
SC 0.1. EL CONCEPTO DE DESARROLLO EMBRIONARIO DE
ORGANISMOS PLURICELULARES. V. Flores
El desarrollo embrionario de los organismos pluricelulares es,
básicamente, un fenómeno de incremento de complejidad:
a)como hecho ontogenético, el desarrollo queda caracterizado por
el conjunto de procesos cuya operación conduce a la génesis de la
complejidad supracelular a partir del nivel de organización unicelular,
la CH; b) como hecho estructural, el desarrollo consiste en la
generación de entidades correspondientes a niveles de organización
supracelulares (tisular, orgánico, etc.) emergentes de interacciones
entre elementos, más simples, de niveles subyacentes y c) como
procesoautoorganizado, la génesis de la complejidad pluricelular,
considerada como totalidad o aun considerando su aspectos
parciales, es el resultado neto de la operación integrada de
combinatorias de CCD que se ejecutan interactivamente y de un
modo temporal y espacialmente organizado. El conjunto de
procesos que acontecen desde el estado inicial (Ei: CH) hasta el final
(Ef: individuo terminalmente diferenciado) incluyen:
a) La generación, por mitosis, de un número de células del orden de
1012 o 1013 células según la especie (a partir de 1 célula).
b) Característica esencial de estas divisiones celulares es que las
células resultantes se mantienen vinculadas por contactosy por
medio de moléculas que secretan al espacio entre ellas. Así, entre
todas generan un medioambiente internodenominado matriz
extracelular. Ambos procesos (desarrollo de uniones intercelulares y
formación de matriz extracelular) llevan a la generación de fuerzas
de cohesión y medios de comunicación (intercambio de
información) entre células.
c) La operación fuerzas interfaciales c-c y c-mec es esencial para
la organización pluricelular. Además, la operación de tales fuerzas, y
su regulación, permiten desplazamientos celulares y la generación
de diversos tipos de arreglos espaciales cambiantes y
característicos, pasando a través de formas de organización
transitorias hasta llegar a las que corresponden a las de los tejidos,
órganos, etc. terminalmente diferenciados.
d) Durante el incremento en el número de células, también se
producen, debido a interacciones entre ellas y con el ambiente,
fenómenos de diferenciación celular generándose varias centenas
de tipos y subtipos celulares.
e) La generación de tipos celulares diferentes involucra también la
expresión de tipos particulares de moléculas que, operando
como señales, sirven al efecto de mediar interacciones entre las
nuevas células de modo que continúen sus interacciones de
desarrollo. Así, unas influyen sobre el desarrollo de las otras.
f) Los procesos de comunicación mediados por señales entre células
que operan durante el desarrollo embrionario tienen
como papel principal instalar una red informativa de mensajes
extracelulares que regula epigenéticamente en forma global y/o
local la operación temporal y espacialmente organizada de los CCD.
g) Al final del desarrollo, la red informativa extracelular posibilita la
comunicación apropiada entre los órganos terminalmente
diferenciados y posibilita, en consecuencia, la integración funcional
de todos los tejidos, órganos, aparatos y sistemas.
h) Durante el desarrollo, la red informativa de mensajes
extracelulares permite que el proceso de diferenciación celular se
realice de un modo organizado en tiempo y espacio. Ello es posible
debido a que muchos de dichos procesos de señalización
espacialmente estructurados están instalados por poblaciones
celulares con función informativa, denominadospoblaciones
celulares organizadoras o centros señalizadores. Estos
organizadores aparecen en momentos y lugares definidos del
embrión e instalan polaridades (distribuciones asimétricas de
moléculas) señal cuyas concentraciones, las células, son
diferencialmente sensibles. Tales estructuraciones se
denominan campos morfogenéticos. Y sirven específicamente al
efecto de organizar en el espacio los procesos de determinación y
diferenciación celular (SC 0.5.
El concepto de determinación.
Potencia y significado evolutivos
; SC El concepto de diferenciación
celular. Criterios que definen el grado de diferenciación
; SC
3.4. Concepto de campo morfogenético).
i) Incluidos entre los CMD epigenéticamente regulados se
encuentran las reprogramaciones o “reseteos” de la información
genética experimentados por las células en momentos y lugares
definidos. Los reseteos irreversibles de la información genética, que
preceden a la diferenciación celular (denominados
de determinación celular), son especialmente importantes ya que
hacen que las células exhiban comportamientos estables durante
períodos prolongados de tiempo, en algunos casos durante toda la
vida del individuo (SC 0.5.
El concepto de determinación. Potencia y
significado evolutivos
; SC El concepto de diferenciación celular.
Criterios que definen el grado de diferenciación
).
j) La determinación celular, por su irreversibilidad,
instala direccionalidad al proceso global. El desarrollo es
un fenómeno vectorial. En prácticamente todas las especies el
desarrollo se describe en términos de sucesión de estados de
complejidad creciente. Ello es así debido a que, si bien es continuo,
posee saltos cualitativos, asociados a las sucesivas
reprogramaciones, que justifican su descripción en términos
de cambios de estado. Tales cambios de estado poseen sentido.
Vale decir, se cumplen en una sucesión temporal
característica pues en cada estado no sólo se elaboran sus
características sino que sesientan las bases del siguiente.
k) Todas las características fenotípicas del individuo terminalmente
diferenciado se hallan implícitas (en potencia) y surgen a partir de
un estado ordenado inicial, constituido con elementos estructurales
e informativos aportados por las gametas e integradas en la CH
(SC
Polaridad de la CH y organización citoplasmática. Evidencias
experimentales
).
l) Debido a que existe un estado inicial y a que en cada estado se
prepara el siguiente, el desarrollo procede ‒en cualquier momento
que se lo considere‒ como si fuera la expresión de un programa de
desarrollo previamente establecido. El desarrollo es, en efecto, la
ejecución de un programa cifrado en términos moleculares y que se
ejecuta por medio de interacciones entre moléculas. De tales
comportamientos moleculares surgen los comportamientos celulares
y los demás niveles de organización.
m) Los CMD característicos de cada estado dependen de moléculas
informativas que interactivamente generan flujos
informativos célula A → matriz extracelular → célula B citoplasma
→ núcleo B → citoplasma B→ matriz extracelular → célula A.
En cada estado, en cada uno de los compartimentos celulares
específicamente involucrados en el desarrollo, se expresan
patrones típicos de moléculas informativas. Estos patrones
típicos de moléculas van cambiando en función del tiempo de un
modo tal que cada uno de ellos conduce, a su vez, a la expresión
de nuevas combinatorias de moléculas y en relación con dichos
cambios moleculares los CCD se van modificando típicamente en
función del tiempo y espacio.
n) Así, el programa global de desarrollo de la mayor parte de las
especies se ejecuta en forma epigenética, integrada y progresiva.
Sin embargo, el programa global se desarrolla en forma de muchos
módulos de programación parcialesejecutados simultáneamente
y en forma integrada (regulada por interacciones múltiples
cooperativas) pero en distintas poblaciones celulares. Ninguna
población celular ejecuta todos los módulos informativos que
componen el programa completo sino módulos parciales de
programación que corren (run) en paralelo, pero el resultado es

integrado y globaldebido a que se ejecutan epigenética o
interactivamente.
ñ) A partir de una configuración inicial, se van generando diversas
formas, ramas o bifurcaciones que son otros tantos modos de
ejecución parcial del programa. Dichas ramas o bifurcaciones se
relacionan con la aparición de diferentes tipos celulares. Así,
cada estirpe celular implementa un módulo en particular de ejecución
del programa global.
El programa global incluye todos los diferentes conjuntos o
combinatorias de CMD característicos de cada uno de los tipos
celulares del individo. Pero cada tipo ejecuta un módulo que
corresponde a un porcentaje muy bajo de la información
contenida en el ADN. El programa es pasible de sufrir en momentos
críticos del desarrollo, a lo largo de su propia ejecución, diversos tipos
de reseteos o reprogramaciones que permiten en forma de
bifurcaciones ir generando los diversos tipos celulares.
SC 0.2. LA FORMA CELULAR Y EL EFECTO MORFOGENÉTICO
DE LOS CAMBIOS DE FORMA CELULAR. V. Flores
Con el objeto de simplificar las descripciones histológicas es común
homologar la forma de las células a formas o cuerpos geométricos
simples. Las células cuyas tres dimensiones son aproximadamente
iguales (isodiamétricas) se denominan esféricas, poliédricas o
cúbicas. A las células anisodiamétricas, cuando una de las
dimensiones es pequeña respecto de las otras dos, se las denomina
“planas” y cuando una de ellas es visiblemente mayor que las otras
dos se las denomina “cilíndricas”. También se las suele comparar con
objetos comúnmente conocidos; así, se describen células fusiformes,
piriformes, estrelladas, etcétera.
La forma de las células depende de varios factores intrínsecos y
extrínsecos. Por un lado depende de la organización del
citoesqueleto. La forma celular que menor energía requiere es la
isodiamétrica. Una esfera es isodiamétrica; su diámetro es el mismo
en todas las direcciones del espacio. Abandonar la forma esférica
implica la operación de fuerzas y, en consecuencia, consumo de
energía. Mantener una forma anisodiamétrica requiere una
organización particular del citoesqueleto e interacciones de
adhesión con algún elemento externo más rígido o consistente
que las células. Las células epiteliales anisodiámétricas (planas o
cilíndricas) puestas en suspensión en un medio de cultivo
rápidamente pierden su forma típica y adquieren una forma que
fluctúa alrededor la esférica. Ello se debe a que estando en un medio
líquido carecen de puntos de apoyo para las fuerzas que
mantienen una forma diferente de la esférica.
Aun cuando el citoesqueleto de célulaa en suspensión puede generar
fuerzas, éstas sólo producen deformaciones suaves o pequeñas
prolongaciones que sobresalen sobre una forma global que fluctúa
alrededor de la esférica. Sólo cuando las células en cultivo toman
contacto unas con otras, forman agregados y se compactan, o
cuando se depositan sobre un sustrato rígido, se adosan y cambian
de forma. Este fenómeno sólo ocurre en tanto existan fuerzas de
adhesión intersuperficiales (interfaciales) entre las células del
agregado o entre las células y el sustrato. En ausencia de fuerzas de
adhesión a elementos que operen como soporte mecánico
(Véase: Placas de adhesión focal, en cualquier texto de Biología
Molecular), las células no pueden cambiar de forma de modo
estable.
La generación y el mantenimiento de formas muy anisodiamétricas
requiere especializaciones del citoesqueleto y de la región de la
superficie celular que interactúa con el sustrato. Las
diferenciaciones de la membrana plasmática de las células
epiteliales que operan como sitios de contacto o anclaje para
elementos del citoesqueleto son los complejos de unión. De éstos
existen varios tipos y cada uno de ellos posee a) un conjunto
particular de proteínas de membrana, b) proteínas que interactúan
con elementos extracelulares y c) proteínas que interactúan con el
citoesqueleto. Estas diferenciaciones de membrana son estables y
proveen mecanismos de adhesión entre células y entre células y
MEC de larga duración. No ocurre lo mismo con las células que
durante el desarrollo embrionario cambian de forma o migran.
Durante el desarrollo, las células disponen de procesos de adhesión
intercelular o célula-mec mediados por diferenciaciones de
membrana menos estables y más dinámicos.
Todo cambio de forma celular requiere una reorganización general
de los elementos del citoesqueleto (microtúbulos, filamentos
intermedios y microfilamentos) y puede implicar el desensamblado
de los elementos existentes y la generación o ensamblaje de nuevos
filamentos y redes. Estas nuevas redes de filamentos tienen
organizaciones espaciales que dependen de las posiciones que
ocupan en la superficie celular los sitios de adhesión a otros
elementos. Las células migratorias, por ejemplo, tienen la capacidad
de realizar cambios muy rápidos de forma y de adhesión diferencial.
Poseen sitios de contactos focales o placas de adhesión focal en
las zonas de contacto o interacción con la mec. Estos contactos
poseen, por un lado, las moléculas implicadas en el
establecimiento del contacto y, por otro, las moléculas que
regulan el ensamblado-desensamblado, o para degradar algunas
de las proteínas de éste. También poseen moléculas receptoras de
señales que inician vías de señalización intracelular. Algunas de
dichas señales disparan la reorganización del citoesqueleto. La
dinámica del citoesqueleto está regulada por proteínas que regulan
el ensamblado-desensamblado, que confieren estabilidad o labilidad
que generan bifurcaciones, etc. Recuérdese que cada uno de los
tipos de elementos del citoesqueleto tiene su propia dinámina y
que ésta se halla influida por moléculas que las regulan.
(Véase: Placas de adhesión o contactos focales, dinámica de los
elementos del citoesqueleto, en cualquier texto de Biologías
Molecular).
Un cambio de forma celular típico, cuando es realizado por
muchas células de la población, puede producir cambios globales
de la población. Éstos pueden ser cambios de forma y también
cambios de posición en el espacio. Existen muchos ejemplos durante
el desarrollo que ilustran estos efectos morfogenéticos del cambio de
forma celular. La mayor parte de los procesos de invaginación de
epitelios involucran este CCD (SC 09
El cierre del tubo neural).
Con el objeto de ilustrar el papel morfogenético que puede tener el
cambio de forma celular, analicemos un ejemplo teórico simple: el
cambio de forma celular en un epitelio cilíndrico simple (
Fig. SC 0-2-
1 A).
{kind=link}
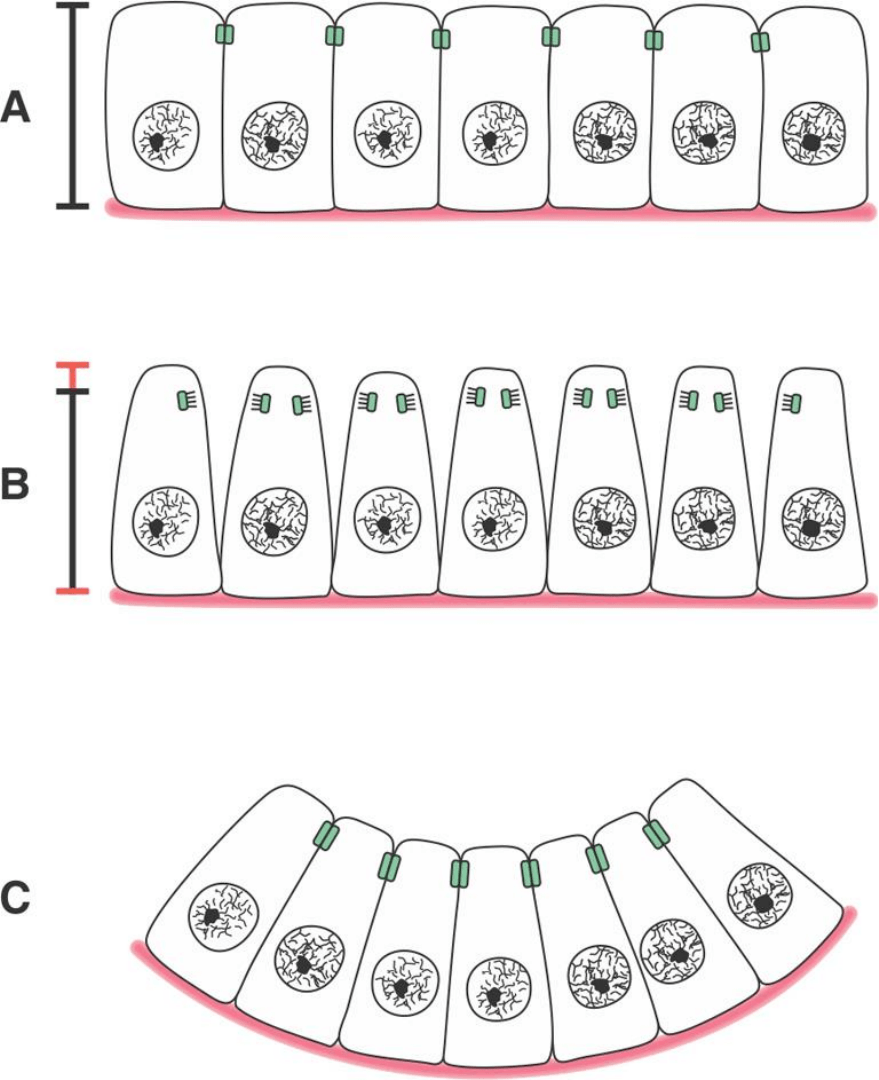
Fig. SC 0-2-1. A. Epitelio cilíndrico simple. La suma de las superficies
apicales es similar a la suma de las superficies basales. B. Resultado
hipótetico de lo que podría ocurrir en un epitelio cilíndrico simple si
las células cambiaran de forma, de cilíndricas a piramidales truncas,
y si no se mantuvieran los complejos de unión. Las regiones apicales
de las células se separarían y las regiones basales retendrían su
adhesión a la membrana basal. El epitelio y su membrana basal
retendrían la disposición planar. C. Resultado del cambio de forma
celular, cilíndrico a pirámide trunca, acompañado de un aumento en
la fuerza de adhesión célula-célula mediada por complejos de unión.
La disminución del área apical y el mantenimiento o incremento de la
fuerza de adhesión intercelular llevan a una curvatura del epitelio y
de la membrana basal en la que apoya.
1) Consideremos un cambio de forma celular tal que las células de
un epitelio simple formado por células prismáticas o cilíndricas se
transformen en piramidales truncas. El cambio de forma de una
célula aislada no permite apreciar el cambio global que podría
producir (
Fig. SC 0-2-1 A y B). Puede apreciarse que el cambio
implica tanto una elongación como un adelgazamiento del extremo
apical de las células. Se propone que este tipo de cambio requiere
una reorganización del citoesqueleto de la región apical de las
células y la generación de fuerzas mecánicas. Los estudios con
microscopia electrónica permiten concluir que este cambio involucra
a microctúbulos, microfilamentos y filamentos intermedios. Los
microtúbulos se disponen preferencialmente en la dirección de la
elongación y el tratamiento de las células con colchicina,un
inhibidor de la polimerización de la tubulina, inhibe la elongación
del extremo apical de estas células. Por otro lado, el adelgazamiento
del extremo apical requiere fuerzas que operen tangencialmente en
el plano del epitelio y que ello implicafenómenos contráctiles en la
red terminal apical. La red terminal es una diferenciación local del
citoesqueleto submembranoso en la región apical. Se trata de
una red densa de microfilamentos y filamentos intermedios. Esta
estructura filamentosa y contrácitil le confiere a la célula una rigidez
suficiente como para actuar como apoyo mecánico para las
diferenciaciones de la membrana apical y también capacidad para
soportar las tensiones que se generan en el plano del epitelio.
Los filamentos de la red terminal se insertan en sitios
especializados, los complejos de unión, de la membrana lateral de
células adyacentes. Se propone que, cuando la red terminal se
contrae, la superficie apical se reduce y dicho extremo se adelgaza
con respecto a la superficie basal. Algunos experimentos de
tratamiento de las células con citocalasina B, un inhibidor de la
polimerización de la actina G, muestran que inhibe el cambio de
forma descrito.
2) Si las células pudieran cambiar de forman independientemente
unas de otras, cosa que podría ocurrir si no existieranfuerzas de
adhesión intersuperficial entre sus membranas laterales, el efecto
global sobre el epitelio sería el que se ilustra en
la figura SC 0-2-1B.
3) Por el contrario, si las células cambian de forma reteniendo los
contactos que los unen a las células adyacentes, el cambio de forma
celular se integra en un resultado global diferente que involucra a
toda la lámina epitelial (figura SC 0-2-1 C). El epitelio abandona la
disposición plana y se pliega.
4) Nótese que existe un conjunto de requisitos teóricos que deben
cumplirse para que ocurra un fenómeno de este tipo:
a) La membrana plasmática apical debe plegarse o disminuir su
extensión. La disminución de su extensión puede producirse por
la remoción de parches de membrana por medio de un
proceso similar a una endocitosis. También puede ocurrir si las
células comparten mayor superficie de contacto a expensas de la
membrana apical. Vale decir, parte de la membrana apical pasaría
a ser lateral generando mayor superficie de contacto. Dado que las
células pueden deformarse y regular su superficie de membrana,
el incremento de la fuerza de adhesión interfacial c-c o c-mec se
asocia a un incremento de las superficies de contacto. Esto se debe
a que fuerzas de adhesión muy intensas hacen que las células se
aplasten unas contra otras (este fenómeno se denomina
habitualmente compactación) o contra la mec.
b) Los epitelios asientan sobre una lámina basal plana, una
especialización de la matriz extracelular que constituye lainterfase
de interacción entre tejidos epiteliales y conectivos (en el adulto)
o mesénquima (en el embrión). Se considera que la membrana basal
es más rígida que la célula y sirve también de asiento o soporte
mecánico para el epitelio. Muchos epitelios poseen diferenciaciones
de unión (hemidesmosomas) entre su membrana plasmática basal
y la lámina basal. Nótese que el plegamiento ilustrado en la
figura SC
0-2-1C
implica un cambio en la disposición de la membrana
basal. Así, es un requisito teórico que las fuerzas desarrolladas por
el citoesqueleto en el seno del epitelio debe ser mayor que la
resistencia de la membrana basal a la deformación y menor que
la fuerza de adhesión intercelular.
También es sabido que la deformación de la lámina basal que
acompaña al plegamiento involucra su remodelación.
Teóricamente, este hecho debe hacer desaparecer la tensión a la
que podría estar sometida durante la deformación.
Por todos estos motivos, algunos estudios biofísicos postulan que la
fuerza fundamental que promueve el cambio de forma celular y el
plegamiento del epitelio es fuerza de adhesión interfacial c-c. El
incremento de esta fuerza produciría un aumento de la superficie de
contacto intercelular en la región apical con lo cual el extremo apical
de las células se adelgazaría. Como se señaló, este aumento en la
superficie de contacto se haría a expensas de la membrana apical.
Nótese que, si no existiera una fuerza de adhesión interfacial c-c

mayor que la fuerza de contracción generada por la red terminal, los
extremos apicales de las células se despegarían y se obtendría el
resultado mostrado en la
figura SC 0-2-1 B.
SC 0.3. LA DIFERENCIACIÓN CELULAR Y EL CONCEPTO DE
PATTERNING. V. Flores
El término “diferenciación” es frecuentemente usado en la literatura
para aludir a fenómenos que corresponden a diferentes niveles de
organización (celular, tisular, orgánico e incluso de aparato sistema).
Así, por ejemplo son comunes expresiones tales como
“diferenciación del hepatocito”, “diferenciación hepática”,
“diferenciación del tubo digestivo” o “diferenciación del aparato
digestivo”.
Desde el punto de vista de la biología celular y molecular, el uso del
término en sentido amplio es un tanto impreciso ya que una
explicación detallada debería llevar, en todos los casos, al nivel
celular. Vale decir, a explicaciones acerca de cómo la expresión
selectiva de proteínas específicas de tipo celular lleva a la aparición
de los diferentes tipos celulares terminalmente diferenciados. Así, la
cuestión quedaría siempre planteada en términos de diferenciación
celular o citodiferenciación, y el resultado final sería la descripción de
modos particulares, para cada tipo celular, de expresión del genoma
y de cómo tal expresión se expresa en el fenotipo de cada tipo celular.
Este modo de concebir la diferenciación, sin embargo, no explica
fenómenos correspondientes a niveles de organización
supracelulares.
La visión descrita sería insuficiente para comprender cómo las
células musculares y las mesenquimáticas interactúan y forman
diferentes tipos de tejidos musculares esqueléticos y también
diferentes tipos de músculos esqueléticos.
Si el interés fuera analizar el desarrollo del tejido muscular o, más
aún, el desarrollo interactivo entre tejidos conectivos y tejidos
musculares de modo que ambos durante el desarrollo se organicen
en un músculo en particular, deberían considerarse fenómenos
adicionales en los que los elementos informativos no radican sólo en
las células musculares sino en el mesénquima, los vasos, los nervios,
etcétera.
Analicemos en detalle este ejemplo. Una descripción genérica de
la diferenciación de la célula muscular esquelética, por completa
que fuera, no explicaría fenómenos del nivel de organización
tisular como por ejemplo la existencia de varios tipos diferentes de
tejidos musculares esqueléticos con diferentes proporciones de fibras
rápidas y lentas, diferente tamaño celular, diferente orientación
espacial, diferente número de núcleos y de placas mioneurales y
diferentes modos de relacionarse con los otros tejidos. Tampoco
explicaría fenómenos correspondientes al nivel de organización
orgánico. Cada músculo posee un nombre propio debido a que es
un órgano en particular definido no sólo por ubicación en relación con
el esqueleto y sus inserciones óseas sino también por el modo como
se integran los tejidos muscular y esquelético, los varios diferentes
tipos de tejidos conectivos (aponeurosis, perimisios, endomisio, etc.),
vasos, nervios, tipos de inervación, tamaño de las unidades motoras,
los tipos de alfamotoneuronas que las inervan, los tipos de fibras del
sistema gamma que recibe, etc. Todas las características
mencionadas son esenciales con respecto a la caracterización de
cada músculo como entidad biológica.
Este último planteo, en última instancia, alude al modo como
se regula la organización en el espacio, a procesos de
determinación, de diferenciación, proliferación, etc. de tejidos
que desarrollan conjuntamente y se ensamblan en el espacio
exhibiendo un patrón de organización peculiar que lo distingue
de otros órganos formados por los mismos tipos celularespero
que constituyen entidades biológicas diferentes.
Este último fenómeno que no se reduce sólo a la citodiferenciación
sino a su organización espacial se denomina, en la literatura inglesa,
“patterning”. Adoptamos este término debido a su amplia difusión en
trabajos científicos.
La esencia del concepto de patterning se advierte cuando se
comparan entidades biológicas integradas con los mismos tipos y
subtipos celulares, los mismos tejidos, etc., pero que son diferentes
debido a que los tejidos y células que los componen tienen distintas
proporciones, distintos números y distintas disposiciones espaciales.
Compárense los dedos de la mano entre sí, compárense manos y
pies o, mejor, compárense los mismos elementos anatómicos del
lado derecho con los del lado izquierdo y se advertirá que la
citodiferenciación no explica dichas diferencias. Con el objeto de
identificar, analizar y realizar la experimentación apropiada para
explicar dichas diferencias espaciales se utiliza la noción
de patterning. Así, esta noción alude al modo como los mismos CCD
y CMD se organizan en espacio y tiempo y conducen a diferentes
estructuras cuyas células sufren procesos de citodiferenciación
básicos similares.

El ejemplo que más claramente resalta la cuestión de la organización
espacial de los procesos biológicos es la comparación entre
las estructuras corporales derechas e izquierdas que poseen
representación bilateral (compárense dedos índice derecho e
izquierdo). Es claro que en ambos casos se utiliza la misma
información genética, que en ambos casos se trata de células que
posen la misma historia de determinaciones, pero son
completamente distintos ya que se estructuraron con polaridades
diferentemente orientadas en el espacio.
Tanto es así que las células que forman el “dedo derecho” podrían
formar el “dedo izquierdo” si estuvieran en una posición diferente en
el sistema de referencia establecido por las polaridades que operan
durante el desarrollo. Este ejemplo ilustra con claridad que se trata
de “el mismo fenómeno diferentemente estructurado en el
espacio”. Las diferencias mencionadas se describen en términos de
diferencias en una propiedad de las células en desarrollo
denominada información posicional o información de posición.
Fenómenos de este tipo son de naturaleza eminentemente
epigénética pues requiere recurrir a información que no es
intrínseca de las células sino que está establecida como
un sistema de referencia espacial que organiza la operación de
CCD. Tal entidad informativa que está plasmada en el espacio en el
que las células ejecutan sus CCD ha sido clásicamente
denominada “patrón” (“pattern” en la literatura inglesa). Se trata
de fenómenos de señalización celular espacialmente organizados
(SC 3.4.
Concepto de campo morfogenético). El concepto de patrón,
en su formulación más simple, alude a unaentidad informativa cuya
función de desarrollo es la organización espacial del proceso de
diferenciación celular. En un sentido más amplio alude también a
la organización espacial de CCD. El concepto de patterning alude
al modo como tal entidad informativa se “traduce” en una entidad
con patrón estructural definido.
SC 0.4. LA ORGANIZACIÓN JERÁRQUICA DE LAS VÍAS
EVOLUTIVAS EN EL PROGRAMA DE DESARROLLO. EL
ÁRBOL DE DETERMINACIONES. V. Flores
La CH origina por mitosis todas las células del organismo. En los
vertebrados "superiores" los individuos terminalmente desarrollados
poseen alrededor de doscientos tipos celulares distintos. La CH
posee entonces la capacidad de originar células que durante el
desarrollo van haciéndose diferentes unas de otras.
En los mamíferos, la CH y las blastómeras (hasta el E8c) poseen la
capacidad de originar todos los tipos celulares. La aparición, a lo
largo del desarrollo, de diferentes tipos celulares implica que existen
varias formas posibles de evolución a partir de la CH. A cada una de
las diversas formas posibles de evolución se denomina vía
evolutiva.
Cada vía evolutiva constituye una modalidad particular de expresión
del programa de desarrollo ejecutada por una estirpe celular en
particular. Todas las células de un organismo poseen la información
genética correspondiente al programa global de desarrollo, pero
ninguna estirpe celular expresa al mismo tiempo el programa de
desarrollo global sino un módulo de expresión del genoma en
particular. Así, la información que utiliza cada célula corresponde sólo
a un pequeño porcentaje del genoma. Si bien es difícil estimar, ya
que varía en diferentes especies, algunas estimaciones indican que
en la especie humana, en promedio, cada célula expresa menos del
5% de la información global del genoma.
Se designa con el nombre de potencialidad o potencia
evolutiva (PE), o simplemente potencia, al conjunto de todas las
formas posibles de evolución que podría exhibir una población
celular a partir de un cierto estado del desarrollo cuando es puesta
en diversas condiciones de desarrollo (SC 0.5.
El concepto de
determinación. Potencia y significado evolutivos
). La PE de una
población celular no alude sólo a los distintos tipos celulares que
puede generar durante el desarrollo. Incluye tambiéna todas las
formas posibles de evolución que podría exhibir (todos los
diferentes tipos celulares que podría originar) en diferentes
condiciones de desarrollo. Alude entonces a todas las diversas
formas de expresión del programa de desarrollo que la
población celular tiene habilitada.
El conjunto particular de vías de desarrollo que una población celular
exhibe (el conjunto de tipos celulares que origina) durante el
desarrollo normal (que en general constituye sólo una parte de su
petencia) se denomina significado evolutivo o también destino
evolutivo. Existen CMD que se ejecutan interactivactivamente por
medio de los cuales subconjuntos particulares de células ingresan,
en momentos definidos del desarrollo, en diferentes vías evolutivas
(SC 0.6.
Concepto de acción celular determinante (A c-c D)) y
progresan a través de ellas diferenciándose unas de otras (SC 0.8.
El
perfil evolutivo del grado de diferenciación celular. El papel de las
acciones celulares permisivas
). Estos procesos
constituyen reprogramaciones o reseteos del programa
de desarrollo por medio de los cuales se habilita una modalidad
de expresión en particular y quedan irreversiblemente
inhabilitadas las otras.
La organización de vías evolutivas del programa de desarrollo
Las vías evolutivas que componen la PE de una población celular no
son independientes entre sí. Con el objeto de ilustrar este concepto
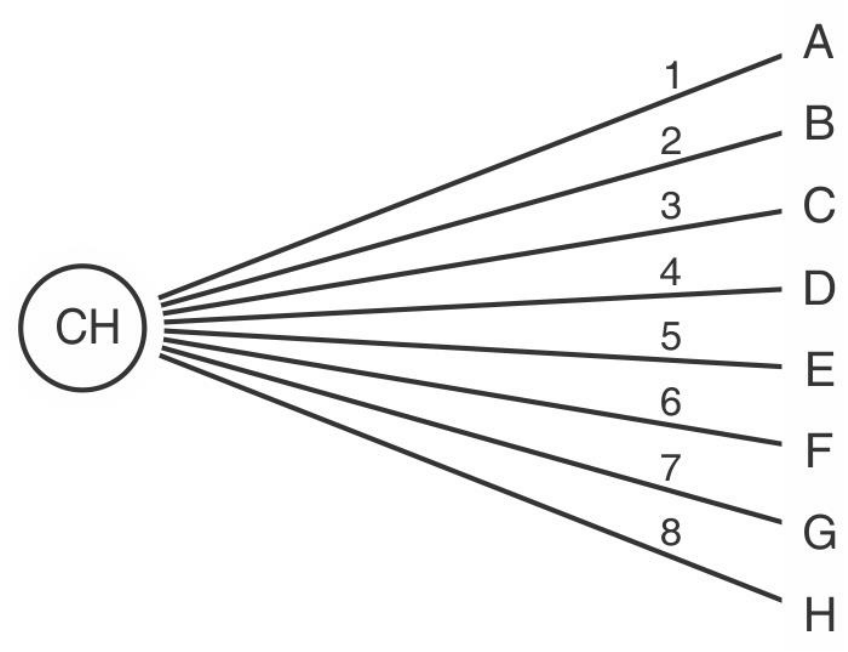
imaginemos una especie hipotética que posee 8 tipos celulares que
se generan por medio de 8 modalidades diferentes e independientes
de expresión del programa de desarrollo. La independencia de las
vías evolutivas estaría representada gráficamente en la
figura SC 0-
4-1
. Existirían 8 modos diferentes de evolución (1 a 8) por medio de
los cuales se generarían los tipos celulares, A a H, a partir de la CH.
Ninguna vía estaría influida por las otras y las células no tendrían
ningún tipo de vínculo entre sí. En una CH hipotética así organizada
todos los módulos de expresión del programa podrían estar
habilitados desde el estado inicial, cada tipo o grupo de células podría
ejecutar uno de ellos, todos los tipos celulares tendrían como única
relación de parentesco (genealogía) su descendencia de la CH; los
diversos tipos celulares podrían determinarse temprana y
simultáneamente y sin la intervención de fenómenos epigenéticos o
interactivos.
Fig. SC 0-4-1. Representación esquemática de las 8 vías evolutivas
(1 a 8) independientes, en un organismo pluricelular hipotético que
{kind=link}
posea 8 tipos celulares diferentes (A a H) no vinculados entre sí.
Todos los tipos celulares estarían determinados desde el principio del
desarrollo.
No disponemos de información que indique que los programas de
desarrollo de organismos superiores y las vías evolutivas que lo
integran estén organizados como ilustra la
figura SC 0-4-1. Los
programas de desarrollo están organizados de modo que las
características del fenotipo se elaboran de lo general a lo
particular y ello hace que las diferentes vías evolutivas se vinculen
entre sí como en una estructura jerárquica de categorías en las
que unas quedan incluidas en, o derivan de, otras.
Analicemos gráficamente este concepto. Consideremos, como en el
ejemplo anterior, un organismo hipotético en cuyo estado de
diferenciación terminal existen ocho tipos celulares diferentes (A a H)
(
Fig. SC 0-4-2). La CH, dado su carácter de estado inicial del
desarrollo, no origina directamente los tipos celulares mencionados,
sino que se comporta como precursora de poblaciones con escaso
grado de determinación que, a su vez, son precursoras de otras cuyo
grado de determinación aumenta en función del progreso del
desarrollo. Así, el desarrollo implica la aparición, en forma sucesiva,
de poblaciones celulares precursoras de otras que tienen cada vez
mayor grado de determinación.
El gráfico muestra que la CH sólo origina a una población 1
que posee toda la potencia del sistema (a-h). Vale decir no ha
experimentado determinación o disminución de su potencia. La
población 1, a su vez, es precursora de las poblaciones 2 y 3, pero
la generación de estas dos poblaciones ya implica una primera
determinación y, en consecuencia, reducción de lapotencia de
cada una de ellas. Así, a partir de la población 2 sólo se pueden
seguir vías evolutivas que conduzcan a la generación de células
del tipo A a D. La población celular 2 redujo su potencia pues ya no
es capaz de originar células del tipo E a H. Éstas, a su vez, caen
dentro de la potencia de la población 3 que ya no puede originar
células del tipo A a D.
En el organismo hipotético descrito se podrán definir tantas vías
evolutivas de citodiferenciación como tipos celulares posea el
individuo adulto. Nótese que ellas se organizan en un árbol de
bifurcaciones. Las diversas secuencias de tipos celulares
identificables a lo largo del desarrollo de dicho organismo
constituyen cada una de las diversas vías evolutivas incluidas
en su programa de desarrollo (
Fig. SC 0-4-3-A).
{kind=link}
{kind=link}
Este documento contiene más páginas...
Descargar Completo
Cap 16 SC Flores.docx
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.