
Fisiología Renal
www.nefrologiaaldia.org.es
1
INDICE
1. Generalidades sobre la función renal
2. Caracteristicas morfo-funcionales de los riñones.
la nefrona
3. Fisiología renal y mantenimiento del ambiente
interno.
4. Formación de la orina. ltrado glomerular
4.1 Proceso de ltración
4.2 Presiones que afectan al ltrado
glomerular.
4.3 Regulación del ltrado glomerular
.4 Tasa de ltración glomerular
5. Formación de la orina. intercambio tubular
5.1 Mecanismos de reabsorción y secreción
tubulares.
5.2 Transporte en el túbulo contorneado distal
y conducto colector
5.3 Secreción de substancias en la formación
de la orina
6. Funcion endocrina del riñon
6.1 Eritropoyetina y hematopoyesis
6.2 Producción de renina, regulación de la
presión sanguínea y el ph
6.3 Hormona paratiroidea y calcitriol
6.4 Otros mediadores asociados a la actividad
renal con función vascular y/o presora
6.5 Otras funciones endocrinas del riñón
7. Reexión sobre la siología renal
1. Generalidades de la función renal
Los riñones son órganos esenciales que, además
de actuar a modo de ltro eliminando productos
metabólicos y toxinas de la sangre, participan
en el control integrado del líquido extracelular,
del equilibrio electrolítico y del equilibrio acido-
básico. Producen hormonas como el calcitriol o la
eritropoyetina, y en ellos se activan metabolitos
como la enzima renina. Por ello, al describir la
siología renal, hay que recordar que va mucho más
allá del estudio del órgano que regula la excreción de
productos de desecho (Tabla 1). Esto es especialmente
relevante en el ámbito de la Nefrología, donde en
ocasiones, la valoración de mantener, aunque solo
sea de forma parcial esta funcionalidad renal, alcanza
una gran importancia.
Como ocurre con el resto de nuestro organismo,
la siología renal está ligada a la estructura del
aparato excretor renal, diseñada para mantener un
ujo unidireccional. Este ujo hará que la orina,
que inicia su formación en los riñones, órganos
principales del sistema, pase a través de los uréteres
a la vejiga urinaria para su almacenamiento, para
que posteriormente pueda ser eliminada a través de
la uretra. Para que esta actividad se lleve a cabo,
los riñones cuentan con una vascularización muy
signicativa, que facilita que, a pesar de su pequeño
tamaño, reciban aproximadamente un 20% del
gasto cardiaco. Además, una destacada inervación
por bras nerviosas simpáticas, regula entre otras
actividades la liberación de renina, el ujo sanguíneo
renal o la reabsorción de Na+ en las células tubulares.
2. caracteristicas morfo-funcionales de los
riñones. La nefrona
Desde un punto de vista macroscópico (Figura
1), los riñones son dos órganos ovalados con una
indentación medial. Miden aproximadamente 11 x
7 x 3 cm y pesan unos 150 g, siendo normalmente
el riñón izquierdo algo mayor que el derecho. Los
riñones se localizan en la zona retroperitoneal, en
la pared posterior del abdomen a ambos lados de
la columna vertebral, desde la altura de la última
vértebra dorsal hasta por encima de la tercera vértebra
lumbar. El riñón derecho suele estar algo más bajo
que el izquierdo, debido a la ocupación del espacio
derecho por otros órganos abdominales, como el
hígado. La cara medial de cada riñón contiene una
región con una muesca, llamada hilio, por la que
Fisiología Renal
Julia Carracedo
1
, Rafael Ramírez
2
1 Departamento de Genética, Fisiología y Microbiología, Universidad Complutense/Instituto de Investigación
Sanitaria Hospital 12 de Octubre (imas12), Madrid. Email: [email protected]
2 Departamento 2 de Biología de Sistemas, Facultad de Medicina y Ciencias de la Salud (IRYCIS),
Universidad de Alcalá, Alcalá de Henares, Madrid. Email: [email protected]

Fisiología Renal
www.nefrologiaaldia.org.es
2
Fisiología Renal
www.nefrologiaaldia.org.es
3
pasan la arteria y la vena renales, los vasos linfáticos,
la inervación y el uréter.
En un corte sagital del riñón pueden observarse
las estructuras que conforman el órgano y que
clásicamente se conocen como corteza externa y
regiones internas de la médula. La médula se divide
en 8-10 masas de tejido en forma de cono llamadas
pirámides renales. La base de cada pirámide se
origina en el borde entre la corteza y termina en la
papila, que se proyecta en el espacio de la pelvis
renal. El borde externo de la pelvis renal se divide
en los cálices mayores, que se extienden hacia abajo
y se dividen en los cálices menores, que recogen la
orina de los túbulos de cada papila.
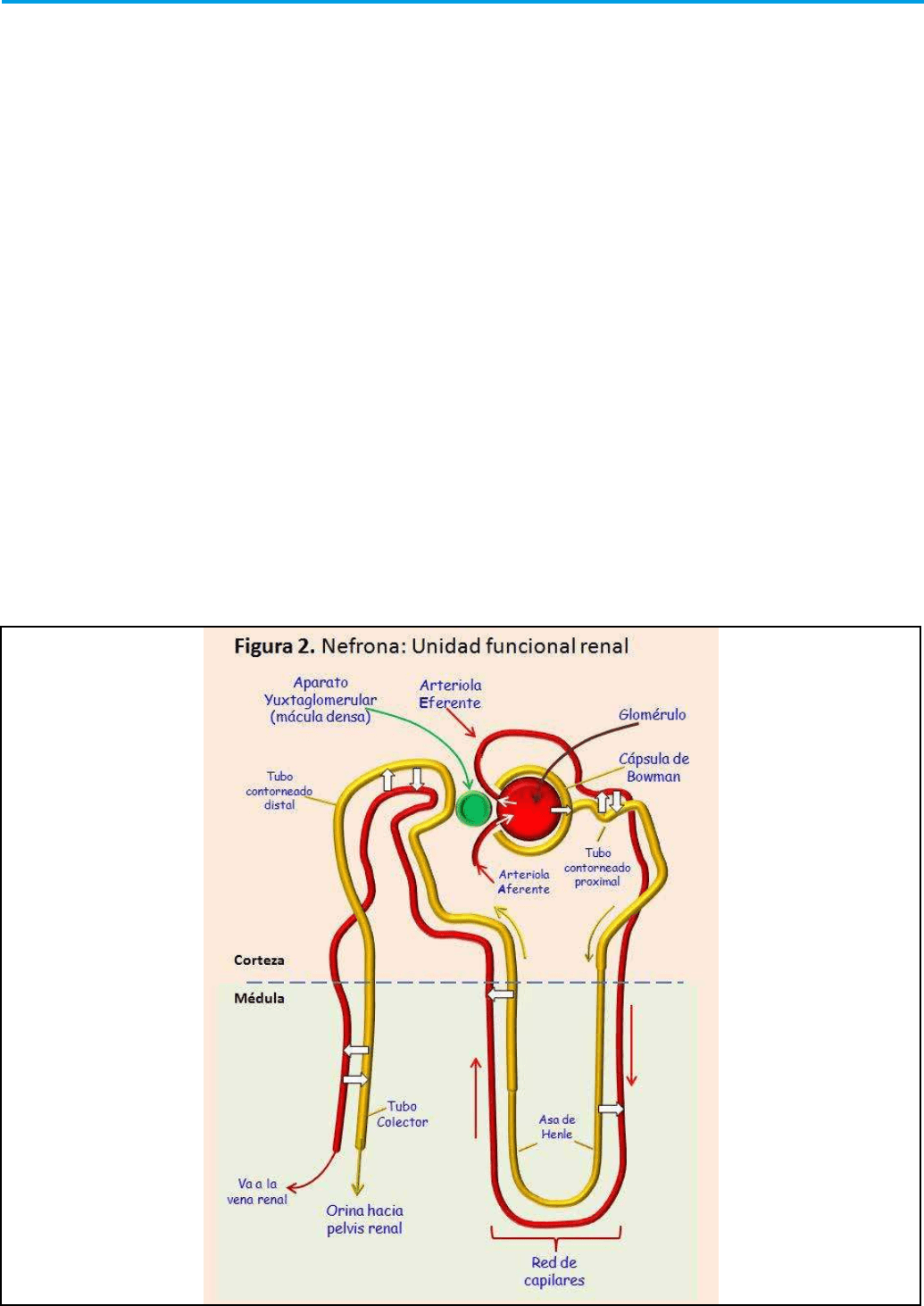
A nivel microscópico, se establece una unidad
funcional renal, la nefrona (Figura 2). Cada riñón
humano contiene alrededor de 800.000 a 1.000.000
nefronas, cada una de las cuales es capaz de formar
orina. A lo largo del envejecimiento renal normal,
por lesión o por enfermedad, el número de nefronas
se puede reducir gradualmente debido a que no
se pueden regenerar. Sin embargo, la pérdida de
nefronas no suele comprometer la función renal
porque se producen cambios adaptativos que suplen
la funcionalidad en el resto del sistema.
Cada nefrona está formada por un agrupamiento de
vasos capilares llamado glomérulo (Figura 3), por
el que se ltran grandes cantidades de líquido desde
la sangre, y por un túbulo largo en el que el líquido
ltrado se convierte en orina en su trayecto hacia la
pelvis renal.
Los capilares glomerulares se ramican y
anastomosan y, comparados con otros capilares
de otros sistemas, tienen una presión hidrostática
elevada (alrededor de 60 mmHg). Todo el glomérulo
está cubierto por la denominada cápsula de Bowman.
El líquido ltrado desde los capilares glomerulares
circula hacia la cápsula de Bowman y después al
túbulo proximal. Estas estructuras de la nefrona se
encuentran en la corteza del riñón. Desde el túbulo
proximal, el líquido ltrado discurre hacia el asa de
Henle, que desciende hasta la médula renal. El asa de
Fisiología Renal
www.nefrologiaaldia.org.es
4
Henle está constituida por una rama descendente y
otra ascendente. Las paredes de la rama descendente
y el segmento inferior de la rama ascendente del asa
de Henle son muy nas, y se llaman segmento no
del asa de Henle. Una vez la rama ascendente del
asa de Henle vuelve a la corteza renal, la pared se
engruesa denominándose segmento grueso del asa
ascendente. En la zona nal del segmento grueso de
la rama ascendente del asa de Henle, se localiza una
placa de células epiteliales especializadas que es la
mácula densa, cuya función es fundamental como
veremos más adelante. A continuación del asa de
Henle, el líquido llega al túbulo distal que se localiza
en la corteza renal.
Al túbulo distal le siguen el túbulo colector cortical.
Hay de 8 a 10 conductos colectores corticales que
se unen para formar un solo conducto colector
mayor que discurre hacia el interior de la médula y
se convierte en el conducto colector medular. Los
conductos colectores se van uniendo y formando
progresivamente conductos cada vez mayores que
vacían su contenido en la pelvis renal.
Dentro de las características anatómo-funcionales
del riñón hay que destacar la importancia de la
vasculatura. La arteria renal entra en el riñón a
través del hilio y después se ramica hasta formar
las arterias interlobulares, las arterias arciformes, las
arterias interlobulillares y las arteriolas aferentes,
que terminan en los capilares glomerulares, donde se
produce la ltración de grandes cantidades de líquido
y solutos para comenzar la formación de orina. Los
extremos distales de los capilares glomerulares
coalescen hasta formar la arteriola eferente, que
llega a la segunda red capilar formando los capilares
peritubulares, que rodean a los túbulos renales. En
denitiva, se puede armar que la circulación renal
tiene dos lechos capilares, los capilares glomerulares
y los capilares peritubulares, que están dispuestos en
serie y están separados por las arteriolas eferentes.
Estas arteriolas participan en la regulación de la
presión hidrostática en los dos grupos de capilares,
ajustando la resistencia de las arteriolas aferente y
eferente. Los capilares peritubulares continúan hacia
los vasos del sistema venoso, que discurren paralelos
a los vasos arteriolares, abandonando la sangre el
riñón junto a la arteria renal y el uréter.

Fisiología Renal
www.nefrologiaaldia.org.es
5
3. Fisiología renal y mantenimiento del
ambiente interno.
Si bien el concepto de “función renal” incluye
todas las actividades desarrolladas en el riñón para
el mantenimiento de la homeostasis (Tabla 1), si
tuviésemos que elegir un concepto que reeje la
función renal, este sería su capacidad para mantener
la homeostasis liquida en nuestro organismo a través
de la capacidad para depurar sustancias circulantes
en el plasma sanguíneo. Esta es una actividad
estrechamente relacionada con la capacidad de los
riñones para regular la concentración de agua, la
composición de iones inorgánicos, y mantener el
equilibrio ácido-base.
Como una consecuencia de esta actividad reguladora
del medio líquido, los riñones excretan productos
como la urea, generada del catabolismo de proteínas,
el ácido úrico producido a partir de ácidos nucleicos,
la creatinina, derivada en gran medida de la actividad
muscular, o productos nales de la degradación de
la hemoglobina. También a través de los riñones, se
eliminan drogas y otras sustancias químicas, como
los aditivos utilizados en alimentación.
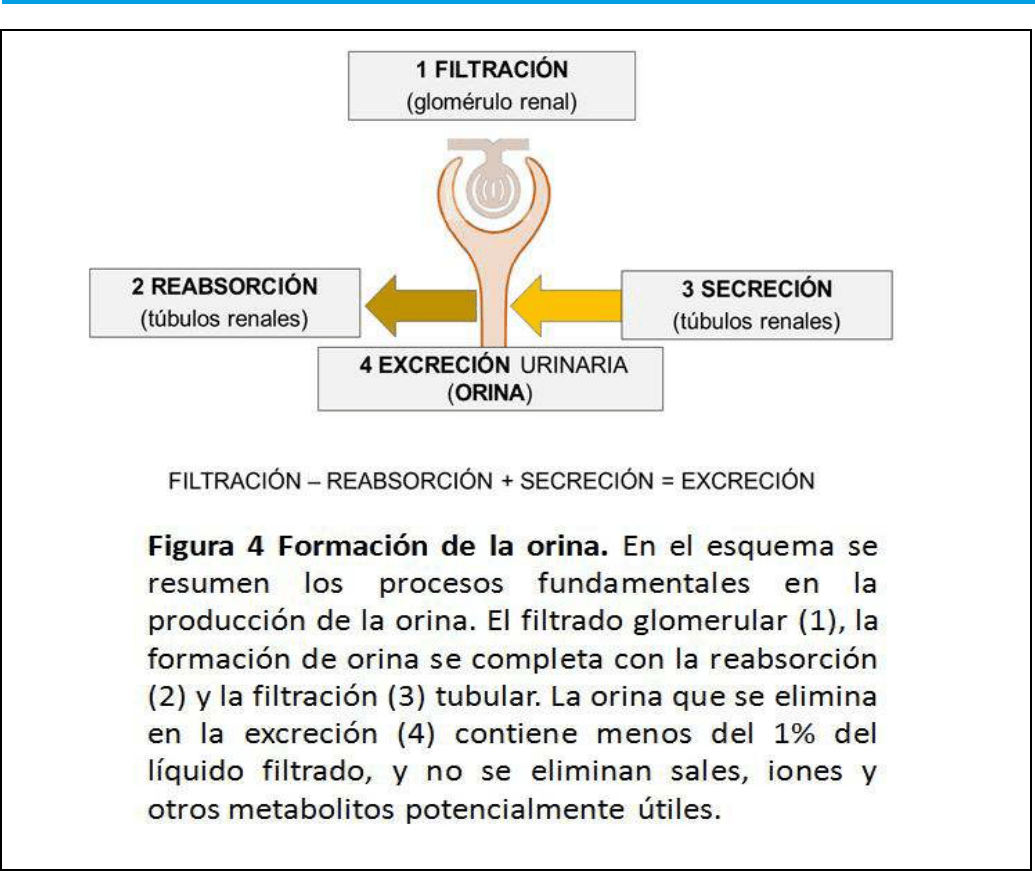
Proceso renal básico: Formación de orina
Los riñones procesan un volumen enorme de sangre
cada día. Cada minuto, el ujo sanguíneo que llega
a los glomérulos renales es de unos 1200 mililitros
de sangre, de los cuales, 650 ml corresponden
a plasma sanguíneo y de este, una quinta parte
aproximadamente será ltrado en el glomérulo. Esto
implica que cada 24 horas, los riñones ltran más de
60 veces todo el plasma sanguíneo.
Para evitar el enorme coste que la perdida de líquidos
y otros elementos esenciales puedan derivarse
del proceso de depuración renal; tras el ltrado
glomerular, la formación de orina se completa con
la reabsorción y ltración tubular, de forma que la
orina contenga nalmente menos del 1% de la parte
liquida ltrada, y no se eliminen sales, iones y otros
metabolitos que puedan ser útiles (Figura 4).
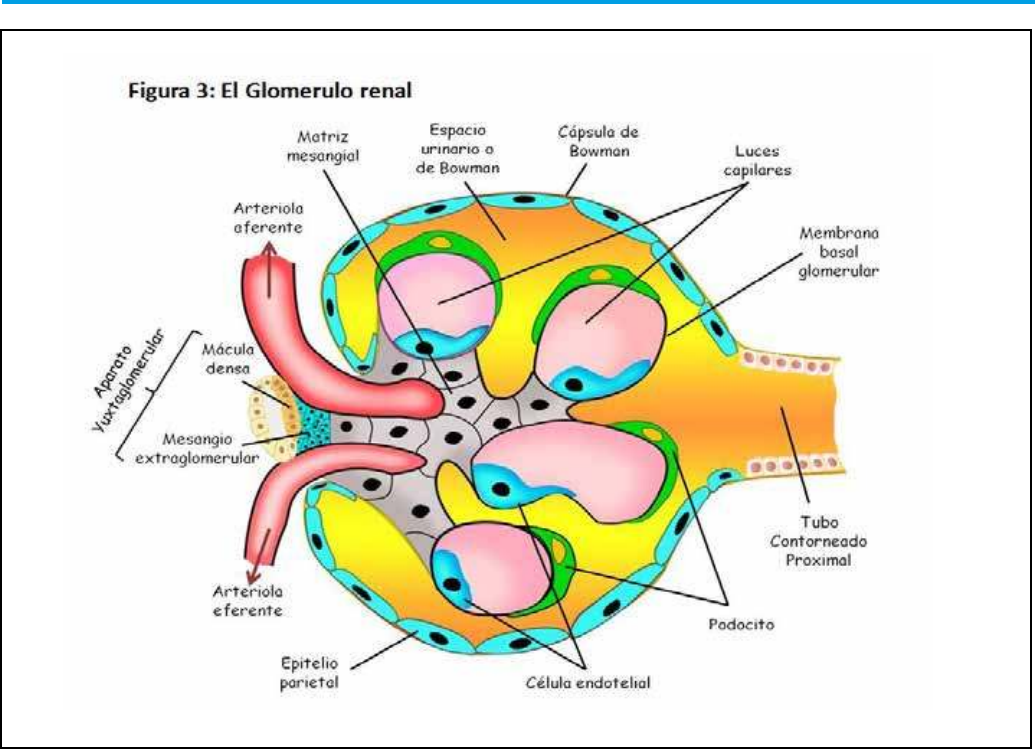
4. Formación de la orina. Filtrado
Glomerular
Desde una perspectiva funcional, la formación de la
orina se inicia en la cavidad glomerular, en la que se
puede distinguir una capa parietal externa de epitelio
escamoso simple, que contribuye a su mantenimiento
estructural pero no participa en la formación de la
orina; y una capa visceral, en la que la red capilar
vascular y las estructuras glomerulares prácticamente
forman una unidad funcional gracias a la actividad de
células epiteliales ramicadas altamente modicadas
llamadas podocitos, que literalmente, van a abrazar a
los capilares glomerulares.
4.1 Proceso de ltración
La ltración glomerular es un proceso pasivo. De
hecho, este proceso de ltración no tiene apenas gasto
energético para el organismo, por lo que podríamos
considerarlo un proceso meramente mecánico en
el que la presión hidrostática de la arteria aferente
empuja literalmente a la sangre contra la membrana
de ltración glomerular.
4.1.1 Membrana de ltración glomerular.
Esta membrana constituye una barrera que evita
el paso al túbulo renal de células y de la mayor
parte de las proteínas plasmáticas, generando un
“ultraltrado” compuesto fundamentalmente por
agua y elementos de pequeño tamaño circulantes en
la sangre. Para realizar esta función, la membrana de
ltración consta de un endotelio capilar fenestrado,
es decir, con poros capilares cuyo tamaño impide el
paso de células o la mayor parte de las proteínas.
4.1.2 Membrana basal.
Situada entra la capa endotelial y la epitelial.
Presenta un grosor de 240 a 340 nm. Está constituida
fundamentalmente por colágenos de tipo IV y V,
glicoproteínas, y proteoglicanos como el heparán
sulfato. Esta composición hace que presente una carga
electro-negativa que repele a pequeñas proteínas
y otros elementos cargados negativamente que
hubiesen atravesado la barrera endotelial fenestrada.
4.1.3 Membrana podocitaria.
Los podocitos son células polarizadas, con una parte
apical orientada hacia el espacio de la cápsula de
Bowman y otra hacia la lámina basal del endotelio.
Presentan un citoplasma aplanado, que emite multitud
de prolongaciones a modo de dedos que literalmente
abrazan a la lámina basal del endotelio, constituyendo
los “pies interdigitados”. Estos pies, suelen contactar
Fisiología Renal
www.nefrologiaaldia.org.es
6
con otros “pies” de podocitos vecinos mediante
complejos moleculares para formar diafragmas de
ranura, a través de los cuales moléculas de pequeño
tamaño y elementos líquidos pasaran de la estructura
glomerular al túbulo.
Los podocitos mantienen una baja actividad mitótica,
y su número permanece prácticamente invariable al
estar reprimida su capacidad replicativa. Pero esto
no quiere decir que sean células funcionalmente
inactivas. De hecho, sintetizan y liberan entre otras
moléculas colágeno de tipo IV, bronectina, laminina,
o heparán sulfato, que hacen que presenten una carga
electro-negativa muy importante y colaboren en la
actividad funcional de las células de la membrana
basal.
Pero quizás la actividad más relevante de los podocitos
en el ltrado glomerular sea la ligada a su función
en la arquitectura de la capsula glomerular. De una
parte, al abrazar mediante los pies interdigitados a
los capilares glomerulares, contribuyen a que estos
soporten la presión hidrostática, ya que los podocitos
tienen un citoesqueleto formado sobre todo por
lamentos de actina que al formar el complejo
actina-miosina modulan la respuesta de los capilares
a la presión hidrostática. Por otra parte, a través
de las ranuras que dejan estos pies podocitarios, el
líquido y las pequeñas moléculas que atraviesan las
membranas precedentes salen del glomérulo renal.
Para ello, resulta clave la distribución homogénea de
las podocitos mantenida gracias a su baja actividad
replicativa, y también que en las ranuras podocitarias
se expresen moléculas de adhesión y proteínas de
supercie que mantienen una constante interacción
entre podocitos vecinos.
Como resultado del proceso de ltración en la
estructura glomerular, las moléculas de menos
Fisiología Renal
www.nefrologiaaldia.org.es
7
de 3 nm de diámetro, como el agua, la glucosa,
aminoácidos y desechos nitrogenados, atraviesan
este ltro; mientras que las moléculas más complejas
y con cargas eléctricas como proteínas o ciertos
oligoelementos, permanecen en la sangre, lo que
resulta clave para el mantenimiento de la presión
coloidal osmótica en la misma, entre otras ventajas.
4.2 Presiones que afectan al ltrado
glomerular.
Al tratarse de un proceso físico dependiente de
presiones, los principios que rigen la ltración
glomerular no dieren de los establecidos para
cualquier otro lecho vascular, y por tanto depende de
los siguientes factores:
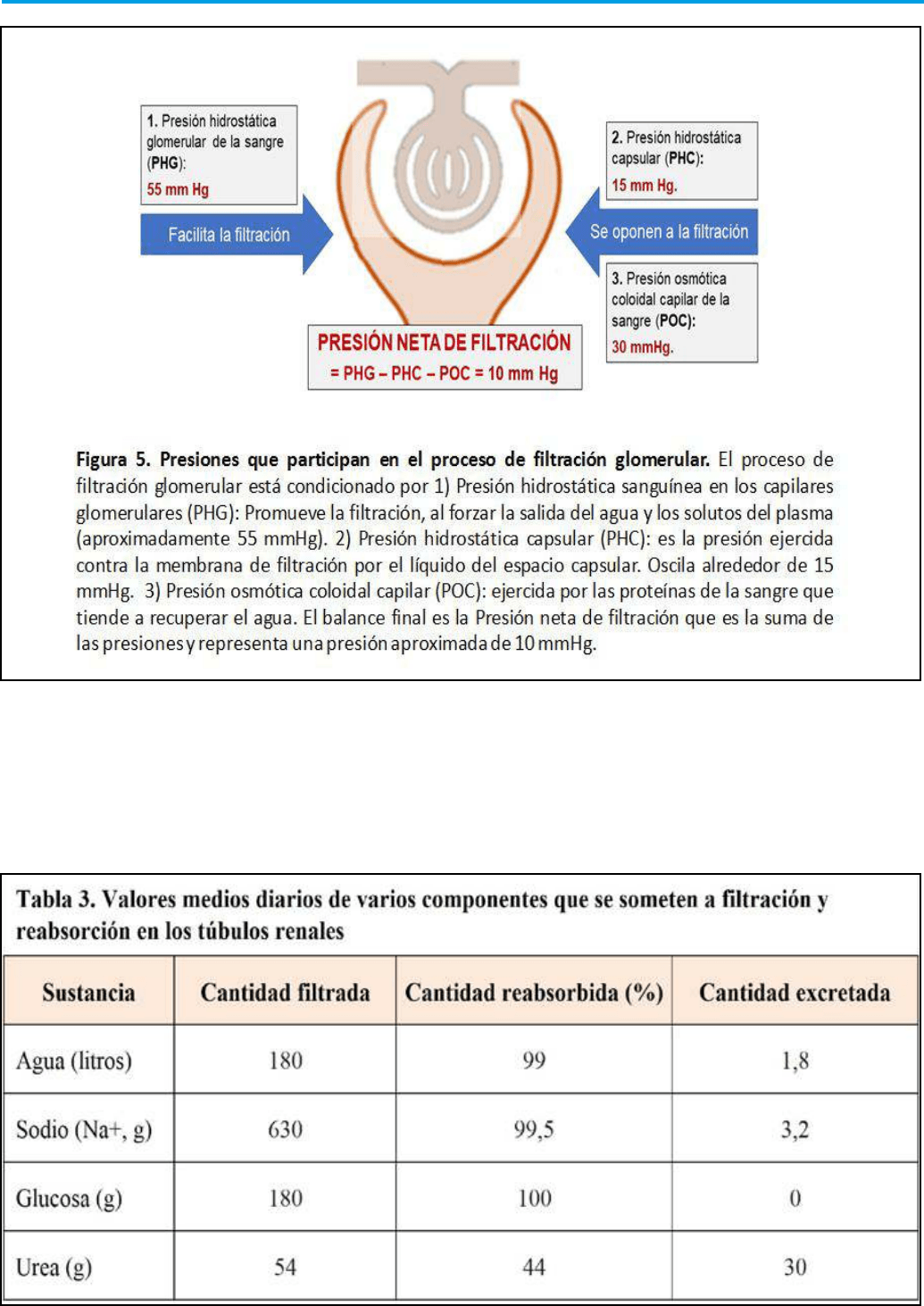
4.2.1 Presión hidrostática sanguínea
La presión hidrostática en capilares glomerulares es
esencialmente la fuerza principal que empuja a la
sangre contra la barrera de ltración glomerular. Por
esta razón, la presión sanguínea en el glomérulo renal
es excepcionalmente elevada (aproximadamente 55
mmHg) en comparación con la observada en otros
lechos capilares que es de 26 mmHg. Para mantener
esta elevada presión hidrostática sanguínea, arteriolas
aferentes y eferentes en el glomérulo presentan unas
diferencias en su diámetro importantes. Esto hace que
se diculte el drenaje capilar por la arteriola eferente,
y se mantenga en todo el lecho capilar glomerular una
elevada presión hidrostática, generándose a nivel de
arteria eferente una presión promedio de 30 mmHg.
4.2.2 La presión hidrostática capsular
Es la presión hidrostática ejercida contra la membrana
de ltración por el líquido del espacio capsular. Esta
presión se opone a la ltración hacia el túbulo renal
y oscila alrededor de 15 mm Hg.
4.2.3 Presión osmótica coloidal en los capilares
glomerulares
Es la presión ejercida por las proteínas en la sangre,
que tiende a recuperar el agua ltrada. A nivel de la
capsula glomerular se mantiene de forma más o menos
constante una presión coloidal de aproximadamente
30 mmHg.
Como se muestra en la (Figura 5), las presiones
anteriores determinan la presión de ltración neta.
Esta representa el resultado de la suma de las presiones
que favorecen o se oponen al ltrado glomerular. En
denitiva, a nivel capilar renal se genera una fuerza
de empuje extremadamente elevada y constante de
unos 10 mmHg.
4.3 Regulación del ltrado glomerular
Como hemos comentado con anterioridad, el proceso
de ltrado glomerular está condicionado por la presión
hidrostática capilar, que tiene que ser constante
para garantizar una actividad funcional ecaz de
los riñones. Pero la presión sanguínea glomerular
depende de la presión sanguínea sistémica, y ambas
presiones no siempre van a ir paralelas. Por ejemplo,

Fisiología Renal
www.nefrologiaaldia.org.es
8
si aumenta la presión hidrostática glomerular, se
producirá más ltrado y en consecuencia, más orina
y una reducción en la volemia con el consiguiente
descenso de la presión sistémica. Por lo tanto, el
mantenimiento de la presión capilar renal depende
de la regulación nerviosa y endocrina que regula
la presión sistémica, pero también cuenta con un
sistema de autorregulación local o intrínseco que
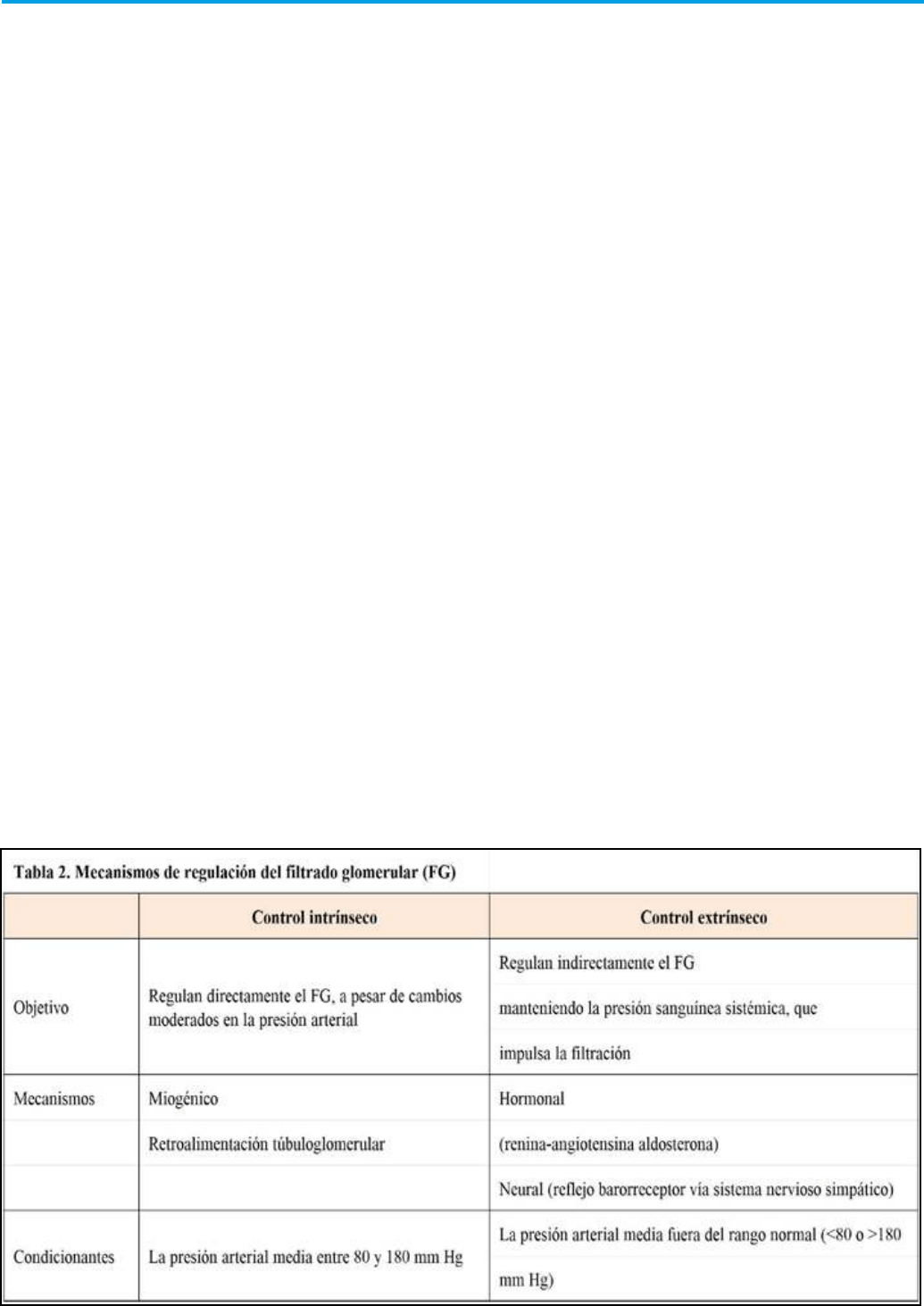
garantiza su mantenimiento (Tabla 2):
4.3.1 Controles intrínsecos.
La arquitectura glomerular requiere que, para
mantener una tasa de ltración casi constante,
la presión hidrostática capilar presente mínimas
diferencias. Para ello, la red capilar glomerular cuenta
con un sistema de autorregulación que compensa las
uctuaciones que pueda sufrir la presión sistémica.
La autorregulación renal utiliza dos mecanismos
diferentes de control:
a) Control miogénico. Depende de las células
musculares lisas presentes en la pared de los
capilares aferentes. Estas células poseen canales de
sodio/calcios sensibles al estiramiento, de forma
que, frente a un aumento de presión sistémica, las
células musculares se contraen disminuyendo la luz
capilar y con ello, el ujo vascular. Por el contrario,
un descenso en la presión sistémica provoca la
dilatación de las arteriolas aferentes.
b) El sistema de retroalimentación túbulo-glomerular
actúa cuando las células de la mácula densa en
el túbulo distal detectan un incremento en sodio
y/o cloruro. En respuesta a este incremento de
electrolitos, las células de la macula densa secretan
sustancias vasoconstrictoras como adenosina,
ATP y/o tromboxano, que por mecanismos mal
conocidos llegan a la arteria aferente provocando
vasoconstricción y con ello un descenso de la presión
capilar glomerular y de la ltración glomerular.
Estos mecanismos de autorregulación son muy ecaces
para mantener la presión hidrostática glomerular
prácticamente constante, con independencia de
las uctuaciones que sufra la presión sistémica;
en consecuencia, mantienen el ujo renal sin estar
condicionados por mecanismos sistémicos cuando
la presión arterial sistémica se mantiene en rangos
entre 80 y 180 mmHg, Sin embargo, son incapaces
de compensar grandes uctuaciones de presión.
Cuando la presión arterial cae por debajo de 80 mm
Hg, cesa la autorregulación intrínseca, y se activan
los mecanismos reguladores sistémicos que tienden
a preservar la actividad de elementos fundamentales
como el corazón o el sistema nervioso central.
4.3.2 Mecanismos de regulación extrínsecos o
sistémicos
a) Mecanismos neurogénicos. El control del
sistema nervioso simpático mantiene la homeostasis
vascular sistémica independiente de lo que ocurra
a nivel renal. Sin embargo, cuando el volumen de
líquido extracelular es extremadamente bajo (como
en el shock hipovolémico durante una hemorragia
severa), la actividad del sistema simpático provoca
una vasoconstricción general, que hace que se limite
el ujo sanguíneo glomerular hasta el punto de dañar
los riñones, con independencia de la actividad del
control intrínseco renal.
b) Sistema renina-angiotensina-aldosterona.
Frente a una caída importante de la presión sistémica,
la secreción de renina hace que se pierda la capacidad
de regulación intrínseca por diferentes vías:
- La actividad de retroalimentación con el sistema
nerviosos simpático.
- Un efecto sobre las células de mácula densa que,
frente al descenso de ltrado glomerular, dejan de
producir elementos vasoconstrictores.
4.4 Tasa de ltración glomerular
La cantidad de ltrado glomerular que se forma en
todos los corpúsculos renales de ambos riñones por
minuto es la tasa de ltración glomerular (TFG), que
suele ser de unos 125 mL/min, en los hombres, y algo
menor, unos de 105 mL/min, en las mujeres. Esta
TFG se mantiene relativamente constante. Si se eleva
de forma importante, el ultraltrado glomerular sería
tan alto que se dicultaría la actividad de reabsorción
tubular que veremos a continuación. Si es muy baja,
un exceso de reabsorción en el túbulo dicultaría la
excreción de productos a este nivel.
La TFG depende básicamente de tres condicionantes
(Figura 5). La presión hidrostática sanguínea con la
que el agua y los solutos son empujados contra la
membrana de ltración; la longitud o extensión de
esta membrana de ltración, y la permeabilidad de
la misma. En condiciones siológicas, la membrana
glomerular presenta muy pocas diferencias en su
Fisiología Renal
www.nefrologiaaldia.org.es
9
extensión o en su permeabilidad, por lo que la
presión de ltración neta, es decir, la presión total que
promueve la ltración, es regulada fundamentalmente
por:
- La presión hidrostática sanguínea en los capilares
glomerulares. Su valor suele ser alto, en torno a los
55 mm Hg.
- La presión hidrostática capsular ejercida contra
la membrana de ltración por el líquido que ha
penetrado en el espacio capsular. Esta presión se

Fisiología Renal
www.nefrologiaaldia.org.es
10
opone a la ltración y su, valor promedio es de 15
mm Hg.
- La presión osmótica coloidal de la sangre. También
se opone a la ltración, y es de unos 30 mm Hg.
Así pues, la presión neta es de unos 10 mm Hg, y
mantenerla estable hace que el ltrado glomerular
sea continuo y ecaz. Por ejemplo, una pérdida
importante de sangre reduce la tensión arterial
media, y con ello, la presión hidrostática glomerular.
Si esta desciende por debajo de 45 mm Hg, cesa el
ltrado, y con él la actividad renal en gran medida.
Sin embargo, en el sentido contrario los elementos
de regulación de la presión glomerular son muy
ecaces. Cuando la tensión arterial sistémica se eleva
por encima de lo normal, la presión de ltración neta
y la TFG aumentan muy poco, y en la práctica, son
capaces de trabajar con rangos de presión sanguínea
entre 80 y 180 mm Hg.
Aspectos relevantes en el proceso de ltración
renal
Se realiza en el corpúsculo renal en la nefrona. Su
principal característica es poseer un dintel máximo de
ltración de aproximadamente 4,5 nm de diámetro,
y mantener una carga electrostática que repele a
proteínas y otros elementos.
Para ello cuenta con: a) Un endotelio capilar
capaz de mantener estable el ujo y a presión
sanguínea, con poros de 50 a 100 µm de diámetro
que permita la salida de elementos del plasma;
b) Una membrana basal glomerular, con células
cargadas electronegativamente; c) Una capa de
células podocitarias, contráctiles, que envuelven a
los capilares regulando la actividad capilar y dejando
a su vez entre ellas espacios libres que permitan el
paso de pequeñas substancias.
Cuanticación del proceso de ltración: Se consigue
calculando la depuración de un soluto indicador que
se ltre totalmente a su paso por el riñón. Por ejemplo,
la inulina o la creatinina, utilizada habitualmente en
clínica que, aunque es secretada parcialmente en los
túbulos (entre un 10 y un 15%), se compensa con
la sobrevaloración entre un 10 y 15% en el método
colorimétrico de su medida. Se calcula que cada
glomérulo mantiene una TFG media de 125 ml/ min.,
que va disminuyendo a medida que aumenta la edad.
La ltración glomerular sigue fundamentalmente
leyes físicas reguladas por presiones que afectan al
ujo plasmático, estando implicadas:
Presión hidrostática del capilar glomerular, se
mantiene muy estable por la capacidad de regulación
de la resistencia de la arteriola aferente como en la
de la eferente. Su valor promedio es de 55 mm de Hg
Presión hidrostática en la cápsula de Bowman, con
un valor prácticamente despreciable en condiciones
siológicas ya que el líquido ltrado pasa
inmediatamente al túbulo.
Presión oncótica de las proteínas plasmáticas,
que, aunque crece ligeramente hacia el nal del
glomérulo por la disminución de volumen líquido
ltrado, suele mantenerse estable salvo en estados de
deshidratación, hipoproteinemia, etc.
5. Formación de la orina. Intercambio
tubular litros
Aproximadamente cada 22 minutos, la totalidad
del plasma sanguíneo ha sido ltrado en los
glomérulos. Esto quiere decir que los riñones ltran
aproximadamente 180 L/día de plasma. Sin embargo,
el volumen de orina en 24 horas suele ser algo inferior
a 1,5 litros, de los que aproximadamente el 95 % es
agua y el 5% restante son sustancias de desecho. Por
tanto, la composición del ultraltrado glomerular,
durante su paso por los túbulos renales es modicado
para que gran parte del agua y los oligoelementos
ltrados sean reabsorbidos y transportados a los
capilares peritubulares para ser reutilizados. Pero
a nivel tubular no solo se produce un proceso de
reabsorción. En sentido contrario, es decir, desde los
capilares peritubulares hacia la luz del túbulo renal,
algunas substancias son secretadas.
Estos procesos de reabsorción y secreción permiten
por ejemplo que, solutos importantes como la
glucosa se recuperen completamente, mientras
que la mayoría de las toxinas son secretadas y no
reabsorbidas. Por lo tanto, son procesos adaptativos
que varían en función de las necesidades siológicas.
En este sentido, prácticamente todos los nutrientes
orgánicos como la glucosa y los aminoácidos se
reabsorben por completo para mantener o restaurar
concentraciones plasmáticas dentro de un rango de
normalidad (Tabla 3).
En el túbulo, se reabsorbe más del 90% del agua y el
cloruro sódico ltrados y aproximadamente el 70%
del potasio y el 80% del bicarbonato. A pesar de que
todo el túbulo está involucrado en la reabsorción
de los diferentes productos, existen diferencias

Fisiología Renal
www.nefrologiaaldia.org.es
11
estructurales en el mismo que condicionan esta
reabsorción. En particular, la reabsorción de Na+ y
agua se verá condicionada por mecanismos de ajuste
en las regiones terminales del túbulo, que permitirán
de manera precisa regular la cantidad de estos
elementos reabsorbidos en función de las necesidades
hídricas del organismo; mientras que, en la región
proximal, más del 65% de Na+ será reabsorbido
y con él, gran parte del agua, aminoácidos o la
glucosa (Tabla 3). La actividad de hormonas en los
túbulos distal y colector condicionará que pequeñas
cantidades de agua y algunos iones que han llegado
hasta aquí (en torno al 10% de sales), se reabsorban
en caso necesario.
5.1 Mecanismos de reabsorción y secreción
tubulares.
5.1.1 Reabsorción de Grandes moléculas
A pesar del dintel de ltración glomerular, unos 30
g de proteínas con peso molecular inferior a 70.000
atraviesan diariamente el glomérulo. Estas proteínas,
junto con otros elementos como pequeñas hormonas
peptídicas, se recuperan en su totalidad en el túbulo
proximal mediante un proceso de endocitosis que
tiene lugar en el borde en cepillo de la cara apical del
túbulo proximal.
5.1.2 Reabsorción de glucosa y otras moléculas
importantes para el organismo
Glucosa, aminoácidos y vitaminas, son también
reabsorbidos en su práctica totalidad en el túbulo
proximal por un mecanismo de transporte activo
secundario, mediante cotransporte con el Na+.
Este mecanismo de transporte utiliza bombas y
otras unidades de transporte especícas situadas
principalmente en la membrana luminal de la célula
tubular, que son diferentes a las encontradas en la
membrana basolateral. Esta distribución polarizada
de transportadores permite un desplazamiento
transcelular neto de substancias.
En general, la máxima absorción de estos elementos
se produce en la primera porción del túbulo
contorneado proximal. En la cara apical de las células
del túbulo, y más concretamente, a nivel del borde
en cepillo de las mismas, transportadores de Na+
ionizado cotransportan estos elementos al interior
de la célula tubular. Desde aquí, los aminoácidos
salen por difusión pasiva o facilitada hacia el líquido
intersticial.
Otros elementos utilizaran bombas de transporte
como la Na, K-ATPasa, o canales de cloro para ser
extraídos al espacio intersticial. Quizás el elemento
de esta familia mejor estudiado es el transporte de
glucosa. Aproximadamente se ltran 180g de glucosa
cada día, de los que más del 99% es reabsorbida a
nivel tubular. Este proceso ocurre a diferentes niveles
en el túbulo y dado que la glucosa no difunde a través
de la bicapa lipídica celular, se hace a expensas de
los transportadores de membrana de sodio y glucosa
SGLT (sodium-glucose transporters). El 90% de la
glucosa se reabsorbe en el segmento contorneado del
túbulo proximal utilizando transportadores SGLT2,
mientras que el 10% restante se reabsorbe en el
segmento recto del túbulo proximal descendente
mediante transportadores SGLT1.
Como indica su nombre, estos transportadores son
proteínas situadas en la membrana luminal de las
células tubulares, que utilizan el movimiento de Na+
por gradiente electroquímico para que glucosa entre
en la célula del túbulo. La unión del Na+ que estaba
en el interior de la célula al SGLT genera un cambio
de conformación que permite la unión de la glucosa
al mismo. El Na+ saldrá de la célula por gradiente
electroquímico para permitir que la glucosa unida a
los SGLT penetre en la célula, y desde allí, de nuevo
los transportadores de glucosa la extraerán hacia
el compartimiento intravascular para que entre por
difusión a los capilares.
De esta manera se genera un balance a través de
SGLT de dos moléculas de Na+, una de glucosa
y 260 moléculas de agua, lo que hace que estos
transportadores soporten una alta tasa de actividad.
De hecho, el valor máximo de transporte de glucosa
es de aproximadamente 320 mg/min. Aunque
no todas las nefronas tienen un mismo índice de
saturación, por lo que el umbral de glucosa se sitúa
en una carga tubular de 220 mg/min, equivalente a
una concentración plasmática de 180 mg/dL (Umbral
renal de glucosa); cifra a partir de la cual comienzan
a aparecer cantidades signicativas de glucosa en
orina. La glucosa plasmática es ltrada y recuperada
por completo. La reabsorción de glucosa está sujeta
a un máximo de transporte tubular y la cantidad
reabsorbida por unidad de tiempo constituye el
transporte de glucosa (Tg), expresado en mg/
min, hasta que se alcanza el máximo de transporte

Fisiología Renal
www.nefrologiaaldia.org.es
12
tubular (Tmg), a partir del cual el transporte se hace
constante y concentraciones superiores conllevan la
eliminación del exceso por la orina. El valor de Tmg
es de 320 mg/min e indica el máximo de glucosa que
puede transportar el sistema. Sin embargo, debido a
que unas nefronas dieren de otras en su capacidad
de saturación, el umbral de glucosa se sitúa en una
carga mg/dL, (Umbral renal) a partir de la cual
comienzan a aparecer cantidades signicativas de
glucosa en orina.
Transporte Máximo
Los sistemas de transporte transcelular para los
diversos solutos requieren un funcionamiento
coordinado y aunque presentan cierta especicidad
para la sustancia que transportan, son saturables. De
ahí que se establezca un transporte máximo para casi
todas las sustancias que se reabsorben utilizando
transportadores de membrana, que se determina
como los mg/min de sustancia trasportada, y que
presenta diferencias para los diferentes solutos. Por
ejemplo, la necesidad de reabsorber casi toda la
glucosa que se ltra hace que el transporte máximo
para esta sustancia sea alto, ya que dispone de muchos
transportadores. Por el contrario, los elementos de
desecho que no tienen transportadores son excretados
en su mayor parte.
El hecho de que la glucosa tenga un transporte
máximo elevado no quiere decir que no pueda
saturarse. En general, cuando los niveles de glucosa
exceden de 80 mg/dl, los trasportadores se saturan y
aparece glucosuria a pesar de que los túbulos renales
siguen funcionando normalmente.
5.1.3 Reabsorción de iones y agua:
Agua, iones y algunos nutrientes son reabsorbidos
fundamentalmente a nivel del túbulo proximal por
diferentes mecanismos. El agua, seguirá un proceso
de transporte pasivo por arrastre osmótico. Por el
contrario, iones y algunos pequeños nutrientes no
reabsorbidos utilizarán trasportadores que necesitan
energía. Esta energía, será aportada en gran medida
por intercambiadores ligados al sodio.
Al igual que ocurría con los restantes elementos útiles
para el organismo, el agua, y la mayor parte de los iones
son recuperados. De hecho, más del 90% del agua y
del cloruro sódico ltrado son reabsorbidos. El 10%
restante tendrá una reabsorción condicionada a las
necesidades del organismo. La actividad del túbulo
depende en gran medida de mantener una presión
osmótica del uido similar a la del plasma. Puesto
que proteínas y otros elementos que participan en
el mantenimiento de la presión osmótica plasmática
no atraviesan la barrera glomerular, en el túbulo, la
presión isoosmótica con el plasma está condicionada
fundamentalmente por el contenido de agua y sal
(disociado como cloro y Na+) del ultraltrado.
a) Transporte de sodio: Como hemos comentado,
la reabsorción de Na+ es el principal motor para el
transporte de múltiples elementos que se acoplan
a los intercambiadores de sodio en las células del
túbulo.
El Na+, es el catión más abundante en el ultraltrado,
y en función de la región del túbulo donde se
reabsorba, parte del Na+ entra por difusión facilitada
a través de canales situados en la cara apical de
las células del túbulo. Pero el mayor porcentaje de
Na+, se reabsorbe en la región proximal del túbulo
utilizando un mecanismo de transporte activo con
gran gasto energético. De hecho, aproximadamente
el 80% de la energía utilizada para el transporte
activo a nivel tubular se consumirá en el transporte
de Na+, de ahí, que las células del túbulo en la región
proximal contengan muchas mitocondrias que serán
necesarias para suministrar el ATP que requerirá el
transporte a este nivel.
- Transporte de sodio a través de la membrana
basolateral
El sodio es transportado fuera de la célula del túbulo
por un transportador activo primario, la bomba
de Na+-K +-ATPasa de la membrana basolateral.
A partir de ahí, el Na+ es arrastrado por el agua
hacia los capilares peritubulares. Este arrastre es
favorecido porque el agua uye rápidamente por
las diferencias de presiones entre un intersticio con
muy baja presión osmótica hacia un capilar que al
contener proteínas y otros coloides presenta una
elevada presión osmótica.
Es importante recordar que la bomba Na+-K+-
ATPasa es una enzima ATPasa que saca tres iones
sodio (Na+) a la vez que ingresa dos iones potasio
(K+) en el interior de la célula, generando un
gradiente electronegativo intracelular
- Transporte de sodio a través de la membrana apical

Fisiología Renal
www.nefrologiaaldia.org.es
13
El gradiente electronegativo generado en el interior
de la célula del túbulo favorece la entrada del Na+
desde el ultraltrado, y para ello, se utilizan diferentes
mecanismos en función de la región del túbulo La
bomba de Na+-K+-ATPasa de la pared basal había
generado un gradiente electronegativo en el interior
de la célula del túbulo, al sacar 3 Na+ hacia fuera
por 2 K+ que entra, por lo que se favorece la entrada
de Na+ desde el borde apical. Pero además este Na+
que entra abre canales de escape para el K+ que
difunde rápidamente hacia el intersticio, acentuando
la electronegatividad intracelular y favoreciendo la
incorporación de Na+.
b) Reabsorción de cloro: Se produce por vía
paracelular, sin requerir energía para su transporte,
en el último segmento del túbulo proximal, y asa
de Henle. Pero un pequeño porcentaje de cloro es
absorbido mediante un cotransportador Na+/2Cl−/
K+, que aprovecha el movimiento de cargas positivas
para recuperar cloro.
c) Transporte del Agua: En el asa de Henle, la
permeabilidad de las células del túbulo cambia, y
esta modicación, tiene una función primordial en la
capacidad renal para concentrar la orina en función
de la dinámica de reabsorción del agua mediante
un mecanismo de intercambio a contracorriente
condicionado en gran medida por su disposición
en forma de U, paralela al conducto colector, y
penetrando hasta la papila de la médula renal, lo que
ocasiona que las nefronas se dispongan de forma
radial en el riñón.
En el asa de Henle se distingue un segmento delgado,
que da continuidad al tubo proximal, formado
por células epiteliales muy delgadas, con pocas
mitocondrias, y en consecuencia baja actividad
metabólica. Sin embrago, en esta rama descendente
siguen estando presente aquaporinas que permitirán
la reabsorción de agua.
La rama delgada del asa, se continua con una rama
ascendente gruesa, de células epiteliales con bombas
de Na•-K•-2cl- en su región apical y bombas de Na
+
-
K
+
-ATPasa en su zona basolateral, que al mantener
una alta actividad metabólica permiten la reabsorción
de iones como el sodio (Na+), cloro (Cl+) y potasio
(K+). Sin embargo, en la región ascendente, las
acuaporinas son escasas, lo que prácticamente
imposibilita la reabsorción de agua a este nivel.
Como consecuencia de estas características
diferenciales en la absorción en el asa de Henle, se
genera un intercambio de solutos y solventes hacia el
espacio intersticial que será clave en la reabsorción
de elementos a este nivel. Aproximadamente el
20% del agua que llega a la rama descendente será
reabsorbida siguiendo un gradiente osmótico, y
como consecuencia por difusión simple sin consumo
de energía. Es más, esta difusión es tan importante
que arrastrara algo de Na+ con el agua que difunde.
Por el contrario, en la rama ascendente la actividad
de transportadores hace que se lleguen a reabsorber
hasta un 25% del Na+, al que acompañaran otros
iones como el K+ o el Cl- o calcio, bicarbonato y
magnesio que también se reabsorben a este nivel.
Como consecuencia, se genera un mecanismo a
contracorriente por el que a medida que el ltrado
glomerular pasa por el asa de Henle, el ultraltrado
pierde agua y va concentrándose. Al llegar a las
ramas ascendentes ocurrirá el proceso opuesto, la
salida de solutos del ltrado al espacio extracelular.
El resultado neto es que en el asa se ha generado
un gradiente osmótico que arrastra agua, que pasa
posteriormente a los capilares peritubulares de las
nefronas, devolviéndola a la circulación sistémica.
Un elemento muy importante en la reabsorción en el
asa de Henle es la urea. La urea, principal producto
nal del metabolismo proteico, es eliminada en
su mayor parte por la orina. Pero una parte será
reabsorbida en la rama ascendente del asa de Henle
contribuyendo a aumentar la concentración de
solutos en el espacio extracelular. Su excreción no
sólo está determinada por la ltración glomerular
sino también por la reabsorción tubular que le
permite jugar un papel importante en la producción
de una orina concentrada y en el mantenimiento del
agua corporal.
5.2 Transporte en el Túbulo contorneado
distal y conducto colector
Mientras que en el túbulo proximal y en el asa de
Henle la dinámica de movimientos de solventes y
solutos solamente estaba condicionada por principios
físicos y la disponibilidad de transportadores; en esta
porción de la nefrona se va a regular la reabsorción
de los elementos presentes en el ultraltrado en

Fisiología Renal
www.nefrologiaaldia.org.es
14
función de los requerimientos homeostáticos.
Fundamentalmente una regulación endocrina a este
nivel permitirá, adaptar la excreción o reabsorción de
agua y otras sales a las necesidades hídricas en cada
momento
5.2.1 Hormona antidiurética (ADH)
Como su nombre indica, la ADH inhibe la secreción
de agua y con ello la producción de orina al
regular la cantidad de aquaporinas permeables
para la reabsorción de agua. Cuando el cuerpo está
sobrehidratado, desciende la osmolalidad del líquido
extracelular, y en paralelo, desciende la secreción
de ADH en la secreción por la hipósis posterior.
Como consecuencia, a nivel de túbulos colectores, la
actividad de las aquaporinas desciende y se excreta
más agua.
Junto a esta actividad sobre las aquaporinas, la ADH
aumenta la reabsorción de urea a nivel de túbulos
colectores como veremos más adelante
5.2.2 Aldosterona
La aldosterona actúa modulando la reabsorción del
Na+ que ha llegado hasta la región terminal de la
nefrona. De hecho, hasta el 5% del Na+ ltrado puede
ser reabsorbido gracias a la acción de la aldosterona.
Aunque desde una perspectiva siológica la función
de la aldosterona se relaciona con la regulación de
la presión arterial, para ejercer esta actividad, esta
hormona que se sintetiza en la corteza de las glándulas
suprarrenales, tiene su efecto principalmente en el
riñón, especícamente a nivel del túbulo contorneado
distal y del túbulo colector donde se expresan
receptores de mineralocorticoides. Tras su unión a
estos receptores, la aldosterona provoca la actividad
de bombas de Na+/K+/ATPasa, provocando la
reabsorción de Na+ y con el de Cl y agua en aquellas
regiones con aquaporinas; y la salida de K`+ hacia
la zona luminal del túbulo que será excretado en la
orina.
5.2.3 Péptido natriurético atrial o auricular
(PNA).
Este péptido forma parte de una familia de al menos
tres péptidos con actividad hipotensora. El PNA
es sintetizado fundamentalmente en los miocitos
auriculares y ventriculares, y tiene un efecto
natriurético disminuyendo la reabsorción del sodio a
nivel de nefrona terminal inhibiendo la expresión de
los transportadores de Na+ en esta región.
Junto a esta actividad modulando la reabsorción
de Na+, y con él, los elementos que pueden
acompañarle, a nivel renal, el PNA actúa sobre el
músculo liso vascular disminuyendo su actividad en
respuesta a substancias vasoconstrictoras; disminuye
la secreción de aldosterona, e inhibe la secreción
de ADH, así como la secreción de renina. Además,
dilata la arteria aferente glomerular; contrae la arteria
eferente glomerular y relaja las células mesangiales,
con incremento de la tasa de ltrado glomerular.
5.2.4 Hormona paratiroidea (PTH)
También actúa a este nivel, aumentando la reabsorción
de Ca2 principalmente en el tubo contorneado distal.
5.3 Secreción de substancias en la
formación de la orina
El plasma vehicula diferentes metabolitos y
elementos que no son útiles para el organismo, y
que, en consecuencia, deben ser eliminados. La
forma más directa es no reabsorberlos a partir del
ultraltrado. Pero en ocasiones además de no ser
reabsorbidas, es necesario que elementos no útiles
del plasma sean secretados hacia la luz del túbulo
para ser excretados en la orina. Sustancias como
H+, K+, NH4 + (ion amonio), creatinina, y algunos
ácidos y bases orgánicos son secretados desde los
capilares peritubulares hacia la luz tubular. Pero
también substancias que se han producido como
resultado de la actividad metabólica en las células el
túbulo en la reabsorción de solventes (como CO
3
H)
son secretadas para ser eliminadas por la orina.
Por tanto, en la formación de la orna es importante
considerar la secreción de:
• Medicamentos y metabolitos, que sobre todo al estar
unidos a proteínas transportadoras no son ltrados, y
por lo tanto, deben secretarse.
• Sustancias siológicamente no útiles o productos
intermedios que se han utilizado en la reabsorción
tubular.
Especialmente importante es la secreción de urea
por su actividad modulando la osmolaridad en
la región medular interna renal. Es obvio que la
urea, vista como un simple metabolito de desecho,
debe ser eliminada. Pero, además, en el proceso de

Fisiología Renal
www.nefrologiaaldia.org.es
15
reabsorción de agua en la nefrona, la actividad de
la urea es clave para formar el gradiente medular,
y en consecuencia, reabsorber el agua en el asa de
Henle. Aproximadamente el 50% de la urea ltrada
en el glomérulo se reabsorbe en el túbulo proximal
a través de una vía transcelular por un mecanismo
pasivo de arrastre de solventes. El resto, continua
sin ser absorbida por la estructura tubular que es
impermeable a la urea, hasta llegar a la parte distal
del tubo colector, donde se expresan transportadores
de urea que hacen a esta región de nuevo permeable
a la urea. Por tanto, la práctica totalidad de la urea
que entra en el túbulo colector baja hasta la médula
interna, incrementando su concentración en el túbulo
colector a medida que desciende. Obviamente, si
además se produce reabsorción de agua a este nivel,
se incrementará aún más la concentración de urea en
la región medular. Como el colector en esta región
cuenta con transportadores de urea, que además
son estimulados entre otros factores por la ADH,
a medida que se incrementa el nivel de urea y/o
se reabsorbe agua, la urea sale del túbulo colector
hacia el intersticio. Esto hace que en la zona medular
interna se produzca una alta concentración de
urea. A medida que se incrementa la concentración
de urea es secretada por difusión al asa de Henle,
incrementando la osmolaridad en esta región, y con
ello el arrastre de agua. Conforme la nefrona va
saliendo de la región medular, el proceso se invierte,
y la urea retorna de nuevo al túbulo colector donde
volverá a ser reconcentrada y reabsorbida.
Por lo tanto, una gran cantidad de urea simplemente
se recicla a lo largo del túbulo, aunque una parte
importante de esta urea pasa desde el líquido
intersticial a los capilares peritubulares para ser
llevada fuera de los riñones. Como la urea reciclada
desde el espacio intersticial al colector sigue al agua
que no se ha reabsorbido, el porcentaje de urea
reciclado depende en gran medida de volumen de
agua en el túbulo colector, y por tanto de la actividad
de la ADH. En ausencia de ADH, los trasportadores
de urea tienen menor actividad. Además, el túbulo
colector es más impermeable al agua. Por lo tanto,
concentración de orina es menor y tiene menos
trasportadores por lo que será menos reabsorbida y
más excretada en la orina. Por el contrario, si aumenta
la secreción de ADH, se incrementa la concentración
de urea en el túbulo colector y se facilita su salida,
con lo que se incrementa el gradiente osmótico
medular formando orina más concentrada
Relevancia de la función tubular renal:
El producto ltrado en el glomérulo se verá sometido
a procesos de secreción (paso de sustancias desde el
plasma a la luz tubular) y procesos de reabsorción
(paso de sustancias desde la luz tubular al plasma en
los diferentes segmentos tubulares; lo que determina
que al nal, la composición de la orina sea diferente
a la inicial ltrada, tanto en los solutos, como en el
contenido de agua.
En la actividad del túbulo, se van a integrar
mecanismos de transporte que incluyen
- Vía intercelular: movimiento de agua y solutos
mediante arrastre por solvente, siguiendo diferencias
osmóticas.
- Vía transcelular: mediante transportadores activos
primarios y secundarios, así como mediante transporte
facilitado (antitransporte y cotransporte). Muchos
de estos transportadores muestran saturación por lo
que tienen un transporte máximo. También se da la
difusión de solutos, muchos de ellos dependiente
del pH pues la forma difusible es la no disociada. El
motor de todos los movimientos de solutos y agua a
través de la supercie transepitelial, son las bombas
de Na/K ubicadas en la membrana basolateral.
Relación anatomo-funcional.
TÚBULO PROXIMAL (TP): En él se realiza la
reabsorción y secreción casi total de los principales
solutos, además de la reabsorción del 70% del agua
ltrada. El motor fundamental en estos procesos son
movimientos de arrastre del sodio por las bombas de
sodio/potasio ubicadas en la membrana basolateral
de sus células.
Junto con el sodio, mediante cotransporte se reabsorbe
la glucosa, los aminoácidos, los fosfatos, sulfatos,
etc. Este tipo de transporte presenta saturación y
por tanto un máximo (Tm). Signica que a partir de
cierta concentración del soluto en plasma (umbral),
todo exceso en la ltración se excretará. Mientras no
se alcance el umbral todo lo ltrado será reabsorbido.
Además, el movimiento de sodio condiciona un
antitransporte sodio/hidrogenión (NHE3) que facilita
la secreción de hidrogeniones y la recuperación del
bicarbonato ltrado (este antitransporte es regulado
hormonalmente, se inhibe con la PTH y el glucagón
y se activa con la angiotensina II, alfa-catecolaminas
o la endotelina.
Este documento contiene más páginas...
Descargar Completo
335_301120201022.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.