Tanto el sistema nervioso como el endocrino tienen el mismo obje-
tivo: conservar las condiciones controladas dentro de los límites
que mantienen la vida. El sistema nervioso regula las actividades
corporales respondiendo con rapidez mediante impulsos nervio-
sos; el sistema endocrino responde con la liberación de hormonas.
Las funciones que desempeñan ambos sistemas en el manteni-
miento de la homeostasis se comparan en el Capítulo 18.
El sistema nervioso tiene también a su cargo nuestras percepcio-
nes, conductas y recuerdos, e inicia todos los movimientos volun-
tarios. Dado que el sistema nervioso es bastante complejo, consi-
deraremos los diferentes aspectos de su estructura y función en
varios capítulos. En este nos dedicaremos a la organización del sis-
tema nervioso y a las propiedades de las neuronas (células nervio-
sas) y neuroglia (células que sostienen la actividad de las neuro-
nas). En los capítulos que siguen examinaremos la estructura y la
función de la médula espinal y los nervios espinales (Capítulo 13)
y del encéfalo y los nervios craneales (Capítulo 14). El sistema ner-
vioso autónomo, la parte del sistema nervioso que opera sin con-
trol voluntario, será tratado en el capítulo 15. El Capítulo 16 des-
cribirá los sentidos somáticos (el tacto, la presión, el calor, el frío,
el dolor y otros) y sus vías sensitivas y motoras para entender cómo
se transmiten los impulsos nerviosos hacia la médula espinal y el
encéfalo, o desde la médula espinal y el encéfalo hacia los múscu-
los y las glándulas. Nuestro estudio del sistema nervioso concluye con una exposición acerca de los senti-
dos especiales: olfato, gusto, visión, audición y equilibrio (Capítulo 17).
La neurología estudia el funcionamiento normal y los trastornos del sistema nervioso. Un neurólogo es
un médico que se especializa en el diagnóstico y tratamiento de los trastornos del sistema nervioso.
447
TEJIDO NERVIOSO
12
EL TEJIDO NERVIOSO Y LA HOMEOSTASIS
Las características excitables del tejido nervioso
permiten la generación de impulsos nerviosos (potenciales de acción) que hacen posible la
comunicación y la regulación de la mayoría de los tejidos del cuerpo.
¿Alguna vez pensó de qué modo el sistema nervioso
humano coordina e integra todos los sistemas
corporales tan rápida y eficientemente?
?
ERRN VPHGL FRVRUJ

12.1 GENERALIDADES DEL
SISTEMA NERVIOSO
OBJETIVOS
• Describir la organización del sistema nervioso.
• Enumerar las tres funciones básicas del sistema nervioso.
Organización del sistema nervioso
Con un peso de sólo 2 kg, alrededor del 3% del peso corporal total,
el sistema nervioso es uno de los más pequeños y, sin embargo, más
complejos de los once sistemas y aparatos del cuerpo. Esta red intrin-
cada de miles de millones de neuronas (incluso la neuroglia) está
organizada en dos subdivisiones principales: el sistema nervioso cen-
tral y el sistema nervioso periférico.
Sistema nervioso central
El sistema nervioso central (SNC) está formado por el encéfalo y
la médula espinal (Figura 12.1a). El encéfalo es la parte del SNC que
se localiza en el cráneo y contiene unos 100 000 millones (10
11
) de
neuronas. La médula espinal está conectada con el encéfalo a través
del foramen magno del hueso occipital y está rodeada por los huesos
de la columna vertebral. La médula espinal contiene unos 100 millo-
nes de neuronas. El SNC procesa diversos tipos de información sensi-
tiva aferente. Es también la fuente de los pensamientos, emociones y
recuerdos. La mayoría de los impulsos nerviosos que estimulan a los
músculos para que se contraigan y a las glándulas para que aumenten
su secreción se originan en el SNC.
Sistema nervioso periférico
El sistema nervioso periférico (SNP) está formado por todo el teji-
do nervioso que se encuentra fuera de la médula espinal (Figura
12.1a). Los componentes del SNP incluyen nervios, ganglios, plexos
entéricos y receptores sensoriales. Un nervio es un haz de cientos de
miles de axones (junto con el tejido conectivo y los vasos sanguíneos
asociados) que se encuentran por fuera del encéfalo y la médula espi-
nal. Doce pares de nervios craneales emergen del encéfalo y 31 pares
de nervios espinales emergen de la médula espinal. Cada nervio sigue
un camino definido e inerva una región específica del cuerpo. Los
ganglios (ganglion-, nudo) son pequeñas masas de tejido nervioso
constituidas por los cuerpos celulares de las neuronas, localizados
fuera del encéfalo y de la médula espinal. Los ganglios están íntima-
mente asociados a los nervios craneales y espinales. Los plexos enté-
ricos son redes extensas de neuronas localizadas en las paredes de los
órganos del tubo digestivo. Las neuronas de estos plexos ayudan a
regular el aparato digestivo (véase la Sección 24.3). El término recep-
tor sensorial se refiere a una estructura del sistema nervioso que con-
trola los cambios en el medio ambiente externo o interno. Los ejem-
plos de receptores sensoriales los constituyen los receptores del tacto
en la piel, los fotorreceptores del ojo y los receptores olfatorios en la
nariz.
El SNP puede ser subdividido, a su vez, en sistema nervioso somá-
tico (soma-, cuerpo) (SNS), sistema nervioso autónomo (auto-, pro-
pio; y -nómico, ley) (SNA) y sistema nervioso entérico (enteron-,
intestino) (SNE). El SNS consiste en 1) neuronas sensitivas que trans-
miten la información desde los receptores somáticos de la cabeza, la
pared corporal y los miembros y desde los receptores para los senti-
dos especiales de la visión, audición, gusto y olfato hacia el SNC, y
2) neuronas motoras que conducen impulsos desde el SNC hacia los
músculos esqueléticos solamente. Como estas respuestas motoras
pueden ser controladas conscientemente, la acción de esta región del
SNP es voluntaria.
El SNA está formado por 1) neuronas sensitivas que transportan
información proveniente de los receptores sensitivos autonómicos
localizados principalmente en órganos viscerales como el estómago y
los pulmones hacia el SNC, y 2) neuronas motoras que conducen
impulsos nerviosos desde el SNC hacia el músculo liso, el músculo
cardíaco y las glándulas. Dado que estas respuestas motoras no están
normalmente bajo control consciente, la acción del SNA es involunta-
ria. La zona motora del SNA tiene 2 ramas: la división simpática y
la división parasimpática. Con pocas excepciones, los efectores
están inervados por ambas divisiones, y habitualmente éstas ejercen
acciones opuestas. Por ejemplo, las neuronas simpáticas aumentan la
frecuencia cardíaca, y las neuronas parasimpáticas la reducen.
Generalmente, la división simpática ayuda a la ejecución de las accio-
nes de emergencia, las llamadas respuestas de “lucha y huida”, y la
división parasimpática tiene a su cargo las actividades de “reposo y
digestión”.
El funcionamiento del SNE, el “cerebro visceral”, es involuntario.
Alguna vez considerado parte del SNA, el SNE contiene más de 100
millones de neuronas situadas en los plexos entéricos, que se distribu-
yen a lo largo de la mayor parte del tubo digestivo. Muchas de las neu-
ronas localizadas en los plexos entéricos funcionan, hasta cierto
punto, en forma independiente del SNA y del SNC, aunque también
se comunican con el SNC por medio de neuronas simpáticas y para-
simpáticas. Las neuronas sensitivas del SNE monitorizan los cambios
químicos que se producen en el tubo digestivo, y también la distensión
de sus paredes. Las neuronas motoras entéricas coordinan la contrac-
ción del músculo liso del tubo digestivo, que estimula la progresión
del alimento a lo largo de él, regulan las secreciones de los órganos
digestivos, como el ácido gástrico, y la actividad de las células endo-
crinas del aparato digestivo, que secretan hormonas.
Funciones del sistema nervioso
El sistema nervioso lleva a cabo un complejo conjunto de tareas.
Permite percibir diferentes olores, hablar y recordar hechos pasados,
también proporciona señales que controlan los movimientos del cuer-
po y regulan el funcionamiento de los órganos internos. Estas activi-
dades diversas pueden ser agrupadas en 3 funciones básicas: sensitiva
(aferente), integradora (de proceso) y motora (eferente).
• Función sensitiva. Los receptores sensitivos detectan los estímu-
los internos, como el aumento de la tensión arterial, y los externos,
como el estímulo que produce una gota de lluvia cuando cae sobre
el brazo. Esta información sensitiva es transportada luego hacia el
encéfalo y la médula espinal a través de los nervios craneales y
espinales.
• Función integradora. El sistema nervioso procesa la información
sensitiva analizando y tomando decisiones para efectuar las res-
puestas adecuadas, actividad conocida como integración.
• Función motora. Una vez que la información sensorial ha sido
integrada, el sistema nervioso puede generar una respuesta moto-
ra adecuada activando efectores (músculos y glándulas) a través
de los nervios craneales y espinales. La estimulación de los efecto-
res produce la contracción de un músculo o estimula una glándula
para aumentar su secreción.
Las tres funciones básicas del sistema nervioso se ponen en eviden-
cia, por ejemplo, cuando usted responde su teléfono celular después
de escuchar que suena. El sonido del teléfono sonando estimula los
receptores sensoriales de sus oídos (función sensitiva). Esta informa-
448 CAPÍTULO 12 • TEJIDO NERVIOSO
ERRN VPHGL FRVRUJ
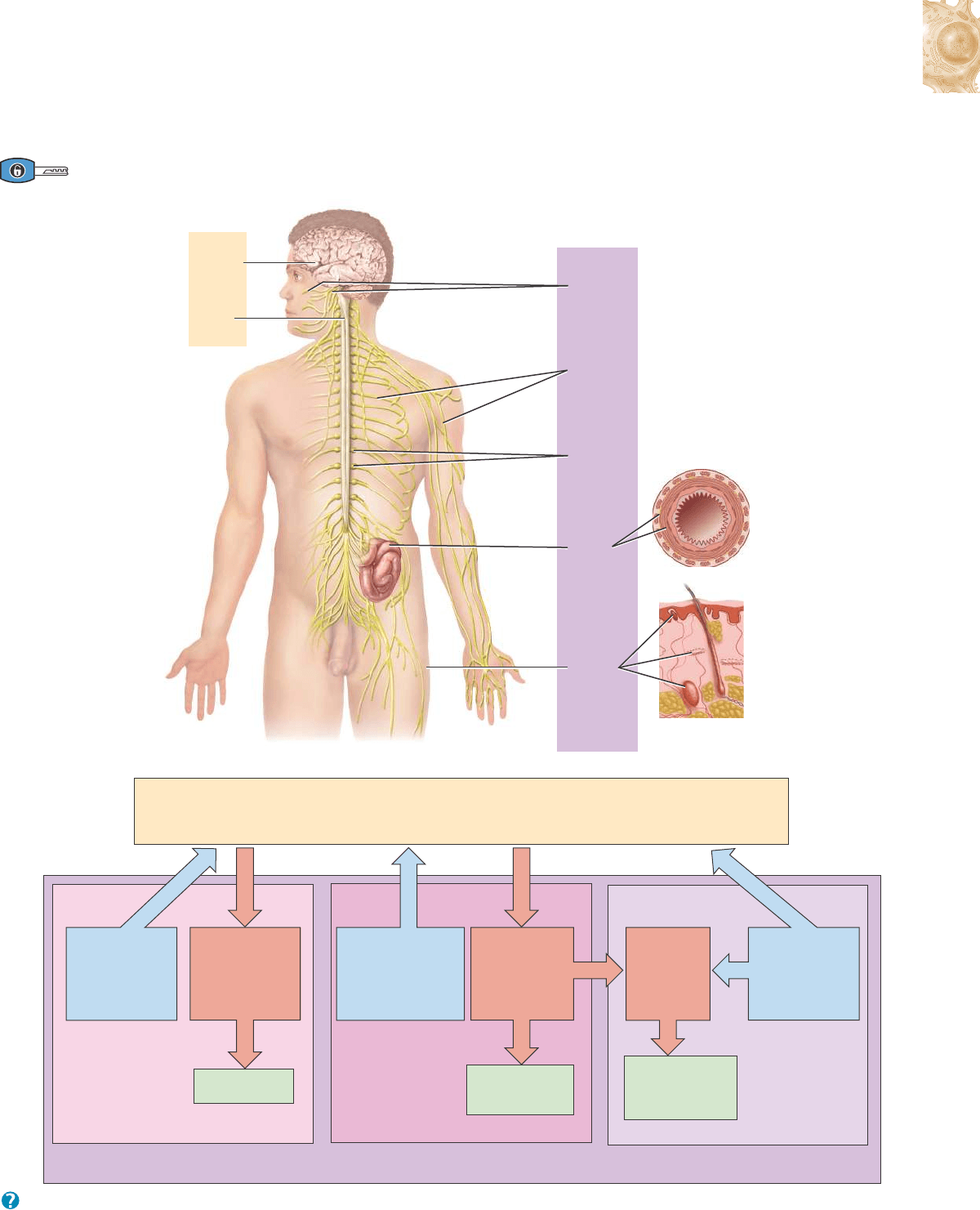
Figura 12.1 Organización del sistema nervioso. a) Subdivisiones del sistema nervioso. b) Diagrama de organización del sistema
nervioso; los recuadros celestes representan componentes sensitivos del sistema nervioso periférico, los rojos, componentes
motores del SNP y los verdes, efectores (músculos y glándulas).
Los dos subsistemas principales del sistema nervioso son: 1) el sistema nervioso central (SNC), constituido por el encéfalo y la médula espinal
y 2) el sistema nervioso periférico (SNP), formado por todo el tejido nervioso que se halla por fuera del SNC.
Nervios
craneales
Nervios
espinales
Ganglios
Recep-
tores
sensitivos
en la piel
Plexos
entéricos
en el
intestino
delgado
Encéfalo
espinal
Médula
SNP:
SNC:
(a)
Receptores
sensitivos
somáticos y es-
peciales, y neu-
ronas sensitivas
somáticas
Neuronas
motoras somáticas
(voluntarias)
Sistema Nervioso Somático (SNS)
Receptores sensitivos
autónomos y neuronas
sensitivas autónomas
Neuronas motoras
autónomas
(involuntarias):
divisiones
simpática y
parasimpática
Músculo liso,
músculo cardíaco
y glándulas
Neuronas
motoras
entéricas
(involuntarias):
en plexos
entéricos
Receptores sensiti-
vos entéricos y
neuronas sensitivas
entéricas en el tubo
digestivo y en los
plexos entéricos
Músculo liso,
glándulas y células
endócrinas del tubo
digestivo
Sistema Nervioso Autónomo (SNA) Sistema Nervioso Entérico (SNE)
SISTEMA NERVIOSO PERIFÉRICO (SNP): todo el tejido nervioso fuera del SNC
SISTEMA NERVIOSO CENTRAL (SNC): encéfalo y médula espinal
(b)
Músculo
esquelético
¿Cuáles son algunas de las funciones del SNC?
ERRN VPHGL FRVRUJ

ción auditiva es transmitida posteriormente a su encéfalo donde es
procesada y se toma la decisión de responder el teléfono (función inte-
gradora). El encéfalo estimula, entonces, la contracción de músculos
específicos que permiten tomar el teléfono y presionar el botón apro-
piado para responder (función motora).
PREGUNTAS DE REVISIÓN
1. ¿Cuál es el propósito de un receptor sensitivo?
2. ¿Cuáles son los componentes y las funciones del SNS, del
SNA y del SNE?
3. ¿Qué subdivisión del SNP controla las acciones volunta-
rias? ¿Cuál controla las acciones involuntarias?
4. Explique el concepto de integración y proporcione un
ejemplo.
12.2 HISTOLOGÍA DEL TEJIDO
NERVIOSO
OBJETIVOS
• Comparar las características histológicas y las funciones de
las neuronas y de la neuroglia.
• Distinguir entre la sustancia gris y la sustancia blanca.
El tejido nervioso tiene dos tipos de células: las neuronas y la neu-
roglia. Estas células se combinan de distintas formas en diferentes
regiones del sistema nervioso. Además de formar las redes de proce-
samiento complejo dentro del encéfalo y la médula espinal, las neuro-
nas también conectan todas las regiones del cuerpo con el encéfalo y
la médula espinal. Como células altamente especializadas capaces de
alcanzar grandes longitudes y de realizar conexiones extremadamente
intrincadas con otras células, las neuronas realizan la mayoría de las
funciones propias del sistema nervioso, como la sensación, el pensa-
miento, el recuerdo, el control de la actividad muscular y la regulación
de las secreciones glandulares. Como resultado de su especialización,
estas células han perdido la capacidad para realizar divisiones mitóti-
cas. Las células de la neuroglia son más pequeñas pero superan en
número a las neuronas, tal vez hasta 25 veces. La neuroglia sostiene,
nutre y protege a las neuronas; además, mantiene el líquido intersticial
que las baña. Al contrario de las neuronas, las células de la neuroglia
se siguen dividiendo durante toda la vida. Las neuronas y la neuro-
glia difieren estructuralmente según su localización en el sistema ner-
vioso central o en el sistema nervioso periférico. Estas diferencias de
estructura se deben a las diferentes funciones del sistema nervioso
central y del sistema nervioso periférico.
Neuronas
Como las células musculares, las neuronas (células nerviosas) tienen
excitabilidad eléctrica: la capacidad para responder a un estímulo y
convertirlo en un potencial de acción. Un estímulo es cualquier cambio
en el medio que sea lo suficientemente importante para iniciar un poten-
cial de acción. Un potencial de acción (impulso nervioso) es una señal
eléctrica que se propaga (viaja) a lo largo de la superficie de la membra-
na plasmática de una neurona. Se inicia y se desplaza por el movimien-
to de iones (como los de sodio y potasio) entre el líquido intersticial y el
interior de la neurona a través de canales iónicos específicos en su mem-
brana plasmática. Una vez que ha comenzado, un impulso nervioso se
desplaza rápidamente y con una amplitud constante.
Algunas neuronas son pequeñas y propagan los impulsos nerviosos a
corta distancia (menos de 1 mm) dentro del SNC. Otras se encuentran
entre las células más largas del cuerpo. Las neuronas motoras que nos
permiten mover los dedos de los pies, por ejemplo, se extienden desde
la región lumbar de la médula espinal (justo por encima del nivel de la
cintura) hasta los músculos de los pies. Algunas neuronas sensitivas son
incluso más largas. Aquellas que hacen posible percibir una pluma
tocando los dedos del pie se extienden desde el pie hasta la porción más
baja del encéfalo. Los impulsos nerviosos recorren estas grandes distan-
cias a velocidades que van desde 0,5 a 130 metros por segundo.
Partes de una neurona
Casi todas las neuronas tienen 3 partes constitutivas: 1) un cuerpo
celular, 2) dendritas y 3) un axón (Figura 12.2). El cuerpo celular,
también conocido como pericarion o soma, contiene el núcleo rodea-
do por el citoplasma, en el que se hallan los típicos orgánulos celula-
res como los lisosomas, las mitocondrias y el complejo de Golgi. Los
cuerpos celulares de las neuronas también contienen ribosomas libres
y condensaciones del retículo endoplasmático rugoso, denominadas
cuerpos de Nissl. Los ribosomas son los sitios donde tiene lugar la
síntesis de las proteínas. Las proteínas recién sintetizadas que se for-
man en los cuerpos de Nissl sirven para remplazar diversos compo-
nentes celulares utilizados en el crecimiento de las neuronas y en la
regeneración de los axones dañados en el SNP. En el citoesqueleto se
encuentran las neurofibrillas, compuestas por haces de filamentos
intermedios que le dan forma y soporte a la célula, y los microtúbu-
los, que participan en el movimiento de materiales entre el cuerpo
celular y el axón. Las neuronas que envejecen también contienen lipo-
fuscina, un pigmento que aparece como acumulaciones de gránulos
amarillentos en el citoplasma. La lipofuscina es un producto de los
lisosomas neuronales que se acumula a medida que la neurona enve-
jece, pero que no parece generar daño a la célula.
Fibra nerviosa es un término general con el que se designa cual-
quier proyección que emerge del cuerpo de una neurona. La mayoría
de las neuronas tiene dos tipos de prolongaciones: las dendritas (múl-
tiples) y el axón (único). Las dendritas (déndron-, árbol) conforman
la porción receptora o de entrada de una neurona. Las membranas
plasmáticas de las dendritas (y los cuerpos celulares) contienen nume-
rosos sitios receptores para la fijación de mensajeros químicos prove-
nientes de otras células. Las dendritas habitualmente son cortas, agu-
zadas y presentan múltiples ramificaciones. En muchas neuronas, las
dendritas adoptan una disposición arborescente de ramificaciones que
se extienden desde el cuerpo celular. Su citoplasma contiene cuerpos
de Nissl, mitocondrias y otros orgánulos.
El axón (áxoon-, eje) de una neurona propaga los impulsos nervio-
sos hacia otra neurona, una fibra muscular o una célula glandular. El
axón es una proyección cilíndrica larga y fina que generalmente se
une con el cuerpo celular en una elevación cónica denominada cono
axónico. El sector del axón más cercano al cono axónico es el seg-
mento inicial. En casi todas las neuronas, los impulsos nerviosos se
originan en la unión entre el cono axónico y el segmento inicial, área
que se denomina zona gatillo, desde donde estos impulsos se dirigen
a lo largo del axón hasta su destino final. Un axón contiene mitocon-
drias, microtúbulos y neurofibrillas. Como no presenta retículo endo-
plasmático rugoso, no puede realizarse la síntesis de proteínas. El
citoplasma de un axón, denominado axoplasma, está rodeado por una
membrana plasmática conocida como axolema (de axón; y -lemma,
vaina o corteza). A lo largo del axón puede haber ramificaciones,
denominadas colaterales axónicas, que forman un ángulo recto con
el axón del que originalmente salieron. El axón y sus colaterales ter-
minan en muchas prolongaciones delgadas que se denominan axón
terminal, o telodendrón (télos-, fin, término; y -déndron, árbol).
450 CAPÍTULO 12 • TEJIDO NERVIOSO
ERRN VPHGL FRVRUJ
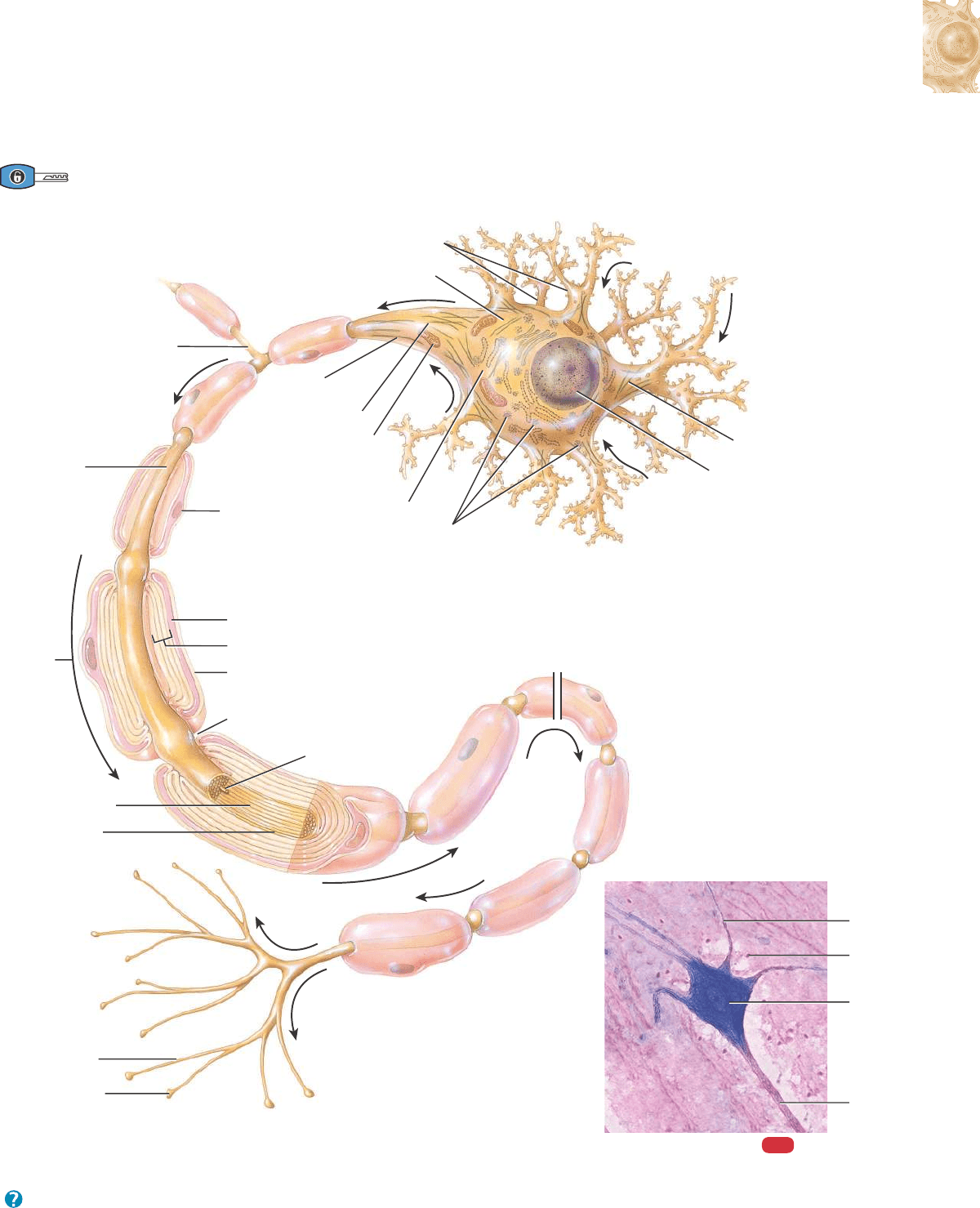
12.2 HISTOLOGÍA DEL TEJIDO NERVIOSO 451
¿Qué roles desempeñan las dendritas, el cuerpo celular y el axón en la comunicación de señales?
Figura 12.2 Estructura de una neurona multipolar (una neurona con un gran cuerpo celular, varias dendritas cortas y un solo
axón largo). Las flechas indican la dirección del flujo de información: dendritas n cuerpo celular n axón n axones
terminales.
Las partes básicas de una neurona son las dendritas, el cuerpo celular y el axón.
AXÓN
Segmento inicial
Cono axónico
Mitocondria
Citoplasma
Neurofibrilla
Núcleo
Cuerpos de Nissl
Núcleo
de la
célula de
Schwann
Célula de Schwann:
Nodo de Ranvier
Neurofibrilla
Axón terminal
Bulbo terminal
sináptico
Axón:
Citoplasma
Vaina de mielina
Axoplasma
Impulso
nervioso
Axolema
Neurolema
DENDRITAS
CUERPO CELULAR
Colateral axónico
(a) Partes de una neurona
Célula
neuroglial
Cuerpo
celular
Axón
Dendrita
400x
MO
(b) Neurona motora
ERRN VPHGL FRVRUJ
El lugar de la comunicación entre dos neuronas o entre una neuro-
na y una célula efectora es la sinapsis (synapsis-, unión, conexión).
Los extremos de algunos terminales axónicos se ensanchan para for-
mar estructuras que por su forma se denominan bulbos sinápticos
terminales; otros muestran una cadena de porciones ensanchadas que
reciben el nombre de varicosidades. Tanto los bulbos sinápticos ter-
minales como las varicosidades contienen gran número de sacos rode-
ados de membrana, las vesículas sinápticas, que almacenan una sus-
tancia química denominada neurotransmisor. Un neurotransmisor es
una molécula liberada desde una vesícula sináptica que excita o inhi-
be otra neurona, una fibra muscular o una célula glandular. Muchas
neuronas presentan dos o incluso tres tipos de neurotransmisores,
cada uno con diferentes efectos sobre las estructuras postsinápticas.
Como algunas de las sustancias que se sintetizan o reciclan en el
cuerpo neuronal son necesarias en el axón o en los axones termina-
les, existen dos tipos de sistemas de transporte que conducen sustan-
cias desde el cuerpo o soma de la célula hasta los axones terminales y
desde éstos hacia el cuerpo celular. El sistema más lento, que traslada
estas sustancias aproximadamente a 1-5 mm por día, se denomina
transporte axónico lento. Este sistema sólo transporta el axoplasma
en una dirección: desde el cuerpo celular hacia los axones terminales.
El transporte axónico lento abastece de axoplasma nuevo a los axones
que están en desarrollo o en regeneración y lo repone en los axones en
crecimiento o en los ya maduros.
El transporte axónico rápido, capaz de conducir sustancias a una
velocidad de 200 a 400 mm por día, utiliza proteínas que funcionan
como “motores” para movilizarlas a lo largo de la superficie de los
microtúbulos del citoesqueleto neuronal. El transporte axónico rápido
conduce materiales en ambas direcciones: desde el cuerpo celular y
hacia él. El transporte axónico rápido que ocurre en dirección anteró-
grada (hacia adelante) conduce orgánulos y vesículas sinápticas
desde el cuerpo celular hacia los axones terminales. Aquel que se pro-
duce en dirección retrógrada (hacia atrás) conduce las vesículas de la
membrana y otros materiales celulares desde los axones terminales
hacia el cuerpo celular, para que sean degradados o reciclados. Las
sustancias que entran en la neurona en los axones terminales también
son conducidas hacia el cuerpo celular, por medio de transporte axó-
nico rápido. Estas sustancias incluyen productos químicos tróficos
como el factor de crecimiento nervioso y agentes nocivos como la
toxina tetánica y los virus que producen rabia, herpes simple y polio-
mielitis.
Diversidad estructural de las neuronas
Las neuronas presentan una gran diversidad de formas y de tama-
ños. Por ejemplo, sus cuerpos celulares tienen un diámetro que va
desde los 5 micrómetros (µm) (ligeramente menor que un eritrocito)
hasta los 135 µm (lo suficientemente grande como para ser percibido
apenas por el ojo humano). El patrón de ramificación de las dendritas
es variado y distintivo de cada clase de neurona en los diferentes sec-
tores del sistema nervioso. Unas pocas neuronas pequeñas carecen de
axón, y muchas otras tienen axones muy cortos. Como ya hemos
visto, la longitud de los axones más largos es casi igual a la estatura
de una persona, ya que abarca una superficie que va desde la parte más
baja del encéfalo hasta los dedos de los pies.
Clasificación de las neuronas
Para clasificar las diferentes neuronas que se encuentran en el cuer-
po, se utilizan tanto sus características funcionales como las estructu-
rales.
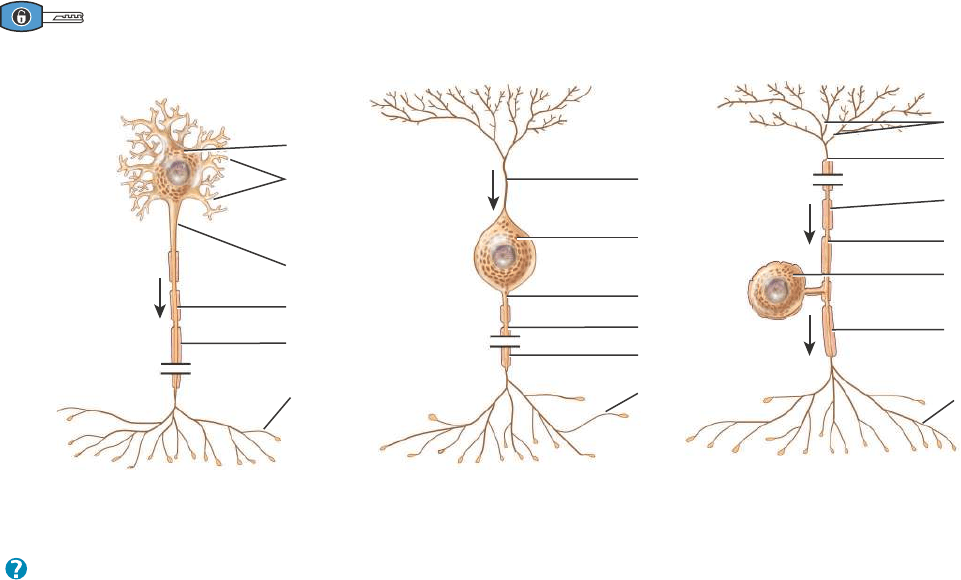
CLASIFICACIÓN ESTRUCTURAL. Las neuronas se clasifican en cuanto a
su estructura, según el número de prolongaciones que afloran de su
cuerpo celular (Figura 12.3).
452 CAPÍTULO 12 • TEJIDO NERVIOSO
Cuerpo celular
Dentritas
Zona gatillo
Axón
Terminación
axónica
(a) Neurona multipolar
Cuerpo celular
Dentritas
Zona gatillo
Axón
Vaina
de mielina
Vaina de mielina
Terminación
axónica
(b) Neurona bipolar
cuerpo celular
Vaina de mielina
Zona gatillo
Prolongación
periférica
Prolongación
central
Terminación
axónica
(c) Neurona unipolar
Dentritas
¿Qué tipo de neurona que se observa en esta figura es el más abundante en el SNC?
Figura 12.3 Clasificación estructural de las neuronas. Las líneas de corte en las figuras indican que los axones son más largos que lo que
se muestra.
Una neurona multipolar tiene muchas prolongaciones que se extienden desde el cuerpo celular; una neurona bipolar tiene dos prolongaciones y
una neurona unipolar tiene sólo una.
ERRN VPHGL FRVRUJ
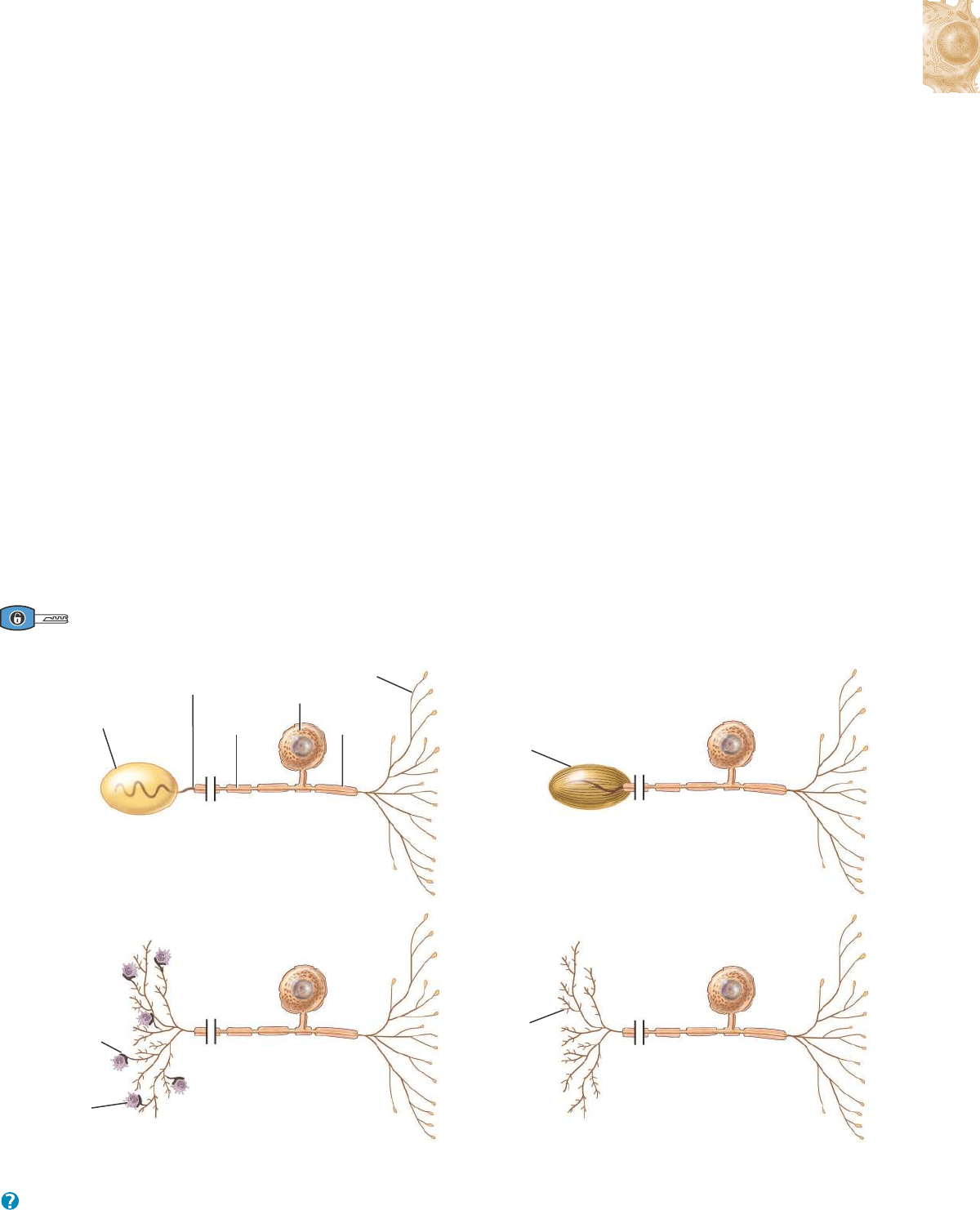
1. Las neuronas multipolares tienen generalmente varias dendritas y
un axón (Figura 12.3a). La mayoría de las neuronas situadas en el
encéfalo y en la médula espinal son de este tipo, como todas las
neuronas motoras (tratadas más adelante).
2. Las neuronas bipolares tienen una dendrita principal y un axón
(Figura 12.3b). Se encuentran en la retina del ojo, en el oído inter-
no y en el área olfatoria del encéfalo.
3. Las neuronas unipolares tienen dendritas y un axón que se fusio-
nan para formar una prolongación continua que emerge del cuerpo
celular (Figura 12.3c). Estas neuronas se denominan más adecua-
damente neuronas seudounipolares porque comienzan en el
embrión como neuronas bipolares. Durante el desarrollo, las den-
dritas y el axón se fusionan en una prolongación única. Las dendri-
tas de la mayoría de las neuronas unipolares funcionan como recep-
tores sensoriales que detectan un estímulo sensitivo como tacto,
presión, dolor o estímulos térmicos (véase la Figura 12.10). La
zona gatillo de los impulsos nerviosos en una célula unipolar se
halla en la unión entre las dendritas y el axón (Figura 12.3c). Los
impulsos se dirigen luego hacia los bulbos terminales sinápticos.
Los cuerpos celulares de la mayoría de las neuronas unipolares
están localizados en los ganglios de los nervios craneales y espina-
les. En la Figura 12.4, se muestran varios ejemplos de receptores
sensoriales que son dendritas de neuronas unipolares.
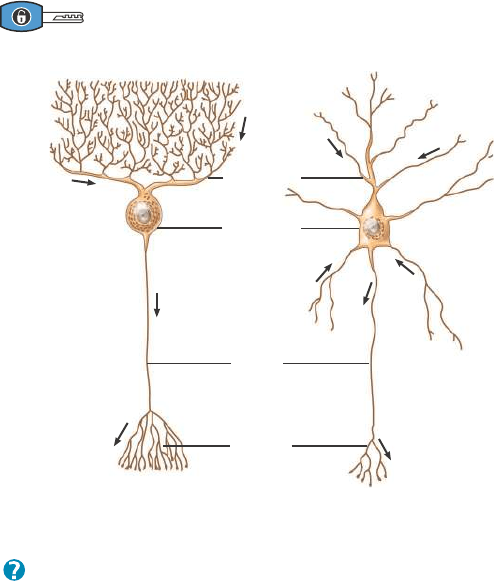
Además del esquema de clasificación que acabamos de describir,
algunas neuronas reciben su nombre en honor al histólogo que las des-
cribió originalmente o por su forma o aspecto; algunos ejemplos de
ello son las células de Purkinje en el cerebelo y las células pirami-
dales, de la corteza cerebral, que poseen cuerpos celulares con forma
de pirámide (Figura 12.5).
12.2 HISTOLOGÍA DEL TEJIDO NERVIOSO 453
Figura 12.4 Ejemplos de receptores sensoriales que son dendritas de neuronas unipolares. Las interrupciones indican que los axones
son más largos que lo que se muestra. a) Un corpúsculo de Meissner es un receptor táctil que consiste en una masa de
dendritas encerradas por una cápsula de tejido conectivo. b) Un disco de Merkel es un receptor táctil compuesto por
terminaciones nerviosas libres (dendritas desnudas) que hacen contacto con células de Merkel del estrato basal de la piel.
c) Un corpúsculo de Pacini es un receptor de presión compuesto por una cápsula con múltiples capas de tejido conectivo que
encierra una dendrita. d) Un nociceptor es un receptor para el dolor que consiste en terminaciones nerviosas libres (dendritas
desnudas). Los termorreceptores (que detectan sensaciones térmicas), los receptores del prurito y los receptores del
cosquilleo se asemejan a los nociceptores en que son neuronas unipolares con terminaciones nerviosas libres que sirven como
receptores sensitivos.
El tipo de receptor sensorial hallado en las dendritas de una neurona unipolar determina el tipo de estímulo sensitivo que puede detectar esa
neurona.
Cuerpo
celular
Axón
Zona
gatillo
Corpúsculo
de Meissner
(corpúsculo
del tacto)
Célula
de Merkel
Disco de
Merkel
(táctil)
Corpúsculo
de Pacini
(laminado)
Nociceptor
Vaina de
mielina
Axón terminal
(a)
(b)
(c)
(d)
¿En que parte del cuerpo se localizan la mayoría de las neuronas unipolares?
ERRN VPHGL FRVRUJ
CLASIFICACIÓN FUNCIONAL. En cuanto a su función, las neuronas se
clasifican según la dirección en la que se transmite el impulso nervio-
so (potencial de acción) con respecto al SNC.
1. Las neuronas sensitivas o aferentes (af-, hacia; y -ferrente, trans-
portado) contienen receptores sensitivos en sus extremos distales
(dendritas) (véase la Figura 12.10) o se localizan inmediatamente
después de los receptores sensitivos, que son células separadas.
Una vez que un estímulo adecuado activa un receptor sensitivo, la
neurona sensitiva produce un potencial de acción en su axón y éste
es transmitido en el SNC, a través de los nervios craneales o espi-
nales. La mayoría de las neuronas sensitivas tienen una estructura
unipolar.
2. Las neuronas motoras o eferentes (ef-, lejos de) transmiten los
potenciales de acción lejos del SNC hacia los efectores (músculos
y glándulas) en la periferia (SNP), a través de los nervios craneales
y espinales (véase la Figura 12.10). Las neuronas motoras tienen
una estructura multipolar.
3. Las interneuronas o neuronas de asociación se localizan funda-
mentalmente dentro del SNC, entre las neuronas sensitivas y moto-
ras (véase la Figura 12.10). Las interneuronas integran (procesan)
la información sensitiva entrante proveniente de las neuronas sen-
sitivas y luego producen una respuesta motora, al activar las neuro-
nas motoras adecuadas. La mayoría de las interneuronas tienen una
estructura multipolar.
Neuroglia
La neuroglia (néuron-, nervio; y -glía, gluten) o glía representa
alrededor de la mitad del volumen del SNC. Su nombre deriva de la
idea que tenían los primeros histólogos, de que era el “adhesivo” que
mantenía unido el sistema nervioso. Ahora sabemos que la neuroglia
no es un mero espectador pasivo, sino que participa activamente en las
actividades del tejido nervioso. En general, las células gliales tienen
menor tamaño que las neuronas, pero son entre 5 y 25 veces más
numerosas. A diferencia de las neuronas, las células gliales no gene-
ran ni propagan potenciales de acción, y se pueden multiplicar y divi-
dir en el sistema nervioso ya maduro. En caso de lesión o enfermedad,
la neuroglia se multiplica para rellenar los espacios que anteriormen-
te ocupaban las neuronas. Los tumores encefálicos derivados de la
glía, denominados gliomas, tienden a ser muy malignos y a crecer con
gran rapidez. De los seis tipos de células gliales, cuatro (los astroci-
tos, los oligodendrocitos, la microglia y las células ependimarias) se
encuentran sólo en el SNC. Los dos tipos restantes (las células de
Schwann y las células satélite) están presentes en el SNP.
Neuroglia del SNC
Las células de la neuroglia del SNC pueden clasificarse según el
tamaño, las prolongaciones citoplasmáticas y la organización intrace-
lular, en cuatro tipos: astrocitos, oligodendrocitos, microglia y células
ependimarias (Figura 12.6).
ASTROCITOS (astro-, estrella; y -cytos, cavidad). Estas células con
forma de estrella tienen muchas prolongaciones celulares y son las
más largas y numerosas de la neuroglia. Existen dos tipos de astroci-
tos. Los astrocitos protoplasmáticos tienen gran cantidad de prolon-
gaciones cortas y ramificadas y se encuentran en la sustancia gris (tra-
tada más adelante). Los astrocitos fibrosos tienen gran cantidad de lar-
gas prolongaciones no ramificadas y se localizan principalmente en la
sustancia blanca (también tratada más adelante). Esas prolongaciones
hacen contacto con capilares sanguíneos, con neuronas y con la pia-
madre (una delgada membrana que se dispone alrededor del encéfalo
y la médula espinal).
Las funciones de los astrocitos son las siguientes: (1) Contienen
microfilamentos que les dan una resistencia considerable y les permi-
ten sostener las neuronas. (2) Las proyecciones de los astrocitos que
envuelven los capilares sanguíneos aíslan las neuronas del SNC de
diferentes sustancias potencialmente nocivas de la sangre, mediante la
secreción de compuestos químicos que mantienen las características
exclusivas de permeabilidad que tienen las células endoteliales de los
capilares. En efecto, las células endoteliales forman la barrera hema-
toencefálica, que restringe el paso de sustancias entre la sangre y el
líquido intersticial del SNC. Los detalles de la barrera hematoencefá-
lica se tratan en el Capítulo 14. (3) En el embrión, los astrocitos secre-
tan sustancias químicas que aparentemente regulan el crecimiento, la
migración y la interconexión entre las neuronas cerebrales. (4) Los
astrocitos contribuyen a mantener las condiciones químicas propicias
para la generación de impulsos nerviosos. Por ejemplo, regulan la
concentración de iones tan importantes como el K
+
, capturan los neu-
rotransmisores excedentes y sirven como conducto para el paso de
nutrientes y otras sustancias entre los capilares sanguíneos y las neu-
ronas. (5) Los astrocitos también desempeñan una función en el
aprendizaje y en la memoria, por medio de la influencia que ejercen
sobre la formación de las sinapsis (véase la Sección 16.5).
OLIGODENDROCITOS (oligo-, poco; y -dendrón, árbol). Estas células
se asemejan a los astrocitos, pero son más pequeñas y contienen
menor cantidad de prolongaciones. Las prolongaciones de los oligo-
dendrocitos son responsables de la formación y mantenimiento de la
vaina de mielina que se ubica alrededor de los axones del SNC. Como
usted verá más adelante, la vaina de mielina es una cubierta con múl-
tiples capas, formada por lípidos y proteínas, que envuelve a ciertos
454 CAPÍTULO 12 • TEJIDO NERVIOSO
Cuerpo
celular
Dendritas
Axón
(a) Célula de Purkinje (b) Célula piramidal
Axón
terminal
Figura 12.5 Dos ejemplos de neuronas del SNC. Las flechas
indican la dirección del flujo de información.
El patrón de ramificación de las dendritas suele ser distintivo
de cada tipo particular de neurona.
¿Cómo obtuvo su nombre la célula piramidal?
ERRN VPHGL FRVRUJ
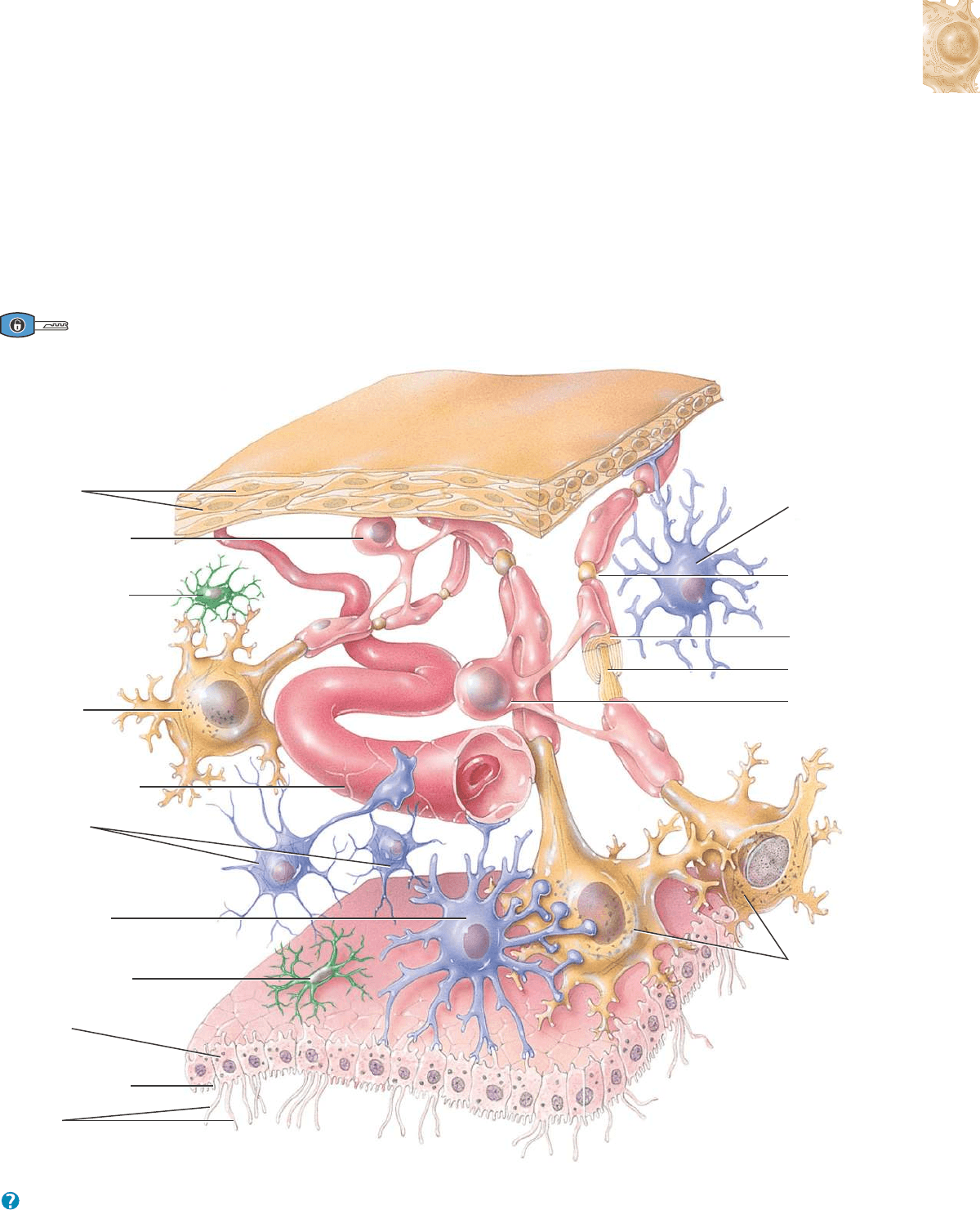
axones, los aísla y aumenta la velocidad de conducción de los impul-
sos nerviosos. Se dice que tales axones están mielinizados.
MICROGLIA (micro-, pequeño). Estas células de la neuroglia son
pequeñas y tienen delgadas prolongaciones que emiten numerosas
proyecciones con forma de espinas. La microglia cumple funciones
fagocíticas. Como los macrófagos de los tejidos, eliminan los detritos
celulares que se forman durante el desarrollo normal del sistema ner-
vioso y fagocitan microorganismos y tejido nervioso dañado.
CÉLULAS EPENDIMARIAS. Las células ependimarias (epen-, encima;
y -dymo, vestidura) tienen forma cuboide o cilíndrica y están distribui-
das en una monocapa con microvellosidades y cilios. Estas células
tapizan los ventrículos cerebrales y el conducto central de la médula
espinal (espacios que contienen líquido cefalorraquídeo, que protege
y nutre al encéfalo y la médula). En cuanto a su función, las células
ependimarias producen, posiblemente monitorizan, y contribuyen a la
circulación del líquido cefalorraquídeo. También forman parte de la
barrera hematoencefálica, que será tratada en el Capítulo 14.
12.2 HISTOLOGÍA DEL TEJIDO NERVIOSO 455
Figura 12.6 Neuroglia del sistema nervioso central (SNC).
Las células gliales del SNC se diferencian de acuerdo con su tamaño, sus prolongaciones citoplasmáticas y su organización intracelular.
¿Qué células de la neuroglia del SNC funcionan como fagocitos?
Células
de la piamadre
Astrocitos
fibrosos
Oligodendrocitos
Célula microglial
Neurona
Capilar sanguíneo
Astrocitos
protoplasmáticos
Célula microglial
Célula
ependimal
Microvellosidad
Cilio
Ventrículo
Nueronas
Oligodendrocito
Nodo de Ranvier
Astrocito
protoplasmático
Vaina de mielina
Axón
ERRN VPHGL FRVRUJ
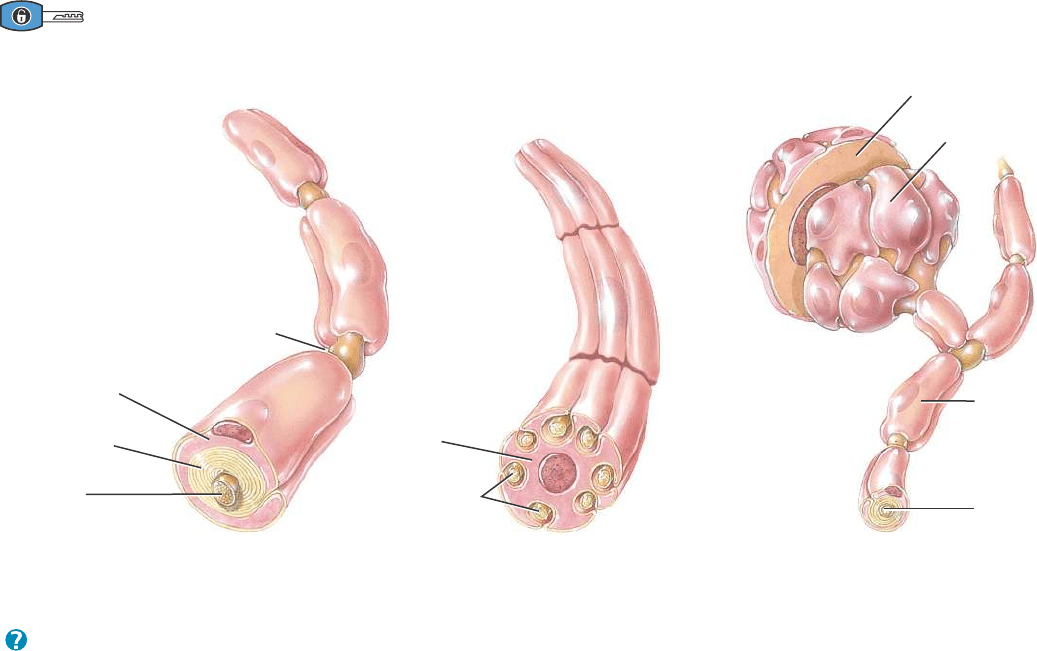
Neuroglia del SNP
La neuroglia del SNP rodea por completo los axones y los cuerpos
celulares. Los dos tipos de células gliales que se hallan en el SNP son
las células de Schwann y las células satélite (Figura 12.7).
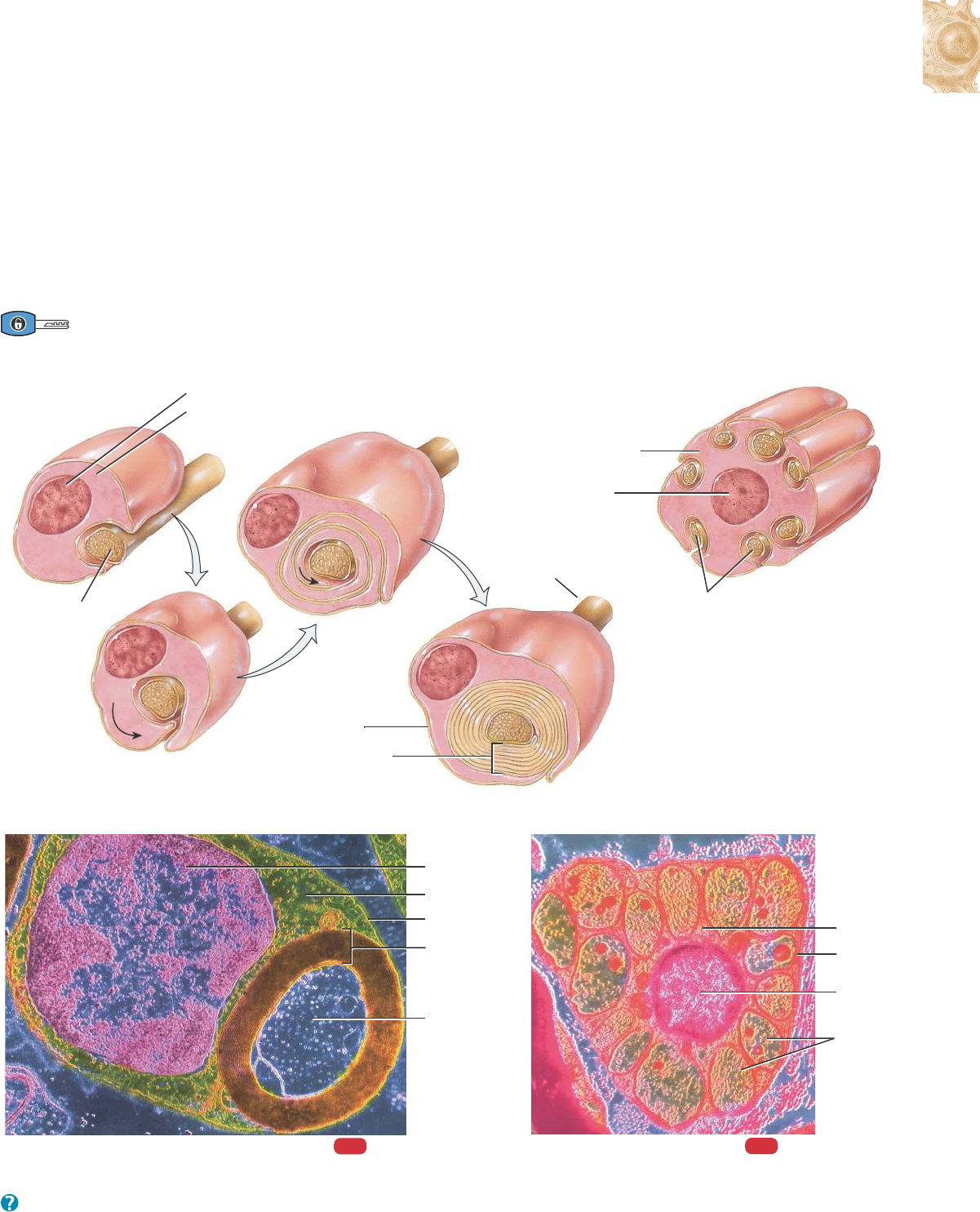
CÉLULAS DE SCHWANN. Estas células rodean los axones del SNP.
Como los oligodendrocitos, forman la vaina de mielina que envuelve
los axones. Sin embargo, un solo oligodendrocito mieliniza a varios
axones, mientras que cada célula de Schwann mieliniza un único
axón (Figura 12.7a; véase también la Figura 12.8a, c). Una sola célu-
la de Schwann también puede rodear 20 o más axones amielínicos
(axones que carecen de la vaina de mielina) (Figura 12.7b). Las célu-
las de Schwann participan en la regeneración axónica, que se alcanza
con más facilidad en el SNP que en el SNC.
CÉLULAS SATÉLITE. Estas células aplanadas rodean los cuerpos
celulares de las neuronas de los ganglios del SNP (Figura 12.7c).
Además de dar soporte estructural, las células satélite también regu-
lan los intercambios de sustancias entre los cuerpos de las neuronas y
el líquido intersticial.
Mielinización
Como ya vimos, se dice que están mielinizados los axones que tie-
nen una vaina de mielina, constituida por múltiples capas de lípidos y
proteínas (Figura 12.8a). Esta vaina actúa como aislante eléctrico del
axón de una neurona y aumenta la velocidad de conducción de los
impulsos nerviosos. Los axones que carecen de esa cubierta se deno-
minan amielínicos (Figura 12.8b).
Hay dos tipos de células gliales que producen vainas de mielina:
las células de Schwann (en el SNP) y los oligodendrocitos (en el
SNC). Las células de Schwann comienzan a formar las vainas de
mielina alrededor de los axones, durante el desarrollo fetal. Cada
célula de Schwann se envuelve varias veces alrededor de un axón y
cubre alrededor de 1 mm de su longitud (Figura 12.8a). Finalmente,
múltiples capas de membranas plasmáticas gliales rodean el axón, y
el citoplasma y el núcleo de las células de Schwann forman parte de
la capa más externa. La porción más interna, constituida hasta por
100 capas superpuestas de la membrana plasmática de las células de
Schwann, forma la vaina de mielina. La capa citoplasmática nuclea-
da externa de la célula de Schwann, que encierra la vaina de mieli-
na, es el neurolema (vaina de Schwann). El neurolema se encuen-
tra solamente alrededor de los axones en el SNP. Cuando un axón
resulta dañado, el neurolema contribuye a la regeneración, median-
te la formación de un conducto que guía y estimula el nuevo creci-
miento del axón. A lo largo de todo el axón se encuentran, a interva-
los regulares, interrupciones de la vaina de mielina denominados
nodos de Ranvier (Figura 12.8; véase también la Figura 12.2). Cada
célula de Schwann envuelve sólo un segmento axónico entre dos
nódulos.
En el SNC, un oligodendrocito mieliniza diferentes segmentos de
varios axones. Cada oligodendrocito emite alrededor de 15 prolonga-
ciones extensas y aplanadas que se enrollan alrededor de los axones
en el SNC y forman la vaina de mielina. Sin embargo, no hay neuro-
lema porque el cuerpo celular y el núcleo del oligodendrocito no
envuelven el axón. Por otra parte, los nodos de Ranvier, aunque pre-
sentes, son menos numerosos. Los axones del SNC tienen poca capa-
456 CAPÍTULO 12 • TEJIDO NERVIOSO
Figura 12.7 Neuroglia del sistema nervioso periférico (SNP).
Las células gliales del SNP rodean completamente los axones y los cuerpos de las neuronas.
¿En qué difieren las células de Schwann y los oligodendrocitos respecto de la cantidad de axones que mielinizan?
Nodo de Ranvier
Cuerpo de célula
neuronal en un ganglio
Célula de
Schwann
Célula de
Schwann
Axones amielínicos
(a) (b) (c)
Vaina de
mielina
Axón
Célula satélite
Célula de
Schwann
Axón
ERRN VPHGL FRVRUJ
cidad para regenerarse después de una lesión. Se piensa que esto se
debe en parte a la ausencia de neurolema y en parte a la influencia
inhibitoria que ejercerían los oligodendrocitos sobre la reconstrucción
de los axones.
La cantidad de mielina aumenta desde el nacimiento hasta la madu-
rez, y su presencia incrementa la velocidad de conducción de los
impulsos nerviosos. Las respuestas de un lactante a un estímulo no
son tan rápidas ni coordinadas como las de un niño mayor o las de un
adulto porque la mielinización, entre otros factores, está todavía en
desarrollo en el lactante.
Colecciones de tejido nervioso
Los componentes del tejido nervioso se agrupan de distintas formas.
12.2 HISTOLOGÍA DEL TEJIDO NERVIOSO 457
Figura 12.8 Axones mielínicos y amielínicos. Se observa que una capa formada por la membrana celular de la célula de Schwann rodea los
axones amielínicos.
Los axones rodeados por una vaina de mielina producida por las células de Schwann en el SNP y por los oligodendrocitos en el SNC se dice
que son mielínicos.
¿Cuál es la ventaja funcional de la mielinización?
Célula de Schwann:
(a) Cortes transversales de los estadios en la formación de una vaina de mielina
Nodo de
Ranvier
Axones amielínicos
Núcleo
Citoplasma
Axón
Neurolema
Vaina de mielina
(b) Corte transversal de axones amielínicos
Núcleo
Citoplasma
Vaina de mielina
Axón amielínico
Neurolema
Célula
de Schwann:
(c) Corte transversal del axón mielínico
MET
5 000x
Núcleo
Citoplasma
Axones
amielínicos
Neurolema
Célula
de Schwann:
(d) Corte transversal de axones amielínico
MET
2 700x
Núcleo
Citoplasma
Núcleo
Célula de Schwann:
ERRN VPHGL FRVRUJ
Los cuerpos de las neuronas, a menudo, se agrupan en conjuntos. Los
axones de las neuronas suelen agruparse juntos en haces. Además,
regiones amplias del tejido nervioso se acumulan como sustancia gris
o sustancia blanca.
Agrupaciones de los cuerpos de las neuronas
Recuerde que un ganglio está constituido por la agrupación de los
cuerpos de células neuronales localizada en el SNP. Como ya mencio-
namos, los ganglios se encuentran íntimamente asociados a los ner-
vios craneales o espinales. Por el contrario, un núcleo es un conjunto
de cuerpos de células neuronales, localizado en el SNC.
Haces de axones
Recordemos que un nervio es un haz de axones localizado en el
SNP. Los nervios craneales conectan el encéfalo con la periferia,
mientras que los nervios espinales conectan la médula espinal con
la periferia. Un tracto es un haz de axones localizado en el SNC.
Los tractos interconectan las neuronas en la médula espinal y el
encéfalo.
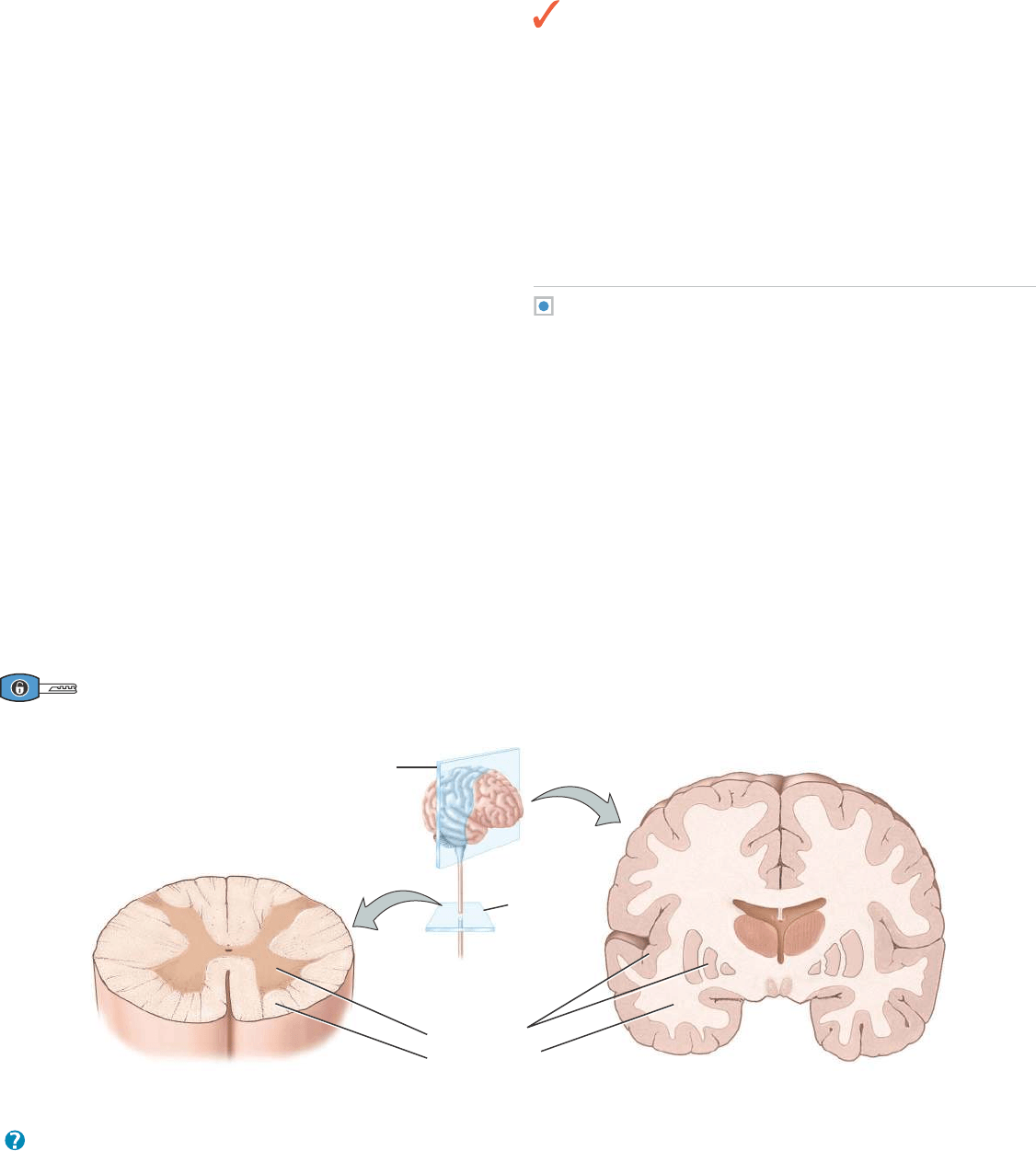
Sustancia gris y sustancia blanca
En los cortes del cerebro o de la médula espinal recientemente dise-
cados, algunas regiones se ven blancas o brillantes, mientras que otras
aparecen grisáceas (Figura 12.9). La sustancia blanca está compues-
ta principalmente por axones mielinícos, y le debe su nombre al color
blanquecino de la mielina. La sustancia gris del sistema nervioso
contiene los cuerpos celulares de las neuronas, dendritas, axones
amielínicos, axones terminales y neuroglia. Tiene un tinte grisáceo
por los cuerpos de Nissl, que le dan ese color, y porque la mielina es
escasa o nula en estas regiones. Tanto la sustancia blanca como la gris
contienen vasos sanguíneos. En la médula espinal, la sustancia blanca
envuelve un núcleo interno de sustancia gris, que dependiendo de su
imaginación, tiene la forma de una mariposa o de la letra H; por su
parte, una delgada capa de sustancia gris cubre la superficie de las
porciones más grandes del encéfalo, el cerebro y el cerebelo (
Figura
12.9). La disposición de la sustancia gris y la blanca en la médula
espinal y en el encéfalo será tratada más extensamente en los
Capítulos 13 y 14, respectivamente.
PREGUNTAS DE REVISIÓN
5. Describa las partes de una neurona y la función de cada
una.
6. Proporcione varios ejemplos de las clasificaciones estruc-
turales y funcionales de las neuronas.
7. ¿Qué es el neurolema? ¿Cuál es su importancia?
8. En referencia al sistema nervioso, ¿qué es un núcleo?
12.3 SEÑALES ELÉCTRICAS EN LAS
NEURONAS
OBJETIVOS
• Describir las propiedades celulares que permiten la comu-
nicación entre las neuronas y sus efectores.
• Comparar los tipos básicos de canales iónicos y explicar
cómo se relacionan con los potenciales de acción y con los
potenciales graduados.
• Describir los factores que mantienen el potencial de mem-
brana de reposo.
• Enumerar secuencia de procesos que conducen a la gene-
ración de un potencial de acción.
Como las fibras musculares, las neuronas son eléctricamente exci-
tables. Se comunican entre sí mediante dos tipos de señales eléctricas:
458 CAPÍTULO 12 • TEJIDO NERVIOSO
Figura 12.9 Distribución de las sustancias gris y blanca en la médula espinal y en el encéfalo.
La sustancia blanca está constituida principalmente por axones mielínicos provenientes de muchas neuronas. La sustancia gris se halla forma-
da por cuerpos celulares, dendritas, terminales axónicos, axones amielínicos y neuroglia.
¿Qué sustancia es responsable del aspecto de la sustancia blanca?
Plano frontal
a través del
encéfalo
Plano
transverso
a través de
la médula
espinal
Sustancia gris
(a) Plano transverso a través de la médula espinal (b) Corte frontal del encéfalo
Sustancia blanca
ERRN VPHGL FRVRUJ
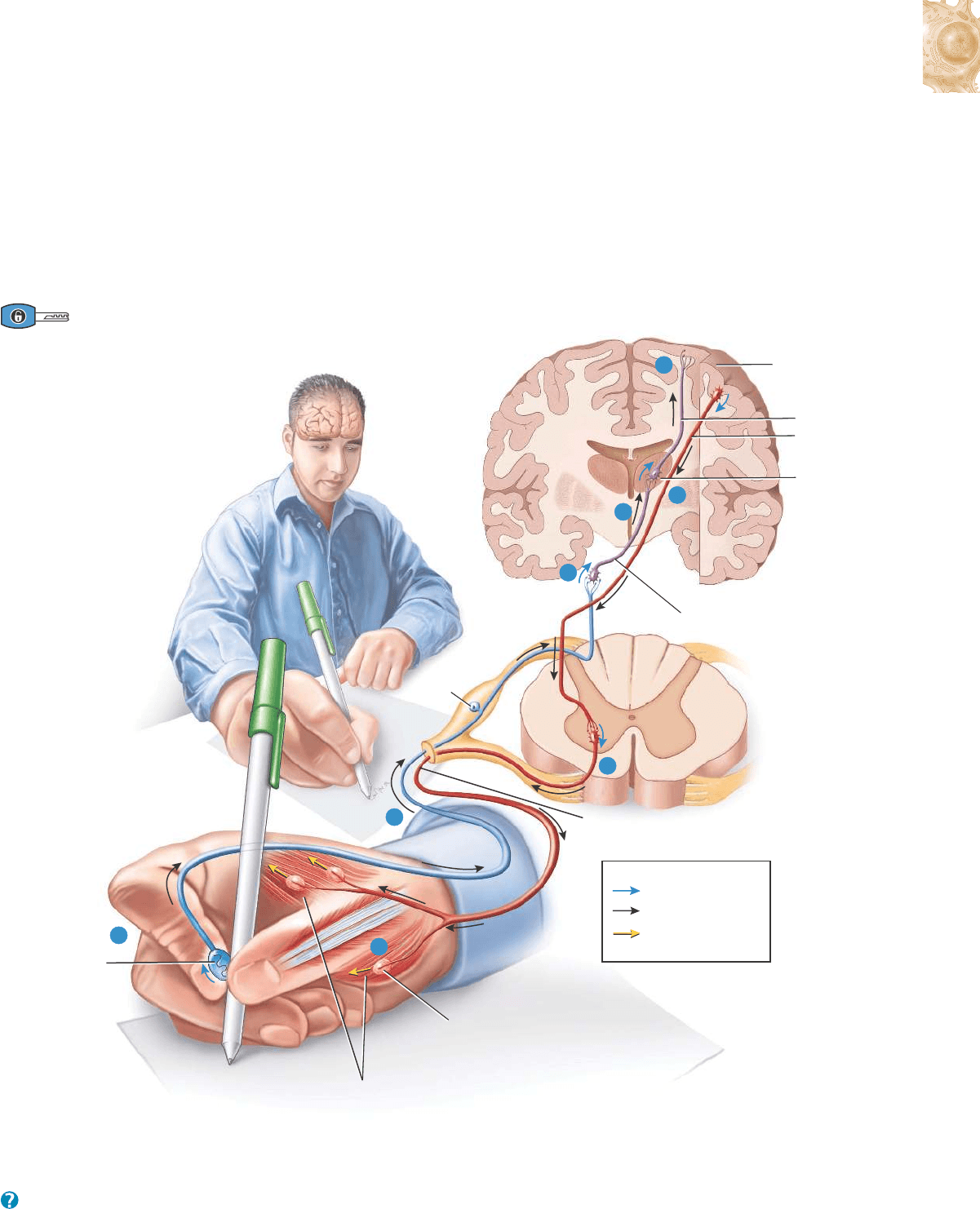
(1) Los potenciales graduados (descritos más adelante) se utilizan
sólo para las comunicaciones a corta distancia. (2) Los potenciales de
acción (también descritos más adelante) permiten la comunicación
con lugares cercanos y lejanos, dentro del cuerpo. Recuerde que un
potencial de acción en una fibra muscular se denomina potencial de
acción muscular. Cuando se produce un potencial de acción en una
neurona (célula nerviosa), recibe el nombre de potencial de acción
nervioso (impulso nervioso). Para comprender las funciones de los
potenciales graduados y de los potenciales de acción, considere cómo
hace su sistema nervioso para lograr que usted perciba la superficie
lisa de un bolígrafo que levantó de una mesa (Figura 12.10):
1
Mientras usted toca el bolígrafo, se desarrolla un potencial gradua-
do en los receptores sensitivos de la piel de sus dedos.
2
El potencial graduado estimula el axón de la neurona sensitiva para
que inicie un potencial de acción nervioso, que se dirige a lo largo
del axón hacia el SNC y, por último, causa la liberación de neuro-
transmisores en la sinapsis con una interneurona.
12.3 SEÑALES ELÉCTRICAS EN LAS NEURONAS 459
Figura 12.10 Generalidades de las funciones del sistema nervioso.
¿En qué región del cerebro se produce principalmente la percepción?
Potencial graduado
Potencial de acción
nervioso
Potencial de acción
muscular
Lado derecho del encéfaloLado derecho del encéfalo
Corteza cerebral
Encéfalo
Tálamo
Neurona motora inferior
Músculos esqueléticos
Unión
neuromuscular
Neurona motora
superior
Interneurona
Médula espinal
Clave:
Interneurona
Neurona
sensitiva
Receptor
sensitivo
5
6
4
3
2
7
1
8
Los potenciales graduados y los potenciales de acción
nerviosos y musculares intervienen en el relevo de estímulos
sensitivos en funciones integradoras como la percepción,
y en actividades motoras.
ERRN VPHGL FRVRUJ
3
El neurotransmisor estimula la interneurona para que inicie un
potencial graduado en sus dendritas y en su cuerpo celular.
4
En respuesta a este potencial graduado, el axón de la interneurona
produce un potencial de acción nervioso. Éste se desplaza a lo largo
del axón, y de ello resulta la liberación de neurotransmisores en la
próxima sinapsis con otra interneurona.
5
Este proceso de liberación de neurotransmisores en la sinapsis,
seguido de la formación de un potencial graduado y luego de un
potencial de acción nervioso ocurre una y otra vez a medida que se
activan interneuronas localizadas en sectores superiores del encéfa-
lo (como el tálamo y la corteza cerebral). La percepción se produce
una vez que se activan las interneuronas de la zona más externa del
cerebro, la corteza cerebral, y usted puede sentir la superficie lisa
de un bolígrafo sostenido entre sus dedos. Como se verá en el
Capítulo 14, la percepción, la noción consciente de una sensación,
es fundamentalmente una función de la corteza cerebral.
Suponga que usted quiere utilizar ese bolígrafo para escribir una
carta. El sistema nervioso respondería de la siguiente manera (
Figura
12.10):
6
Un estímulo en el cerebro produce un potencial graduado en las den-
dritas y en el cuerpo celular de una neurona motora superior, un
tipo de neurona motora que hace sinapsis con una neurona motora
inferior situada más abajo en el SNC, y de esta forma provoca la
contracción de un músculo esquelético. Este potencial graduado
genera luego la producción de un potencial de acción nervioso en el
axón de la neurona motora superior, seguido de la liberación de neu-
rotransmisores.
7
El neurotransmisor origina un potencial graduado en una neurona
motora inferior, un tipo de neurona motora que inerva directamen-
te las fibras musculares esqueléticas. El potencial graduado desen-
cadena un potencial de acción nervioso y luego la liberación de neu-
rotransmisores en la unión neuromuscular que se forma con las
fibras musculares esqueléticas que controlan el movimiento de los
dedos.
8
El neurotransmisor estimula la formación de un potencial de acción
muscular en estas fibras musculares, que provocan la contracción de
las fibras musculares de los dedos; esto le permite a usted escribir
con el bolígrafo.
La generación de potenciales graduados y de potenciales de acción
depende de dos características fundamentales de la membrana plas-
mática de las células excitables: la existencia de un potencial de mem-
brana de reposo y la presencia de tipos específicos de canales iónicos.
Como ocurre en todas las otras células del cuerpo, la membrana plas-
mática de las células excitables presenta un potencial de membrana,
una diferencia de potencial eléctrico (voltaje) a través de esa membra-
na. En las células excitables, este voltaje se denomina potencial de
membrana de reposo. El potencial de membrana es similar al volta-
je que se almacena en un acumulador. Si se conectan los terminales
positivo y negativo de un acumulador con un cable, los electrones flui-
rán a lo largo de éste. Ese flujo de partículas cargadas se denomina
corriente. En las células vivas, el flujo de iones (más que de electro-
nes) constituye las corrientes eléctricas.
Los potenciales graduados y los potenciales de acción se producen
porque las membranas de las neuronas contienen muchos tipos dife-
rentes de canales iónicos que se abren o se cierran, en respuesta a estí-
mulos específicos. Como la bicapa lipídica que constituye la membra-
na plasmática es un buen aislante eléctrico, las vías principales que
utiliza el flujo iónico para pasar a través de la membrana son los cana-
les iónicos.
Canales iónicos
Cuando los canales iónicos están abiertos permiten el paso de iones
específicos a través de la membrana plasmática, los cuales siguen su
gradiente electroquímico: una diferencia en la concentración de sus-
tancias químicas sumada a una diferencia de cargas eléctricas.
Recuerde que los iones se mueven desde áreas en las que se encuen-
tran en mayor concentración hacia regiones en las que se hallan en
menor concentración (el componente químico del gradiente). Además,
los cationes con cargas positivas se desplazan hacia zonas con cargas
negativas, y los aniones con cargas negativas se dirigen hacia áreas
con cargas positivas (el componente eléctrico del gradiente). A medi-
da que los iones se mueven, crean una corriente eléctrica que puede
cambiar el potencial de membrana.
Los canales iónicos se abren y se cierran por la presencia de “com-
puertas”. Una compuerta es una parte de la proteína del canal que
puede cerrar el poro o hacerse a un lado para abrirlo (véase la Figura
3.6). Las señales eléctricas que producen las neuronas y las fibras
musculares dependen de cuatro tipos de canales: canales pasivos,
canales dependientes de ligandos, canales accionados mecánicamente
y canales dependientes del voltaje:
1. Las compuertas de los canales pasivos alternan al azar entre las
posiciones abierta y cerrada (Figura 12.11a). Habitualmente, las
membranas plasmáticas tienen muchos más canales pasivos para el
ion potasio (K
+
) que para el ion sodio (Na
+
), y los canales pasivos
para el ion potasio son más permeables que los canales pasivos para
el ion sodio. De tal forma, la permeabilidad de la membrana
para el K
+
es mucho más alta que para el Na
+
. Los canales pasivos
se encuentran en casi todas las células, que incluyen las dendritas,
los cuerpos celulares y los axones de todos los tipos de neuronas.
2. Los canales dependientes de ligandos se abren y se cierran en res-
puesta a estímulos ligandos (químicos) específicos. Una amplia
variedad de ligandos químicos (como neurotransmisores, hormonas
y algunos iones en particular) pueden abrirlos o cerrarlos. El neuro-
transmisor acetilcolina, por ejemplo, abre canales catiónicos que per-
miten la difusión hacia el interior de la célula de iones Na
+
y Ca
2+
y
la difusión hacia afuera de iones K
+
(Figura 12-11b). Los canales
dependientes de ligandos se localizan en las dendritas de algunas
neuronas sensitivas, como los receptores del dolor, y en las dendritas
y en los cuerpos celulares de interneuronas y neuronas motoras.
3. Los canales accionados mecánicamente se abren o se cierran
luego de una estimulación mecánica, que puede tomar la forma de
una vibración (como las ondas sonoras), el tacto, la presión o el
estiramiento de un tejido (Figura 12.11c). La fuerza ejercida distor-
siona la posición de reposo del canal y abre su compuerta. Entre los
ejemplos de canales operados mecánicamente están aquellos que se
encuentran en los receptores auditivos del oído, en los receptores
que monitorizan el estiramiento de los órganos internos y en los
receptores para el tacto y la presión de la piel.
4. Los canales dependientes del voltaje se abren en respuesta a un
cambio en su potencial de membrana (voltaje) (Figura 12.11d).
Estos canales participan en la generación y conducción de los
potenciales de acción, en los axones de todos los tipos de neuronas.
En el Cuadro 12.1 se presenta un resumen de los cuatro tipos
principales de canales iónicos de las neuronas.
Potencial de membrana en reposo
El potencial de membrana en reposo es la consecuencia de la peque-
ña acumulación de iones negativos en el citosol a lo largo de la super-
460 CAPÍTULO 12 • TEJIDO NERVIOSO
ERRN VPHGL FRVRUJ

12.3 SEÑALES ELÉCTRICAS EN LAS NEURONAS 461
Figura 12.11 Canales iónicos en la membrana plasmática: a) Los canales pasivos se abren y cierran al azar. b) Un estímulo químico
(aquí, el neurotransmisor acetilcolina) abre un canal dependiente de ligando. c) Un estímulo mecánico abre un canal activado
mecánicamente. d) Un cambio en el potencial de membrana abre los canales de K+ dependientes de voltaje durante un
potencial de acción.
Las señales eléctricas producidas por neuronas y fibras musculares se basan en cuatro tipos de canales iónicos: canales pasivos, canales
dependientes de ligando, canales activados mecánicamente y canales dependientes del voltaje.
¿Qué tipo de canal con compuerta es activado por un roce en el brazo?
(b) Canal dependiente de ligando
(a) Canal pasivo
El canal se abre y cierra
El estímulo químico
abre el canal
Canal pasivo de
potasio cerrado
Canal pasivo de
potasio abierto
Acetilcolina
Canal dependiente
de ligando abierto
Canal dependiente
de ligando cerrado
Na
+
Ca
2+
Na
+
K
+
K
+
(d) Canal dependiente de voltaje
Cambio en el potencial
de membrana abre el canal
(c) Canal activado mecánicamente
El estímulo mecánico
abre el canal
Canal de potasio
dependiente de
voltaje cerrado
Canal de potasio
dependiente de
voltaje abierto
Voltaje
= –50 mV
Voltaje= –70 mV
K
+
K
+
Canal activado
mecánicamente cerrado
Canal activado
mecánicamente
abierto
Ca
2+
Líquido
extracelular
Membrana
plasmática
Citosol
en forma aleatoria
K
+
ERRN VPHGL FRVRUJ
Este documento contiene más páginas...
Descargar Completo
Anatomia-y-fisiologia. Nervioso I..pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.