SINAPSIS
La funcion del SNC depende de la operacion de multiples circuitos neuronales. Estos circuitos se
definen por las neuronas que los integran y sus conexiones. La operacion de los circuitos esta en la
base de las conductas animales y las respuestas de estos frente a los cambios del medio. La salida
de los mismos esta determinado por las neuronas que las componen, por la arquitectura de la red
y por las propiedades de esas conexiones: el contacto sinaptico.
-Se identifica un sentido del flujo de informacion, donde definimos una neurona pre-sinaptica que
va a influenciar sobre una segunda neurona del circuito, que es la neurona post-sinaptica. Por lo
tanto, el flujo de informacion va desde la neurona pre sinaptica hacia la post sinaptica. Esta
transmision de informacion depende de dos mecanismos fundamentales, uno es la CONDUCCION
a nivel del axon y la TRANSMISION SINAPTICA.
-Sinapsis: se define como una union especializada entre neuronas o entre una neurona y otra
celula excitable o no, donde la informacion se transmite de una a otra con gran presicion espacial
y temporal. La sinapsis no solo ocurre entre neuronas, sino que aveces uno de los participantes no
necesariamente tiene que ser una neurona, como por ej, la transmision a nivel de la placa motora
que se da entre la neurona y el musculo. El SNC no es el unico sistema donde se da el manejo de la
informacion, sino que el sistema humoral tambien realiza una funcion similar, siendo la diferencia
la precision espacial y temporal con que lo hace el SNC.
Las neuronas son celulas independientes, que estan completamente rodeadas por membrana
plasmatica, por lo tanto no hay continuidad citoplasmatica entre los diferentes componentes del
SNC. La corriente asociada a un PA no va a atravesar la segunda neurona ya que el medio
extracelular ofrece una resistencia electrica muy baja a diferencia de la resistencia que presentan
las membranas biologicas. Entonces, las corrientes responsables de la generacion del PA no van a
atravesar la membrana de la segunda neurona por este motivo, y toda la corriente se va a fugar
hacia el medio extracelular y muy poca corriente va a atravesar la segunda neurona para modificar
el Vm de esa neurona.
-Identificamos dos grandes mecanismos para la transmision de informacion entre neuronas:
--Transmision Electrica: transmision directa de corriente de una neurona a la otra.
--Transmision Quimica: la transmision de informacion no es directa, sino que esta mediado por un
intermediario quimico, que es el neurotransmisor.
--Transmision Mixta: multiples contactos sinapticos emplean ambas modalidades en forma
concomitante.
-TRANSMISION ELECTRICA-
Hay un flujo directo de corriente. Para que esto sea efectivo es necesario la existencia de una
region de contacto altamente especializada. Este contacto asegura que una parte sustancial de
corriente atraviesa un contacto especializado, fluye hacia la segunda neurona modificando
entonces el Vm de la primera y de la segunda. La corriente desencadena los PA, y parte de esta
corriente, a demas de atravesar la membrana de la neurona que esta generando el PA, tambien
fluye a traves de estos contactos hacia la segunda neurona para atravesar la membrana post
sinaptica y modificar tambien su Vm. El flujo de informacion en el SNC viene determinado por
cambios del Vm de las distintas neuronas que conforman los circuitos.
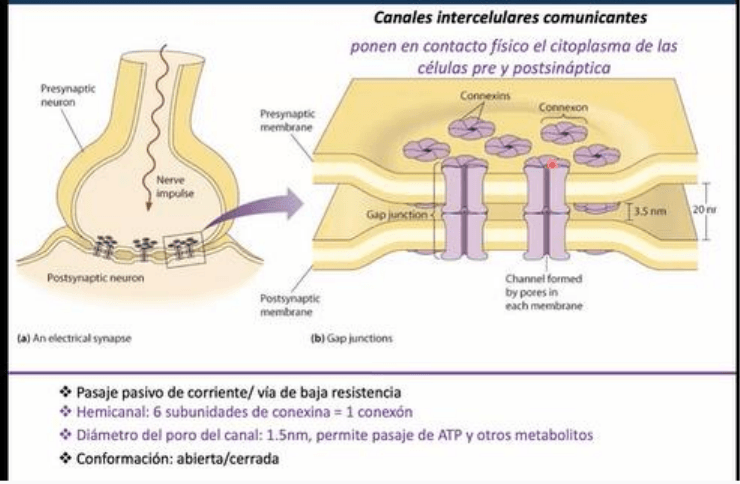
En este tipo de sinapsis, desde el punto de vista estructural, el sustrato anatomico son las uniones
GAP. Estas estructuras son agregados macromoleculares que conforman canales ionicos. Las
uniones GAP son cumulos de canales especiales de membrana, que estan formadas por un canal
intercelular. Un canal intercelular esta conformado por dos canales dispuestos en serie, donde uno
esta dado por una celula y el otro por la siguiente. El alineado y anclaje de estos dos canales forma
un canal funcional de las uniones gap (en hendidura). Hay un estrechamiento considerable del
espacio extracelular, con una distancia de 3.5nm. Esta distancia viene determinada por la distancia
minima necesaria para que se de la interaccion a nivel de los canales intercelulares en el medio
extracelular.
A diferencia
del resto de los canales, el poro acuoso es considerablemente mas grande que por ej, los canales
de Na y K. Este tamaño permite el pasaje de iones, ATP y otros metabolitos. Estos contactos que
permiten los flujos de particulas cargadas (iones) sustentan corrientes ionicas pero tambien
permiten el acople metabolico, que es una modalidad de señalizacion entre neuronas que no
implica cambios del Vm, sino que señalizacion quimica de moleculas o mensajeros generado en
una de las neuronas, que influencian en otra neurona.
-A su vez, cada canal intercelular esta formado por dos canales, se denomina hemicanal a cada
canal de la celula, entendiendo que un canal completo son ambos canales interactuando y
conformando un canal funcional. Cada hemicanal, conocido como CONEXON, esta conformado por
6 subunidades que pueden ser identicas o no, es decir, es un hexamero dispuestos alrededor de
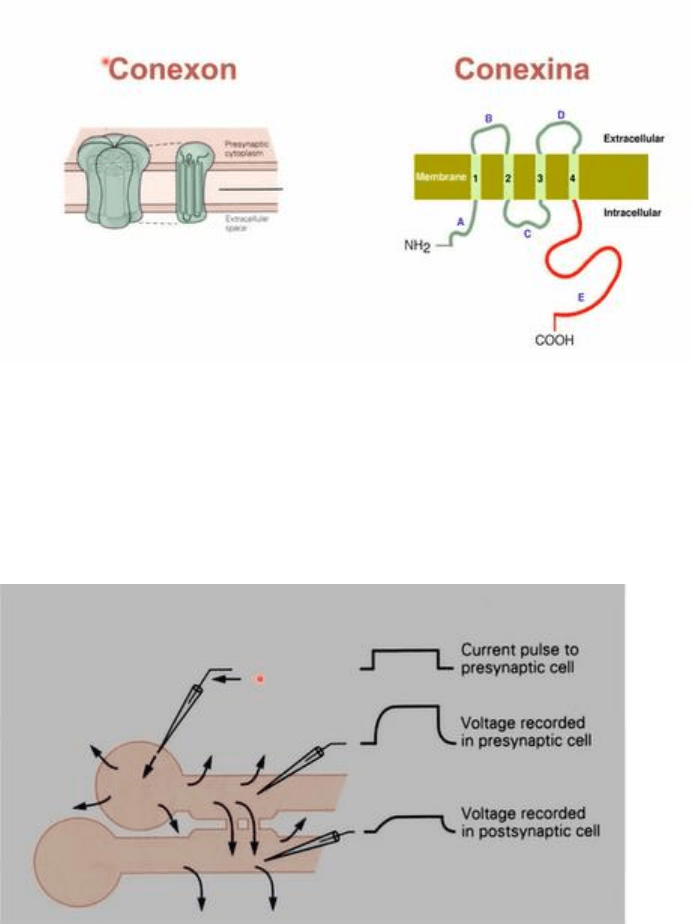
un poro central, que es el poro del canal. Cada una de las 6 subunidades se denomina CONEXINA,
y presenta 4 dominios transmembrana constituidos por alfa helice, dos bucles extracelulares entre
el dominio 1y2 y el 3y4. Estos bucles en cuanto a su secuencia aminoacidica son altamente
conservados ya que permiten la interaccion con el hemicanal de la otra celula. Del lado intracelular
quedan los extremos amino y carboxi terminal y un bucle entre los segmentos transmembrana
2y3. Se identifico que el extremo carboxi T es de gran importancia en la regulacion y trafico de
protenias. En mamiferos se han identificado genes que codifican diferentes conexinas y que
conforman diferentes canales, con propiedades diferentes en cuanto a la permeabilidad, tamaño,
modulacion (transicion entre abierto y cerrado) y presentan un patron de expresion especifico de
acuerdo a los diferentes tejidos y tipos celulares. La gran mayoria de los contactos sinapticos
donde por lo menos una celula es una neurona, esta mediado por la CONEXINA 36 o Cx36.
-Metodos de estudio de las sinapsis electricas-
Una modalidad tipica requiere un registro simultaneo intracelular, donde se tiene acceso al lado
interno de las neuronas mediante microelectrodos. En una configuracion mas compleja se puede
utilizar un electrodo que permita la inyeccion de corriente para modificar el Vm de una de las
neuronas, y un electrodo a cada lado de la sinapsis, de forma de monitorizar los cambios del Vm
provocado por las inyecciones de corriente.
Se observa esquematizado
un pulso rectangular de corriente para diagnosticar si existe una via de baja resistencia que
permita el flujo de corriente entre las celulas. Se aplico un pulso rectangular de corriente mediante
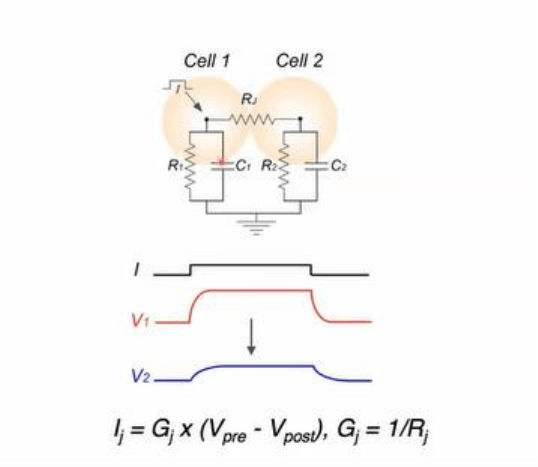
un microelectrodo, que corresponde a un pulso despolarizador, donde la punta del electrodo se
hace positiva respecto al electrodo extracelular y esto establece una circulacion de corriente que
se esquematiza con las flechas. La mayor parte de la corriente de acuerdo a la Ley de Kirchoff, que
tiende a circular por las ramas de menor resistencia, dice que la mayor parte de la corriente va a
fluir a traves de la membrana de la neurona inyectada, produciendo una modificacion del Vm de
acuerdo a sus propiedades pasivas. En caso de que exista un contacto electrico, una fraccion de la
corriente inyectada tambien va a fluir a traves de estos contactos dado que el mismo representa
una via paralela para el flujo de corriente. La corriente que atraviesa la primera celula hacia la
segunda, va a salir por la membrana de esta segunda celula para cerrar el circuito. Es esta
corriente, la que atraveso el contacto y la membrana de la segunda celula, la que va a modificar el
potencial transmembrana de la segunda celula. La señal en la segunda neurona nunca va a ser
mayor que la primera ya que es un circuito pasivo, donde esta señal es considerada menor que la
modificacion que ocurrio en la primera celula, por el motivo de que LA MAYOR PARTE DE LA
CORRIENTE SE FUGA atravesando la membrana de la neurona inyectada (primera neurona). A
demas de presentar menor amplitud y mismo signo ya que es una despolarizacion en ambos
casos, las señales post sinapticas tienen un curso temporal mas lenta en la segunda respecto a la
primera neurona.
Se emplea un circuito analogo, donde cada neurona es representada por un circuito con sus
respectivas resistencias y capacitancias globales de membrana. El unico elemento introducido es la
resistencia Rj. Esta representacion corresponde a una resistencia Ohmica. Muchos de los
contactos electricos dentro de ciertos rangos de voltaje se comportan como una resistencia
Ohmica, aplicando la Ley de Ohm dando una relacion entre la resistencia y voltaje de corriente. Si
aplicamos un pulso de corriente en la celula 1, vemos que la corriente va a fluir mayoritariamente
a traves de la resistencia y capacitancia de la neurona pre sinaptica. Ambas ramas de la celula 1 y 2
interconectadas con Rj, representa una via en paralelo. Parte de la corriente inyectada en el nodo
de la celula 1 fluira a traves del Rc de la celula 1 y en menor medida a traves del contacto electrico
Rj y de R2 y C2 (celula 2). Dado que la mayor parte de la corriente atraviesa la celula 1 y se
encuentran dos elementos dispuestos en serie, una parte menor de la corriente va a fluir por la
celula 2. Entonces, se genera un cambio de voltaje en la celula 1 y un cambio de voltaje de menor
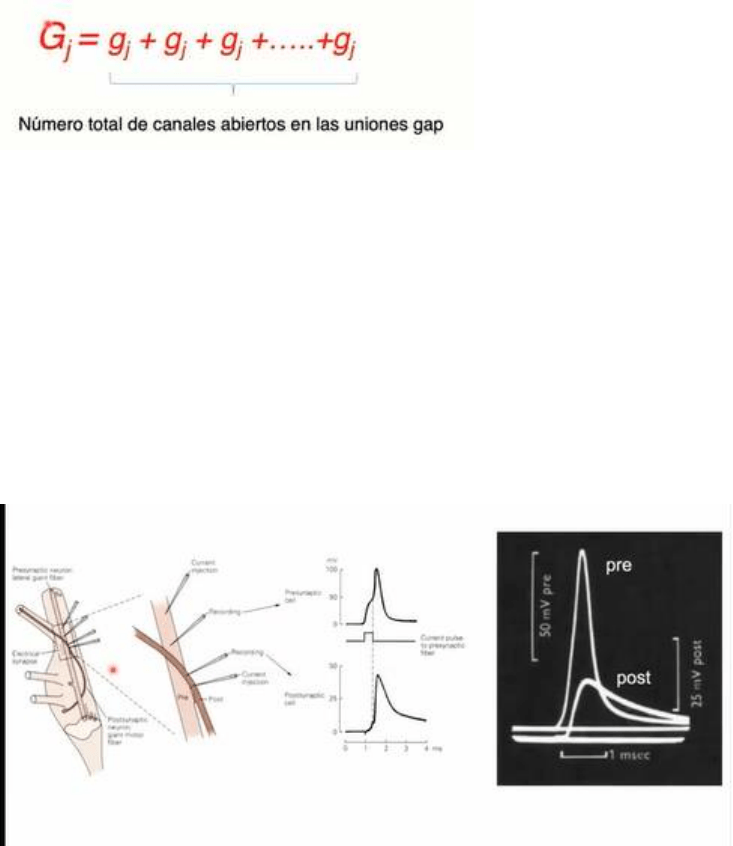
amplitud en la celula 2.
-Podemos identificar los determinantes de la magnitud de la corriente sinaptica. Ij es igual a la
conductancia multiplicada por la diferencia de voltaje entre las celulas acopladas. Esto indica que
una vez que se establezca una diferencia de potencial entre las celulas acopladas a traves de sus
citoplasmas, se va a establecer la circulacion de corriente. El sentido de circulacion de corriente
sera desde la celula mas electropositiva a la mas electronegativa. En este caso concreto, dado que
el pulso fue despolizador, el flujo de corriente sera desde la celula 1 a la celula 2 (entrada de
cargas + en la celula 1 ).
-Cada contacto es un agregado de muchos canales intercelulares, podemos definir una
conductancia unitaria de los canales, representada como gj y una conductancia global o
macroscopica de los contactos, representada como Gj. Gj es igual a la suma de las conductancias
unitarias. El numero total de canales abiertos presentes en los contactos nos determina la
conductancia macroscopica de los contactos.
Los determinantes de la eficacia sinaptica, que es la conductancia de los contactos, va a estar
determinada por la conductancia unitaria de cada canal, el estado de los canales (abierto o
cerrado) y por la cantidad de canales que se encuentren en estado abierto.
-Este tipo de contactos electricos (sinapsis) son rapidos, ya que mas alla de la existencia de
contactos es necesario que se establezca una diferencia del Vm entre las celulas acopladas de las
membranas, donde esto basta para que se establezca de forma instantanea el flujo de corriente
desde una celula a otra (de pre sinaptica a post sinaptica). Si bien el flujo de corriente es
instantaneo, no significa que el cambio de potencial tambien lo sea, ya que esto se determina por
la constante de tiempo de la membrana post sinaptica. La modalidad de transmision electrica es
sensiblemente mas rapida que la modalidad quimica. En muchos casos, practicamente no hay
retardo sinaptico.
Se inyecto y
midio el Vm de ambos elementos que conforman la union entre ambas neuronas. El objetivo del
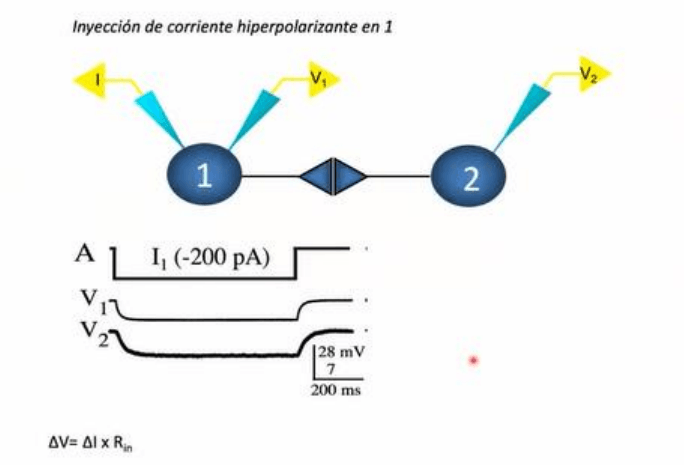
estudio era identificar la naturaleza de la transmision de informacion y en base a la diferencia de
tiempo, se concluyo que la transmision no podia ser de tipo quimico sino que de tipo electrico,
basado en las medidas de tiempo entre la señal pre sinaptica y la señal post sinaptica. La
aplicacion de un pulso supra umbral de corriente en la celula pre sinaptica genera un PA en esta
celula que con una latencia practicamente inexistente, se establece un potencial sinaptico en la
neurona post sinaptica. Se observa que el pie del PA en la celula pre sinaptica practicamente
coincide en el tiempo con el inicio del potencial sinaptico, llamado potencial de acoplamiento en la
neurona post sinaptica. Se determino que no existia contacto quimico a este nivel dado que los
retardos sinapticos de tipo quimico rondan el ms, donde aqui es muchisimo menos.
-Bidireccionalidad: esta transmision se basa fundamentalmente en el flujo directo de corriente de
una neurona a la otra a traves de un contacto especializado, determina de acuerdo a la Ley de
Ohm, que la transmision pueda ser bidoreccional. El flujo de corriente va a estar determinado por
cual de las dos neuronas es electropositiva respecto de la otra.
Por ej en este caso, se
registro en forma simultanea dos neuronas para estudiar la direccionalidad de la transmision. Se
aplico corriente con un microelectrodo en una de las celulas, y se aplico una corriente
hiperpolarizadora que hace mas negativo el Vm de la neurona 1. La medida de voltaje de la
inyeccion del microelectrodo se encuentra representada en V1, que le corresponde la calibracion
de 28 mV. En forma concomitante, a esta modificacion del Vm (V1) en una segunda neurona
proxima a la primera, registrada tambien en forma intracelular mediante un amplificador
representado en V2, en forma concomitante, la modificacion del Vm de la primera neurona es
acompañada por una modificacion casi inmediata del Vm de la segunda neurona, aunque con un
curso temporal mas lento. La modificacion del Vm de la segunda neurona es de menor amplitud,
donde corresponde a 7mV (V2).
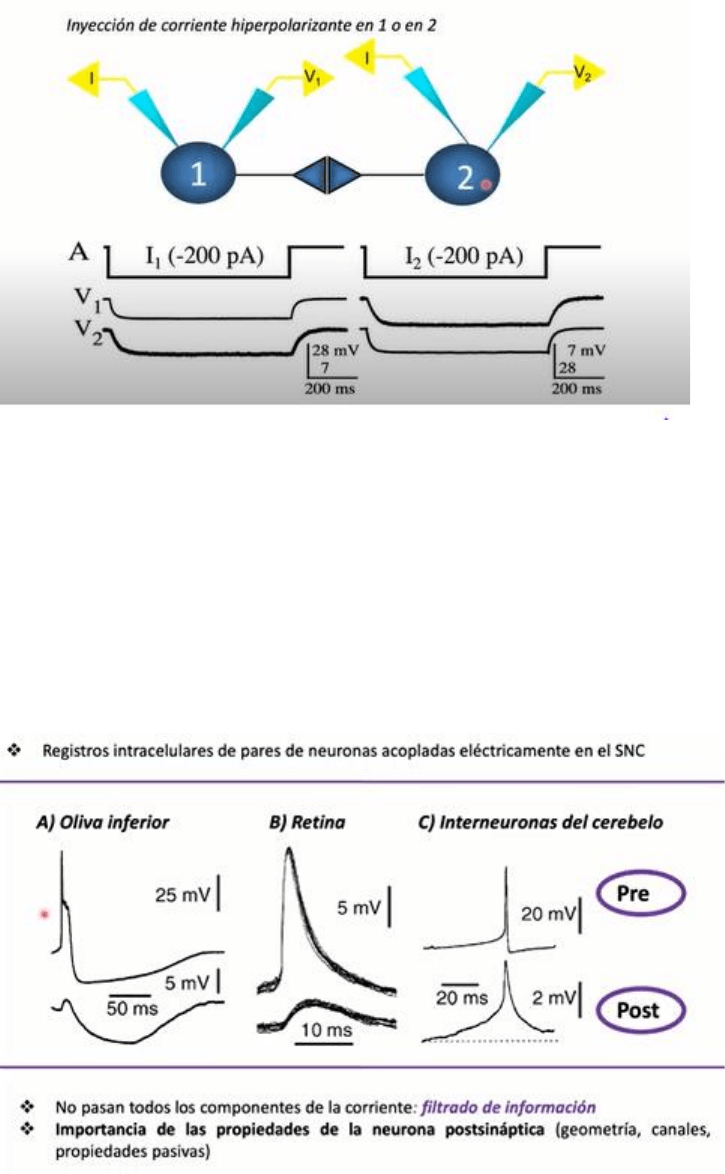
Cuando se hace la
maniobra simetrica pero la inyeccion de corriente es en la celula 2, se aplica un pulso de la misma
intensidad (-200pA), tenemos una modificacion del Vm de la celula 2 que produce una
modificacion de menor amplitud y curso temporal en la celula 1. Esto nos demuestra que esta
modalidad de transmision permite la comunicacion en forma bidireccional. Lo unico que va a estar
determinando el sentido del flujo de informacion sera cual es la celula que recibio la modificacion
inicial del Vm ya sea la 1 o la 2. Es un aspecto distintivo de esta modalidad de transmision sinaptica
electrica.
-Si bien se creia que esta transmision electrica era casi exclusiva de los invertebrados, tambien se
pudo demostrar que este tipo de transmision estaba tambien presente en todas las regiones del
SNC de los mamiferos.
Ej, nucleo de la
oliva inferior representado en A , se observa la señal de un PA que genera una meseta producto de
una corriente de Ca que produce una hiperpolarizacion muy prolongada producto de la activacion
de una corriente de K dependiente de Ca, y una recuperacion de la hiperpolarizacion por la
activacion de una corriente de tipo IH. Esta forma de onda relativamente compleja del PA pre
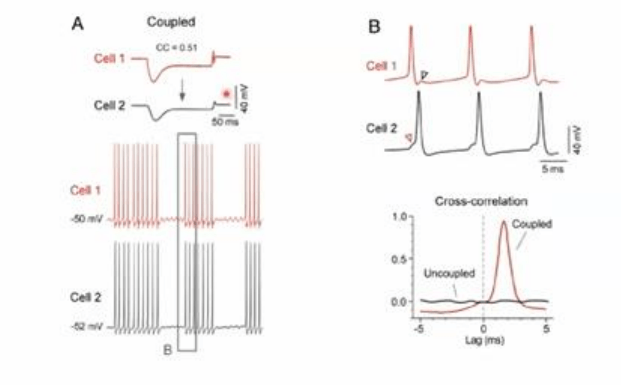
sinaptica se refleja en la neurona post sinaptica, con una fase despolarizadora y otra fase
hiperpolarizadora.
-Resumen-
La transmision sinaptica electrica se basa esencialmente en un flujo pasivo de corrientes de una
celula a la otra. No hay necesariamente un mecanismo activo, donde simplemente esta gobernado
por la diferencia de potencial entre las celulas acopladas y la conductancia de contacto que esta
determinada a su vez por la conductancia unitaria de los canales, numero de canales y el estado
de estos. Alguna de las consecuencias de este estado de transmision es que es una modalidad
rapida, donde la latencia es practicamente inexistente. El cambio del Vm en la celula post sinaptica
es proporcional al cambio de Vm pre sinaptico y permite la transmision de señales no solo las
despolarizadoras (asociadas al PA) sino que tambien hiperpolarizadoras. La gran mayoria de los
contactos sinapticos electricos funcionan de manera bidireccional permitiendo el pasaje reciproco
de una celula a la otra.
-Otra de las funciones a demas de la transmision rapida, la combinacion de la rapidez de la
transmision y la posibilidad de la interaccion reciproca entre las neuronas que conforman los
circuitos, permite la sincronizacion.
Se observa que las
neuronas estan fuertemente acopladas, con una modificacion del Vm de una de las celulas con un
pulso hiperpolarizante y el efecto sobre el Vm de la celula siguiente. Cuando estas dos celulas
descargan PA, estas descargas son altamente sincronicas (A). Si bien hay una celula comando y una
que la sigue, la relacion entre las descargas es extremadamente breve y muy sincronica, es decir,
el retardo entre una y otra es constante (B). En el grafico inferior derecho, se cuantifica mediante
la funcion de correlacion cruzada, donde se mide la probabilidad de descarga de la celula 2, dado
que la celula 1 descargo un PA en un instante 0. Este pico centrado alrededor de 2ms, nos indica
que la celula 2 presenta una altisima probabilidad de disparo cuando la celula 1 genera una espiga,
con un retardo de 1 o 2 ms. Esta misma cuantificacio en neuronas no acopladas, nos da una
funcion chata cercana a 0, indicando que no hay correlacion entre ambas neuronas.
-Otra funcion muy importante, es la capacidad de estos contactos de promover la actividad en
forma no solo sincronica sino que a demas automatica, que esta en la base de generacion de
ritmos cerebrales de actividad automatica y ritmica de algunos complejos neuronales. Ej: en
neuronas de la oliva inferior, las neuronas que se encuentran acopladas electricamente, una de las
consecuencias del acople electrico es el alto grado de sincronismo y ademas, las propias
oscilaciones y automatismo, surgen de esta modalidad de comunicacion neuronal. Presenta una
consecuencia funcional relevante, ya que la salida de este nucleo constituye una de las dos vias
mas importantes aferentes (via trepadora).
-TRANSMISION QUIMICA-
El mecanismo es netamente diferente, basado en un aspecto estructural en el cual NO hay un
nexo o contacto que permita un flujo directo de corriente (no hay continuidad citoplasmatica). La
estructura neuronal esta totalmente aislada (pre sinaptica) de la otra neurona (post sinaptica),
donde son independientes, no hay un flujo directo de corriente ni de moleculas. Tiene que existir
un mecanismo que permita que el mensaje contenido en el cambio del Vm de la celula pre
sinaptica se traduzca en un cambio del Vm de la celula post sinaptica para que ocurra el flujo de
informacion en los circuitos neuronales. El mediador en este caso es una sustancia quimica
almacenada en la terminal de la neurona pre sinaptica, y ante cambios del Vm en esta neurona, se
da la liberacion del neurotransmisor que se encontraba contenido en las vesiculas pre sinapticas, y
su liberacion va a interactuar con receptores especificos de la neurona post sinaptica para generar
un efecto variable, plastico , pero que implica un cambio del Vm.
-A nivel de la sinapsis quimica, la ultraestructura es totalmente diferente a la electrica (sustrato
estructural). Las uniones intercelulares se caracterizan por la presencia de vesiculas sinapticas con
presencia de zonas activas electrondensas, que resultan de la agregacion de un complejo
multimolecular complejo y critico para la funcion de liberacion y fusion exocitotica de vesiculas y
liberacion de su contenido vesicular. A nivel de la otra celula, se encuentra una densidad post
sinaptica (PSD) que tambien representa un agregado multimolecular denso y complejo entre los
que se destacan los receptores que reconocen en forma especifico el contenido de las vesiculas.
La diferenciacion morfologica entre el elemento pre y post sinaptico es el primer determinante al
hecho de que la transmision de informacion NO es direccional, sino que es unidireccional desde el
elemento pre sinaptico que contiene las vesiculas con el elemento neurotransmisor, hacia el
elemento post sinaptico en donde se encuentran los receptores post sinapticos que reconocen de
forma especifica dicho neurotransmisor.
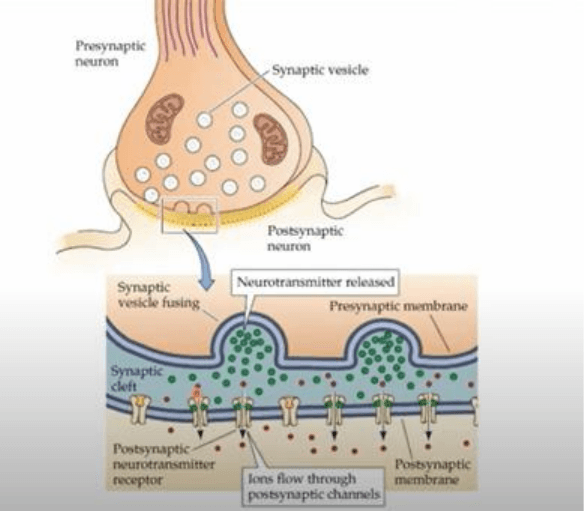
Desde el punto de vista
estructural, como sabemos, a nivel pre sinaptico encontramos las vesiculas que almacenan en
paquetes al neurotransmisor a una concentracion relativamente alta mediante mecanismos
activos. Ante la llegada de un PA a la terminal y la invasion del mismo, vemos que por un
mecanismo que depende de un flujo de Ca desde el medio extracelular al intracelular,
promoviendo la fusion exocitotica de las vesiculas con la membrana superficial, y por lo tanto, la
liberacion del contenido vesicular al medio extracelular. Una vez en el medio extracelular, por
difusion simple el contenido vesicular (neurotransmisor) difunde por el medio extracelular para
alcanzar los receptores post sinapticos anclados en la membrana de la celula post sinaptica. Estos
nt se unen a los receptores post sinapticos para promover la apertura de los mismos. Una vez que
se une el nt a sitios especificos del receptor se produce un cambio conformacional de los canales,
donde estos se abren y de acuerdo a su permeabilidad y selectividad especifica se da el flujo de
corriente (movimiento de carga) de acuerdo a su selectividad y se da el movimiento de iones
permeables. Estos canales denominados ionotropicos que median las acciones sinapticas quimicas
de tipo rapido, como el colinergico nicotinico, glutamato, AMPA y NMDA, los podemos concebir
como canales similares a los dependientes de voltaje, solo que en vez de ser sensibles a cambios
de potencial a traves de la membrana, estos son operados a traves de la presencia de un ligando
en el medio extracelular. Una vez que el neurotransmisor se unio al receptor (region especifica del
canal), este se abre y se establece un flujo de corriente entrante, donde el efecto en la celula post
sinaptica, sera una despolarizacion de su Vm. De alguna manera, vemos que una señal electrica
mediante un cambio de Vm es transformada en una liberacion de un mediador quimico, el cual a
su vez a nivel de la membrana post sinaptica, vuelve a generar una modificacion del potencial de la
membrana de la celula post sinaptica.
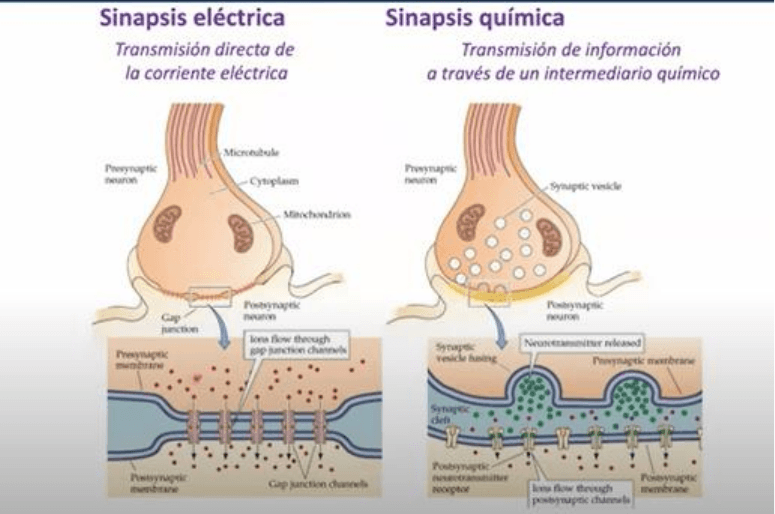
En la sinapsis electrica, la transmision es directa gracias a la presencia de cumulos de canales
intercelulares, que forman los contactos electricos (uniones GAP) donde la posicion estrecha de
ambas mebranas celulares permiten la interaccion y anclaje de los hemicanales para formar estas
vias de flujo de corriente directo entre una celula y la otra. En la transmision de tipo quimica NO
hay un flujo directo de corriente, sino que tenemos una señal electrica que desencadena la
liberacion del transmisor el cual a su vez interactua con un receptor especifico, que culmina con la
apertura de un canal y la generacion de una señal electrica en la celula post sinaptica. Al
neurotransmisor entonces lo consideramos un mediador de la transmision entre ambos elementos
neuronales. En la transmision electrica hay un flujo directo de corriente gracias a la presencia de
una via de baja resistencia (alta conductancia) y en la sinapsis quimica, la ausencia de esta via de
comunicacion directa requiere de la existencia de un mediador, que una vez que se despolariza la
terminal sinaptica, va a liberarse y modificar el Vm de la neurona post sinaptica. La modificacion
del Vm de una neurona modifica de alguna manera ya sea del flujo de corriente o la intervencion
de un mediador quimico, el Vm de la segunda celula del circuito.
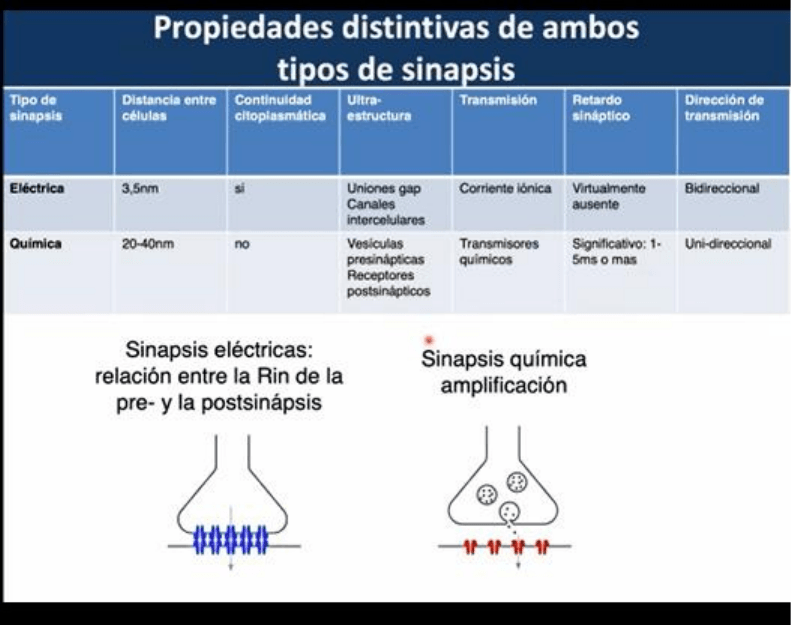
En la transmision quimica, depende de la existencia de un PA pre sinaptico y que el terminal pre
sinaptico sea invadido por este, ya que es la señal que dispara el ingreso de Ca al medio y la
liberacion del neurotransmisor. Mientras que un PA promueve la señalizacion a nivel post
sinaptico, una señal sub umbral que no alcance el umbral para que el Ca pueda ingresar, no va a
tener ningun efecto en la post sinapsis asi como tampoco una hiperpolarizacion de la celula pre
sinaptica. Se entiende que la transmision de tipo quimica es episodica dada que esta asociada
estrictamente a la generacion y ocurriencia de PA de la celula pre sinaptica.
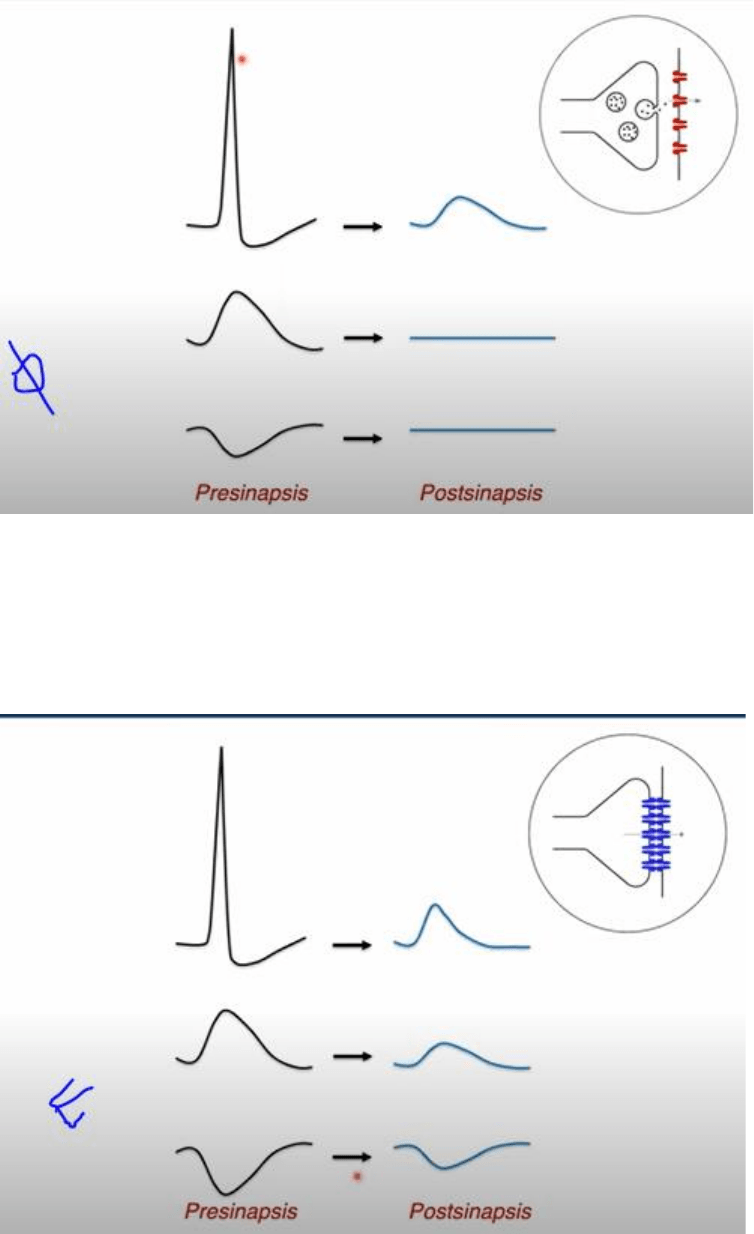
En contraposicion, la transmision electrica es mas continua. En un determinado rango de tiempo
donde no haya un cambio significativo de la cantidad de canales abiertos, la transmision opera de
forma continua. Durante un PA tenemos un potencial de acoplamiento con la celula post sinaptica,
pero no es indispensable la generacion de un PA dado que señales mas chicas van a generar
tambien una transmision de informacion. Cambios hiperpolarizadores del potencial de la celula
pre sinaptica tambien se reflejan cambios en el mismo sentido en la celula post sinaptica.
-LIBERACION DEL NEUROTRANSMISOR- SINAPSIS QUIMICA
La invasion de un PA en la terminal pre sinaptica induce la liberacion del contenido vesicular pre
sinaptico donde se almacena el neurotransmisor. Este nt es liberado y una vez en el espacio
sinaptico difunde hacia la membrana post sinaptica para interactuar con los receptores especificos
y generar un efecto a nivel de dicha membrana. Un cambio del Vm de la membrana pre sinaptica
es capaz de inducir entonces la liberacion de este neurotransmisor.
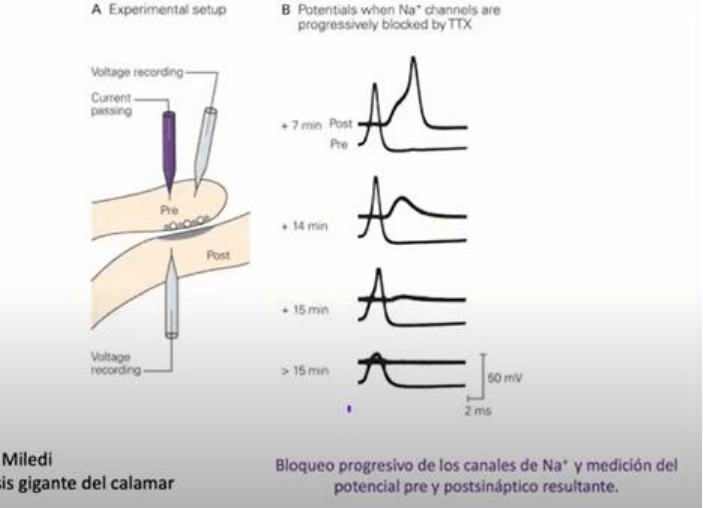
Los experimentos
realizados por investigadores en el axon gigante de calamar, se aplicaron pulsos de corriente para
despolarizar la membrana de la celula pre sinaptica simulando y sustituyendo un PA y elaborando
registros de los cambios del Vm ya sea un PA o de inyeccion de corriente de los electrodos. A nivel
de la celula post sinaptica se realizaron registros del Vm que permitieron monitorizar la cantidad
de transmisor liberado por la celula pre sinaptica. Las evidencias del PA pre sinaptico se
obtuvieron mediante experimentos farmacologicos donde los PA fueron bloqueados mediante la
aplicacion en el baño extracelular de la tetradotoxina (TTX). A concentraciones muy bajas TTX es
capaz de bloquear en forma altamente selectiva los canales de Na dependientes de voltaje que
estan involucrados y son los responsables en la generacion del PA en la celula pre sinaptica. Como
se ve en la secuencia de registros, en el trazado superior se ilustra el cambio del Vm de la celula
post sinaptica y en el inferior el cambio del Vm en la celula pre sinaptica. Se puede ver al inicio de
la aplicacion de TTX cuando aun no tuvo ningun efecto, un PA producido por la despolarizacion de
la corriente que fue inyectada por el electrodo, y produciendo a nivel de la celula post snaptica
con cierto retardo, un potencial post sinaptico excitador y a partir de un punto, este PPSE supera
el umbral para la generacion del PA y vemos una espiga. A medida que el tiempo transcurre y la
droga comienza a hacer efecto bloqueando los canales de Na, lo que vemos es que la amplitud de
la espiga comienza a reducirse progresivamente (pre sinaptica). Concomitantemente con la
reduccion de la amplitud de la espiga pre sinaptica, vemos una reduccion del potencial post
sinaptico excitador hasta que se encuentra totalmente bloqueado.
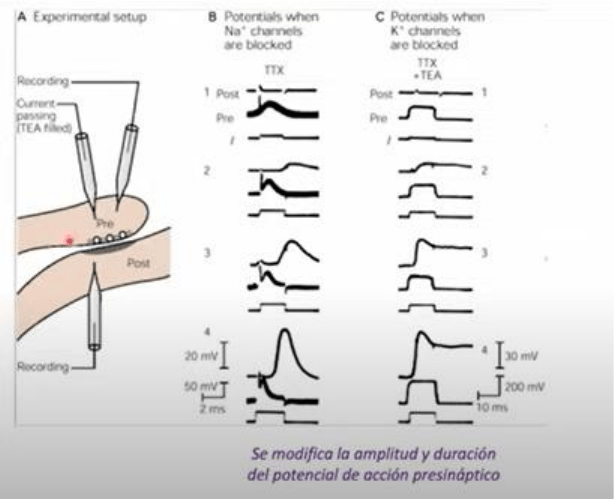
En el mismo preparado
experimental, en un preparado donde primero bloquearon los canales de Na mediante TTX, se le
aplico corriente con un electrodo en el estado de bloqueo total, y se muestra en el trazado inferior
de cada panel. En el trazado inferior observamos entonces el pulso de corriente aplicado con un
electrodo (i), el registro del Vm registrado con el otro microelectrodo en la celula pre sinaptica
(pre) y en la parte superior vemos el registro del Vm registrado con otro microelectrodo en la
celula post sinaptica. Los canales de Na en todo el registro estan completamente bloqueadas.
Conforme se incrementa la intensidad del estimulo de la corriente inyectada con el micro
electrodo pre sinaptico, vemos una respuesta en la membrana pre sinaptica que no puede ser
mediada por canales de Na dado que estos estan bloqueados por accion de TTX. A medida que la
corriente inyectada se incrementa progresivamente, vemos que esta respuesta se hace cada vez
mayor. A nivel de la celula post sinaptica, vemos que las inyecciones progresivamente mayores
generan una modificacion , es decir, un potencial post sinaptico excitador en la celula post
sinaptica. Estos experimentos estan indicando que la entrada de Na a traves de los canales de Na
vdep, NO ES NECESARIA PARA LA LIBERACION DEL NEUROTRANSMISOR Y POR LO TANTO PARA LA
GENERACION DE LOS POTENCIALES POST SINAPTICOS.
-Un experimento similar donde a demas de TTX se agrega TEA, que es un bloqueador de los
canales de K vdep (rectificador tardio) que media la repolarizacion del PA. Vemos 4 paneles en el
cual se aplica progresivamente un pulso de corriente a nivel pre sinaptico de intensidad
progresivamente mayor. La inyeccion produce una respuesta sostenida en el tiempo a nivel de la
celula pre sinaptica debido a la ausencia de mecanismos repolarizadores. A nivel post sinaptico, las
corrientes despolarizadoras inyectadas en la celula pre sinaptica de mayor intensidad, son capaces
de provocar un cambio en el Vm en la celula post sinaptica, indicando que hubo liberacion del
transmisor desde la celula pre hacia el espacio extracelular. Estos experimentos nuevamente
muestran que LO QUE IMPORTA PARA LA LIBERACION DEL NEUROTRANSMISOR ES LA
DESPOLARIZACION DE LA TERMINAL PRE SINAPTICA Y NO EL INGRESO DE Na NI LA SALIDA DE K.
POR LO TANTO,PARA LA LIBERACION DEL NEUROTRANSMISOR NO ES NECESARIO EL FLUJO DE
IONES Na NI DE K.
Este documento contiene más páginas...
Descargar Completo
2 SINAPSIS.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.