancias en
las que células del mesénquima cefálico desempeñan papeles
importantes durante el desarrollo del sistema visual.
a) El efecto más temprano conocido es la escisión del campo ocular
(retiniano) del extremo cefálico (prosencefálico) de la placa neural en
dos porciones, derecha e izquierda, con capacidad para formar un
par de ojos con simetría bilateral. Tempranamente, dicha región tiene
capacidad de campo y forma un único ojo medial. Ulteriormente es
escindido en dos mitades simétricas y bilaterales. Esta función al
parecer es cumplida por células del mesodermo axil precordal
(proceso cefálico o notocorda anterior) que, a través de la
señalización celular vía Shh, que es un importante morfógeno de la
región ventral del tubo neural, tienen un efecto en la generación de la
línea media y la formación de estructuras especulares o bilaterales
respecto de ella. Las alteraciones en este proceso de señalización
conducen a malformaciones en las que no es posible distinguir una
línea media como la ciclopía aislada o asociada a otras
malformaciones más graves como la holoprosencefalia. Por otro
lado, se ha postulado que este mesénquima también contribuye con
células musculares a la musculatura extrínseca del ojo.
b) El siguiente efecto es su participación, como generador de señales
determinantes y localizadoras del área preplacodal y luego, durante
la formación de la línea de Wolff, como generadora de señales
determinantes y localizadoras del campo lental en el ectodermo del
área preplacodal (SC 10.2. El campo lental del extremo cefálico del
embrión. Interacciones celulares y vías de señalización involucradas
en su constitución y evolución). Este efecto, denominado
“localizador” de la capacidad formadora de lente y diferenciación de
la placoda lental es muy importante ya que, inicialmente, el campo
lental es bastante más extenso que la placoda y se extiende a zonas
aledañas en las que coexisten células que luego forman parte de
otras placodas. Esta delimitación más precisa de regiones se ha
denominado también como “refinamiento” en la distribución de
potencias de desarrollo.
c) Luego de dicho efecto, el mesénquima cefálico desaparece casi
por completo de la región interpuesta entre la placoda lental y la
vesícula óptica. De esta forma ambas estructuras epiteliales, ya
determinadas, interactúan directamente e inician una serie de
interacciones que poseen tanto efecto morfogenético como

histogenético (SC 10.5. El papel morfogenético del contacto placoda
lental-vesícula óptica).
d) El contacto entre los epitelios de la placoda lental y de la vesícula
óptica produce efectos de desarrollo sobre ambos. Por un lado, la
vesícula óptica actúa como población celular permisiva sobre el
campo lental y estimula su diferenciación en placoda lental. Por otro
lado, señales de la placoda operan en forma determinante y/o
permisiva sobre el neuroepitelio de la vesícula óptica y hacen que se
diferencie en sentido de retina neural. Esta acción de la placoda se
pone de manifiesto cuando se impide el contacto de la vesícula óptica
con el campo lental; en ese caso la región superficial de la vesícula
óptica no se diferencia en retina neural sino que adquiere las
características del epitelio pigmentario de la retina.
e) El hecho señalado indica que el mesénquima cefálico periocular
tiene la capacidad de redireccionar la evolución del neuroepitelio de
la vesícula óptica en sentido de epitelio pigmentario. Esta acción se
pone de manifiesto en experimentos en los que se elimina la placoda
lental y toda la vesícula es rodeada por el mesénquima cefálico
periocular. En esta situación, la vesícula óptica no se invagina y todo
el epitelio de la vesícula se diferencia en una capa de epitelio
pigmentario. Vale decir, no se forma retina neural. Este efecto se
deduce también observando el desarrollo normal; normalmente la
región de vesícula óptica que entra en contacto con la placoda lental
se diferencia en retina neural, mientras que la que queda rodeada por
mesénquima periocular se diferencia en epitelio pigmentario de la
retina. Estas acciones diferentes de la placoda y el mesénquima
periocular tienen efecto morfogenético e histogenético ya que
garantizan que en la hoja interna de la copa óptica (que recibió la
acción de la placoda) se forme la retina y que la hoja externa (que
recibe la acción del mesénquima) rodee externamente a la retina
neural.
f) El mesénquima periocular desempeña también un papel central en
el desarrollo de la retinotopía; vale decir, en la instalación de
las polaridades (dorsoventral y nasotemporal) que definen
el patterning de las células ganglionares de la retina necesario para
la elaboración de los mapas de proyección retinogeniculada derecha
e izquierda. La función visual requiere que las conexiones
retinogeniculadas se realicen entre neuronas que ocupan puntos
correspondientes de la retina y del núcleo geniculado lateral. Esto es
posible debido a que en ambos órganos se instalan polaridades
espaciales queasignan propiedades específicas de lugar o
posición a las neuronas que ocupan diferentes posiciones a lo largo
de dos ejes espaciales. Estos ejes son instalados en la población de

células ganglionares de la retina por medio de sistemas de
señalización celular espacialmente organizados o gradientes
generados por el mesénquima periocular.
g) El mesénquima cefálico-periocular participa también en
el patterning de la lente controlando la ejecución diferencial, en
función del espacio, de las actividades de proliferación y
diferenciación celular (SC 10.1. La influencia de la asimetría sobre la
organización espacial de los CCD: el eje superficie-profundidad de la
vesícula lental).
h) Finalmente, desde el punto de vista estructural, el mesénquima
cefálico periocular participa en el desarrollo del ojo con una
versatilidad llamativa. Analizado a lo largo del eje radial del ojo,
genera una variedad de tejidos conectivos estructuralmente
diferentes y con distintas funciones; considérense las diferentes
cubiertas oculares y el humor vítreo. Por otro lado, cada una de estas
cubiertas posee diferencias estructurales y funcionales típicas en
diferentes regiones a lo largo de la circunferencia que va desde el
centro de la córnea al nervio óptico. También participa en la
generación de estructuras específicas del ojo como la zónula de Zinn,
cápsulas de la lente y del humor vítreo, entre otras.
SC 10.4. El campo ocular del extremo cefálico de la placa
neural. V. Flores
Algunos experimentos clásicos mostraron que el extremo cefálico de
la placa neural posee potencia para generar retina neural y que,
además, posee propiedades de campo.
Cuando el extremo cefálico (prosencefálico) de la placa neural, de las
etapas de fines de la gastrulación, es disecado y llevado a un medio
de cultivo rodeado de mesénquima (explanto), se diferencia en una
estructura vesicular epitelial similar a la vesícula óptica. El explanto
así cultivado, pese a que a) está formado por las dos mitades,
derecha e izquierda, del extremo de la placa neural, y que b) durante
el desarrollo embrionario normalmente origina dos vesículas ópticas
que luego generan dos retinas con simetría bilateral, en las
condiciones de cultivo señaladas solo origina una vesícula óptica.
Cuando en el explanto arriba señalado (extremo anterior de la placa
neural) antes del cultivo es dividido en sus dos mitades, derecha e
izquierda, cada mitad exhibe capacidad para originar una vesícula
óptica.
Estos experimentos muestran, tempranamente, durante la
gastrulación, que el extremo anterior de la placa neural se comporta
como un campo morfogenético. Por dicho motivo, el extremo

anterior de la placa neural fue denominado clásicamente, por algunos
investigadores, como territorio presuntivo ocular de la placa
neural y, por otros, como campo ocular (Fig. SC 10-4-1 A).
Cuando los experimentos arriba descritos se repiten en función del
tiempo, se constata que la propiedad de campo se mantiene durante
un cierto período y luego desaparece. Sobre la base de otros
resultados experimentales de cultivos de explantos, combinados con
análisis de biología celular y molecular, se ha postulado que la
escisión del campo ocular en dos campos, uno derecho y uno
izquierdo, depende de una acción ejercida por el mesodermo axil
precordal que normalmente subyace en dicha región de la placa
neural. Por un lado, las alteraciones espontáneas en la formación de
dicho mesodermo se asocian estadísticamente a fallas en la escisión
del campo ocular y, por otro, cuando se interfiere experimentalmente
la migración del mesénquima precordal se mantiene por más tiempo
la propiedad de campo.
Las observaciones mencionadas permitieron proponer que la ciclopía
(presencia de un único ojo de ubicación medial) podría ser el
resultado de una falla en el proceso por medio cual el campo ocular
se escinde en dos. En tales circunstancias se formaría una sola
vesícula óptica y, en consecuencia una sola copa óptica y, en rededor
de la misma, el mesénquima cefálico se organizaría formando las
cubiertas de un único ojo medial.
Varias observaciones recientes, con nueva tecnología, llegan a
conclusiones similares. Confirman la existencia de una región del
extremo cefálico de la placa neural precursora de las células que
integrarán las vesículas ópticas. Esta región puede ser identificada
por la expresión local selectiva de las proteínas factores de
transcripción Pax2 y Pax6. Poco tiempo luego de la expresión de
estos factores, las células de la placa precordal, que se continúa
caudalmente con el mesénquima precorda,l inician la síntesis de
la proteína señal Shh. Al parecer, la activación de la vía de
señalización de la señal Shh en las células de la región medial del
campo ocular, de un modo no conocido, inhibe el desarrollo en
sentido ocular y escinde el territorio o campo ocular inicial en dos
regiones laterales con potencia para formar retina (Fig. SC 10-4-1 B).
Una vez activada la señalización vía Shh en la línea media,
desaparece en dicha región la expresión de los factores de
transcripción Pax2 y Pax6, en tanto que en las regiones derecha e
izquierda continúa su expresión. El factor de transcripción Pax2 se
expresa selectivamente en la región ocupada por células que
quedarán confinadas al pedículo óptico, y el factor Pax6 se expresa
{kind=link}
en células que integrarán la copa óptica. Éstas últimas son las que,
dependiendo de interacciones celulares ulteriores, con el
mesénquima cefálico regional o con la placoda lental, originarán
diferentes tipos celulares de la retina (SC10.3. El papel del
mesénquima cefálico-periocular como integrador de la morfogénesis
y la histogénesis ocular).
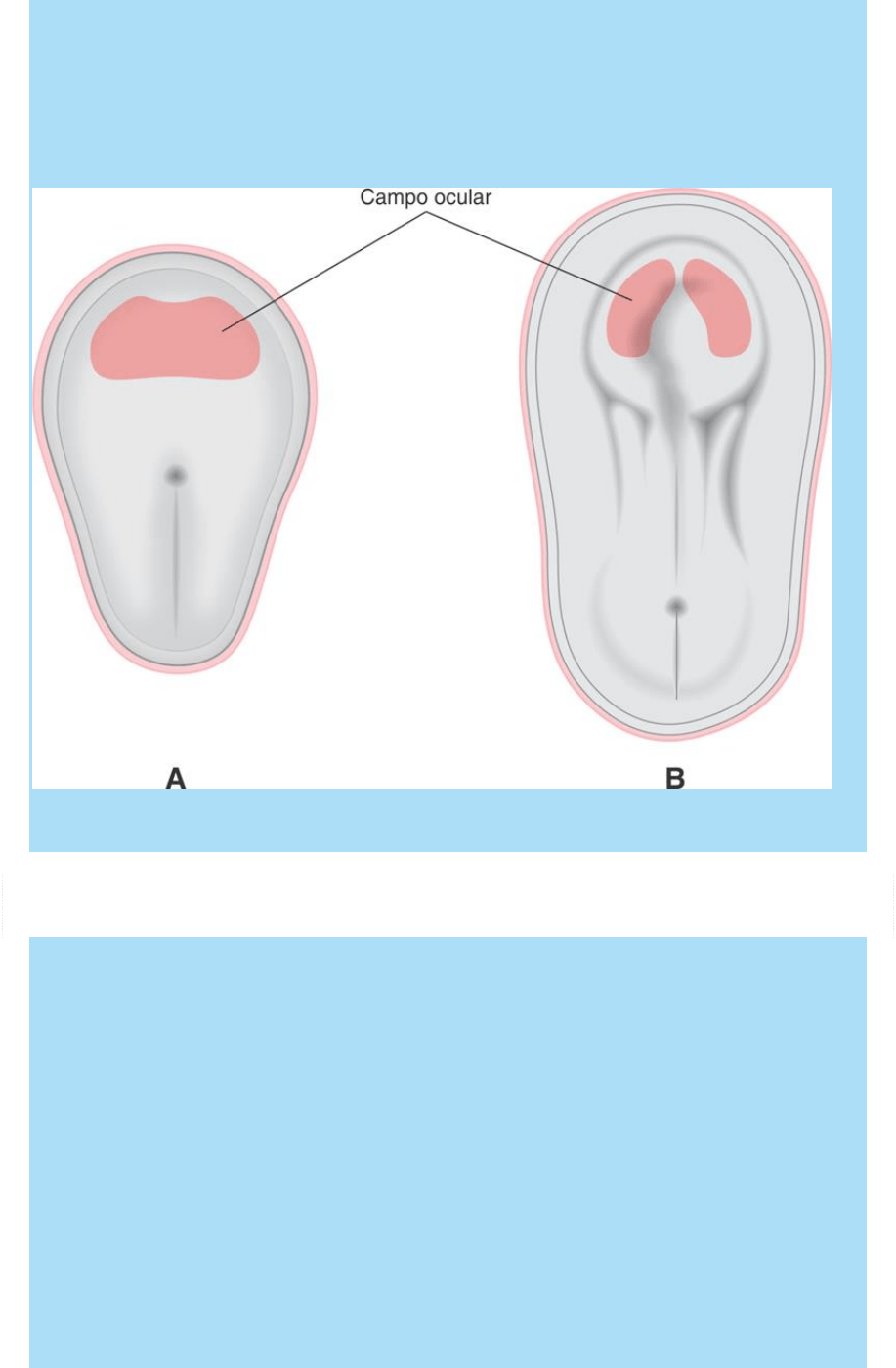
Fig. SC 10-4-1. Representación esquemática de la ubicación del
campo ocular. Vista dorsal. A. Mediados de 3ª SD. B. Fines de 3ª SD.
SC 10.5. El papel morfogenético del contacto placoda lental-
vesícula óptica. V. Flores
Las interacciones placoda lental-vesícula óptica no poseen solo
papeles permisivos o determinantes referidos al destino futuro de las
células que los componen. Las propiedades adhesivas y las
intensidades de las fuerzas interfaciales entre los epitelios
interactuantes también poseen un papel morfogenético. Tal papel
resulta del hecho de que las fuerzas desarrolladas a) modelan
temprana y transitoriamente a las poblaciones interactuantes, b) los
cambios de forma tempranos, al ser base de procesos futuros,
repercuten más adelante en la morfogénesis global y c) generan
disposiciones espaciales de los tejidos que posibilitan interacciones
futuras con otras poblaciones celulares.
Entre las interacciones con papel morfogenético merecen
comentarse las siguientes (varias otras figuran en la literatura):

a) La adhesión entre las superficies basales del ectodermo prelental
y del epitelio neural de la vesícula óptica no es solo resultado de la
intensidad de la fuerza interfacial entre ambas. Este proceso es
facilitado por un desplazamiento de las células del mesénquima
cefálico, fuera del área de contacto epitelial, que favorece la
adhesión. El desplazamiento del mesénquima fuera del área de
contacto y la consiguiente adhesión entre los dos epitelios permiten
que las fuerzas morfogenéticas necesarias para la invaginación,
generadas en cualquiera de los dos epitelios, se transmita también al
otro. Se considera que este fenómeno explica cómo ambos epitelios
se curvan, simultáneamente, como láminas concéntricas con el
mismo radio de curvatura.
b) Otro fenómeno involucrado en el control de la invaginación
conjunta de ambos epitelios es la tensión que ejerce el ectodermo
perilental sobre los bordes de la placoda lental durante la
invaginación. Durante la invaginación aparece una tensión, en el
plano del epitelio perilental, que se revela por el hecho de que si, en
el momento de la invaginación, se realiza una pequeña incisión
rectilínea de orientación tangencial al borde de la placoda, la línea de
incisión se transforma rápidamente en un ojal. Este hecho revela que
los bordes de la incisura están sometidos a tensión (Fig. SC 10-5-1).
La importancia morfogenética de la tensión ejercida sobre los bordes
de la placoda se revela por el hecho de que una incisión realizada en
derrededor de la placoda ocasiona una invaginación acelerada y
excesiva. La vesícula lental, en lugar de quedar ubicada en posición
normal, rodeada por los bordes de la copa óptica, queda
completamente incluida dentro de ella. Vale decir, completamente
rodeada por la hoja interna de la copa óptica, ocupando el lugar que
correspondería al humor vítreo. Así, la tensión y el tiempo durante el
cual la placoda se mantiene unida al ectodermo parece poseer un
efecto sobre la intensidad del plegamiento de ambos epitelios.
{kind=link}
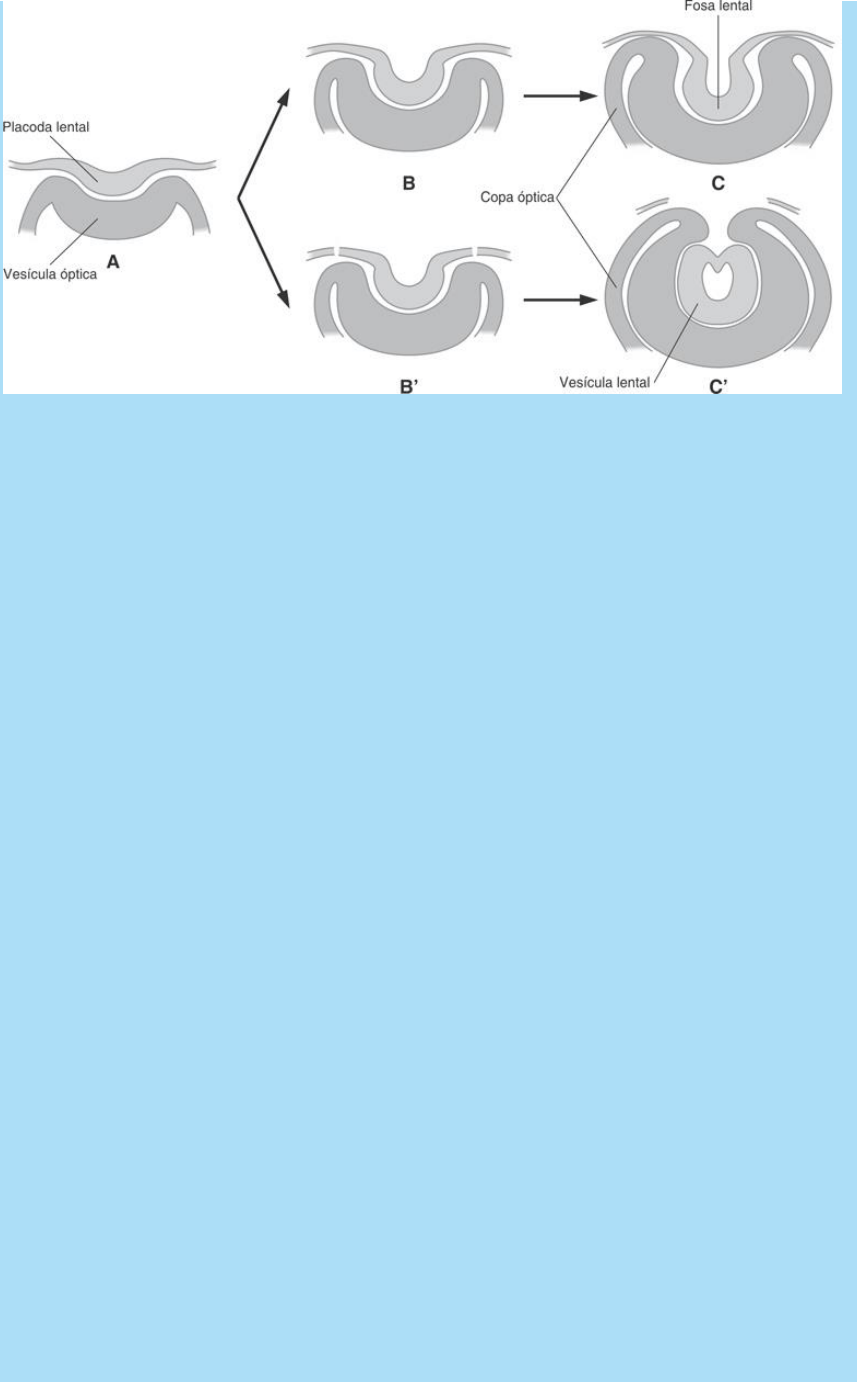
Fig. SC 10-5-1. Representación esquemática de la experiencia de
sección del epitelio ectodérmico perilental. A-C. Invaginación normal
de la placoda lental. B’. Ilustra el estado inmediato posterior a la
cirugía. C’. Evolución luego de la cirugía. La invaginación de la copa
óptica es exagerada y encierra a la vesícula lental.
c) El contacto entre las superficies basales de ambos epitelios
permite interacciones que generan cambios en el patrón de proteínas
que ambos epitelios expresan en sus superficies basales y que
secretan hacia el mesénquima y conforman sus respectivas
membranas basales. La evidencia experimental indica que la
composición molecular de dichas membranas basales es importante
en cuanto a definir interacciones futuras con el mesénquima
circundante: 1) en el caso de la hoja interna de la copa óptica, con la
cara profunda del cristalino y el mesénquima precursor del cuerpo
vítreo y 2) en el caso de la cara anterior del cristalino y la superficie
basal del ectodermo (futura córnea) con el mesénquima que forma
los estromas de la córnea y del iris (véase siguiente punto).
d) Un fenómeno similar al descrito ocurre en la zona de contacto
transitorio entre las superficies basales del epitelio del borde de la
fosa o la vesícula lental recién formada y el epitelio ectodérmico
superficial antes de que ambos se desprendan. En dicha interfaz, al
principio, no se introduce el mesénquima circundante. Durante dicho
tiempo de interacción, los epitelios interactuantes generan
membranas basales que programan el reingreso de mesénquima en
dicha zona. El reingreso del mesénquima cefálico se produce en
forma de dos capas deslaminadas: 1) una de las láminas migra sobre
la superficie anterior de la lente y forma la membrana
iridopupilar; 2) la otra lámina migra sobre la membrana basal del
ectodermo superficial y formará el estroma de la córnea. Este
fenómeno modela también la cámara anterior del ojo que queda
ubicada entre las dos láminas de mesénquima mencionadas. Estos
ejemplos ilustran cómo las interacciones mencionadas
en c y dprograman futuras interacciones con otros tejidos y
estructuraciones espaciales futuras de éstos (Fig. SC 10-5-2).
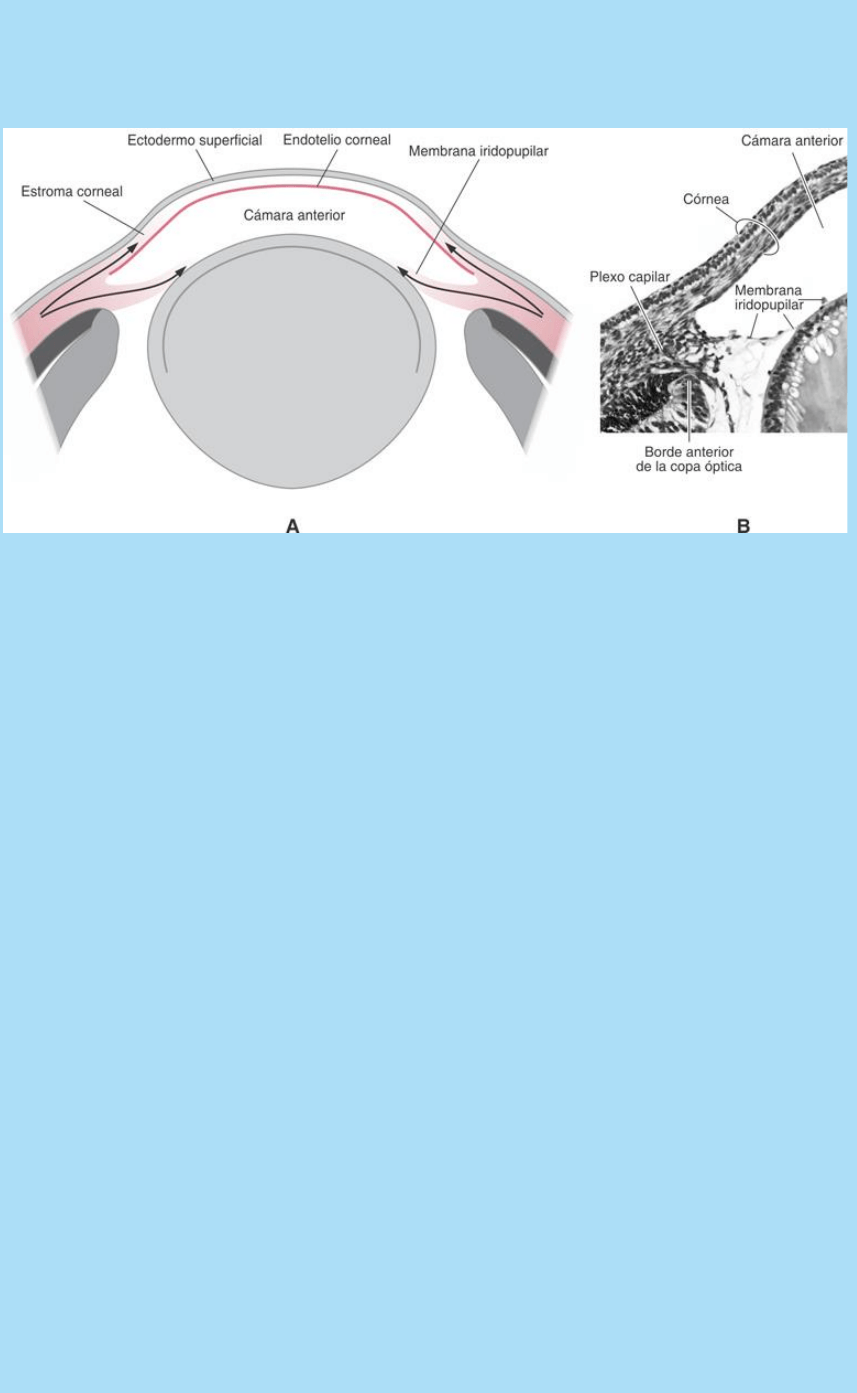
Fig. SC 10-5-2. A. Esquema que ilustra las oleadas migratorias de
mesénquima cefálico que forman el estroma de la córnea y del iris.
La primera oleada formó el endotelio corneal. La segunda oleada
forma el estroma del iris y la tercera oleada forma el estroma corneal.
B. Fotografía de corte histológico del borde superficial de la copa
óptica, el limbo esclerocorneal y la vesícula lental (Embrión humano;
8ª SD). Entre el endotelio corneal y la membrana iridopupilar en
formación se está formando la cámara anterior. En el limbo
esclerocorneal se observa un plexo capilar que se comunica con
ramas del plexo vascular hialoideo. La hoja externa de la copa óptica
es un epitelio pigmentado.
e) Se considera que la extensión del área de contacto entre la
vesícula óptica y la placoda lental posee papel morfogenético. Dado
que se trata de poblaciones celulares que, desde el punto de vista
físico, se comportan como materiales maleables, la intensidad de la
fuerza interfacial opera como variable que regula el área de contacto
entre ambas superficies. Por otro lado, se considera que la presión
hidrostática del líquido contenido en cavidades epiteliales también
posee papel morfogenético ya que genera una tensión tangencial
sobre las células que puede modificar significativamente sus
comportamientos de desarrollo. En el caso de la vesícula óptica se
considera que la presión hidrostática posee una connotación
adicional. El proceso morfogenético de adhesión entre el ectodermo
prelental y la vesícula óptica, y la extensión del área de
contactoentre ambos, depende, por un lado, de la intensidad de la
fuerza de adhesión interfacial entre ambas y, por otro, de la
{kind=link}

posibilidad de que cada uno de dichos epitelios se adapte a la forma
del otro; cuanto mejor se adapten uno al otro, mayor será la superficie
de contacto entre ellas. Una vesícula óptica con presión interna
normal tiene un área de contacto diferente del de una vesícula
“desinflada” que se amolda mejor al epitelio. Este efecto se pone de
manifiesto cuando se realiza un pequeño orificio en la pared de la
vesícula que permite la filtración del líquido interno. Esta situación
conduce a diversos tipos de malformaciones oculares. Se ha
postulado que una presión intravesicular relativamente baja permitiría
una zona de contacto amplia y ello conduciría a vesículas lentales
grandes y hojas internas de la copa óptica también de mayor tamaño.
Por el contrario, una presión intraocular alta llevaría a situaciones
inversas. En ambos casos, en los demás tejidos se producirían
modificaciones compensatorias en el crecimiento de los tejidos
tendientes a lograr la armonía entre todos los elementos que
componen el ojo. La imposibilidad de lograr tal armonía llevaría a
diversas alteraciones que incluyen tamaño anormal de los ojos.
f) El hecho planteado en el punto precedente ha sido señalado
también como una característica peculiar del ojo en el sentido de que
la influencia recíproca entre epitelio ectodérmico prelental y epitelio
de la vesícula óptica, además del carácter localizador (SC 10.2. El
campo lental del extremo cefálico del embrión. Interacciones
celulares y vías de señalización involucradas en su constitución y
evolución. Véase también el punto precedente) posibilita una cierta
armonía en las proporciones de los órganos derivados de ellos. Este
hecho se pone de manifiesto cuando, experimentalmente, se realizan
disociaciones de los epitelios prelental y vesículas ópticas de
animales que tienen ojos grandes y pequeños y se los reasocia en
recombinaciones recíprocas. De dichas asociaciones surgen copas
ópticas y lentes armónicos que no poseen el tamaño de los ojos
grandes ni pequeños de los animales de los que se obtuvieron los
elementos interactuantes sino tamaños intermedios.
Todos los hechos experimentales señalados ponen de manifiesto la
enorme importancia y los diversos efectos de desarrollo, que poseen
las interacciones mediadas por contactos a veces muy transitorios
entre poblaciones celulares en desarrollo.
SC 10.6. La elaboración de mapas de proyección receptor
periférico-centro: el mapa retino-geniculado. V. Flores
La visión es el sentido que con mayor precisión y rapidez permite
analizar información espacial. La visión integra información
proveniente de todos los puntos del espacio que conforman el campo
visual. Para la conducta animal, es vital discriminar con precisión las

posiciones relativas de los puntos que emiten, o reflejan, luz. Ello
permite percibir los objetos del mundo en sus posiciones relativas
reales.
La retina, y el resto de las estructuras oculares, poseen una
organización que permite discriminar y registrar con eficiencia los
puntos del campo visual. La información visual “registrada” en la
retina en forma espacialmente discriminada es luego enviada al
centro por medio del nervio óptico. De nada serviría la discriminación
espacial lograda en la retina si la ésta no se conservara a lo largo de
la vía visual hasta llegar a la corteza visual. La información visual
captada por cada célula fotorreceptora es transferida hasta las
columnas corticales del área visual primaria de la corteza occipital, a
través de cadenas de neuronas conectadas por varias sinapsis.
La información visual es procesada, en parte, en la propia retina y,
desde la neurona ganglionar de la retina (NGR) es llevada al núcleo
de relevo, el núcleo geniculado lateral (NGL), que luego la transfiere
al área 17 de la corteza occipital o área visual primaria o V1. Dado el
carácter polisináptico de la vía visual, existen estrategias de
establecimiento y mantenimiento de contactos sinápticos tendientes
a lograr que la discriminación espacial lograda en la retina no se
pierda en el trayecto desde las células fotorreceptoras a las columnas
corticales del área V1. Es decir, que la información proveniente de
distintos puntos del campo visual no se “mezcle” pues se perdería la
información referida a las posiciones relativas de dichos puntos en el
campo visual.
A lo largo de la evolución se han seleccionado estrategias de
desarrollo que tienden a lograr una correspondencia topográfica
entre “puntos” de los neuroepitelios receptores de los órganos de los
sentidos y “puntos” de las áreas centrales a las que dicha información
es proyectada. Tales correspondencias topográficas entre puntos de
uno y otro se denominan mapas de proyección. En el caso de la vía
visual existe un mapa de proyección retino-geniculado y también
un mapa de proyección genículo-cortical (Recomendamos
consultar en textos de Anatomía la organización de la vía visual
y, en textos de Histología y Fisiología, la organización de la
retina, del núcleo geniculado lateral y de la corteza visual).
La posibilidad de elaborar mapas de proyección entre neuroepitelios
receptores periféricos y áreas de proyección centrales depende de
procesos que operan en diferentes niveles de regulación del
desarrollo de circuitos en el sistema nervioso central: a) la definición
de categorías de neuronas que relacionen un neuroepitelio receptor
Este documento contiene más páginas...
Descargar Completo
Cap 16 SC Flores.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.