
11
Capítulo
ADN: la molécula de la herencia
䊱
¿Toro común o el increíble Hulk? Un cambio diminuto del ADN
marca toda la diferencia.
Estudio de caso
Músculos, mutaciones
y miostatina
NO, EL TORO que aparece en la parte superior
de la fotografía no levanta pesas: es un toro de
raza Belgian Blue, una raza que tiene músculos
prominentes. ¿A qué se debe que un Belgian Blue
parezca un fisicoconstructivista en comparación
con las reses ordinarias, como el Herford de la parte
inferior de la fotografía?
Cuando un mamífero se desarrolla, sus células se
dividen muchas veces, se agrandan y se especializan
para una función determinada. El tamaño, forma y
tipo de las células de los órganos están regulados
con precisión durante el desarrollo, de modo que
uno no adquiere una cabeza del tamaño de una
pelota de basquetbol ni le crece pelo en el hígado. El
desarrollo muscular no es la excepción. Cuando eras
pequeño, las células destinadas a formar tus músculos
se dividieron, se unieron y formaron células largas
y relativamente gruesas con numerosos núcleos y
sintetizaron las proteínas especializadas que hacen que
los músculos se contraigan para mover el esqueleto.
Una proteína llamada miostatina, que se encuentra en
todos los mamíferos, detiene este proceso. La palabra
“miostatina” significa de forma literal “hacer que los
músculos se queden iguales” y eso es exactamente
lo que hace. Cuando los músculos se desarrollan, la
miostatina lentifica y al final detiene las divisiones
de estas células premusculares. La miostatina regula
también el tamaño definitivo de las células musculares
y, por tanto, su fuerza. Un fisicoconstructivista aumenta
su volumen muscular levantando pesas, lo cual
agranda sus células musculares pero, por lo regular,
no adiciona muchas células nuevas.
El Belgian Blue tiene más células musculares que
son más grandes que las de las reses comunes. ¿Por
qué? Es posible que ya lo hayas imaginado: porque
no producen miostatina normal. ¿Y por qué no? Como
veremos en este capítulo, las proteínas se sintetizan
según las instrucciones genéticas contenidas en el
ácido desoxirribonucleico (ADN). El ADN del Belgian
Blue es significativamente diferente del ADN del ganado
común: tiene un cambio, una mutación en el ADN
del gen de la miostatina. Por consiguiente, produce
miostatina defectuosa. Las células premusculares del
Belgian Blue se dividen más de lo normal y se hacen
muy grandes a medida que se diferencian, lo que
produce este ganado de gran tamaño.
¿Cómo es que el ADN contiene las instrucciones
de rasgos como el tamaño de los músculos,
color de las flores o la determinación del sexo?
¿Cómo se transmiten estas instrucciones de
generación en generación sin alterarse? ¿Por qué a
veces estas instrucciones cambian? Las respuestas se
encuentran en la estructura y la función del ADN.

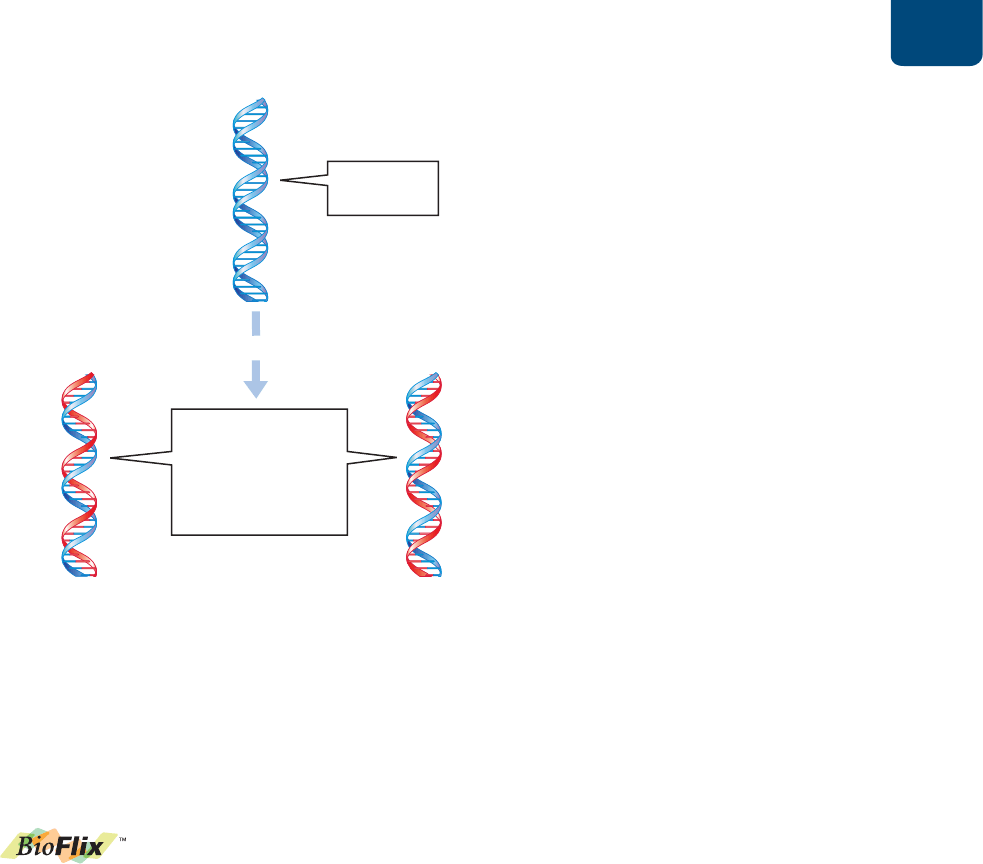
ADN: la molécula de la herencia Capítulo 11 201
11.1
¿CÓMO DESCUBRIERON
LOS CIENTÍFICOS QUE LOS GENES
ESTÁN HECHOS DE ADN?
A finales del siglo XIX, los científicos habían aprendido que la
información genética se encuentra en unidades individuales que
llamaron genes. Sin embargo, no sabían qué eran los genes; sólo
sabían que los genes determinan muchas diferencias hereditarias
en los organismos de una especie. Por ejemplo, los genes del co-
lor de las flores determinan si las rosas son rojas, rosas, amarillas
o blancas. A comienzos del siglo XX, los estudios sobre la división
celular proporcionaron evidencias sólidas de que los genes son
parte de los cromosomas (véanse las páginas 149-150, 174-175).
Poco después, los bioquímicos encontraron que los cromosomas
eucariontes están compuestos únicamente de proteínas y ADN.
Uno de estos compuestos debía llevar el plano hereditario de las
células, pero ¿cuál?
Bacterias transformadas revelaron
el vínculo entre genes y ADN
A finales de la década de 1920, un científico inglés de nombre
Frederick Griffith trataba de elaborar una vacuna para prevenir la
neumonía bacteriana, una de las principales causas de muerte en
la época. Es muy difícil preparar una vacuna contra ciertos tipos de
bacterias infecciosas (por ejemplo, las vacunas modernas contra
el ántrax no son totalmente seguras ni eficaces por completo),
pero esto no se sabía en 1920. Algunas vacunas antibacterianas
constan de una cepa debilitada de las bacterias, que no causan la
enfermedad. Al inyectar esta cepa debilitada pero viva en un ani-
mal, se estimula la inmunidad contra el patógeno. Otras vacunas
están hechas de bacterias virulentas que fueron muertas mediante
exposición al calor o a compuestos químicos.
Griffith trataba de hacer una vacuna con dos cepas de la bac-
teria Streptococcus pneumoniae. Una cepa, R, no causó neumonía al
inyectarla en ratones (
FIGURA 11-1a). La otra cepa, S, resultó mor-
tal al inyectarla: causó neumonía y mató a los ratones en uno o
dos días (
FIGURA 11-1b). Como era de esperarse, cuando se mató
a las bacterias de la cepa S y se inyectaron en los ratones, no causa-
ron la enfermedad (
FIGURA 11-1c). Por desgracia, ni la cepa viva R
ni la cepa muerta S proporcionaron inmunidad contra las bacterias
vivas de la cepa S.
Griffith trató igualmente de mezclar bacterias de la cepa R
con bacterias de la cepa S muertas por calor e inyectar la mezcla a ra-
tones (
FIGURA 11-1d). Como ninguna de estas cepas causaba neu-
monía por sí misma, esperaba que los ratones conservaran la salud.
Para su sorpresa, los ratones se enfermaron y murieron. Cuando
hizo la autopsia de los animales, obtuvo bacterias vivas de la cepa S.
Griffith planteó la hipótesis de que algo de las bacterias muertas por
calor de la cepa S había cambiado las bacterias vivas e inofensivas
de la cepa R y las había convertido en la mortífera cepa S, por un
proceso llamado transformación. A continuación, las células de la
cepa S transformadas se multiplicaron y causaron neumonía.
Griffith nunca descubrió una vacuna eficaz contra la neu-
monía y, en ese sentido, sus experimentos fueron un fracaso
(de hecho, hasta finales de la década de 1970 no se desarrolló
una vacuna eficaz y segura contra la mayor parte de las formas
de Streptococcus pneumoniae). Sin embargo, los experimentos de
Griffith fueron un punto de inflexión en nuestra comprensión
de la genética, porque otros investigadores sospechaban que aque-
llo que causó la transformación podía ser la muy buscada mo-
lécula de la herencia.
Estudio de caso Músculos, mutaciones y miostatina
11.1 ¿Cómo descubrieron los científicos
que los genes están hechos de ADN?
Bacterias transformadas revelaron el vínculo entre genes
y ADN
11.2 La estructura del ADN
El ADN está compuesto por cuatro nucleótidos
El ADN es una doble hélice con dos hebras de nucleótidos
Investigación científica El ADN es la molécula
de la herencia de los bacteriófagos
Enlaces de hidrógeno entre bases complementarias
mantienen unidas las dos cadenas de ADN en la doble
hélice
11.3 ¿Cómo codifica la información el ADN?
Investigación científica El descubrimiento de la doble
hélice
Estudio de caso continuación Músculos, mutaciones
y miostatina
De un vistazo
11.4
¿Cómo es que la replicación del ADN
garantiza la constancia genética durante
la división celular?
La replicación del ADN es un acontecimiento fundamental en
el ciclo celular
La replicación del ADN produce dos dobles hélices de ADN,
cada una con la hebra original y una nueva
Estudio de caso continuación Músculos, mutaciones
y miostatina
De cerca Estructura y replicación del ADN
DNA Replication (disponible en inglés)
11.5 ¿Cómo ocurren las mutaciones?
La replicación precisa y la revisión producen ADN casi sin errores
Los errores pueden ocurrir
Las mutaciones van de cambios en pares de nucleótidos
simples a desplazamientos de grandes piezas de cromosomas
Las mutaciones tienen efectos diversos en las funciones
Estudio de caso otro vistazo Músculos, mutaciones y
miostatina
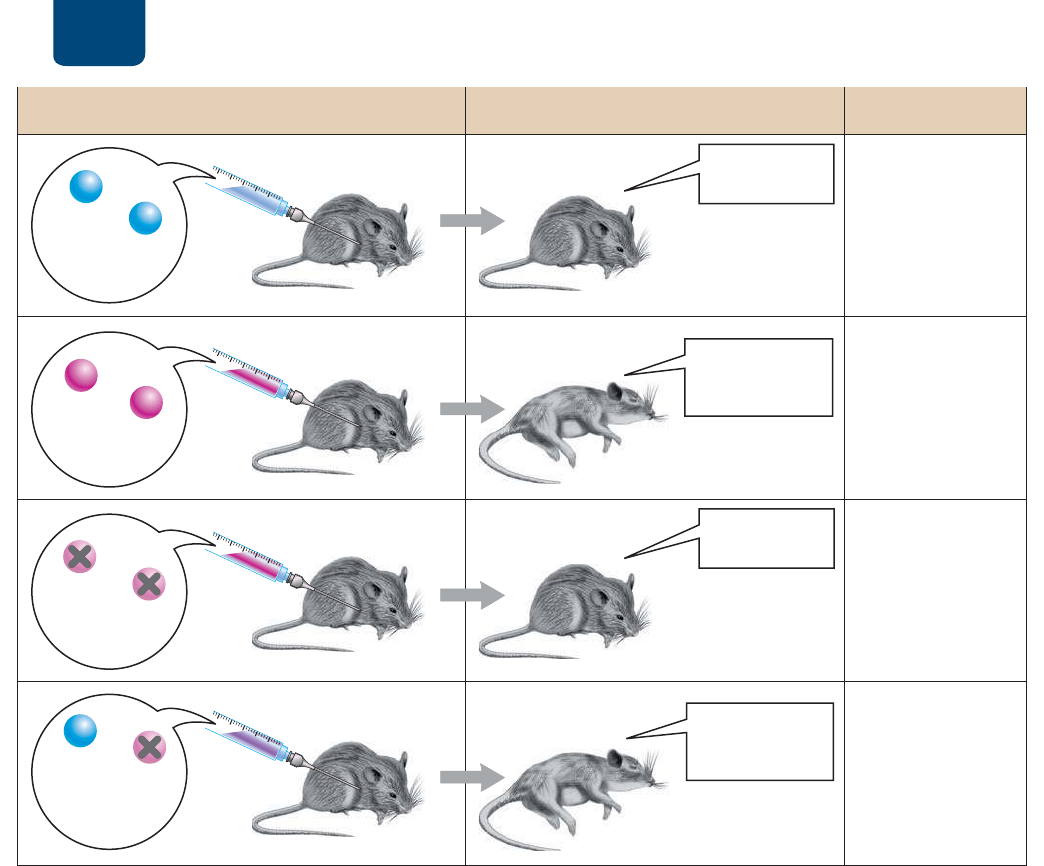
202 UNIDAD 2 Herencia
(OUDWʼnQHVWÀ
VDQR
(OUDWʼnQHVWÀ
VDQR
(OUDWʼnQFRQWUDH
QHXPRQķD\
PXHUH
&HSD
9YLYD
&HSD
:YLYD
&HSD:PXHUWD
SRUFDORU
0H]FODGH
FHSD9YLYD\
FHSD:PXHUWD
SRUFDORU
(OUDWʼnQFRQWUDH
QHXPRQķD\
PXHUH
H
I
J
K
*LWHZKLIHJ[LYPHZPU`LJ[HKHZHYH[VULZ 9LZ\S[HKVZ *VUJS\ZPVULZ
/DFHSD9
QRFDXVD
QHXPRQķD
/DFHSD:FDXVD
QHXPRQķD
/DFHSD:PXHUWD
SRUFDORUQR
FDXVDQHXPRQķD
$OJRGHODFHSD:
PXHUWDSRUFDORU
WUDQVIRUPDOD
FHSD9LQRIHQVLYD
HQXQDFHSD:
PRUWDO
䊱
FIGURA 11-1 La transformación “bacteriana” El descubrimiento de Griffith de que las bacterias
pueden transformarse de inofensivas en mortales estableció las bases para el hallazgo de que los genes
están compuestos de ADN.
La molécula de la transformación es el ADN
En 1933, J.L. Alloway descubrió que los ratones no desempeñaron
ninguna función en la transformación, como lo que ocurrió en las
cajas de Petri de cultivo, cuando las bacterias vivas de la cepa R
se mezclaron con las bacterias muertas de la cepa S. Una década
más tarde, Oswald Avery, Colin MacLeod y Maclyn McCarty des-
cubrieron que la molécula de la transformación es el ADN. Avery,
MacLeod y McCarty aislaron el ADN de bacterias de la cepa S, mez-
cladas con bacterias de la cepa R y produjeron bacterias vivas de la
cepa S. Para mostrar que la transformación era causada por ADN y
no por restos de proteínas que hubieran contaminado el ADN,
trataron algunas muestras con enzimas que destruyeron las pro-
teínas. Estas enzimas no evitaron la transformación; en cambio, al
tratar las muestras con enzimas que destruyen el ADN, se impidió
la transformación.
Este descubrimiento nos sirve para interpretar los resulta-
dos de los experimentos de Griffith. Al calentar las células de la
cepa S, murieron, pero no se destruyó completamente su ADN.
Cuando las bacterias muertas de la cepa S se mezclaron con bacte-
rias vivas de la cepa R, fragmentos del ADN de las células muertas
de la cepa S pasaron a algunas células de la cepa R y se incorpo-
raron al cromosoma de las bacterias de la cepa R (
FIGURA 11-2).
Si estos fragmentos de ADN contenían los genes necesarios para
causar la enfermedad, una célula de la cepa R se transformaría en
una célula de la cepa S. Así, Avery, MacLeod y McCarty concluye-
ron que los genes están hechos de ADN.
El ADN, no la proteína, es la molécula de la herencia
Ahora bien, no todos se sintieron convencidos. Algunos pensaban
todavía que los genes estaban hechos de proteínas y que la transfor-
mación de las moléculas de ADN de bacterias de la cepa S habían
causado una mutación en los genes de las bacterias de la cepa R.
Otros formularon la hipótesis de que el ADN podría ser la molécu-
la de la herencia de las bacterias, pero no de otros organismos. Sin
embargo, se siguieron acumulando las evidencias de que el ADN es
el material genético de muchos, o quizá de todos los organismos.
Por ejemplo, antes de dividirse, una célula eucarionte duplica sus
cromosomas (véanse las páginas 151-152) y replica exactamente el
contenido de ADN, como sería de esperar si los genes están hechos
de ADN. Por último, casi todos los escépticos que quedaban se
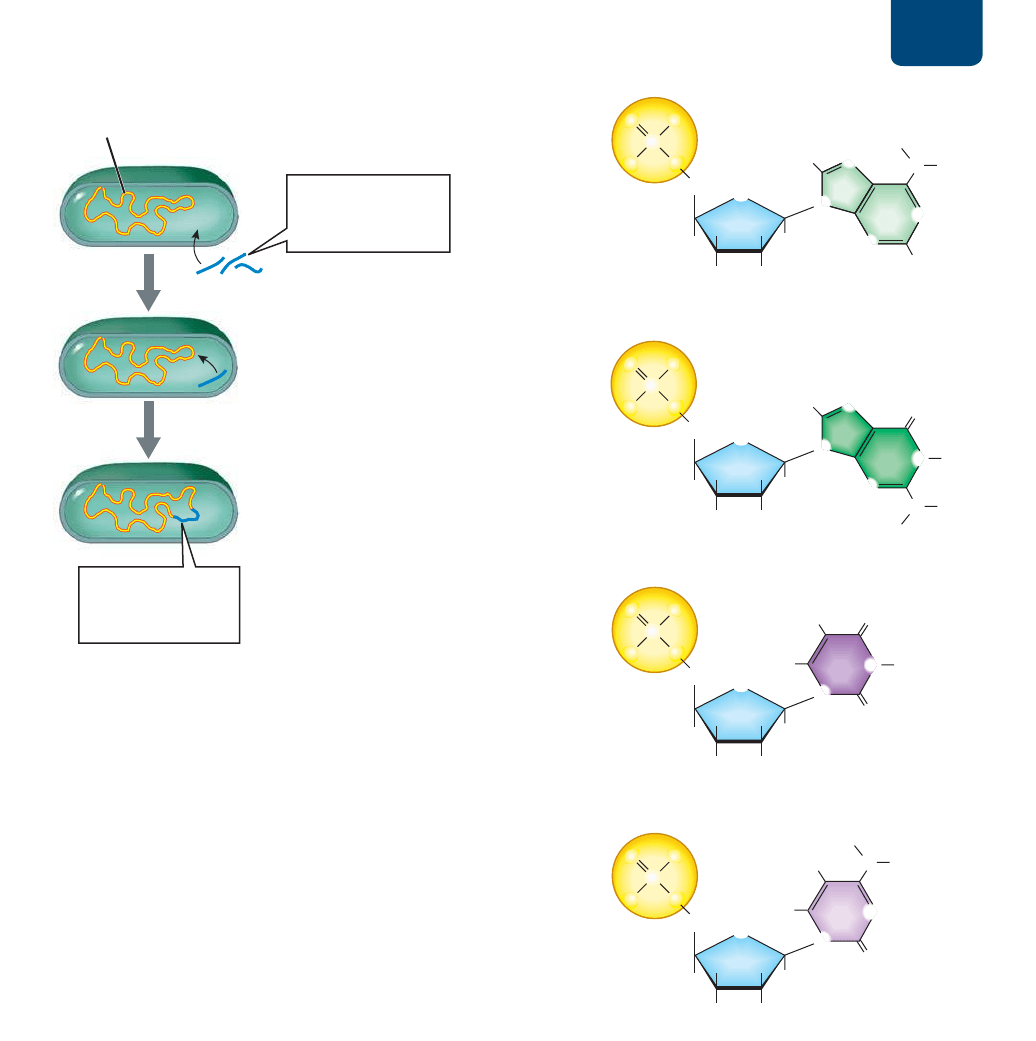
ADN: la molécula de la herencia Capítulo 11 203
XQIUDJPHQWRGH
$'1VHLQFRUSRUD
DOFURPRVRPD
IUDJPHQWRVGH$'1
VRQWUDQVSRUWDGRV
GHQWURGHODEDFWHULD
FURPRVRPD
EDFWHULDQR
䊱
FIGURA 11-2 Mecanismo molecular de la transformación
Casi todas las bacterias tienen un único cromosoma circular
hecho de ADN. Ocurre la transformación cuando una bacteria
viva toma parte del ADN de su ambiente y lo incorpora en su
cromosoma.
convencieron con un grupo soberbio de experimentos de Alfred
Hershey y Martha Chase, en los cuales demostraron en forma con-
cluyente que el ADN es la molécula de la herencia de ciertos virus
(véase el apartado “Investigación científica: “El ADN es la molécula
de la herencia de los bacteriófagos”, en las páginas 204-205).
11.2
LA ESTRUCTURA DEL ADN
Saber que los genes están hechos de ADN no responde pregun-
tas cruciales sobre la herencia. ¿Cómo codifica la información el
ADN? ¿Cómo se replica el ADN de modo que la célula transmita
la información hereditaria a sus células hijas? Los secretos del fun-
cionamiento del ADN y, por consiguiente, de la herencia en sí, se
encuentran en la estructura tridimensional de la molécula de ADN.
El ADN está compuesto por cuatro nucleótidos
Como vimos en el capítulo 3, el ADN consta de cuatro pequeñas
unidades llamadas nucleótidos. Cada nucleótido del ADN tiene
tres partes: un grupo fosfato, un azúcar llamado desoxirribosa y
una de cuatro bases nitrogenadas: adenina (A), guanina (G),
timina (T) o citosina (C) (
FIGURA 11-3).
En la década de 1940, cuando el bioquímico Erwin Chargaff
de la Columbia University (Universidad Columbia) analizó las
cantidades de las cuatro bases del ADN de organismos tan diver-
sos como bacterias, morsas, peces y seres humanos, vio que mos-
traban una curiosa constancia. El ADN de cualquier especie tenía
las mismas cantidades de adenina y timina, así como las mismas
cantidades de guanina y citosina. Esta constancia, que suele lla-
marse la “regla de Chargaff”, pareció ser importante, pero pasaría
casi otra década hasta que alguien entendiera lo que significaba
con respecto a la estructura del ADN.
El ADN es una doble hélice con dos cadenas
de nucleótidos
No es tarea fácil determinar la estructura de cualquier molécula
biológica, ni siquiera para los científicos contemporáneos. Sin em-
bargo, a finales de la década de 1940 varios científicos comenzaron
a investigar la estructura del ADN. Los científicos ingleses Maurice
Wilkins y Rosalind Franklin aplicaron el método de la difracción
por rayos X al estudio de la molécula de ADN. Bombardearon cris-
tales de ADN purificado con rayos X y tomaron nota de cómo re-
H
O
H
O
H
O
H
O
H
H
H
H
H
N
N
N
N
N
O
P
O
O
O
O
P
O
O
O
O
P
O
O
O
O
P
O
O
O
D]ŜFDU
IRVIDWR
H
H
N
N
N
D]ŜFDU
IRVIDWR
H
H
H
N
N
N
N
N
D]ŜFDU
IRVIDWR
CH
3
N
N
D]ŜFDU
IRVIDWR
EDVHHKLUPUH
EDVHN\HUPUH
EDVH[PTPUH
EDVHJP[VZPUH
H
H
H
H
H
OH
CH
2
O
H
H
H
H
H
OH
CH
2
O
H
H
H
H
H
OH
CH
2
O
H
H
H
H
H
OH
CH
2
O
䊱
FIGURA 11-3 Nucleótidos de ADN
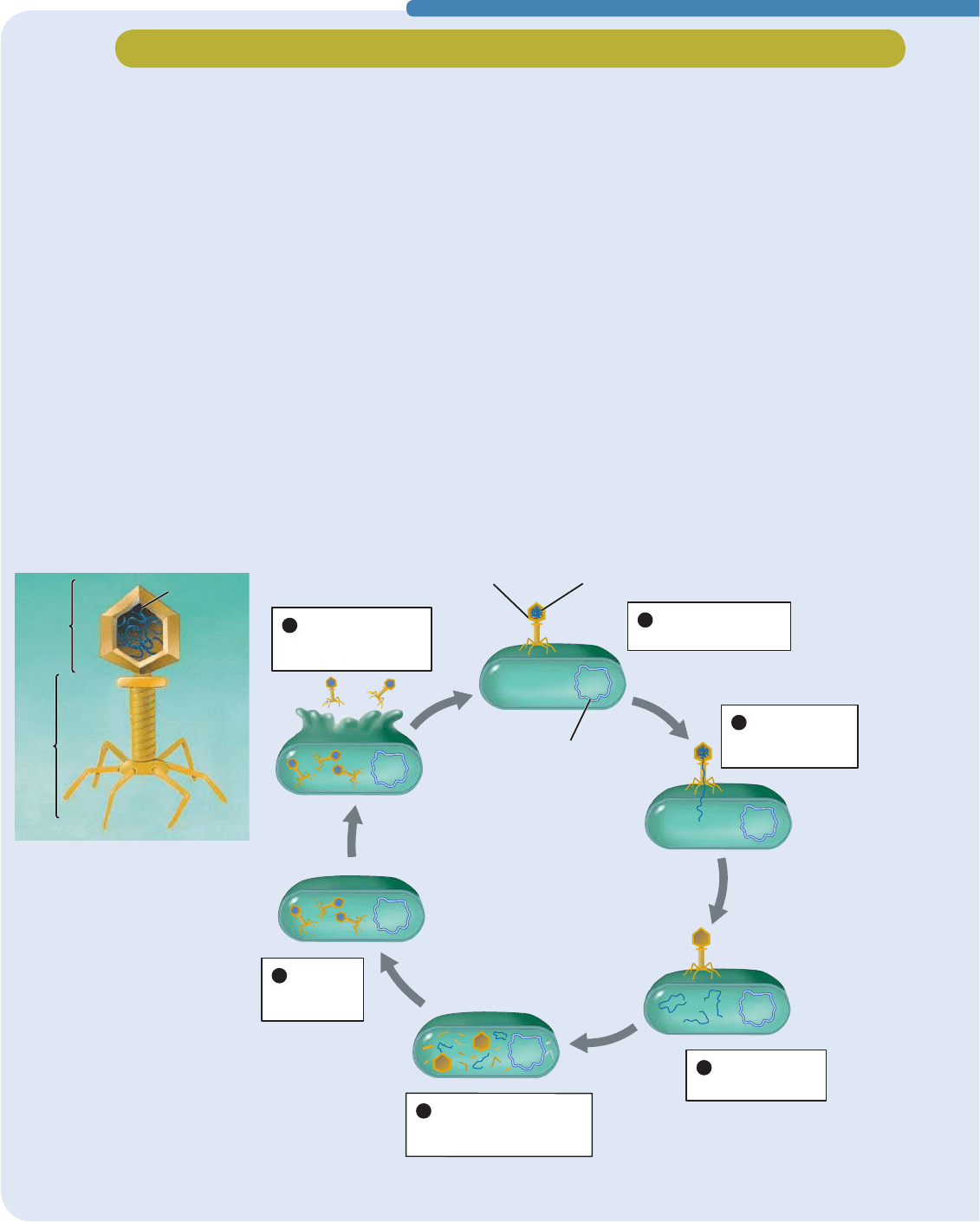
Investigación científica
El ADN es la molécula de la herencia de los bacteriófagos
Ciertos virus infectan sólo bacterias y se llaman bacteriófagos,
lo que significa “comedores de bacterias” (FIGURA E11-1). Un
bacteriófago (“fago” para abreviar) depende de su bacteria
huésped para cubrir todos los aspectos de su ciclo de vida
(
FIGURA E11-1b). Cuando un fago encuentra una bacteria,
se adhiere a la pared celular de ésta y le inyecta su material
genético. La envoltura externa del fago queda fuera de la pared
celular. La bacteria no distingue los genes del fago de sus
propios genes, así que “interpreta” los genes del fago y usa la
información para producir más fagos. Por último, uno de los
genes del fago dirige la síntesis de una enzima que rompe la
bacteria, lo que deja en libertad a los fagos recién constituidos.
Aunque muchos bacteriófagos tienen estructuras intrincadas
(véase la FIGURA E11-1a), químicamente son muy simples, pues
sólo constan de ADN y proteína. Por tanto, una de estas dos
moléculas debe ser el material genético del fago. A comienzos
de la década de 1950, Alfred Hershey y Martha Chase
aprovecharon la sencillez química de los bacteriófagos para
deducir que su material genético es el ADN.
Hershey y Chase sabían que las bacterias infectadas deben
contener material genético del fago, así que si pudieran “marcar”
el ADN y la proteína del fago, y separar las bacterias infectadas
de la envoltura del fago que queda fuera, podrían ver qué
molécula entra en la bacteria (FIGURA E11-2). Como vimos
en el capítulo 3, tanto el ADN como las proteínas contienen
carbono, oxígeno, hidrógeno y nitrógeno. El ADN contiene
también fósforo, pero no azufre, mientras que las proteínas
contienen azufre (en los aminoácidos metionina y cisteína), pero
no fósforo.
Hershey y Chase forzaron a una población de fagos a
sintetizar ADN usando fósforo radiactivo, con lo cual marcaron
su ADN. Otra población fue forzada a sintetizar proteínas
con azufre radiactivo y así quedaron marcadas. Cuando las
bacterias fueron infectadas por los fagos que contenían proteínas
marcadas radiactivamente, las bacterias no se volvieron
radiactivas. En cambio, cuando las bacterias fueron infectadas
por los fagos que contenían ADN radiactivo, las bacterias se
hicieron radiactivas. Hershey y Chase concluyeron que el ADN,
no las proteínas, es el material genético de los fagos.
Hershey y Chase también infirieron que parte del material
genético marcado de los fagos “originales” podría incorporarse
en el material genético de los fagos “descendientes” (veremos
más sobre este tema en la sección 11.4). En un segundo grupo
de experimentos, los investigadores volvieron a marcar el
ADN de una población de fagos y las proteínas en otra población,
y dejaron que los fagos infectaran bacterias. Cuando transcurrió
el tiempo suficiente para que los fagos se reprodujeran, los
investigadores abrieron las bacterias y los fagos descendientes
fueron separados de entre los restos bacterianos. En los fagos
descendientes encontraron ADN radiactivo, pero no las proteínas
radiactivas. El segundo experimento confirmó los resultados del
primero: el ADN es la molécula de la herencia.
IDJR
$'1GHOIDJR
EDFWHULD
FURPRVRPD
EDFWHULDQR
H,Z[Y\J[\YHKL\UIHJ[LYP}MHNV
I*PJSVKL]PKHKL\UIHJ[LYP}MHNV
(OIDJRVHDGKLHUH
DXQDEDFWHULD
(OIDJRLQ\HFWD
VX$'1DOD
EDFWHULD
6HUHSOLFDHO
$'1GHOIDJR
3DUWHVGHOIDJRVRQ
VLQWHWL]DGDVSRUPHGLRGHO
PHWDEROLVPRGHODEDFWHULD
6HIRUPDQ
IDJRV
FRPSOHWRV
6HGHVWUX\HOD
SDUHGGHODEDFWHULD
ORVIDJRVTXHGDQOLEUHV
$'1
FDEH]D
FROD
䊱
FIGURA E11-1 Bacteriófagos
(a) Muchos bacteriófagos tienen
una estructura compleja que
comprende una cabeza con
material genético y fibras caudales
con las que se adhieren a la
superficie de una bacteria, así
como un elaborado aparato para
inyectar en la bacteria su material
genético. (b) Ciclo de vida de un
bacteriófago. El bacteriófago usa
el metabolismo de la bacteria para
producir más fagos.
204
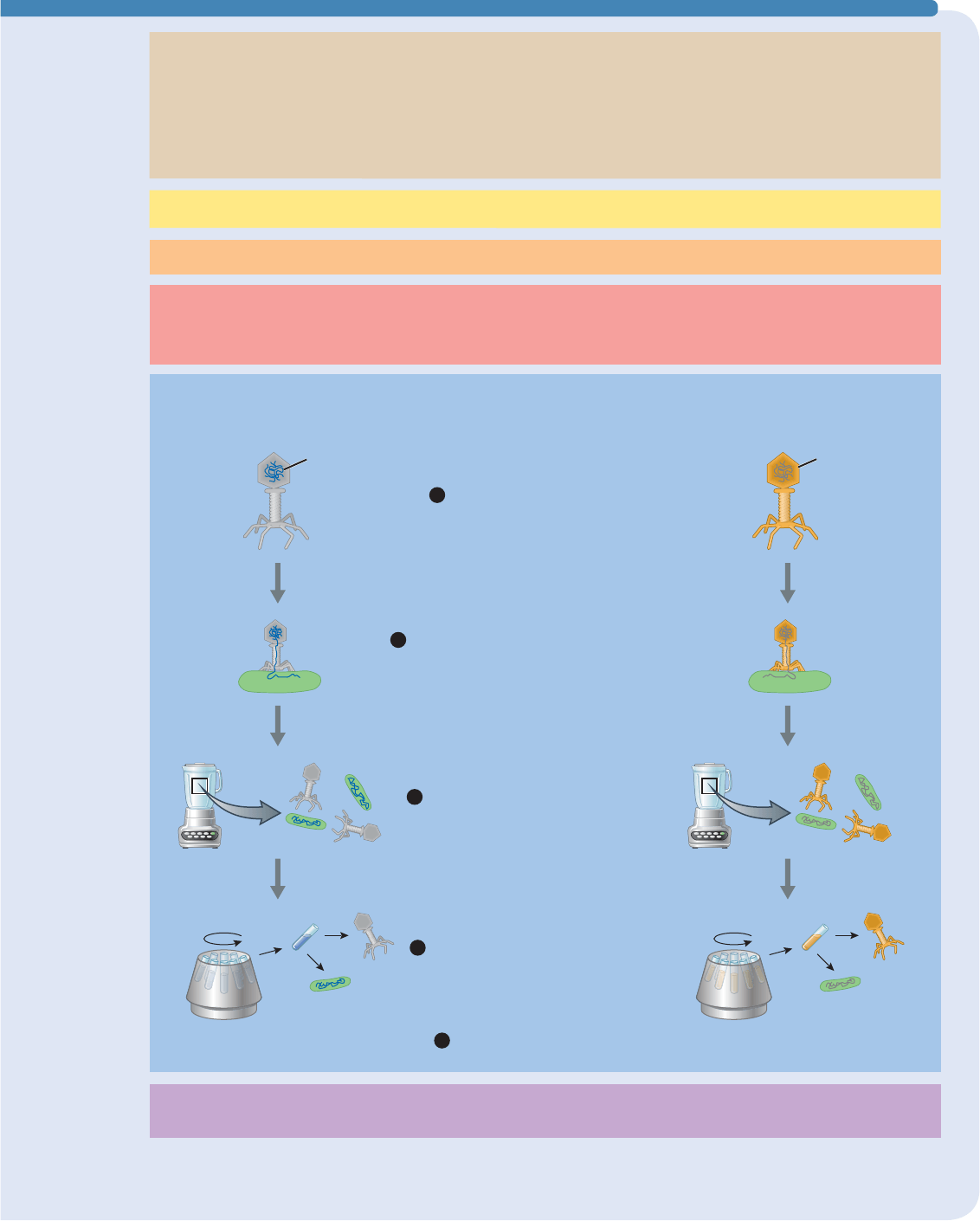
䊱
FIGURA E11-2 Experimento de Hershey-Chase Al marcar radiactivamente el ADN o las proteínas
de bacteriófagos, Hershey y Chase comprobaron si el material genético de los fagos está en el ADN (lado
izquierdo del experimento) o en las proteínas (lado derecho).
Se marcan los fagos con
32
P o
35
S
Se infectan las bacterias con los fagos marcados;
los fagos inyectan su material genético a las bacterias
Se mide la radiactividad de la
envoltura de los fagos y las bacterias
Se centrifuga para separar la envoltura
de los fagos (baja densidad: se quedan en
el líquido) de las bacterias (alta densidad:
se hunden al fondo como “gránulos”)
Se agita en una mezcladora para separar
de las bacterias la envoltura de los fagos
Resultados: las bacterias son
radiactivas: la envoltura de los fagos, no
Conclusión:
Las bacterias infectadas quedan marcadas con fósforo radiactivo, pero no con azufre radiactivo, lo
que apoya la hipótesis de que el material genético de los bacteriófagos es el ADN, no las proteínas.
Observaciones:
1. Los virus bacteriófagos constan únicamente de ADN y proteínas
2. Los bacteriófagos inyectan el material genético a las bacterias y las obligan
a sintetizar más fagos
3. La envoltura exterior de los bacteriófagos se queda fuera de las bacterias
4. El ADN contiene fósforo pero no azufre
a. El ADN se puede marcar con fósforo radiactivo
5. Las proteínas contienen azufre, pero no fósforo
a. Las proteínas se pueden marcar con azufre radiactivo
Pregunta:
¿El ADN o las proteínas son el material genético de los bacteriófagos?
Hipótesis:
Fósforo radiactivo (
32
P)
Resultados: las envolturas de los fagos
son radiactivas; las bacterias, no
Azufre radiactivo (
35
S)
ADN radiactivo
(azul)
Proteína
radiactiva
(dorada)
El ADN es el material genético.
Predicción:
1. Si las bacterias son infectadas por los bacteriófagos que contienen ADN marcado
radiactivamente, se volverán radiactivas
2. Si las bacterias son infectadas con bacteriófagos que contienen proteínas marcadas
radiactivamente, no se volverán radiactivas
Experimento:
2
3
4
5
1
205

206 UNIDAD 2 Herencia
botaban los rayos en la molécula de ADN (FIGURA 11-4a). Como
se ve, el patrón de “difracción” no suministra una imagen directa
de la estructura del ADN. Sin embargo, expertos como Wilkins y
Franklin (
FIGURA 11-4b,c) supieron extraer del patrón mucha in-
formación sobre el ADN. En primer lugar, una molécula de ADN
es larga y delgada, con un diámetro uniforme de unos dos nanó-
metros (dos mil millonésimas de metro). Segundo, el ADN es he-
licoidal, es decir, está torcido como un sacacorchos o una escalera
de caracol. Tercero, la molécula del ADN consta de unidades que
se repiten.
Los datos químicos y de difracción de rayos X no propor-
cionaron suficiente información a los investigadores para dilucidar
la estructura del ADN; también hicieron falta algunas conjeturas
afortunadas. Al combinar los datos de Wilkins y Franklin con un
conocimiento de cómo se unen las moléculas orgánicas complejas
y la intuición de que “los objetos biológicos importantes se pre-
sentan en pares”, James Watson y Francis Crick propusieron un
modelo de la estructura del ADN (véase la sección “Investigación
científica: El descubrimiento de la doble hélice” en la página 208).
Propusieron que la molécula de ADN consta de dos polímeros de
nucleótidos enlazados llamados hebras (
FIGURA 11-5). En cada
hebra de ADN, el grupo fosfato de un nucleótido se une con el
azúcar del siguiente nucleótido en la misma hebra. Esta sucesión
de enlaces produce una “columna vertebral” en que alternan azú-
cares y fosfatos unidos por enlaces covalentes. De esta columna de
azúcar y fosfato se proyectan bases de nucleótidos.
Todos los nucleótidos de una hebra de ADN tienen la mis-
ma orientación; por tanto, los dos extremos de la hebra son dife-
rentes: un extremo tiene un azúcar “libre”, es decir, que no está
enlazada, y el otro extremo tiene un fosfato “libre”, sin enlazar
(véase la figura 11-5a). Imagínate una fila larga de automóviles
detenidos una noche en una concurrida calle de un sentido. Los
faros delanteros de los coches (fosfatos libres) apuntan siempre al
frente y las luces traseras (azúcares libres) apuntan hacia atrás. Si
los coches están muy embotellados, un peatón situado adelante
de la fila sólo vería los faros delanteros del primer vehículo, pero
si estuviera situado atrás, sólo vería las luces traseras del último.
Enlaces de hidrógeno entre bases complementarias
mantienen unidas las dos cadenas de ADN en la
doble hélice
Watson y Crick propusieron que dos hebras de ADN se mantienen
unidas por enlaces de hidrógeno que se forman entre las bases que
se proyectan de cada hebra (véase la figura 11-5a). Estos enlaces con-
fieren al ADN la estructura de escalera, con las columnas de azúcar y
fosfato en la parte exterior (formando los verticales de la escalera)
y las bases de nucleótidos en el interior (haciendo los escalones de
la escalera). Ahora bien, las hebras del ADN no son rectas, sino que
giran una alrededor de la otra de modo que forman una doble hé-
lice que asemeja una escala doblada a lo largo, con el aspecto de
una escalera de caracol (véase la
FIGURA 11-5b). Además, las dos
hebras de la doble hélice de ADN están orientadas en direcciones
opuestas o antiparalelas. En el diagrama de la figura 11-5a, observa
que la hebra a mano izquierda tiene un grupo fosfato libre en la
parte superior y un azúcar libre en la inferior, mientras que ocurre
lo contrario en la hebra a mano derecha. Imagina de nuevo el em-
botellamiento, pero esta vez en una calle congestionada de doble
sentido. El piloto de un helicóptero de tránsito que sobrevolara la
calle vería únicamente los faros delanteros de los automóviles de un
carril y las luces traseras de los vehículos del carril contrario.
Veamos más de cerca los pares de bases que forman los
peldaños de la escalera de la doble hélice. La adenina forma en-
laces de hidrógeno únicamente con la timina, y la guanina forma
enlaces de hidrógeno sólo con la citosina (véanse las figuras 11-5a
y 11-5b). Estos pares A-T y C-G se llaman pares de bases com-
plementarias. Todas las bases de las dos hebras de una doble hé-
lice de ADN son complementarias unas de las otras. Por ejemplo,
si una hebra está organizada A-T-T-C-C-A-G-G-C-T, la otra hebra
debe ir T-A-A-G-G-T-C-C-G-A.
(
a
)
Patrón de difracción del ADN
(
b
)
Maurice Wilkins
(
c
)
Rosalind Franklin
䊱
FIGURA 11-4 Estudios del ADN por difracción de rayos X a) La X formada por manchas oscuras es
característica de las moléculas helicoidales, como el ADN. Las mediciones de varios aspectos del patrón indican las
dimensiones de la doble hélice; por ejemplo, la distancia entre las manchas oscuras corresponde a la distancia entre
los giros de la hélice. (b) Maurice Wilkins y (c) Rosalind Franklin descubrieron muchas características del ADN por
medio del examen meticuloso de los patrones de difracción de los rayos X. Wilkins compartió el premio Nobel de
Fisiología y Medicina con Watson y Crick en 1962. Franklin murió en 1958. Como los premios Nobel no se conceden
póstumamente, muchas veces las aportaciones de Franklin no reciben el reconocimiento que se merecen.
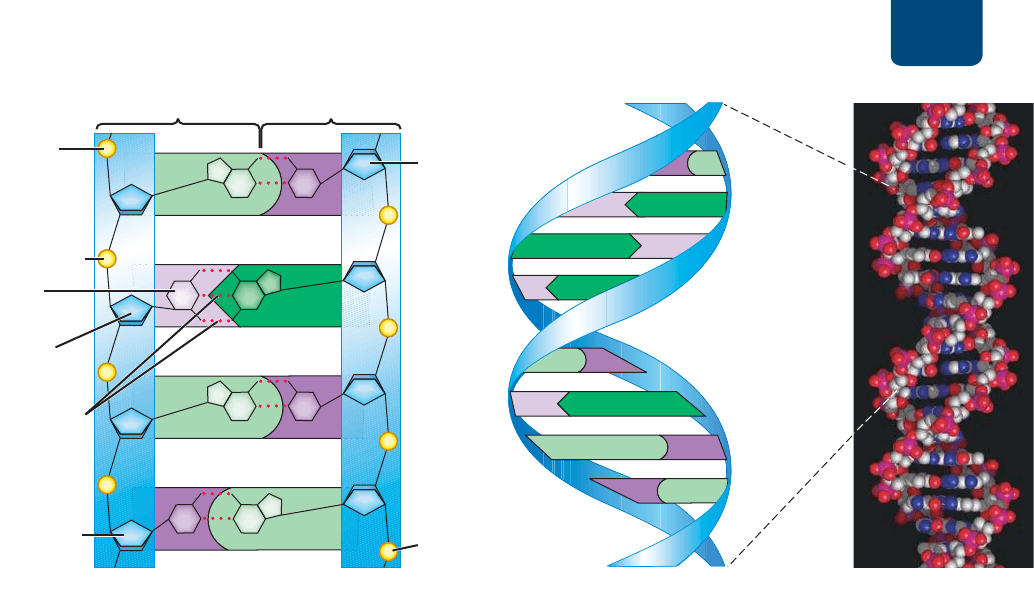
ADN: la molécula de la herencia Capítulo 11 207
fosfato
libre
fosfato
enlaces de
hidrógeno
azúcar
libre
fosfato
libre
nucleótido nucleótido
base
(citosina)
azúcar
azúcar
libre
TA
C
(a) Enlaces de hidrógeno sostienen las bases de pares
complementarios del ADN
(b) Las dos hebras del ADN forman
una doble hélice
(c) Cuatro giros de la
doble hélice de ADN
G
GC
C
G
T
T
T
A
A
A
C
A
A
T
T
G
CG
AT
䊱
FIGURA 11-5 El modelo de Watson y Crick de la estructura del ADN (a) Enlaces de hidrógeno
entre pares de bases complementarias unen las dos hebras del ADN. Tres enlaces de hidrógeno unen la
guanina con la citosina y dos enlaces unen la adenina con la timina. Observa que cada hebra tiene un fosfato
libre en un extremo y un azúcar libre en el extremo opuesto. Además, las dos hebras corren en direcciones
opuestas. (b) Las hebras del ADN se enrollan una sobre la otra en una doble hélice, como una escalera de
caracol, con la columna de fosfato y azúcar que forma las verticales y los pares de bases complementarias,
los peldaños. (c) Modelo volumétrico de la estructura del ADN.
PREGUNTA ¿Qué crees que sería más difícil de romper, un par de bases A-T o uno C-G?
Los pares de bases complementarias explican la “regla de
Chargaff: que el ADN de una especie contiene cantidades iguales
de adenina y timina, así como cantidades iguales de citosina y
guanina. Como una A de una hebra de ADN se empareja siempre
con una T de la otra hebra, la cantidad de A es igual a la cantidad
de T. Del mismo modo, como una G de una hebra se une siempre
con una C de la otra hebra de ADN, la cantidad de G siempre es
igual a la cantidad de C.
Por último, observa el tamaño de las bases. Como la ade-
nina y la guanina constan de dos anillos fusionados, son gran-
des, mientras que la timina y la citosina, formadas por un solo
anillo, son pequeñas. Como la doble hélice tiene únicamente
pares A-T y G-C, todos los peldaños de la escalera del ADN tie-
nen el mismo ancho; por tanto, la doble hélice tiene un diáme-
tro constante, como lo había predicho el patrón de difracción
de rayos X.
La estructura del ADN quedó dilucidada. El 7 de marzo de
1953, en el Bar Eagle de Cambridge, en Inglaterra, Francis Crick
proclamó ante la multitud que se había reunido para comer:
“Descubrimos el secreto de la vida”. Esta afirmación no estaba le-
jos de la verdad. Aunque se necesitaban más datos para confirmar
los detalles, en apenas unos años el modelo del ADN revolucio-
nó la biología, incluidas la genética, la evolución y la medicina.
Como veremos en capítulos posteriores, la revolución continúa
al día de hoy.
11.3
¿CÓMO CODIFICA LA INFORMACIÓN
EL ADN?
Volvamos a la estructura del ADN que se muestra en la figura 11-5.
¿Ves por qué tantos científicos tenían problemas para creer que el
ADN pudiera ser el portador de la información genética? Piensa en
todas las características de un solo organismo. ¿Cómo es posible
que el color de las plumas de un pájaro, el tamaño y la forma de
su pico, su habilidad para construir un nido, su canto y su capaci-
dad de emigrar estén determinados por una molécula con cuatro
unidades simples?
La respuesta es que lo importante no es el número de uni-
dades, sino la secuencia. En una hebra de ADN, las cuatro bases
pueden disponerse en cualquier orden, y cada peculiar secuencia
de bases representa un conjunto único de instrucciones genéticas.
Una analogía será útil para comprender esto. No se necesitan mu-
chas letras para conformar una lengua. El idioma castellano tiene
27 letras, el hawaiano tiene 12 y el lenguaje binario de las compu-
tadoras tiene sólo dos “letras” (0 y 1, o “apagado” y “encendido”).
Sin embargo, los tres idiomas pueden formar miles de palabras di-
ferentes. Un tramo de ADN de sólo 10 nucleótidos tiene más de un
millón de secuencias posibles de las cuatro bases. Como un orga-
nismo tiene desde millones de nucleótidos (en las bacterias) hasta
miles de millones (en las plantas o los animales), las moléculas del
ADN pueden codificar una cantidad asombrosa de información.

208 UNIDAD 2 Herencia
Investigación científica
El descubrimiento de la doble hélice
A comienzos de la década de 1950, muchos biólogos se dieron
cuenta de que la clave para entender la herencia se encontraba
en la estructura del ADN. También sabían que quien dedujera
la estructura correcta del ADN recibiría reconocimientos que
quizá incluyeran el premio Nobel. Linus Pauling, del California
Institute of Technology (Instituto de Tecnología de California),
era la persona que más probabilidades tenía de resolver el
misterio de la estructura del ADN. Es de creer que Pauling sabía
más que cualquier otra persona con vida acerca de la química
de las moléculas orgánicas grandes. Como Rosalind Franklin y
Maurice Wilkins, Pauling era experto en técnicas de difracción
de rayos X. En 1950 se valió de estas técnicas para mostrar
que muchas proteínas estaban enrolladas en hélices de una
hebra (véase la figura 3-20b). Pero Pauling tenía dos grandes
desventajas. En primer lugar, durante años se había concentrado
en la investigación de las proteínas y, por tanto, tenía pocos
datos sobre el ADN. En segundo lugar, participaba activamente
en el movimiento pacifista. En ese entonces, algunos funcionarios
del gobierno estadounidense consideraban que tales actividades
podían ser subversivas y amenazar la seguridad de aquel país.
Esta segunda desventaja resultaría decisiva.
Los siguientes competidores con más probabilidades eran
Wilkins y Franklin, los científicos ingleses que se habían dado a la
tarea de determinar la estructura del ADN mediante patrones de
difracción de rayos X. De hecho, eran los únicos científicos que
tenían buenos datos sobre la forma general de la molécula de ADN.
Por desgracia para ellos, su abordaje metodológico era lento.
La puerta se abrió para los que finalmente descubrieron la doble
hélice: James Watson y Francis Crick, dos científicos que no tenían
los extensos conocimientos de los enlaces químicos que tenía
Pauling ni la experiencia de Franklin y Wilkins en el análisis por
rayos X. Watson y Crick no experimentaban en el sentido ordinario
de la palabra, sino que pasaban el tiempo pensando en el ADN,
mientras trataban de construir un modelo molecular que fuera
lógico y en el que se acomodaran los datos. Como trabajaban en
Inglaterra y como Wilkins les mostró los datos de Franklin (quizá
contra sus deseos), Watson y Crick estaban familiarizados con toda
la información radiográfica relacionada con el ADN.
Los datos de rayos X eran lo que le hacía falta a Pauling.
En virtud de sus tendencias supuestamente subversivas, el
Departamento de Estado de Estados Unidos se negó a expedir un
pasaporte para que Pauling pudiera salir de dicho país, así que
no pudo asistir a las reuniones en las que Wikins presentó los
datos de rayos X ni fue a Inglaterra a hablar directamente con
Wilkins y Franklin. Watson y Crick sabían que Pauling trabajaba
en la estructura del ADN y los impulsaba el miedo de que pudiera
vencerlos. En su libro The Double Helix (La doble hélice), Watson
cuenta que estaba convencido de que Pauling había visto las
imágenes de rayos X, de modo que “cuando mucho en una
semana, Linus habría desentrañado la estructura”.
Quizá estás pensando que eso no es justo, que la meta de
la ciencia es avanzar en el conocimiento y que todos deberían
tener acceso a todos los datos. Tal vez, pero después de todo,
los científicos también son personas. Prácticamente todos los
científicos quieren ver que su disciplina progresa y que derrama
beneficios a la humanidad, pero a la vez cada individuo quiere ser
el factor principal de ese progreso y recibir el crédito y la fama.
Linus Pauling se quedó a oscuras sobre los datos de los rayos X y
fue abatido en la carrera por encontrar la estructura correcta.
Poco después de que Watson y Crick propusieran la doble
hélice (FIGURA E11-3), Watson la describió en una carta a Max
Delbruck, amigo y consejero del California Institute of Technology
(Instituto de Tecnología de California). Cuando Delbruck le contó
a Pauling sobre el modelo de la doble hélice para el ADN, Pauling
felicitó gentilmente a Watson y Crick por su brillante solución. La
carrera había terminado.
Estudio de caso continuación
Músculos, mutaciones y miostatina
Todos los mamíferos “normales” tienen una secuencia de ADN
que codifica una proteína miostatina funcional que limita el
crecimiento de los músculos. El ganado Belgian Blue tiene
una mutación que cambia un gen leve por uno absurdo que
ya no codifica una proteína funcional, de modo que sufre un
desarrollo muscular excesivo.
Desde luego, para que tengan algún sentido, las letras de un
idioma deben estar en el orden correcto. Del mismo modo, un gen
debe tener las bases indicadas, en la secuencia precisa. Así como
“huésped” y “hostil” significan cosas diferentes y “huopli” no quie-
re decir nada, diferentes secuencias de bases del ADN codifican
información muy diferente o no codifican ninguna información.
En el capítulo 12 vamos a explicar cómo se toma la infor-
mación del ADN para formar la estructura de las células vivas.
En el resto del capítulo, examinaremos cómo se replica el ADN
durante la división celular para que se copie correctamente esta
información genética.
䊱
FIGURA E11-3 El descubrimiento del ADN James
Watson y Francis Crick con un modelo de la estructura del ADN.
ADN: la molécula de la herencia Capítulo 11 209
Estudio de caso continuación
Músculos, mutaciones y miostatina
Gracias a un complejo mecanismo que abarca muchas otras
moléculas, la miostatina evita que las células premusculares
repliquen su ADN. Por consiguiente, las células dejan de
dividirse y se limita el número de células maduras. La
miostatina mutada del ganado Belgian Blue no inhibe
la replicación del ADN, así que las células premusculares
siguen dividiéndose y producen mayor masa muscular.
11.4
¿CÓMO ES QUE LA REPLICACIÓN
DEL ADN GARANTIZA LA CONSTANCIA
GENÉTICA DURANTE LA DIVISIÓN CELULAR?
La replicación del ADN es un acontecimiento
fundamental en el ciclo celular
En 1850, el patólogo austriaco Rudolf Virchow se daba cuenta de
que “todas las células vienen de células”. Los billones de células
de tu cuerpo son descendientes de otras células, que se remontan
al momento en que fuiste un óvulo fecundado. Además, casi todas
las células de tu cuerpo contienen información genética idéntica, la
misma información presente en el óvulo fecundado. Cuando las
células se reproducen por división mitótica, cada célula hija reci-
be una copia casi perfecta de la información genética de la célula
original. Por consiguiente, antes de la división, la célula original
debe sintetizar dos copias exactas de su ADN. Un proceso llamado
replicación del ADN produce estas dos dobles hélices idénticas.
La replicación del ADN produce dos dobles
hélices de ADN, cada una con la hebra
original y una nueva
¿Cómo copia exactamente la célula su ADN? En el documento en
el que describieron la estructura del ADN, Watson y Crick escribie-
ron una de las frases más insuficientes de la ciencia: “No se nos ha
es ca pa do que el emparejamiento específico [de las bases] que he-
mos postulado sugiere inmediatamente un posible mecanismo de
copia del material genético”. De hecho, el emparejamiento de las
bases es la base de la replicación del ADN. Recuerda que las reglas
para el emparejamiento de bases son que la adenina de una hebra
debe unirse con una timina en la otra hebra y que la citosina debe
emparejarse con una guanina. Si una de las hebras tiene, por ejem-
plo, A-T-G, la otra hebra debe tener T-A-C. Por tanto, la secuencia
de bases de cada hebra contiene toda la información necesaria para
replicar la otra hebra.
En teoría, la replicación del ADN es bastante simple (
FIGU-
RA 11-6
). Los componentes esenciales son: (1) las hebras de ADN
originales, (2) nucleótidos libres sintetizados previamente en el
citoplasma e introducidos en el núcleo, y (3) diversas enzimas
que desenrollan y abren la doble hélice de ADN original y que
sintetizan nuevas hebras de ADN.
Primero, las enzimas llamadas ADN helicasas (lo que sig-
nifica “enzimas que separan la doble hélice”) abren la doble hé-
lice de ADN original, de modo que las bases de las dos hebras de
ADN ya no forman pares de bases una con la otra. Hay que sinteti-
(;
(;
(
(
;
;
(;
(;
(;
(;
(; ;
;
;
(;
;
;
(;
(
((
;
(
(;
(;
*
*
*
.
*.
*
*.
.
*.
.
*.
*.
.*.
*
*
.
.
*.
.
*.
*.
(
(
;
(;
(;
*.
*.
*.
.
.
QXFOHʼnWLGRVOLEUHV
(O$'1RULJLQDOR
SDUHQWDOVHGHVHQUROOD
6HVLQWHWL]DQ
QXHYDVKHEUDVGH$'1
FRQODVEDVHV
FRPSOHPHQWDULDVGH
ODVKHEUDVRULJLQDOHV
RSDUHQWDOHV
&DGDQXHYDGREOHKÒOLFH
HVWÀFRPSXHVWDSRUXQD
KHEUDRULJLQDORSDUHQWDOD]XO
\XQDQXHYDKHEUDKLMDURMR
'REOHKÒOLFH
GH$'1SDUHQWDO
䊱
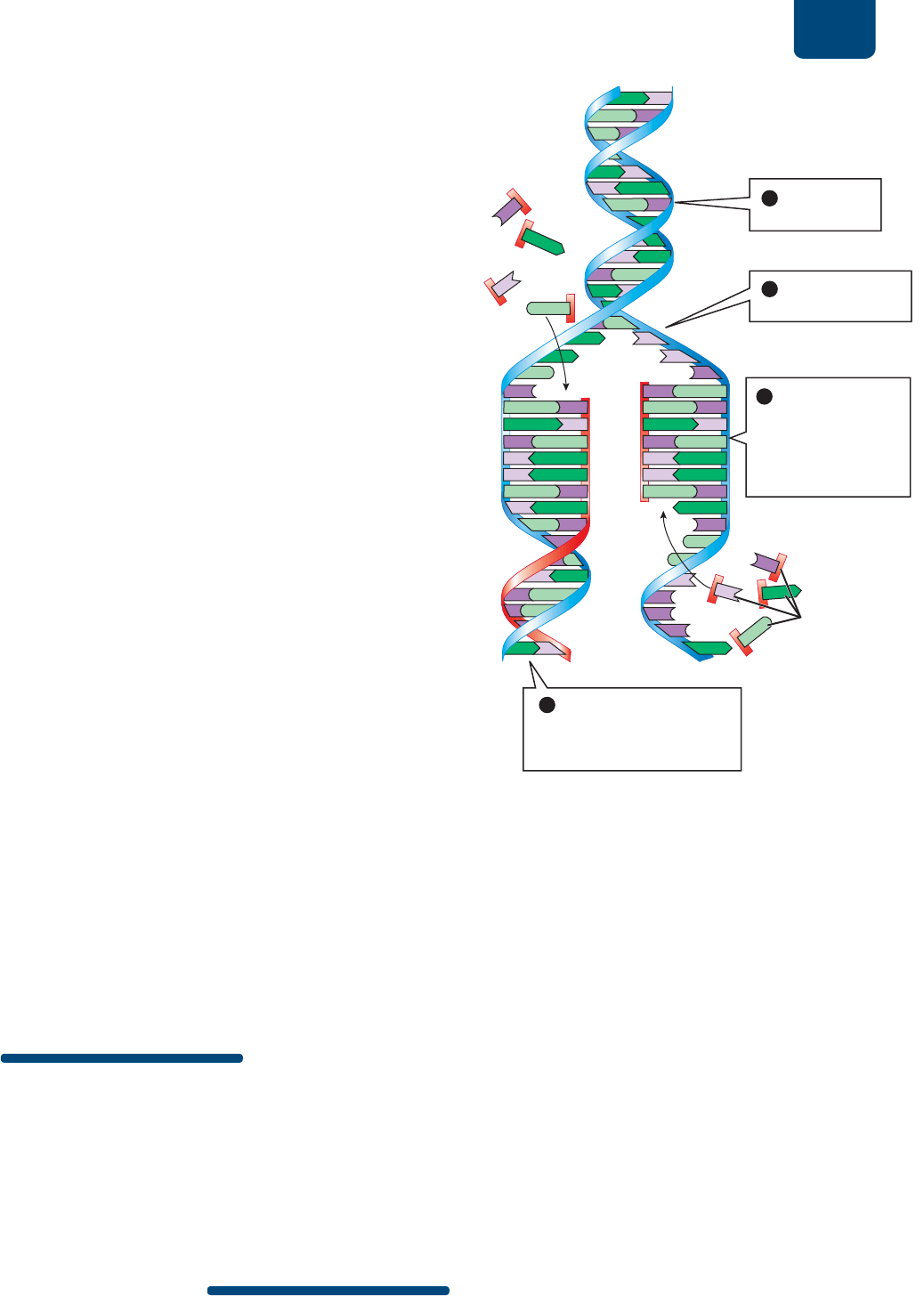
FIGURA 11-6 Elementos básicos de la replicación del
ADN En la replicación se separan las dos hebras de la doble hélice
del ADN parental. Los nucleótidos libres que son complementarios
de los que se encuentran en cada hebra se unen para hacer hebras
hijas. Cada hebra original y su nueva hebra hija forman una nueva
doble hélice.
zar hebras complementarias de las originales. Otras enzimas, lla-
madas ADN polimerasas (“enzimas que sintetizan un polímero
de ADN”), avanzan por cada hebra separada del ADN original y
emparejan sus bases con los nucleótidos libres complementarios.
Por ejemplo, la ADN polimerasa empareja una adenina expuesta
en la hebra original con una timina. La ADN polimerasa también
conecta estos nucleótidos libres unos con otros para formar dos
nuevas hebras de ADN, cada una complementaria de una de las
hebras del ADN original. Así, si una hebra de ADN original lleva
T-A-G, la ADN polimerasa sintetiza una nueva hebra de ADN con
la secuencia complementaria A-T-C. Para más información sobre
cómo se replica el ADN, consulta el apartado “De cerca: Estructu-
ra y replicación del ADN” en las páginas 210-212.
Al terminar la replicación, una hebra del ADN original y
su hebra de ADN hija recién formada se enredan en una doble
hélice. Al mismo tiempo, la otra hebra original y su hebra hija
se enredan en una segunda doble hélice. Al formar las nuevas
dobles hélices, la replicación del ADN conserva una hebra del

210 UNIDAD 2 Herencia
De cerca Estructura y replicación del ADN
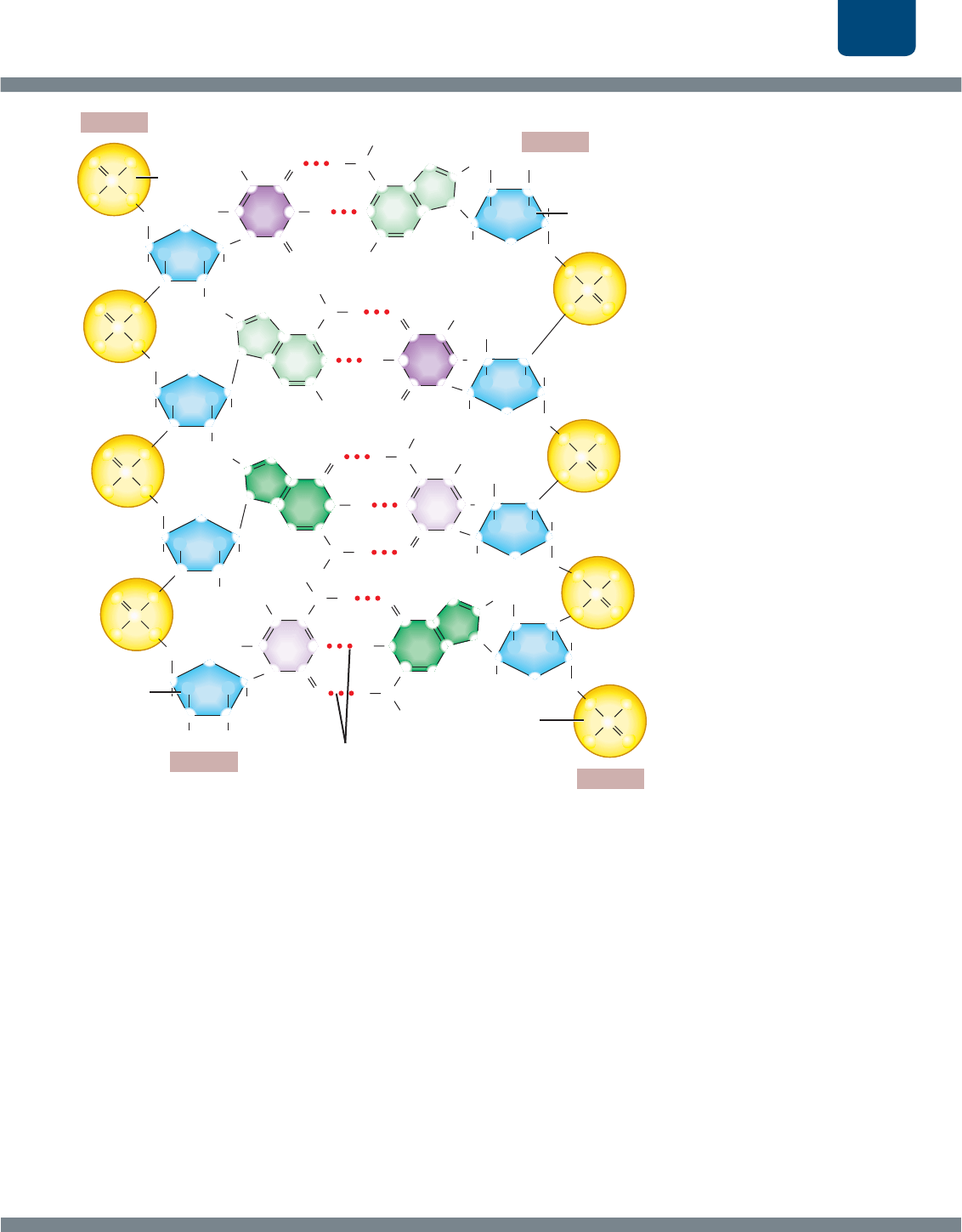
Estructura del ADN
Para entender la replicación del ADN, tenemos que regresar a
su estructura. Los bioquímicos siguen el rastro de los átomos
de una molécula compleja numerándolos. En el caso de un
nucleótido, los átomos que forman las “esquinas” de la base
están numerados del 1 al 6 para citosina y timina que tienen
un solo anillo, y del 1 al 9 para adenina y guanina que
llevan un anillo doble. Los átomos de carbono del azúcar
se numeran del 1 al 5. Se usa el símbolo de prima () para
distinguir los átomos del azúcar de los átomos de la base. Los
carbonos del azúcar se llaman “1 prima” a “5 prima” (
FIGURA
E11-4
).
El azúcar de un nucleótido tiene dos “extremos” que
pueden participar en la síntesis de la columna de azúcar
y fosfato de la hebra de ADN, un extremo 3 que tiene un
grupo libre —OH (hidroxilo) unido al carbono 3 del azúcar,
y un extremo 5 que tiene un grupo fosfato unido al carbono
5. Cuando se sintetiza una hebra de ADN, el fosfato de
un nucleótido se une con el grupo hidroxilo del azúcar del
siguiente nucleótido (
FIGURA E11-5).
Con esto queda todavía un grupo hidroxilo libre en el
extremo 3 de un nucleótido y un grupo fosfato libre en
el extremo 5 del otro nucleótido. No importa cuántos
nucleótidos se unan, siempre queda un hidroxilo libre en el
extremo 3 de la hebra y un fosfato libre en el extremo 5.
Las columnas de azúcar y fosfato de las dos hebras de
una doble hélice son antiparalelas, lo que significa que corren
en direcciones opuestas. Por tanto, en un extremo de una
doble hélice una hebra tiene un hidroxilo libre en el azúcar (el
extremo 3), mientras que la otra hebra tiene un fosfato libre
(el extremo 5). En el otro extremo de la doble hélice, los
extremos de la hebra están invertidos (
FIGURA E11-6).
Replicación del ADN
La replicación del ADN consta de tres acciones principales
(
FIGURA E11-7). En primer lugar, la doble hélice del ADN
debe abrirse para que sea posible interpretar la secuencia de
bases. Enseguida, es necesario sintetizar nuevas hebras
de ADN con la secuencia de bases complementaria de las
dos hebras originales. En las células eucariontes, estas
nuevas hebras de ADN se sintetizan en secciones muy cortas.
Por tanto, el tercer paso de la replicación del ADN es unir las
secciones para formar una nueva hebra continua de ADN. Cada
paso es realizado por un conjunto diferente de enzimas.
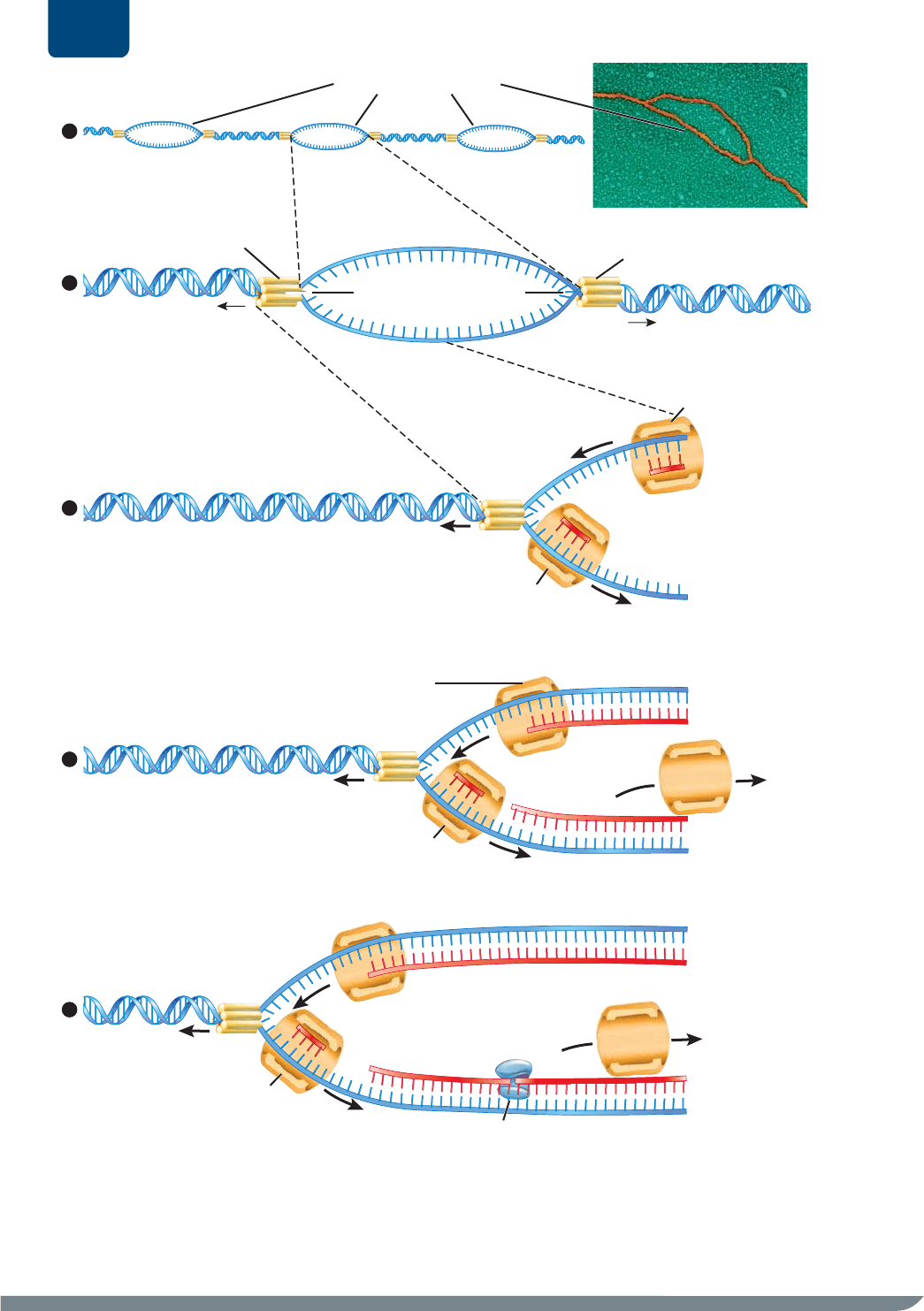
La ADN helicasa separa las hebras
del ADN original o parental
En sintonía con otras enzimas, la ADN helicasa rompe los
enlaces de hidrógeno entre pares de bases complementarias
que mantienen unidas las dos hebras del ADN original.
Con esto se separa y desenrolla la doble hélice original y
se forma una “burbuja” de replicación (
FIGURA E11-7 ❶
y ❷). La burbuja de replicación contiene una “horquilla” de
replicación en cada extremo, donde las dos hebras del ADN
original apenas comienzan a desenrollarse. En la burbuja de
replicación, las bases de las hebras del ADN original dejan
de estar emparejadas.
La ADN polimerasa sintetiza nuevas hebras de ADN
Las burbujas de replicación son esenciales porque permiten
que una segunda enzima, la ADN polimerasa, pase a las bases
de cada hebra de ADN (
FIGURA E11-7 ❸). En cada horquilla de
replicación, un complejo de ADN polimerasa y otras proteínas
se unen a cada hebra original. Por tanto, habrá dos complejos
de ADN polimerasa, uno en cada hebra original. La ADN
polimerasa reconoce una base sin par de la hebra original
y la une a una base complementaria en un nucleótido libre.
Luego, la ADN polimerasa cataliza la formación de nuevos
enlaces covalentes que unan el fosfato del nucleótido libre
nuevo (el extremo 5) con el azúcar del nucleótido añadido
más recientemente (el extremo 3) de la hebra hija que va
creciendo. De esta manera, la ADN polimerasa sintetiza la
columna de azúcar y fosfato de la hebra hija.
¿Por qué se forman burbujas de replicación en lugar de
simplemente empezar en un extremo de la doble hélice y
H
O
H
O
O
P
O
O
O
H
C
H
C
H
C
OH
C
CH
3
NC
CC
N7C
H
H
CH
2
O
䊱
FIGURA E11-4 Numeración de los átomos de
carbono en un nucleótido
H
O
H
O
O
P
O
O
O
H
C
H
C
H
CC
CH
3
NC
CC
NTC
H
H
CH
2
extremo 5
O
5
4
3 2
1
1
5
2
4
63
H
H
N
H
O
P
O
O
O
H
C
H
C
H
C
OH
C
NC
CC
N
N
N
AC
C
H
H
H
CH
2
O
5
4
3 2
1
3
5
7
8
9
2
6
41
extremo 3
Enlace entre el
azúcar y el
nucleótido
superior y entre el
fosfato y el
nucleótido inferior
䊱
FIGURA E11-5 Numeración de los átomos de carbono
en un dinucleótido
ADN: la molécula de la herencia Capítulo 11 211
dejar que una molécula de ADN polimerasa copie el ADN de una
sección continua hasta el otro extremo? Bueno, los cromosomas
eucariontes son muy largos: los cromosomas humanos van de
unos 23 millones de bases en el diminuto cromosoma Y a unos
246 millones de bases en el cromosoma 1. El ADN eucarionte se
copia a un ritmo de unos 50 nucleótidos por segundo, así que
se necesitarían de cinco a 57 días para copiar los cromosomas
humanos en una sección continua. Para replicar un cromosoma
entero en un tiempo razonable, muchas enzimas de ADN
helicasa abren muchas burbujas de replicación, a modo de que
las enzimas de ADN polimerasa copien las hebras originales en
secciones muy pequeñas. Las burbujas crecen conforme avanza
la replicación del ADN y se unen cuando entran en contacto unas
con otras.
La ADN polimerasa siempre se aleja del extremo 3 de una
hebra de ADN original (es decir, del extremo con el grupo
hidroxilo libre del azúcar). Siempre se agregan nuevos nucleótidos
al extremo 3 de la hebra hija. En otras palabras, la ADN
polimerasa mueve 3 →5 en una hebra
original y, simultáneamente, mueve
5 → 3 en la hebra hija. Por último,
como las dos hebras de la doble hélice
de ADN original están orientadas en
direcciones opuestas, las moléculas de la
ADN polimerasa se mueven igualmente
en sentido contrario en las dos hebras
originales (véase la figura E11-7 ❸).
La ADN helicasa y la ADN polimerasa
colaboran (
FIGURA E11-7 ❹). Una
ADN helicasa “aterriza” en la doble
hélice y la recorre para desenrollarla y
separar las hebras. Como las dos hebras
de ADN corren en direcciones opuestas,
conforme una enzima de ADN helicasa
se mueve hacia el extremo 5 de una
hebra original, avanza simultáneamente
hacia el extremo 3 de la otra hebra
original. Ahora visualicemos dos ADN
polimerasas que se depositan en las
dos hebras separadas de ADN. Una
ADN polimerasa (llamémosla polimerasa
1) puede seguir a la helicasa hacia el
extremo 5 de la hebra original y puede
sintetizar una hebra de ADN hija de
forma continua hasta que entra en otra
burbuja de replicación. Esta hebra
hija de ADN que se sintetiza de forma
continua se llama hebra líder. Ahora
bien, en la otra hebra original, la
polimerasa 2 se aleja de la helicasa:
en ❸ de la figura E11-7, observa que
la helicasa se mueve a la izquierda,
mientras que la ADN polimerasa 2 lo
hace a la derecha. Por tanto, la síntesis
del ADN en esta hebra será discontinua:
la ADN polimerasa 2 sintetizará una
nueva hebra corta de ADN llamada hebra
rezagada, pero entre tanto, la helicasa
sigue su movimiento a la izquierda y
desenrollando más de la doble hélice
(véase figura E11-7 ❹ y ❺). Otras ADN polimerasas (número
3, 4, etc.) deben depositarse en esta hebra y sintetizar más
hebras rezagadas cortas.
La ADN ligasa une los segmentos de ADN
Numerosas ADN polimerasas sintetizan secciones de ADN de
diversa longitud. Cada cromosoma puede formar cientos de
burbujas de replicación. En cada burbuja habrá una hebra líder,
de cientos de miles de pares de nucleótidos, y de docenas
a miles de hebras rezagadas, cada una con alrededor de
100 a 200 pares de nucleótidos. Por tanto, una célula puede
sintetizar millones de secciones de ADN cuando replica un
único cromosoma. ¿Cómo se unen todos estos fragmentos?
Es lo que hace la tercera enzima importante, la
ADN ligasa
(“enzima que une el ADN”;
FIGURA E11-7 ❺). Muchas enzimas
de ADN ligasa zurcen los fragmentos de ADN hasta que la
hebra hija consta de un único polímero largo y continuo de
ADN.
N
N
C
O
P
O
O
O
CC
CC
NC
CC
N 7 C
O
O
O
HH
H
CH
2
O
CC
CC
O
H
H
N
H
NC
CC
N
N
$C
N
C
O
P
O
O
O
O
P
O
O
O
O
P
O
O
O
O
P
O
O
O
CC
CC
NC
CC
N & C
O
H
H
CH
2
O
CC
CC
O
H
N
N
HO
N
H
NC
CC
N*C
O
P
O
O
O
CC
CC
NC
CC
N7C
O
O
HH
H
H
H
extremo 5
extremo 5
extremo 3
O
CC
CC
O
H
H
N
H
H
NC
H
CC
N
N
$ C
extremo 3
N
C
O
P
O
O
O
O
P
O
O
O
CC
C
OH
C
NC
CC
N&C
H
CH
2
O
CC
CC
O
O
H
N
H
NC
CC
N
N
*
C
N
C
HH
H
H
H
HH
H
H
H
H
H
H
H
H
H
H
H
HH
HH
HH
H
H
H
H
H
H
H
H
H
HH
HH
H
H
HH
OH
H
H
H
CH
2
CH
3
CH
3
H
2
C
H
2
C
H
2
C
H
2
C
IRVIDWR
OLEUH
IRVIDWR
OLEUH
HQODFHVGHKLGUʼnJHQR
HQWUHODVEDVHV
D]ŜFDU
OLEUH
D]ŜFDU
OLEUH
䊱
FIGURA E11-6 Las dos hebras de una doble hélice de ADN son antiparalelas
(continúa)
212 UNIDAD 2 Herencia
KRUTXLOODVGHUHSOLFDFLʼnQ
$'1KHOLFDVD
$'1
EXUEXMDVGHUHSOLFDFLʼnQ
$'1KHOLFDVD
$'1SROLPHUDVD
$'1
SROLPHUDVD
OD$'1SROLPHUDVD
FRQWLQŜDSRUODKHEUD
GHO$'1RULJLQDO
$'1
SROLPHUDVD
DOHMDPLHQWRGHOD
$'1SROLPHUDVD
$'1
SROLPHUDVD
DOHMDPLHQWRGHOD$'1
SROLPHUDVD
OD$'1OLJDVDXQHODV
KHEUDVKLMDVGH$'1
F
R
Q
W
L
Q
X
D
V
ķ
Q
W
H
V
L
V
V
ķ
Q
W
H
V
L
V
G
L
V
F
R
Q
W
L
Q
X
D
V
ķ
Q
W
H
V
L
V
F
R
Q
W
L
Q
X
D
V
ķ
Q
W
H
V
L
V
G
L
V
F
R
Q
W
L
Q
X
D
䊱
FIGURA E11-7 Replicación del ADN ❶ Las enzimas de ADN helicasa separan las hebras originales de un cromosoma para formar
burbujas de replicación. ❷ Cada burbuja de replicación consta de dos horquillas de replicación entre las cuales hay hebras de ADN
desenrolladas. ❸ Las enzimas de ADN polimerasa sintetizan nuevas secciones de ADN. ❹ La ADN helicasa avanza por la doble hélice del ADN
original, la desenrolla y ensancha la burbuja de replicación. Las ADN polimerasas de la burbuja de replicación sintetizan hebras de ADN hijas.
❺ La ADN ligasa une los pequeños fragmentos de ADN en una única hebra hija.
PREGUNTA Durante la síntesis, ¿por qué la ADN polimerasa se aleja de la horquilla de replicación en las dos hebras?
ADN: la molécula de la herencia Capítulo 11 213
'RVKÒOLFHVGREOHV
GH$'1LGÒQWLFDV
FDGDXQDFRQXQD
KHEUDRULJLQDOR
SDUHQWDOD]XO\XQD
QXHYDKHEUDKLMDURMD
'REOHKÒOLFH
GH$'1
UHSOLFDFLʼnQGHO$'1
䊱
FIGURA 11-7 Replicación semiconservativa de ADN
ADN original y sintetiza una hebra nueva. Por tanto, el proceso se
llama replicación semiconservativa (
FIGURA 11-7).
Si no se cometieron errores, la secuencia de bases de las dos
hélices dobles de ADN es idéntica a la secuencia de bases de la
doble hélice de ADN original y, desde luego, una a la otra.
DNA Replication (disponible en inglés)
11
.
5
¿CÓMO OCURREN LAS MUTACIONES?
Nada es perfecto, ni siquiera el ADN de tus células. Los cambios
en la secuencia de bases del ADN suele dar por resultado un gen
defectuoso, lo que se llama una mutación. En la mayor parte de
las células, las mutaciones se reducen al mínimo por la replicación
extremadamente precisa del ADN, la “revisión de originales” del
ADN recién sintetizado y la reparación de todos los cambios
del ADN que puedan ocurrir aun cuando no se replica el ADN.
La replicación precisa y la revisión
producen ADN casi sin errores
La especificidad de los enlaces de hidrógeno entre pares de bases
complementarias hace que la replicación del ADN sea muy exacta.
La ADN polimerasa incorpora bases incorrectas aproximadamente
una vez en cada mil a 100 mil pares de bases; sin embargo, las
hebras completas de ADN contienen apenas alrededor de un error
cada 100 a mil millones de pares de bases (en los seres humanos,
es menos de uno por cromosoma por replicación). Esta tasa de
error fenomenalmente baja es obra de una variedad de enzimas
de reparación del ADN que revisan cada hebra hija durante y des-
pués de la síntesis. Por ejemplo, algunas formas de ADN polimera-
sa reconocen un error en el emparejamiento de una base cuando se
comete. Este tipo de ADN polimerasa se detiene, corrige el error y
continúa con la síntesis del ADN.
Los errores pueden ocurrir
Pese a esta sorprendente exactitud, ni los seres humanos ni otros
organismos tienen ADN exento de errores. Además de los raros
errores cometidos durante la replicación normal del ADN, diversas
condiciones ambientales pueden dañar el ADN. Por ejemplo, cier-
tos compuestos químicos (como los que conforman el humo del
cigarro) y algunas formas de radiación (como los rayos ultravioleta
de la luz solar y los rayos X) aumentan la frecuencia de errores en
el emparejamiento de las bases durante la replicación o aun indu-
cen cambios en la composición del ADN entre replicaciones. Casi
todos estos cambios en la secuencia del ADN los arreglan enzimas
reparadoras de las células. Sin embargo, es inevitable que queden
algunos.
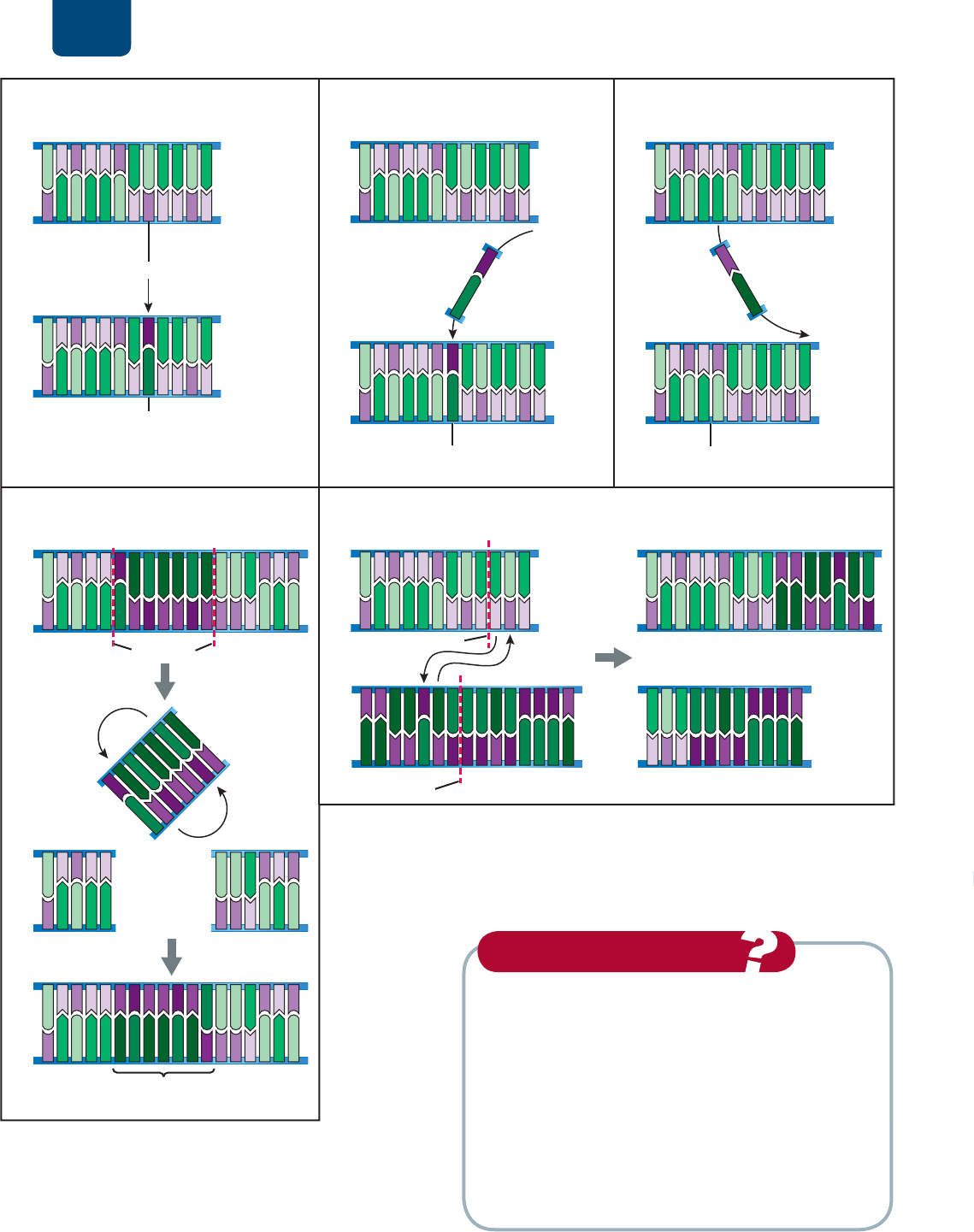
Las mutaciones van de cambios
en pares de nucleótidos simples
a desplazamientos de grandes
piezas de cromosomas
En la replicación, ocasionalmente no concuerda un par de bases.
Por lo general, las enzimas de reparación reconocen el desajuste,
cortan el nucleótido equivocado y lo sustituyen con un nucleóti-
do que lleva una base complementaria. Sin embargo, a veces, las
enzimas reemplazan el nucleótido original en lugar del equivoca-
do. El par de bases que se produce es complementario, pero es in-
correcto. Estas sustituciones de nucleótidos también se llaman
mutaciones puntuales porque cambian nucleótidos individua-
les de la secuencia del ADN (
FIGURA 11-8a). Ocurre una muta-
ción por inserción cuando uno o más pares de nucleótidos se
insertan en la doble hélice de ADN (
FIGURA 11-8b). Se produce
una mutación por supresión cuando se eliminan uno o más
pares de nucleótidos de la doble hélice (
FIGURA 11-8c).
También ocurre que se reorganicen secciones de un cromo-
soma con tamaño variable de un único nucleótido a secciones
enormes del ADN. Ocurre una inversión cuando una sección de
ADN se corta de un cromosoma, se invierte y se reinserta en el es-
pacio (
FIGURA 11-8d). Por último, hay una translocación cuan-
do se remueve un segmento de ADN, por lo regular muy grande,
de un cromosoma y se inserta en otro (
FIGURA 11-8e).
Las mutaciones tienen efectos diversos
en las funciones
Las mutaciones suelen ser nocivas, tal como algunos cambios aza-
rosos a la mitad de Hamlet de Shakespeare interrumpirían el curso
de la obra. Si una mutación es muy perjudicial, es posible que la cé-
lula del organismo que la hereda muera rápidamente. Pero algunas
mutaciones no tienen efecto o, en casos muy raros, son benéficas,
como veremos en el capítulo 12. Las mutaciones que son benéfi-
cas, al menos en ciertos ambientes, pueden ser favorecidas por la
selección natural y son el fundamento de la evolución de la vida en
la Tierra (véase la unidad 3).
214 UNIDAD 2 Herencia
(
;
(
;
*
.
*
.
(
;
*
.
*
.
(
;
(
;*
.
*
.
*
.
(
;
*
.
*
.
(
;
*
.
*
.
(
;
(
;
(
;*
.
*
.
*
.
VHFXHQFLDGH$'1RULJLQDO
H:\Z[P[\JP}UKLU\JSL}[PKVZ
VXVWLWXFLʼnQ
SDUGHQXFOHʼnWLGRVFDPELDGRGH$7D7$
(
;
(
;
*
.
*
.
(
;
*
.
*
.
(
;
(
;*
.
*
.
*
.
(
;
*
.
*
.
(
;
(
;
*
.
*
.
(
;
(
;
(
;
(
;*
.
*
.
*
.
VHFXHQFLDGH$'1RULJLQDO
SDUGHQXFOHʼnWLGRV7$
LQVHUWDGR
I4\[HJP}UWVYPUZLYJP}U
(
;
(
;
*
.
*
.
(
;
*
.
*
.
(
;
(
;*
.
*
.
*
.
(
;
*
.
*
.
(
;
(
;
*
.
*
.
(
;
(
;*
.
*
.
*
.
VHFXHQFLDGH$'1RULJLQDO
SDUGHQXFOHʼnWLGRV&*
HOLPLQDGRV
J4\[HJP}UWVYZ\WYLZP}U
(
;
(
;
*
.
*
.
(
;
*
.
(
;
(
;
*
.
*
.
(
;
(
;*
.
*
.
*
.(
;
(
;*
.
(
;
(
;
(
;
(
;
*
.
(
;
(
;*
.
*
.
*
.
*
.
*
.
*
. (
;
(
;*
.(
;*
.
(
;
(
;
(
;
*
.
(
;*
.
*
.
*
.(
;
(
;*
.
VHFXHQFLDGH$'1RULJLQDO
VHURPSH
VHJPHQWRGH$'1
LQYHUWLGR
K0U]LYZP}U
(
;
*
.
*
.
(
;
*
.
*
.
(
;
(
;
(
;
*
.
*
.
(
;
*
.
*
.
(
;
(
;
*
.
(
;
(
;*
.
*
.
*
.
VHFXHQFLDGH$'1RULJLQDO
L;YHUZSVJHJP}U
VHURPSH
VHURPSH
VHJPHQWRV
GH$'1
LQWHUFDPELDGRV
(
;
(
;
*
.
*
.
*
.*
.
*
.
*
.(
;
(
;
(
;
(
;
(
;
(
;
(
;
(
;
*
.
*
.
*
.
(
;
(
;
(
;*
.
(
;
(
;
(
;
*
.
*
.
*
.
*
.
*
.
(
;
(
;
(
;*
.
*
.
*
.
*
.
*
.
䊱
FIGURA 11-8 Mutaciones (a) Sustitución de nucleótidos.
(b) Mutación por inserción. (c) Mutación por supresión. (d) Mutación
por inversión. (e) Translocación de secciones de ADN entre dos
cromosomas diferentes. En las partes (a) a (d), las bases del ADN
original se muestran con colores pálidos y letras negras; las
mutaciones aparecen con colores oscuros y letras blancas.
¿Te has preguntado…
cuánto influyen los genes en las capacidades
deportivas?
Acéptalo: nunca vas a nadar como Michael Phelps. Pero, ¿cuánto
de esta fantástica capacidad es hereditaria? En algunos casos,
es evidente que los genes marcan una enorme diferencia.
Por ejemplo, las mutaciones de la miostatina pueden acelerar
la velocidad y acrecentar la fuerza, como explicamos en el
apartado “Estudio de caso otro vistazo: Músculos, mutaciones
y miostatina”. Sin embargo, el efecto de la mayor parte de
los genes es pequeño. Más de 240 genes contribuyen al
rendimiento deportivo. ¿Los superdeportistas tienen los mejores
“alelos atléticos”? ¿Cuántos alelos atléticos tiene Phelps y
cuántos tienes tú? Por ahora, nadie lo sabe.
Este documento contiene más páginas...
Descargar Completo
Transmision sináptica215.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.