BIOLOGÍA CELULAR III
SISTEMA DE ENDOMEMBRANAS I
● Sistema de endomembranas, también llamadas sistema vacuolar citoplasmático
está constituido por:
Vesículas
Cisternas (sacos aplanados)
Túbulos
● Estas estructuras al combinarse dan origen a las distintas organelas que forman el
sistema de endomembranas
● Las organelas guardan entre sí una interconexión funcional que suele ser indirecta
pero también existe un ejemplo de comunicación directa que veremos mas
adelante
● El sistema de endomembranas se encuentra distribuido por gran parte del
citoplasma celular dando lugar en él a la formación de dos compartimientos:
● Compartimiento intramembranoso: Corresponde a la luz o el lumen de las
organelas del SDE
● Compartimento citosólico: Mientras que la porción citoplasmática que queda
por fuera recibe el nombre de citosol, citoplasma fundamental o matriz
citoplasmática
● Citosol y citoplasma no son estrictamente sinónimos
Sistema de endomembranas:
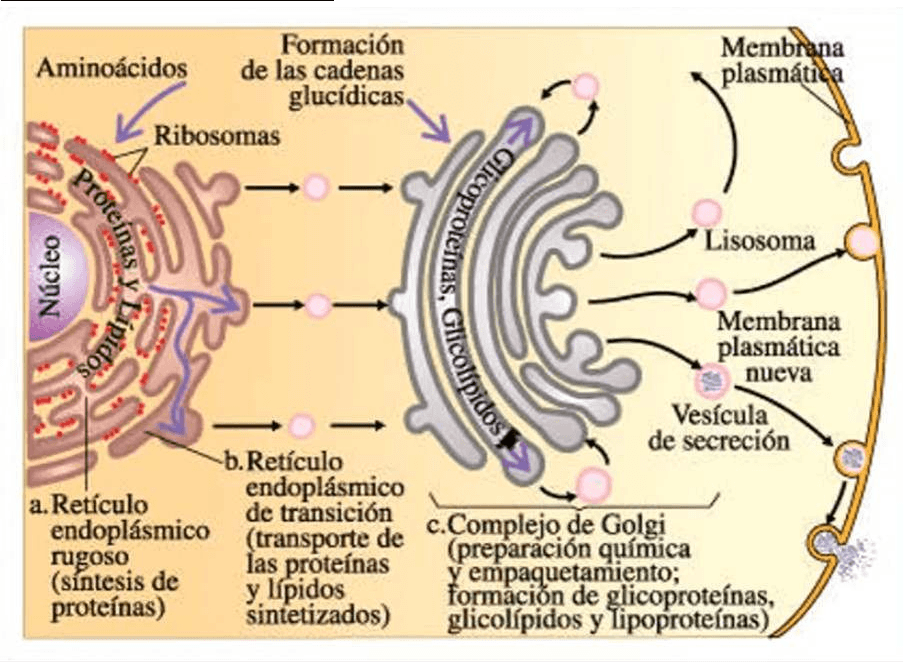
● El sistema de endomembranas comienza por la carioteca o la envoltura nuclear
que está constituido por una membrana interna y otra membrana externa, el
retículo endoplasmático que presenta dos regiones diferentes (REL o REG) (hay un
región de transición entre el REL y REG), luego tenemos el aparato de Golgi,
estructuras membranosos túbulo-vesiculares que corresponden al sistema de
digestión intracelular integrado por el endosoma primario o temprano, el endosoma
secundario o tardío, los lisosomas y luego tenemos un conjunto de vesículas que
fundamentalmente cumplen funciones de transporte a través del citoplasma.
Tenemos por ejemplo: vesículas que transportan sustancias producidas por el REL
hacia el aparato de Golgi, vesículas que comunican las distintas cisternas del
dictiosoma que forma el aparato de Golgi, tenemos vesículas que desde el Golgi
van a la membrana plasmática
● Hablamos entonces de una interconexión funcional, estas vesículas forman parte
de mediadores de esa comunicación o interconexión indirectas entre distintas
organelas del SDE y la comunicación directa la encontramos en la membrana
externa de la carioteca que se continua directamente con las membranas de las
cisternas del REG, incluso la luz que se interpone entre las membranas interna y
externa de la envoltura nuclear o carioteca se continúa directamente con la luz de
las cisternas del RE
● Sobre la superficie externa de la envoltura nuclear aparecen ribosomas, se
sintetizan proteínas que forman parte de la envoltura nuclear o carioteca
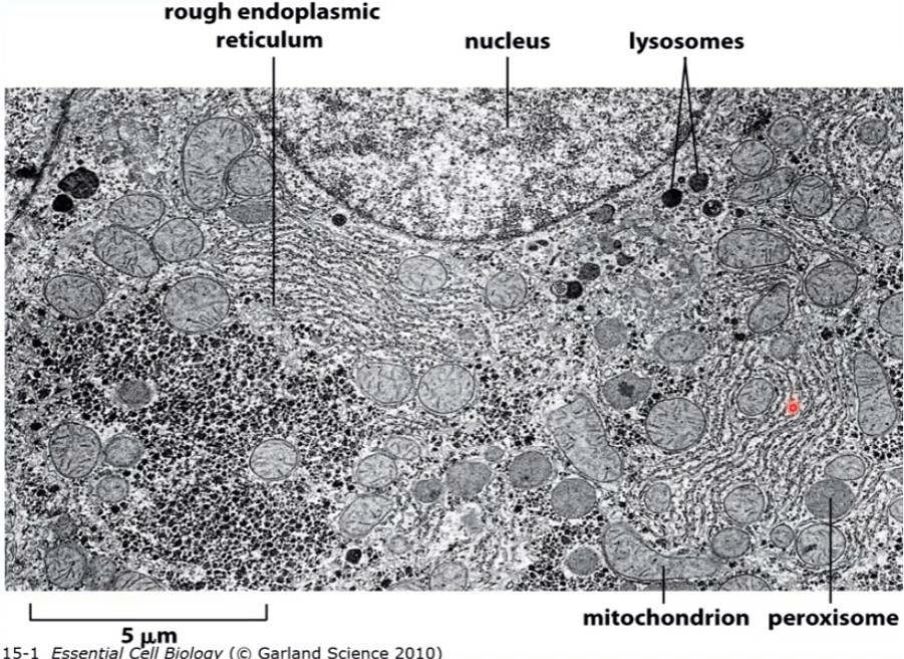
Micrografía electrónica de un hepatocito - organoide membranoso:
Componentes del SDE:
● Envoltura nuclear o carioteca
● Retículo endoplasmático (RE)
● Granular o rugoso (REG o RER)
● Agranular o liso (REL)
● Aparato de Golgi
● Endosomas
● Lisosomas
● Vesículas secretoras
● Sistema vesicular de transito y endocitosis
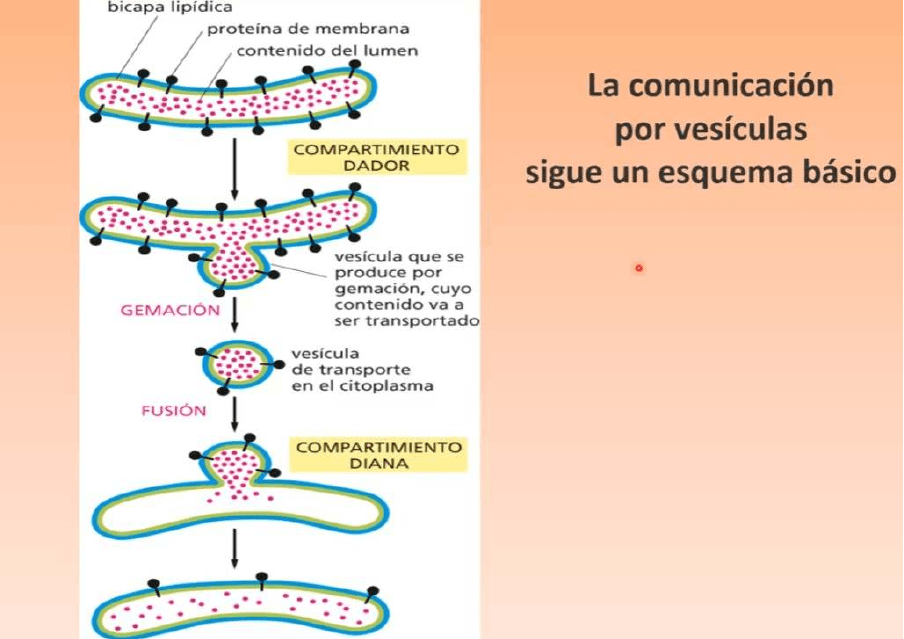
La comunicación vesicular sigue un esquema básico:
● Un compartimento dador que es el que genera la vesícula
● Compartimento diana es quien recibe la vesícula
● La vesícula se origina por GEMACIÓN, se produce una envaginación que se
profundiza cada vez más hasta que queda constituida la vesícula que pierde
contacto con la superficie de la membrana que le dio origen
● Esta vesícula se dirige entonces hacia el compartimento diana, se producirá la
fusión de la membrana de la vesícula con la membrana del órgano que la recibe
● Es muy complejos, existen vesículas que toman direcciones diferentes entonces es
necesario que existe cierta especificidad, existen vesículas con distintas cubiertas
proteicas (seminario 4), van a haber vesículas con cubierta de catrinas, cubierta de
clatomeros de tipo I y de tipo II y además las vesículas tienen expresadas en su
membrana proteínas que actúan como proteínas receptores
Se excluyen del sistema de endomembranas:
● Mitocondrias: organela de doble membrana
● Peroxisomas: organela vesicular formada por una membrana única
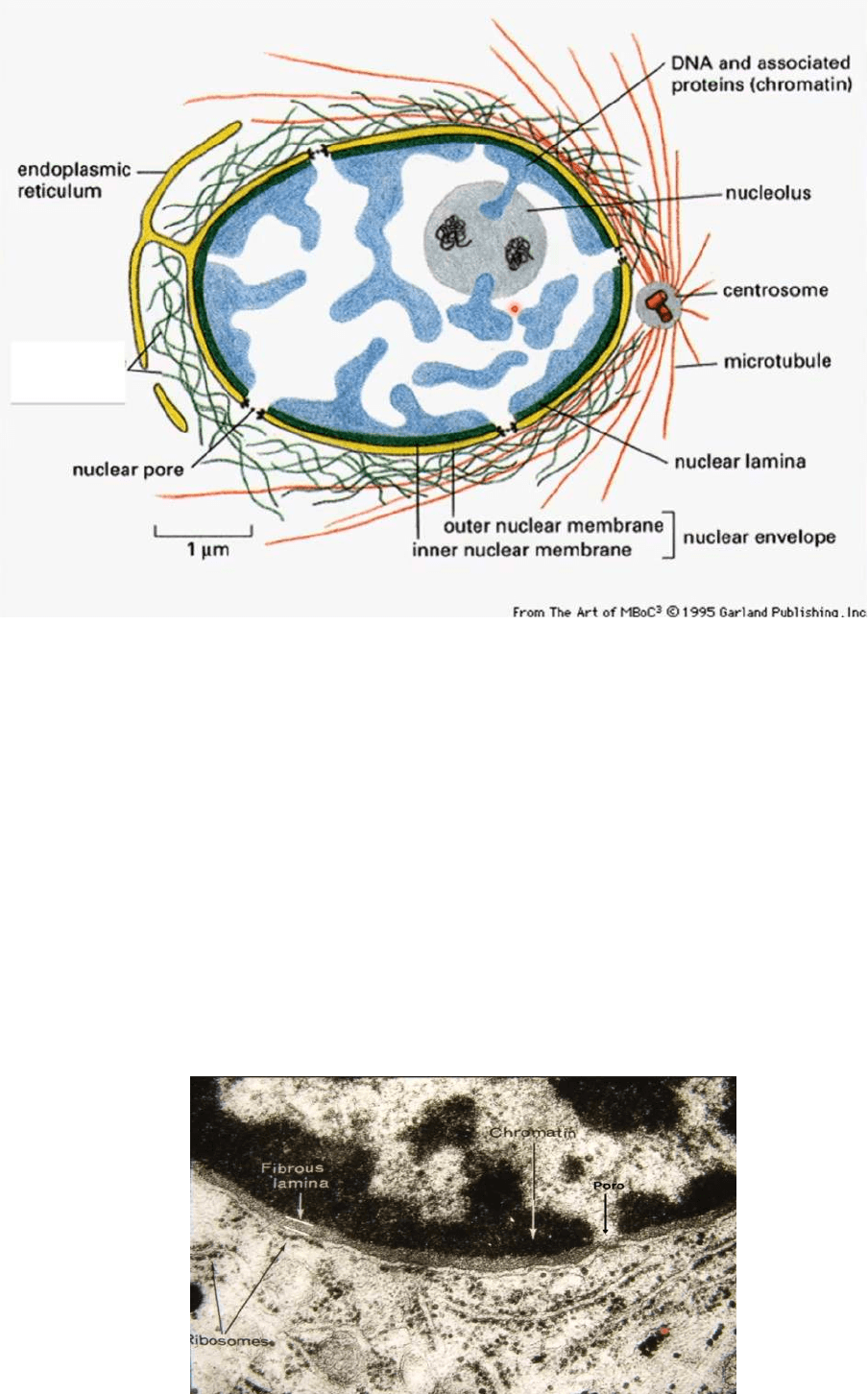
ENVOLTURA NUCLEAR O CARIOTECA
● Se ve el nucleoplasma, se destaca el nucleolo y este núcleo está rodeado por
sáculos aplanados (cada uno de ellos presenta una doble membrana, interna y
externa) y esta membrana está interrumpida en sitios múltiples por los llamados
poros nucleares que permiten la comunicación desde el núcleo hacia el citoplasma
y viceversa, estos poros no están vacíos sino que están ocupados por una serie de
proteínas que llamamos COMPLEJO DEL PORO
● La membrana externa se continúa en forma directa con la membrana del RE y
también hay una continuidad directa entre la luz que media entre la membrana
externa e interna del núcleo y la luz de la cisterna del RER
● La superficie externa de la envoltura nuclear esta rodeada por filamentos que
corresponden a componentes del citoesqueleto y si observamos la superficie
interna de la envoltura nuclear vemos que está revestida por una estructura
verdosa e indica la presencia de la llamada LAMINA NUCLEAR
● La lamina nuclear está formada por filamentos intermedios del núcleos constituidos
por la laminina de tipo A, B y C
● Por lo general en todos los núcleos existe cromatina condensada sobre la
superficie interna de la carioteca aun cuando ese núcleo posea cromatina laxa
● Se ve como se continua la membrana externa de la carioteca con el RER y
ribosomas sobre la membrana externa de la carioteca
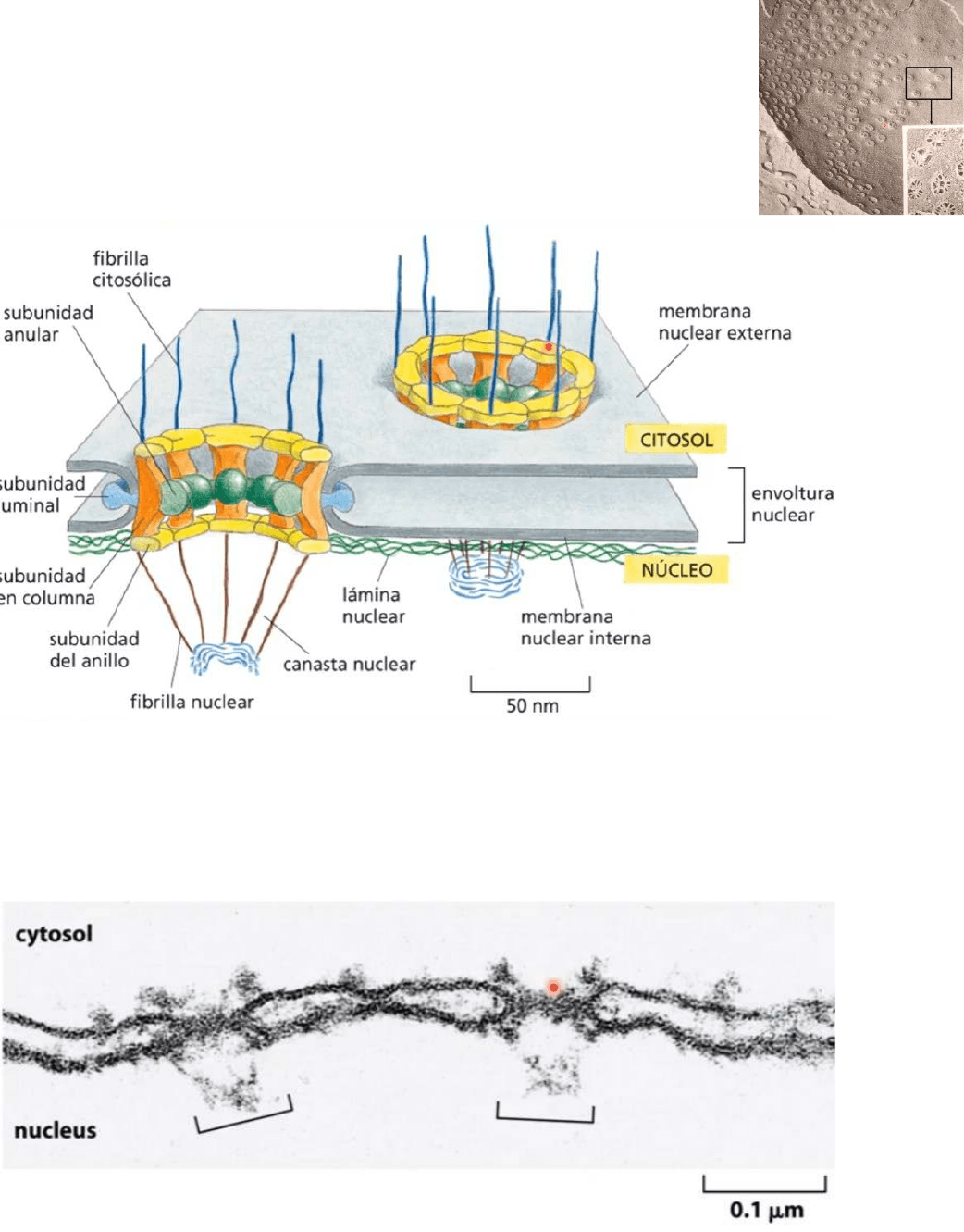
Poro nuclear:
● Los poros nucleares estan ocupados por complejos (NPCs)
nucleoproteicos formado por 30 proteínas diferentes conocidas
como nucleoporinas o proteínas NPC
● El perímetro del poro está tapizado por proteínas columnares que tienen un
dominio ubicado hacia el citosol, otro dominio ubicado hacia el interior del núcleo,
una porción transmembrana
● Existen otras proteínas asociadas como las proteínas fibrilares orientadas tanto al
citosol como al interior del núcleo y cumplen una función importante en el
transporte de moléculas desde el citosol al núcleo y desde el núcleo al citosol
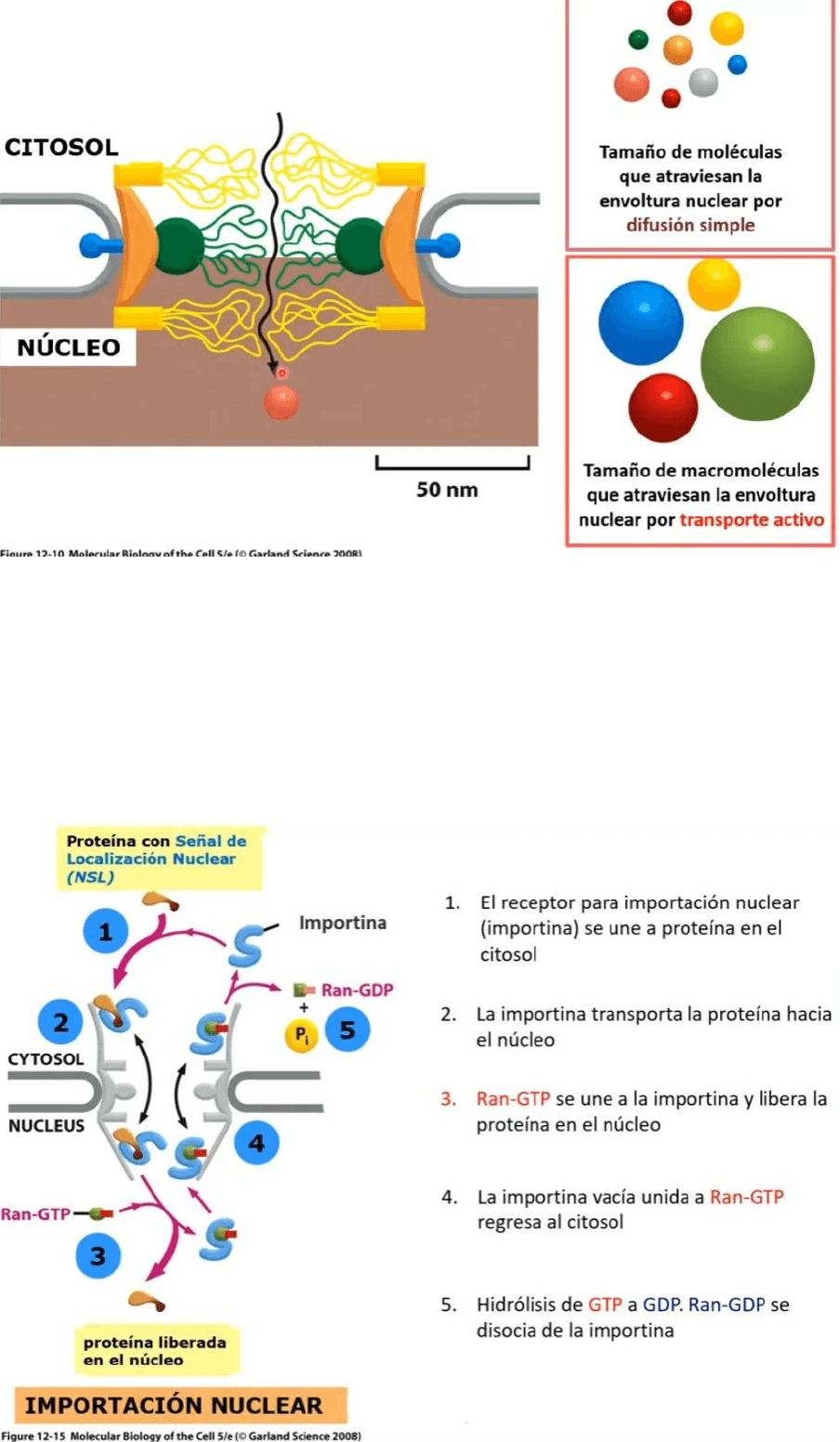
● El transporte activo o pasivo a través de los poros nucleares depende en parte del
tamaño de las moléculas
● Algunas moléculas podrán atravesar por difusión simple y algunas necesitarán
energía
IMPORTACIÓN NUCLEAR - Transporte de proteínas citosol al núcleo
● Este tema se desarrolla más adelante
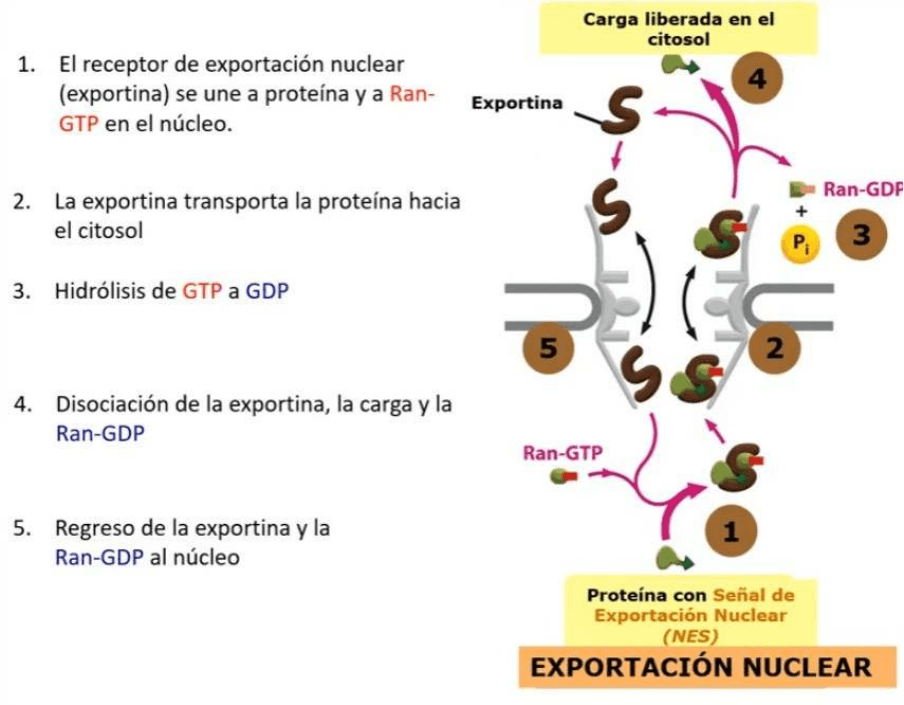
● Vamos a transportar una proteína que esta localizada en el citosol y que tiene que
llegar al núcleo
● La célula sabe que esta proteína debe ser transportada al núcleo porque tiene una
secuencia de aminoácidos especifica que constituyen una señal de localización
nuclear
● Esta proteína es reconocida por una proteína receptora que se llama IMPORTINA
(el núcleo importa: MNEMOTECNIA)
● La importina se une a la proteína y ambas atraviesan el poro nuclear desde el
citosol hasta el núcleo, cuando llega al núcleo se va a unir a una proteína RAN que
es una GTPasa, esta GTPasa cuando esta activa está ligada a GTP
● Tenemos un conjunto formado por la importina, la proteína que fue transportada al
núcleo y el Ran-GTP
● Inicialmente la proteína va a ser liberada al núcleo y continúan unidas la importina
con el Ran-GTP que van a pasar del núcleo al citosol
● Una vez que la importina unida al Ran-GTP llegan al citosol se produce la hidrólisis
del GTP, ahora Ran queda unida a GDP, esto induce la separación del Ran-GDP
de la importina, la importina queda libre en el citosol y puede ser utilizada
nuevamente en un transporte desde el citosol al núcleo
EXPORTACIÓN NUCLEAR - Transporte de proteínas y ARNm nuclear al citosol
● Desde el núcleo al citosol podemos transportar una molécula de ARNm, el núcleo
ahora exporta
● Esta proteína tendrá una secuencia de aminoácidos especifica que constituye la
llamada “señal de exportación nuclear”. Esa señal es reconocida por una proteína
que llamaremos EXPORTINA
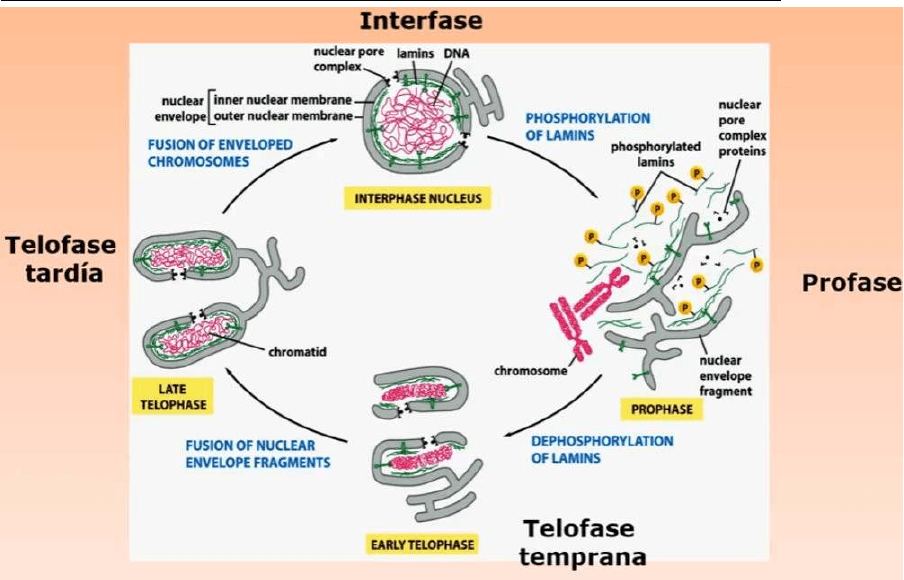
● Ahora tenemos la proteína que tiene que ser transportada del núcleo hacia el
citosol, la exportina y a ello se una Ran-GTP
● Atravesamos el poro nuclear, pasamos al citosol, en el citosol se producirán la
hidrólisis del GTP a GDP, ahora tenemos Ran-GDP que se libera de la
EXPORTINA y la proteína que debe quedar en el citosol
● Esta EXPORTINA será transportada ahora en sentido inverso desde el citosol
hasta el núcleo, en el núcleo podrá volver cumplir un nuevo ciclo de transporte
desde el núcleo hasta el citosol
Ciclo de la carioteca y de la lámina nuclear a través del ciclo celular:
● Sufre cambios o modificaciones cíclicas durante la vida de una célula
● Durante la interfase la características de la membrana nuclear son diferentes a lo
que le sucede durante el periodo de división celular
● La célula en interfase cuando comienza la división celular en un momento dado las
proteínas que forman la envoltura nuclear serán fosforiladas, las proteínas de la
lamina nuclear también serán fosforiladas y esa fosforilación conduce a la
despolimerización tanto de la envoltura nuclear como de la lamina nuclear
● Mas adelante producida la división del material genético entre los núcleos de las
futuras células hijas se va a reconstituir esa envoltura nuclear en torno al material
genético, se produce la REPOLIMERIZACIÓN de la lámina nuclear y de la
envoltura nuclear
RETÍCULO ENDOPLASMÁTICO
● Está constituido por dos porciones diferentes: granular o rugoso y agranular o liso
RETÍCULO ENDOPLASMÁTICO RUGOSO
● Tiene como función principal la síntesis de proteínas
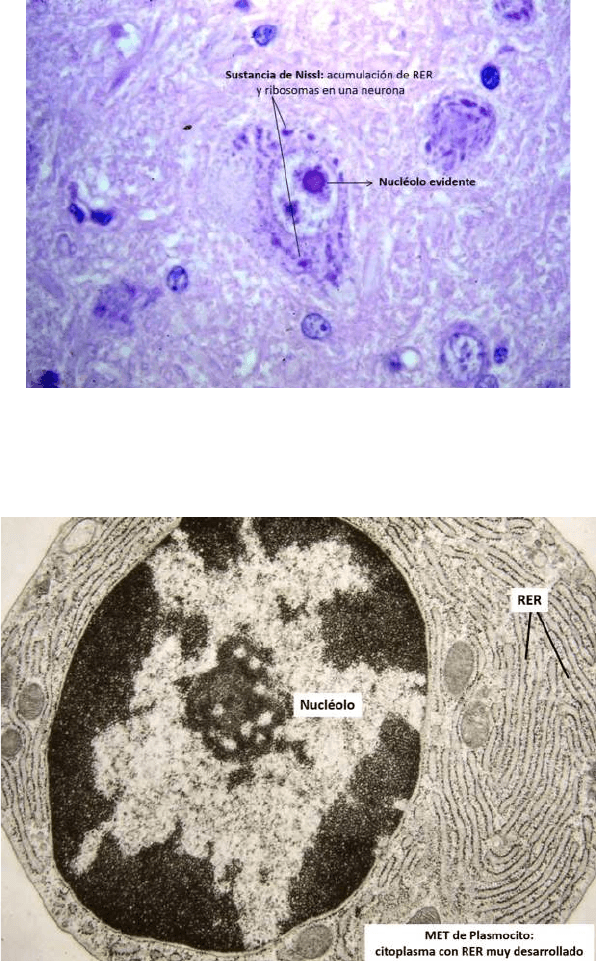
● REG en célula de cultivo método de inmunofluorescencia
● Microscopía óptica de luz convencional con tinción de Nissl, permite que nosotros
veamos en el citoplasma de la células la acumulación de REG y de ribosomas, las
zonas azules son REG y ribosomas libres
● En el núcleo vemos cromatina condensada heterocromatina. NO ESTA SIENDO
TRANSCRITA PORQUE ESTAN EMPAQUETADAS (negra) y cromatina laxa
eucromatina (blanco)
● PLASMOCITO
● Vemos un citoplasma lleno de cisternas del REG, este citoplasma presentaría una
intensa basofilia en M.O porque los ribosomas corresponden a ARN
● La cromatina laxa nos habla de alto nivel transcripcional, descondensada,
desenrollada, data presente en los núcleos de todas aquellas células en donde la
transcripción tiene un nivel importante debido a que las enzimas que llevan a cabo
la transcripción del ADN necesitan introducirse entre las hebras de cromatinas
● Nucleolo esta evidente porque mucha de las funciones del Plasmocito es el
ensamblado de los componentes que constituyen las subunidades ribosomales
menores y mayores
● Existe una estrecha correlación entre función de la célula, su ultraestructura y la
tinción de la célula en un preparado de histología
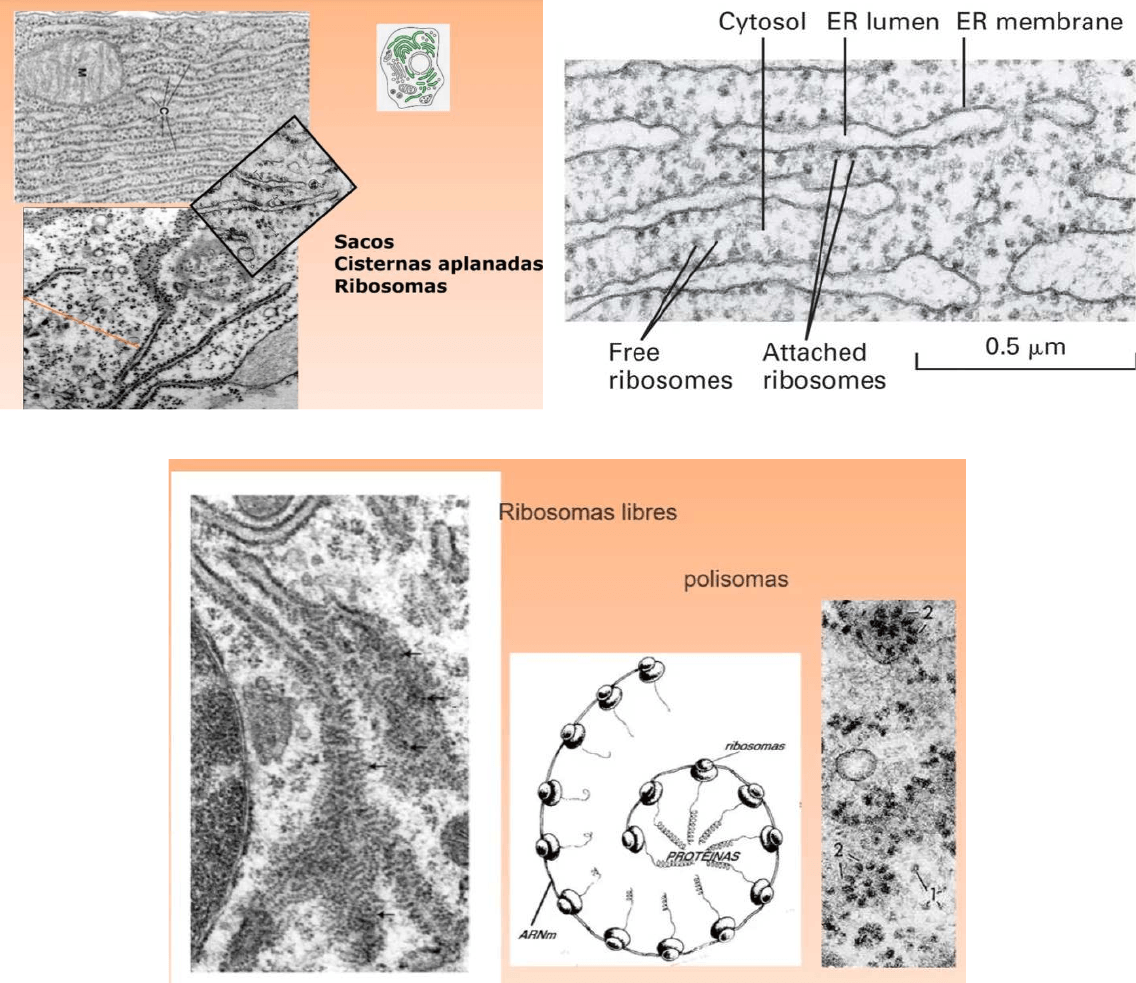
● Esta célula es muy activa en la síntesis de proteínas por eso tiene MUY
desarrollado el REG
● Sintetiza inmunoglobulina o anticuerpos
● Aqui vemos una estructura de polisomas o polirribosomas, estas estructuras se
encuentran en el citosol y podemos observarlo tambien sobre la superficie de
membrana del REG, el ARNm está siendo leído por varios ribosomas
● REG: síntesis de proteínas de membrana y exportación
● Ribosomas: síntesis de proteínas citosólicas, nucleares
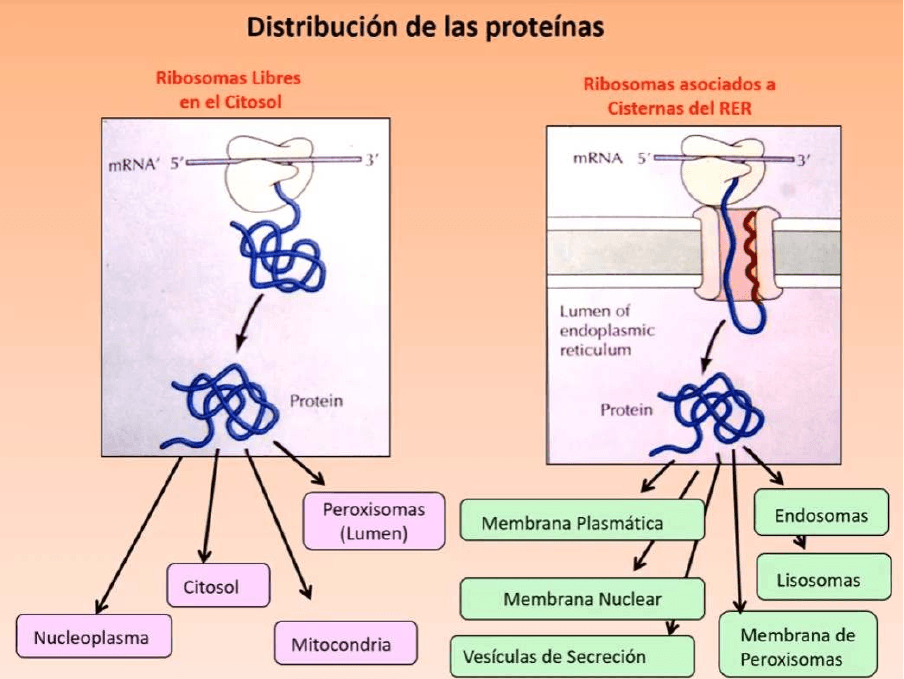
Distribución de las proteínas de los ribosomas libres y los ribosomas asociados:
● Algunas proteínas se sintetizan en ribosomas libres en el citosol, otras en
ribosomas unidos a cisternas del REG
● En la izquierda, los ARNm que se leen en forma completa en ribosomas libres en el
citosol son aquellos mensajeros que codifican para:
● Las enzimas oxidativas, PEROXISOMAS
● Proteínas mitocondriales
● Proteínas del citosol
● Proteínas del núcleo plasma
● En la derecha, los ribosomas asociados a cisternas del REG:
● Proteínas del endosoma
● Proteínas de los lisosomas (hidrolasas ácidas)
● Proteína que forma la membrana de los peroxisomas
● Proteínas de membrana plasmática
● Proteínas de vesículas de secreción
● Proteínas de membrana nuclear
● En todos estos casos de arriba el ARNm comienza a ser leído en ribosomas libres
pero en algún momento determinado esa traducción se detiene y el ribosoma es
llevado al REG, se asocia a su membrana y ahí se reanuda la síntesis de ese
mensaje
Unión de la partícula de Reconocimiento de la Señal (PRS) a la subunidad mayor de
los ribosomas:
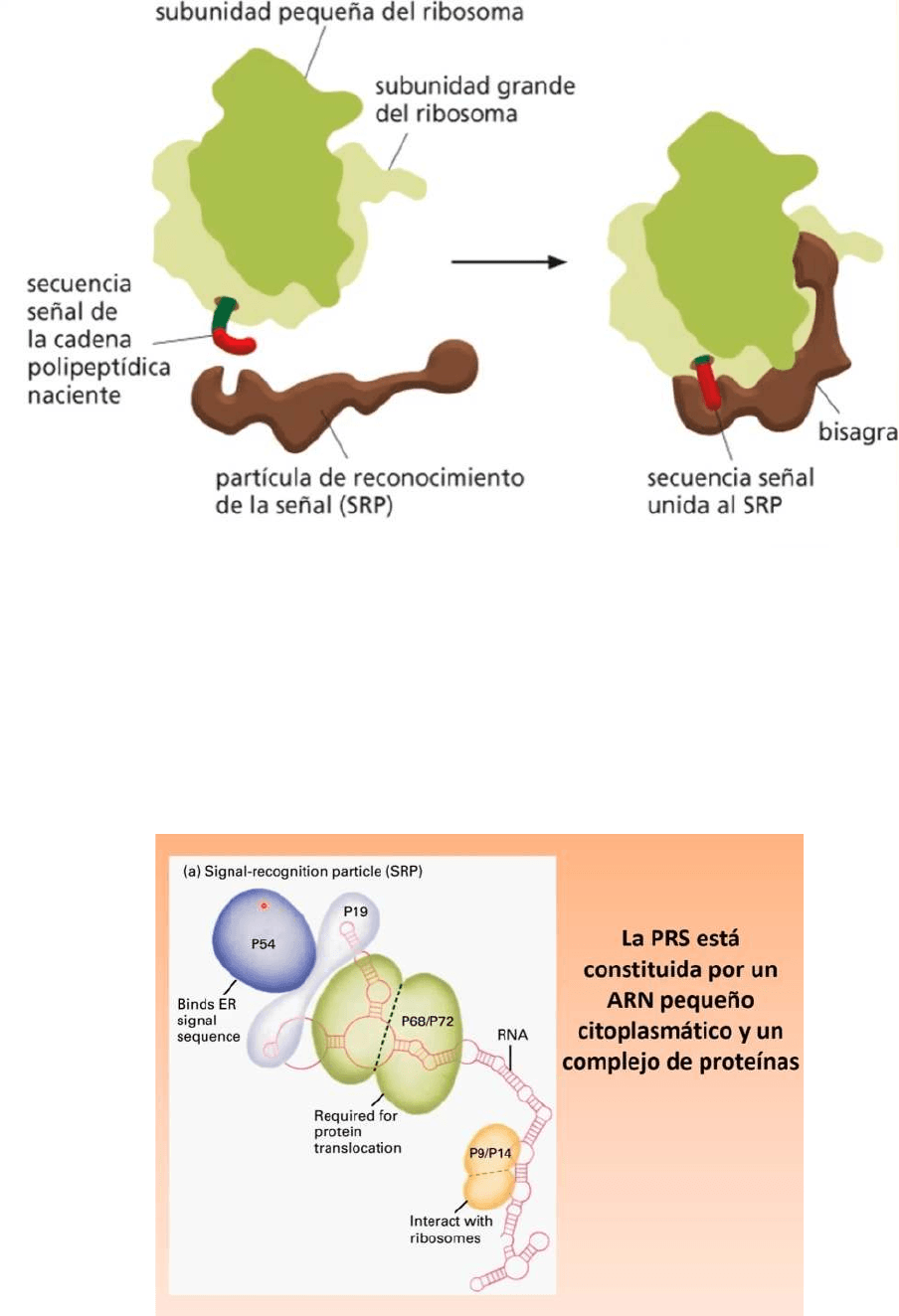
● Un ARNm sabe que será leído de forma completa en ribosomas libres o que
terminará de ser leído en el REG porque existe una partícula de reconocimiento
de la señal (PRS)
● Este PRS se une al péptido señal, el péptido señal es una secuencia de
aminoácidos que se encuentra generalmente en el extremo aminoterminal de una
proteína, todo ARNm comienza a ser leído en ribosomas libros en el citosol,
cuando aparece este péptido señal este péptido señal es reconocido por el PRS,
esta se una al péptido señal, esto determina la inhibición transitoria de la lectura del
ARNm y este ARNm terminará de ser leído en ribosomas adosados al REG
● La PRS está formado por un ARN citoplasmático pequeño de 7S unido a 6
proteínas diferentes
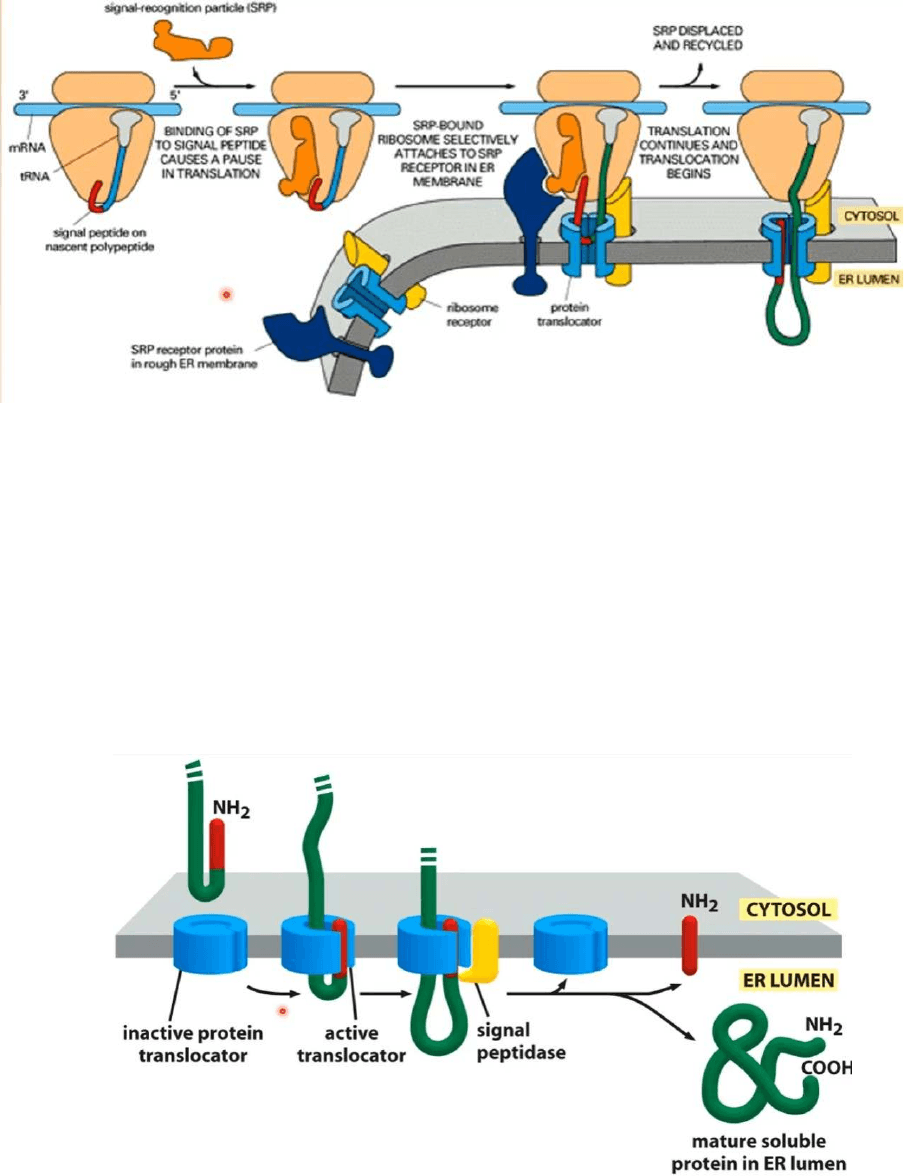
PÉPTIDO SEÑAL Y PRS
● Aqui vemos un ARNm comenzó a ser leído en un ribosoma libre en el citosol, se
esta formando una proteína, apareció un péptido señal, este péptido señal es
reconocido por la PRS, la PRS se une al péptido señal, se inhibe la lectura del
mensajero, todo este complejo es llevado al REG, en el REG tendremos los
siguientes receptores:
● Riboforinas 1 y 2: receptores para la subunidad ribosómica mayor
● Receptor para la PRS: reconocerá a la PRS
● Se une el ribosoma a la cisterna del REG, en consecuencia se reanuda la lectura o
traducción de este ARNm
Translocación de una proteína a la luz de la cisterna del REG
● Proteínas que se estan sintetizando en ribosomas adosados al cisternas del REG y
que estan destinadas a quedarse en la luz de la cisterna
● Tenemos una proteína extremo amino-terminal, extremo carboxilo-terminal, el NH2
es el péptido señal que normalmente está ubicado en el extremo amino-terminal de
la proteína
● Esta proteína será traslocada a través de este canal llamado TRANSLOCON que
permite que la proteína atraviese la membrana del REG y llegue hasta la cisterna
● El péptido señal se separa del resto de la proteína, es degrado en los aminoácidos
constituyentes y esta proteína queda localizada en la luz de la cisterna del REG
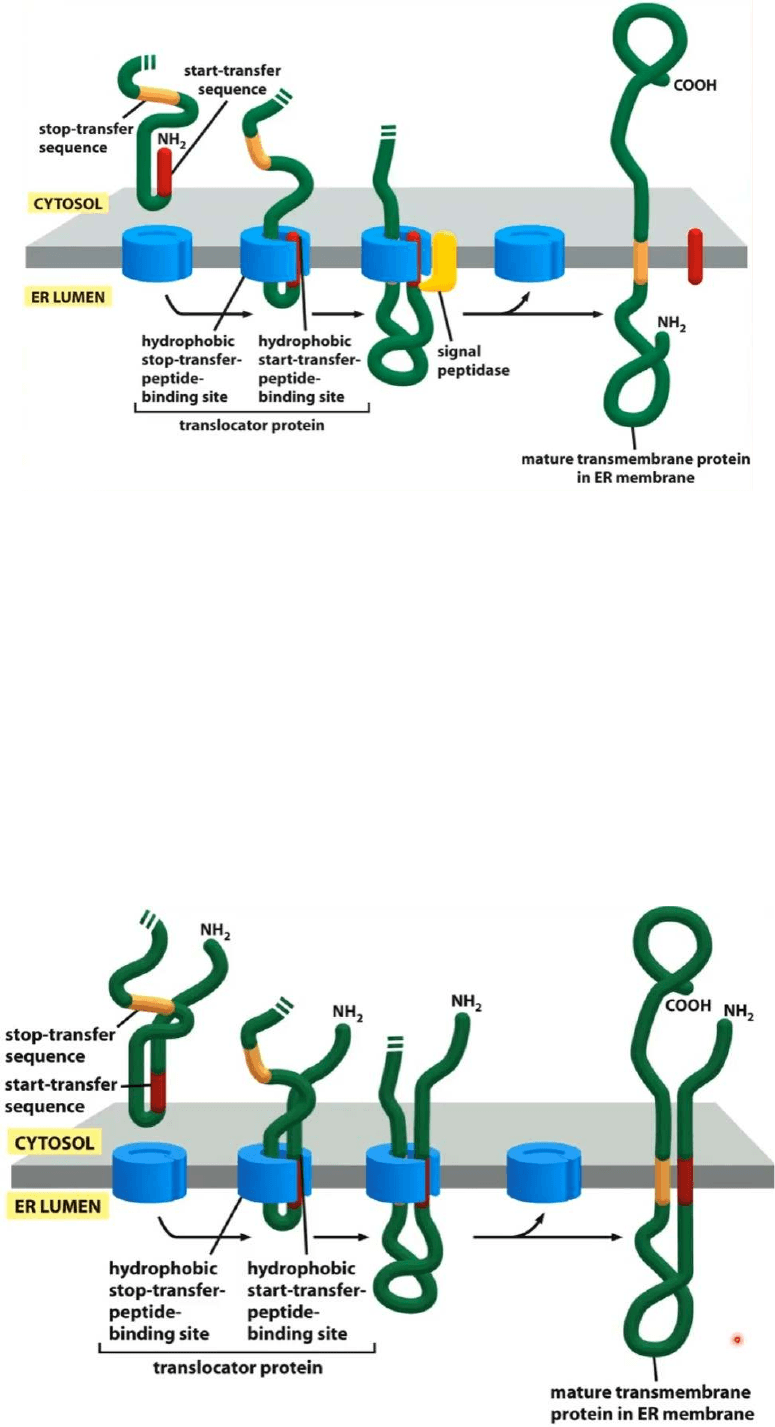
Translocación de una proteína de paso único de membrana en el REG
● Aqui tenemos una proteína que presenta dos señales, en rojo el extremo amino-
terminal tenemos el péptido señal, en el medio del mensaje tenemos esta señal
amarilla
● El mensaje terminará de ser leído en ribosomas adosados en las cisternas del REG
● Cuando la proteína comienza a ser translocada aparece esta señal amarilla que es
una señal de STOP de la translocación, por lo tanto va a ser insertada en la
membrana de la cisterna del REG, esta señal de STOP hace que se detenga esta
señal la translocación y esta proteína no queda como proteína soluble en la luz de
la cisterna del REG como en el anterior sino que ahora queda como proteína
transmembranosa de paso único en la membrana de la cisterna del REG
Translocación de una proteína transmembranosa de doble paso en la membrana de
la cisterna del REG
● La señal roja vuelve a ser el péptido señal, el péptido señal normalmente esta
ubicado en el extremo amino-terminal de una proteína, cuando el péptido señal
esta ubicado en el medio del mensaje además de actuar como péptido señal
también actúa como péptido de STOP que interrumpe la translocación de la
proteína
● Lo que vemos en este caso en lo que vemos es que el péptido señal lo que
determina es la inserción de la proteína en la membrana de la cisterna del REG,
cuando aparece un segundo mensaje vuelve a detener su translocación y ahora
tenemos una proteína que atraviesa dos veces el espesor de la membrana de la
cisterna del REG
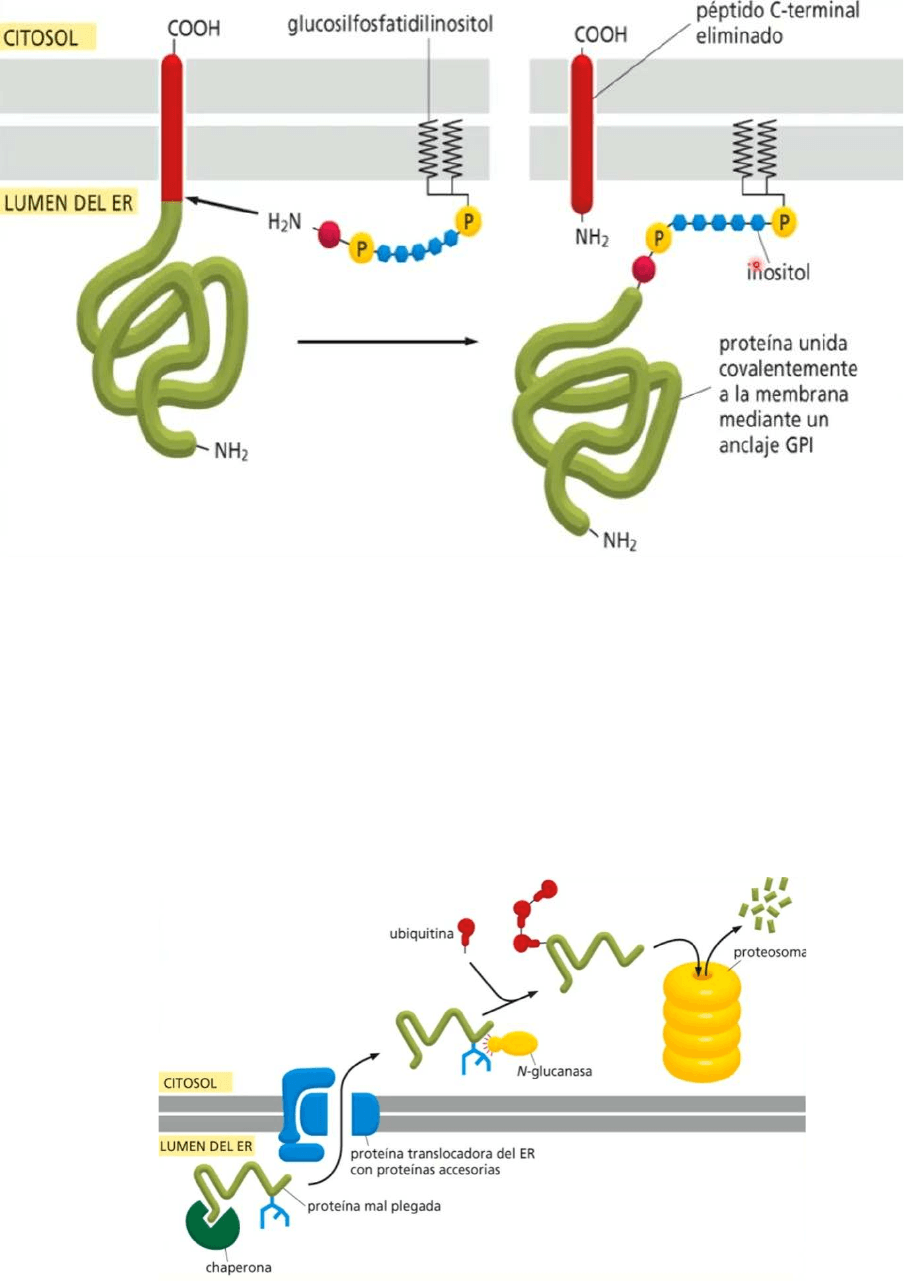
Proteínas con anclaje GPI (Gliceril fosforil inositol)
● Cuando hablamos de síntesis del proteínas en el REG ocurren otras funciones
entre ellas el agregado de este recibo que forma las proteínas con anclaje GPI
(gliceril fosforil inositol)
● Tienen en apariencia una proteína que parece periférica pero en realidad tienen
una unión importante con los lípidos que forman la membrana, en este caso, la
membrana del REG, por lo tanto es un tipo de proteína intrínseca y la forma en que
esta unida es a través de este gliceril fosforil inositol está unida al fosfatidil inositol
de la monocapa interna de la membrana
Control de calidad en el REG
Este documento contiene más páginas...
Descargar Completo
BIOLOGÍA CELULAR III.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.