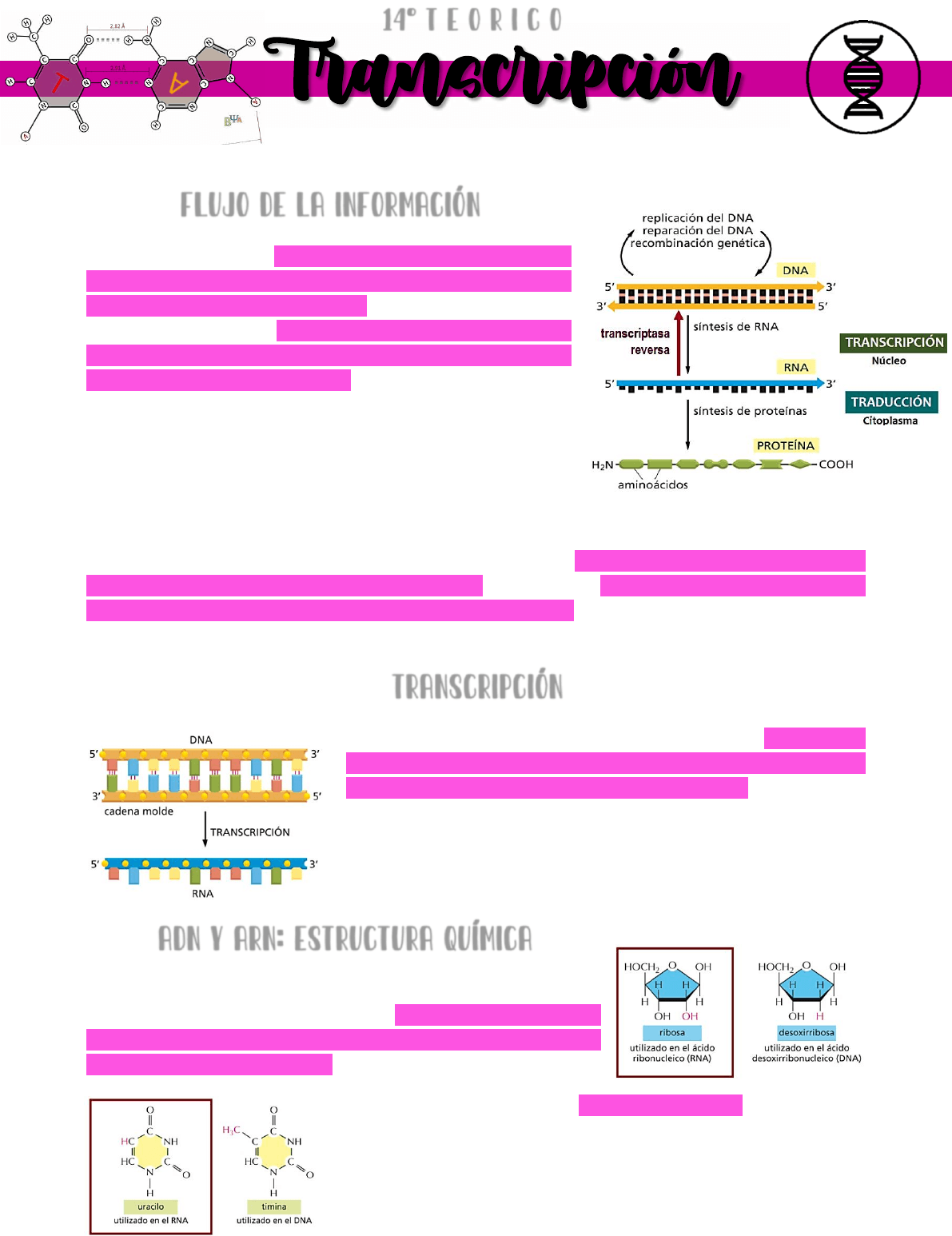
Flujo de la información
Crick estableció que a partir de una molécula de ADN
se puede sintetizar una molécula de ARN mediante un
proceso llamado TRANSCRIPCIÓN → el cual ocurre en el
NÚCLEO. Y además → partir de una molécula de ARN se
puede sintetizar una molécula de proteína mediante un
proceso llamado TRADUCCIÓN → el cual ocurre en el
CITOPLASMA.
Este flujo de la información genética → que va desde la
molécula de ADN al ARN y a proteínas → se conoce
como
DOGMA CENTRAL DE LA BIOLOGÍA.
Años más tarde se comprobó que el flujo de la información genética no era compatible
con el dogma central de la biología → se observa que a partir de una molécula de ARN
viral se puede sintetizar una molécula de ADN. Se aisló una ENZIMA LLAMADA TRANSCRIPTASA
REVERSA → EL PROCESO SE LLAMA TRANSCRIPCIÓN REVERSA. DOGMA CENTRAL DE LA BIOLOGÍA
AMPLIADO.
Transcripción
La transcripción es el proceso mediante el cual se copia un
determinado fragmento de la secuencia de nucleótidos de
ADN a una secuencia de nucleótidos de ARN. No se copia
todo el ADN sino un fragmento → esa parte copiada se
conoce con el nombre de
GEN → se copian nucleótidos de
ADN (desoxirribonucleótidos) a nucleótidos de ARN
(ribonucleótidos).
ADN y ARN: estructura química
Los nucleótidos de ARN son RIBONUCLEÓTIDOS → tienen
ribosa en lugar de desoxirribosa → la ribosa presenta en el
Carbono 2 un OH que hace a la molécula mucho más
reactiva que la desoxirribosa.
Los ARN contienen URACILO en lugar de Timina → si bien son
muy similares en su estructura, la timina tiene un grupo metilo
que carece el uracilo.
14° T E O R I C O

Tanto en el ADN como en el ARN → LOS NUCLEÓTIDOS SE UNEN POR ENLACES FOSFODIÉSTER.
U-A y T-A se unen mediante dos enlaces PdH → enlaces covalentes débiles que permiten
la estabilización de la estructura tridimensional de la molécula.
LAS DIFERENCIAS QUÍMICAS ENTRE EL ADN Y EL ARN SON MÍNIMAS.
El
ADN ES UNA DOBLE CADENA unida por complementariedad
molecular, estabilizada por PdH y fuerzas de Van der Waals →
forma una
DOBLE HÉLICE EN EL ESPACIO.
El
ARN ES UNA CADENA SIMPLE que
tiene la capacidad de
PLEGARSE
SOBE SÍ MISMA → los
plegamientos están estabilizados
por PdH y fuerzas de Van der
Waals → y le confieren a los distintos tipos de ARN
funciones estructurales y catalíticas que los
diferenciarán de otros ARN.
Principales tipos de ARN
Existen distintos tipos de ARN:
los más conocidos son el
ARN
MENSAJERO (ARNm)
→ que
codifica para proteínas; el
ARN RIBOSOMAL (ARNr) → que
forma parte de la estructura
de los ribosomas y catalizan
la síntesis de proteínas; y el
ARN DE TRANSFERENCIA (ARNt)
→ es importante en la síntesis
proteica ya que es un
adaptador entre la cadena
de ARN mensajero y los aa.
También hay ARN involucrados en el procesamiento de otros tipos de ARN → como los
ARN PEQUEÑOS NUCLEARES, los ARN PEQUEÑOS NUCLEOLARES y los ARN PEQUEÑOS DE CAJAL.
También hay ARN que intervienen en la regulación de la expresión génica → como los
MICROARN o los ARN DE INTERFERENCIA (este último anula la expresión de un gen).
Hay
ARN NO CODIFICANTE → participan en distintos procesos celulares como la síntesis de los
telómeros, la inactivación del cromosoma X y el transporte de proteínas al retículo.
TODOS LOS ARN SE SINTETIZAN POR UN PROCESO LLAMADO TRANSCRIPCIÓN → EL CUAL OCURRE EN
EL NÚCLEO DE LAS CÉLULAS EUCARIONTES.
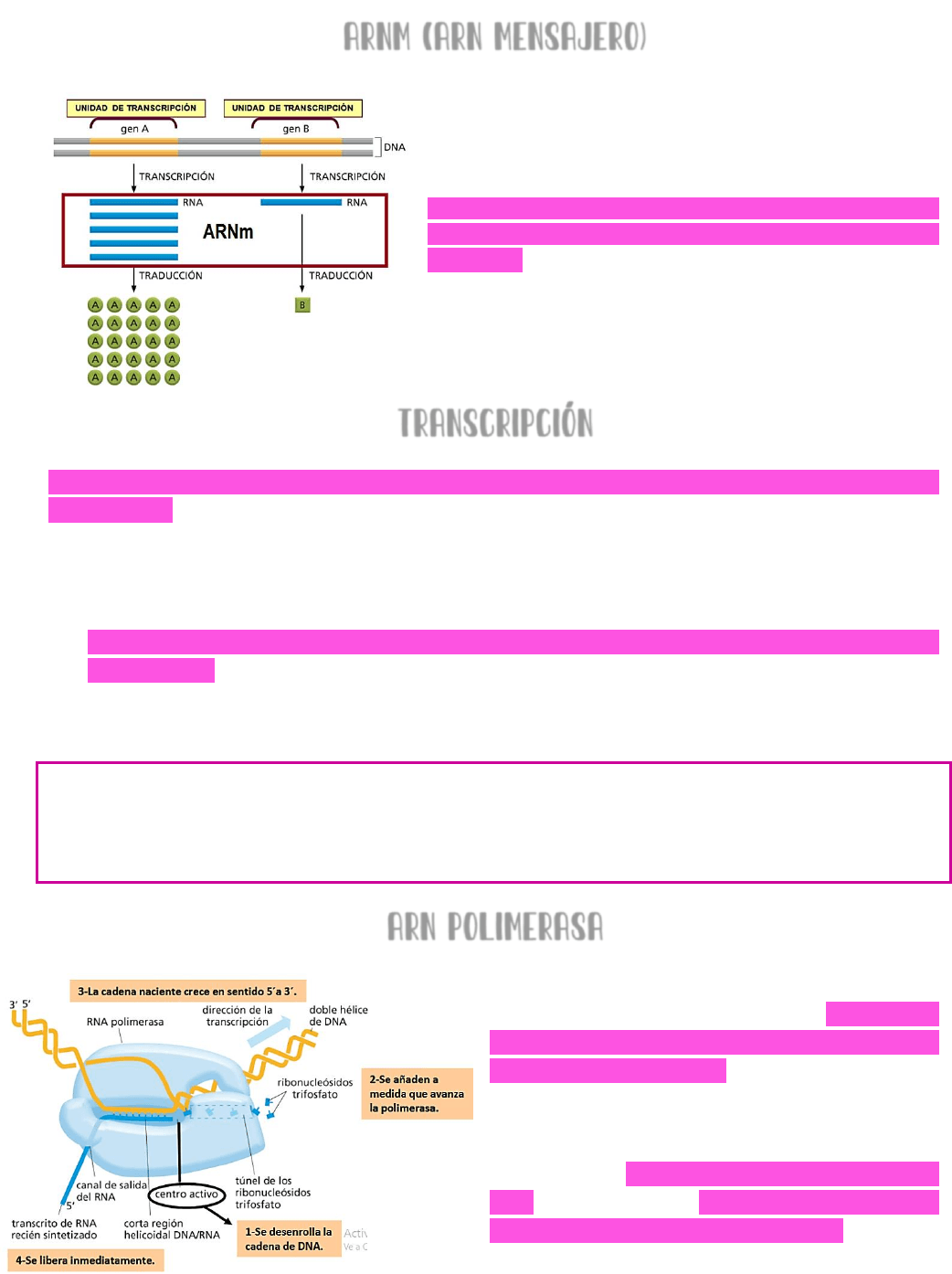
ARNm (ARN mensajero)
El ARN mensajero → se sintetiza por medio del
proceso de transcripción en el núcleo → a partir de
un fragmento de ADN (llamado gen o unidad
transcripcional) se copiará una molécula de ARN →
ESTA MOLÉCULA DE ARN MENSAJERO, EL CUAL CODIFICA
PARA PROTEÍNAS → EN EL CITOSOL SE TRADUCIRÁ A
PROTEÍNAS.
Todos los genes se pueden expresar con diferentes
eficiencias y esto va a depender del tejido en el
que estén y de la función que deba cumplir el gen.
Transcripción
La transcripción es el proceso mediante el cual un fragmento de la cadena de ADN se
copia a ARN.
1. Lo primero que debe suceder es la APERTURA Y EL DESENROLLAMIENTO de una zona de la
doble hélice del ADN.
2. Luego → UNA DE LAS DOS CADENAS DEL ADN ACTÚA COMO MOLDE PARA LA SÍNTESIS DEL ARN.
3. Los nucleótidos comenzarán a incorporarse por complementariedad de bases con la
hebra molde.
4. Proceso de ELONGACIÓN → una enzima, llamada ARN POLIMERASA, cataliza la
formación de
ENLACES FOSFODIÉSTER entre nucleótidos para elongar la cadena.
Durante la síntesis de la cadena de ARN → la cadena de ARN, al ser una cadena simple,
no va a permanecer unida por enlaces PdH a la hebra molde del ADN. En la replicación si
permanecen unidas. Las moléculas de ARN son mucho más cortas que las de ADN → por
eso se copia un fragmento del ADN y no toda la molécula.
ARN polimerasa
La ARN polimerasa es la enzima que CATALIZA
LA TRANSCRIPCIÓN IN VIVO → cataliza la
formación de enlaces fosfodiéster que unen
a los nucleótidos entre sí.
Esta enzima tiene
DISTINTAS SUBUNIDADES →
donde se encuentra un
SITIO ACTIVO → lugar
en el cual se desenrollará la doble hélice de
ADN; luego hay un túnel por donde van a
ingresar los ribonucleótidos trifosfato → se van
a ir añadiendo a medida que avanza la
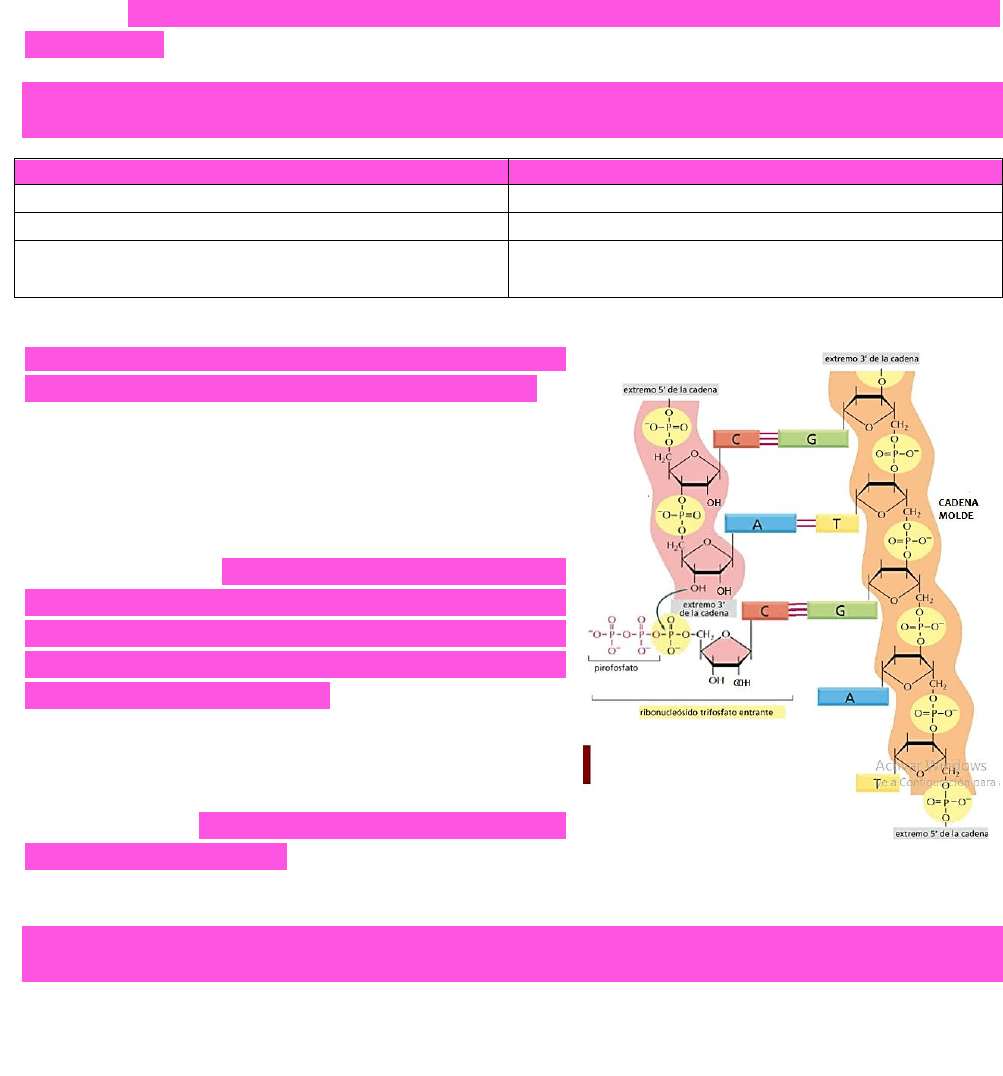
polimerasa y se van a ir uniendo por complementariedad molecular a la cadena molde →
luego, la cadena naciente de ARN recién sintetizado irá saliendo por un canal de salida
de la enzima.
LA ENERGÍA NECESARIA PARA IMPULSAR LA FORMACIÓN DE LOS ENLACES FOSFODIÉSTER SE OBTIENE
A TRAVÉS DE LA HIDROLISIS DE LOS ENLACES DE ALTA ENERGÍA QUE TIENEN LOS GRUPOS FOSFATOS.
ARN POLIMERASAS
ADN POLIMERASAS
Catalizan la unión de ribonucleótidos
Catalizan la unión de desoxirribonucleótidos
No necesitan cebador
Necesitan cebador
Cometen 1 error cada 10
4
nucleótidos
incorporados
Cometen 1 error cada 10
7
nucleótidos
incorporados
ESTAS ENZIMAS NO ESTÁN ESTRUCTURALMENTE RELACIONADAS → NO TIENEN EL MISMO ORIGEN.
LA INCORPORACIÓN DE NUCLEÓTIDOS EN LA CADENA
DE ARN SINTETIZADA SE DA A PARTIR DEL EXTREMO 3’ →
en este se tiene un OH el cual es capaz de atacar
al enlace fosfato del grupo fosfato en posición
alfa del nucleótido a incorporar → se produce la
hidrólisis del enlace, liberándose energía →
ESTA
ENERGÍA SERÁ UTILIZADA EN LA SÍNTESIS DEL ENLACE
FOSFODIÉSTER → la formación de un enlace
fosfodiéster es una reacción no espontánea →
necesita ser acoplada a una reacción
espontánea (como la hidrólisis del grupo fosfato)
para que la reacción ocurra.
LA SÍNTESIS DE ARN SE DA EN SENTIDO 5’ → 3’ YA QUE LA
CADENA CRECE POR EL OH DEL EXTREMO 3’.
Sin embargo → la lectura de la cadena molde es
inversa → es de 3’ → 5’; esto mismo ocurre en la
replicación del ADN.
LA CADENA DE ARN QUE ESTA SINTETIZANDOSE ES COMPLEMENTARIA Y ANTIPARALELA A LA HEBRA
MOLDE.
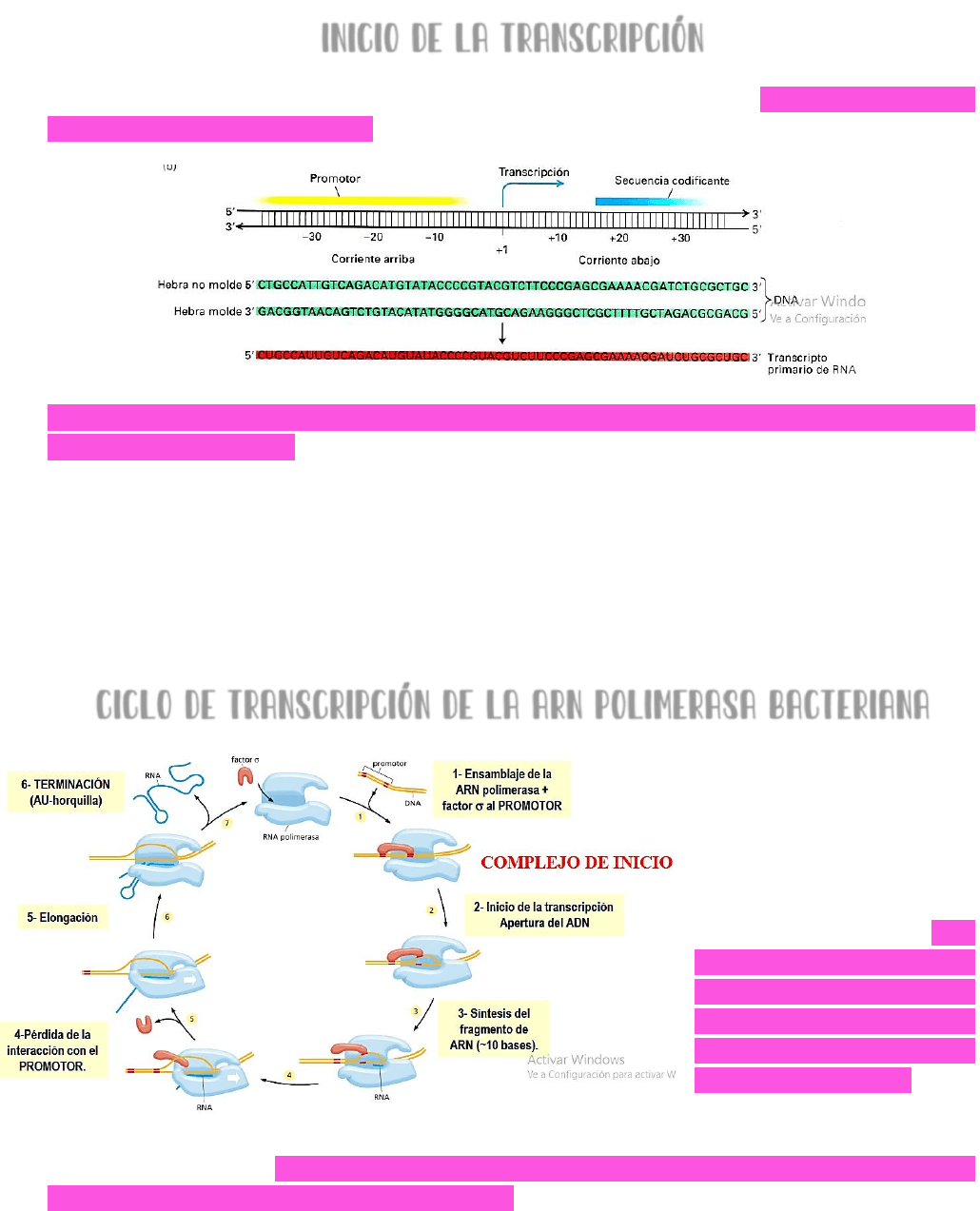
INICIO DE LA TRANSCRIPCIÓN
Existen en el ADN → secuencias especiales, llamadas PROMOTORES → que indican el punto
de inicio para la síntesis del ARN.
Estas secuencias se encuentran en corriente arriba de la posición +1 que es la posición de
inicio de la transcripción → y presentan:
una SECUENCIA DE DOBLE CADENA → ya que es una secuencia que forma parte del ADN
de doble cadena.
Presentan ASIMETRÍA.
Orientan a la polimerasa en el sentido en que debe realizarse la transcripción.
Determinan la velocidad de transcripción.
CICLO DE TRANSCRIPCIÓN DE LA ARN POLIMERASA BACTERIANA
En los PROCARIOTAS → el
proceso de transcripción
es mucho más sencillo que
en los eucariontes.
Se tiene un solo tipo de
ARN polimerasa → esta
enzima se ENSAMBLA AL
PROMOTOR ayudada por un
factor de inicio de la
transcripción conocido
como FACTOR SIGMA → se
forma el
COMPLEJO DE
INICIO DE LA TRANSCRIPCIÓN
→ luego se inicia la transcripción con la apertura de las
cadenas de ADN → en el sitio activo de la enzima se incorporan los nucleótidos y se
comienza la síntesis del fragmento de ADN → este fragmento tiene 10 bases y es un
proceso muy lento debido a que la polimerasa está anclada al sitio del promotor.
SI LA
TRANSCIPCIÓN ES EFICIENTE → OCURRE UNA PERDIDA DE LA INTERACCION CON EL PROMOTOR → SE
DESENSAMBLA EL FACTOR SIGMA Y LA POLIMERASA PUEDE AVANZAR Y CONTINUAR CON LA
ELONGACIÓN DE LA CADENA
→ el proceso de elongación continua hasta que en la cadena

molde de ADN se lea una secuencia de terminación → CUANDO LA POLIMERASA LEE LA
SECUENCIA DE TERMINACIÓN → se detiene la transcripción, se desprende la cadena de ARN
transcripta del ADN molde y se da por finalizada la transcripción.
ARN POLIMERSAS EN CÉLULAS EUCARIOTAS
En EUCARIONTES → se tiene 3 tipos de ARN polimerasas involucradas en el proceso de
transcripción → que trascriben a los distintos tipos de ARN.
ARNm → la polimerasa involucrada en su proceso de transcripción es la ARN
polimerasa II.
En eucariontes se necesita un
CONJUNTO DE FACTORES GENERALES DE TRANSCRIPCIÓN para
que se inicie el proceso → mientras la polimerasa bacteriana solo necesita el factor σ.
Además, en eucariotas también se debe resolver el problema del
EMPAQUETAMIENTO DEL
ADN
.
LOS SEGMENTOS DE ADN QUE NO DEBEN TRANSCRIBIRSE SE ENROLLAN SOBRE SÍ MISMOS EN UN
PROCESO QUE SE CONOCE COMO CONDENSACIÓN DE LA CROMATINA.
Sin embargo → aquellas hebras de ADN que se están
transcribiendo representan a la cromatina menos
condensada (cromatina laxa).
Imagen con hematoxilina y eosina → se observan
núcleos con cromatina laxa y núcleos con cromatina
condensada.
FACTORES GENERALES DE LA TRANSCRIPCIÓN
Los factores generales de la transcripción ayudan en cada uno de los procesos necesarios
durante la transcripción → facilitan la correcta colocación de la ARN polimerasa II sobre el
promotor; ayudan a separar las dos hebras del ADN y liberan a la ARN polimerasa del
promotor permitiendo la elongación de la cadena.
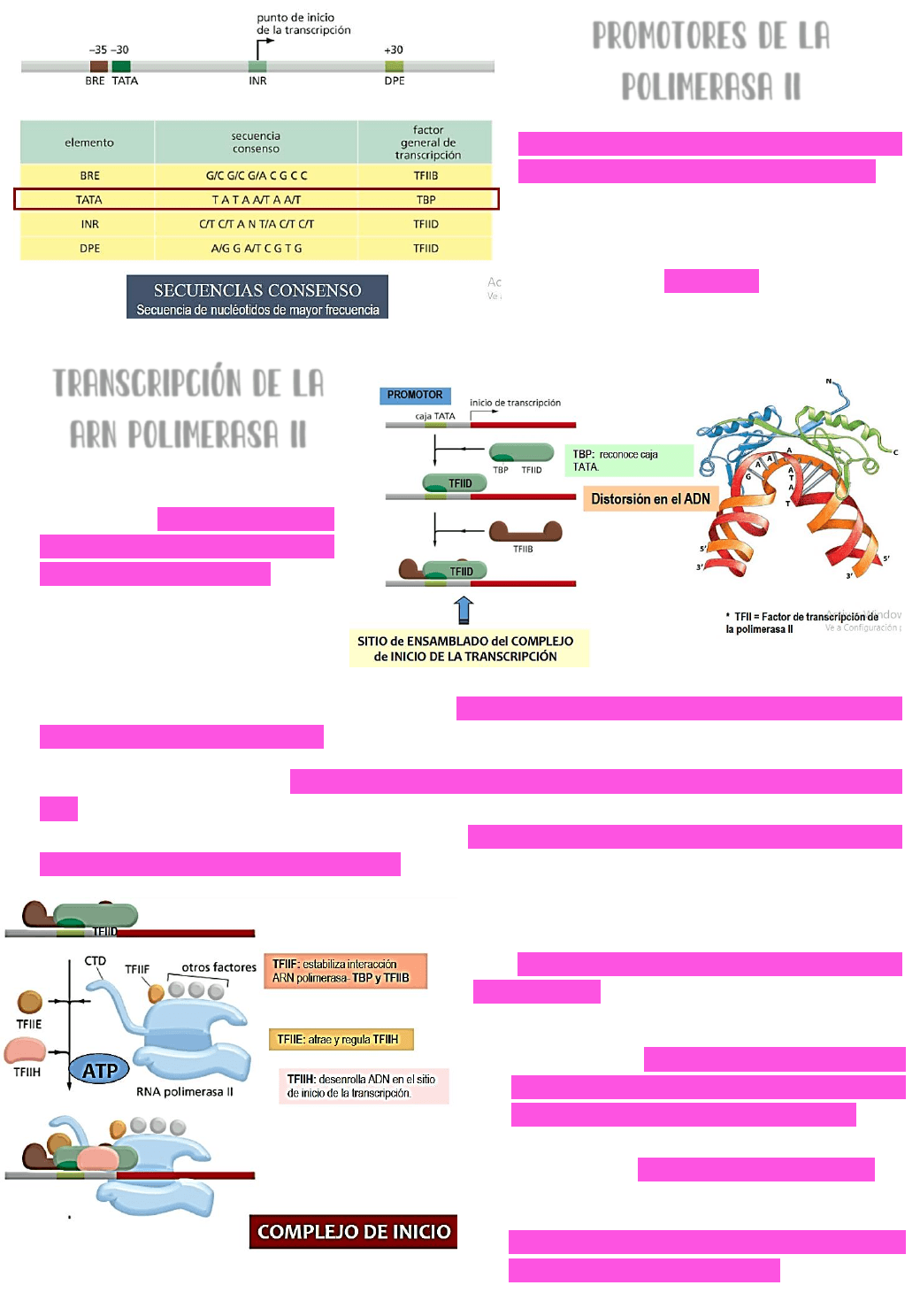
PROMOTORES DE LA
POLIMERASA II
Hay varios promotores que pueden ser
reconocidos por la ARN polimerasa II →
la mayoría de estos promotores se
ubican
CORRIENTE ARRIBA DEL PUNTO DE
INICIO DE LA TRANSCRIPCIÓN
. El más
conocido es la CAJA TATA.
TRANSCRIPCIÓN DE LA
ARN POLIMERASA II
En el gen a transcribir → se
identifica el sitio de inicio de la
transcripción y la zona del
promotor (caja TATA). Una
PROTEÍNA DE UNIÓN A LA CAJA
TATA
(TBP) reconocerá y unirá
a otro factor de transcripción
(factor de transcripción de la polimerasa II D →
TFIID) → este factor de transcripción junto
con la proteína de unión a caja TATA → reconocerán a la caja TATA y se unirán a la
secuencia promotora del gen.
Esta unión provocará una DISTORSIÓN EN EL ADN QUE FACILITARA LA APERTURA DE LA CADENA DE
ADN. Luego → el factor de transcripción de la polimerasa 2 B (TFIIB) se unirá al complejo
que se estaba formando en el promotor → esto va a marcar el sitio de ensamblado del
complejo del inicio de la transcripción.
La
ARN POLIMERASA 2 se unirá al sitio promotor
junto con los otros factores de transcripción
→ y reclutará a más factores de inicio de la
transcripción como:
el factor de inicio de la polimerasa 2F
(TFIIF) → es el encargado de estabilizar la
interacción entre la ARN polimerasa y los
factores de inicio de la transcripción.
el factor de inicio de la polimerasa 2E
(TFIIE) → que atrae y regula al factor H.
el factor de inicio de la polimerasa 2F
(TFIIF
) → es el encargado de estabilizar la
interacción entre la ARN polimerasa y los
factores de inicio de la transcripción.
el factor de inicio de la polimerasa 2E
(TFIIE) → que atrae y regula al factor H.
Factor H (TFIIH) → es el encargado de
desenrollar la cadena de ADN en el sitio
de inicio de la transcripción → o sea, en
el sitio activo de la ARN polimerasa II.
DE ESTA MANERA QUEDA FORMADO EL COMPLEJO DE INICIO DE LA TRANSCRIPCIÓN.
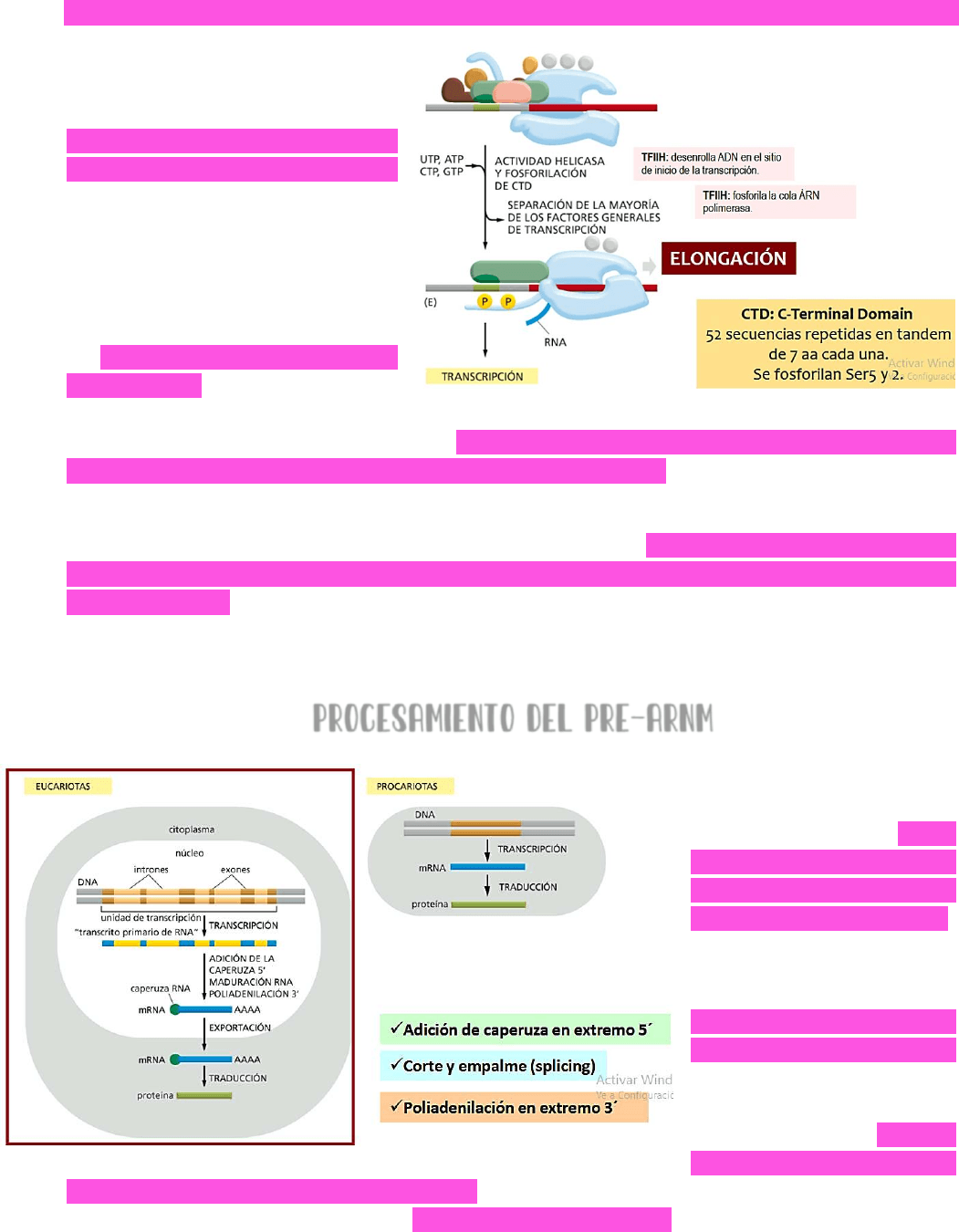
Una vez formado el complejo de
iniciación →
LA POLIMERASA
COMIENZA LA TRANSCRIPCIÓN
→ el
factor H va a desenrollar al ADN
en el sitio activo de la polimerasa
→ ingresarán los nucleótidos y se
va a sintetizar la cadena de ARN.
Al principio, como ocurre en
procariontes →
ESTA SÍNTESIS ES UN
PROCESO SUMAMENTE LENTO
ya que
la polimerasa está anclada al
sitio promotor. Luego → el factor
H va a fosforilar la cola de ARN polimerasa → permitiendo que se desensamblen los
factores de inicio de la transcripción → AL DESENSAMBLARSE DEL COMPLEJO, LO QUE OCURRE ES
QUE LA POLIMERASA PIERDE LA INTERACCIÓN CON EL PROMOTOR → esto le va a permitir a la
POLIMERASA AVANZAR Y ELONGAR LA CADENA DE ARN.
El DOMINIO C TERMINAL de la cola de la ARN polimerasa → son 52 secuencias repetidas en
tándem de 7 aminoácidos cada una y que pueden fosforilarse en las posiciones de la
Serina5 y Serina2 → estas fosforilaciones son importantes para permitir el desensamblaje de
los factores de inicio de la transcripción y QUE ASÍ LA CADENA PUEDA ELONGARSE; pero
también dirigen el procesamiento del pre ARNmensajero que se está sintetizando.
PROCESAMIENTO DEL PRE-ARNM
En PROCARIOTAS se tiene un
gen que se transcribe a un
ARN mensajero → y ese
ARN mensajero sin ninguna
otra modificación → se va
a traducir en una proteína.
Sin embargo → en
EUCARIONTES se tiene un
gen formado por intrones
(no codificantes) y exones
→ el gen se transcribe a un
ARN (
TRANSCRIPTO
PRIMARIO) → en la
transcripción se
transcriben tanto intrones como exones → pero para obtener una proteína que sea
funcional, se necesita que los intrones sean eliminados → ese ARN necesita un
procesamiento.
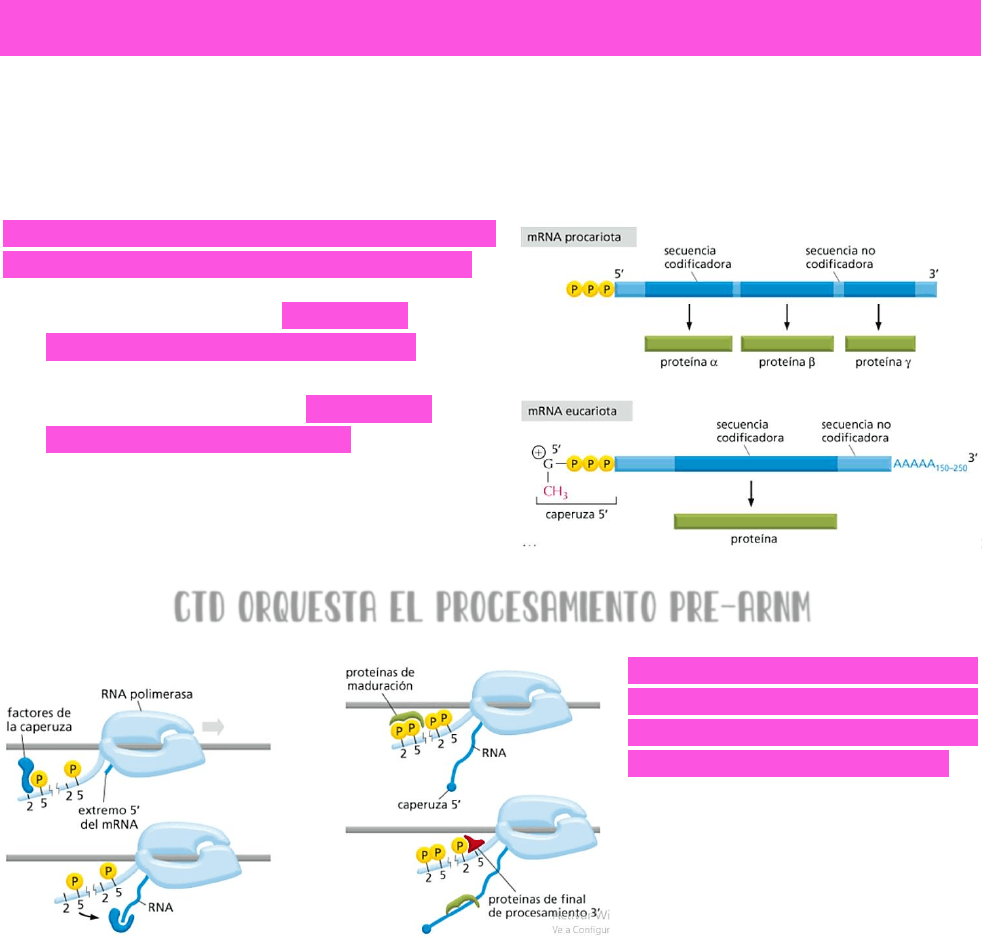
EL ARN MENSAJERO DEBE SUFRIR UNA SERIE DE PROCESOS PARA LLEGAR A SER UN ARN MADURO QUE
LUEGO PUEDA SER TRADUCIDO A UNA PROTEÍNA:
se le debe adicionar una CAPERUZA en el extremo 5’.
Se deben cortar los intrones y empalmar los exones → PROCESO DE CORTE Y EMPALME O
SPLICING
.
Debe sufrir la POLIADENILACIÓN en el extremo 3’ → COLA POLI-A.
Una diferencia importante entre ARN
mensajeros procariontes y ARN eucariontes:
un ARN mensajero procarionte puede
codificar para diferentes proteínas.
los ARN mensajeros eucariontes solo
codifican para una proteína.
CTD ORQUESTA EL PROCESAMIENTO PRE-ARNM
Las fosforilaciones en el dominio
C terminal de la cola de la ARN
polimerasa van a orquestar el
procesamiento del pre ARNm →
estas fosforilaciones ocurren en
las serinas en posición 2 y en
posición 5.
Si se FOSFORILAN LAS SERINAS EN POSICIÓN 5 → se reclutarán los factores necesarios para
la adición de la caperuza.
Si además están FOSFORILADAS LAS SERINAS EN POSICIÓN 2 → se reclutarán proteínas
necesarias para la maduración de ese ARNm → o sea, proteínas que intervendrán en
el corte y empalme.
Y además → si se DESFOSFORILAN LAS SERINAS EN POSICIÓN 5 → se reclutarán proteínas
que intervengan en el final del procesamiento 3’ → es decir, en la poliadenilación del
extremo 3’ del ARN mensajero.
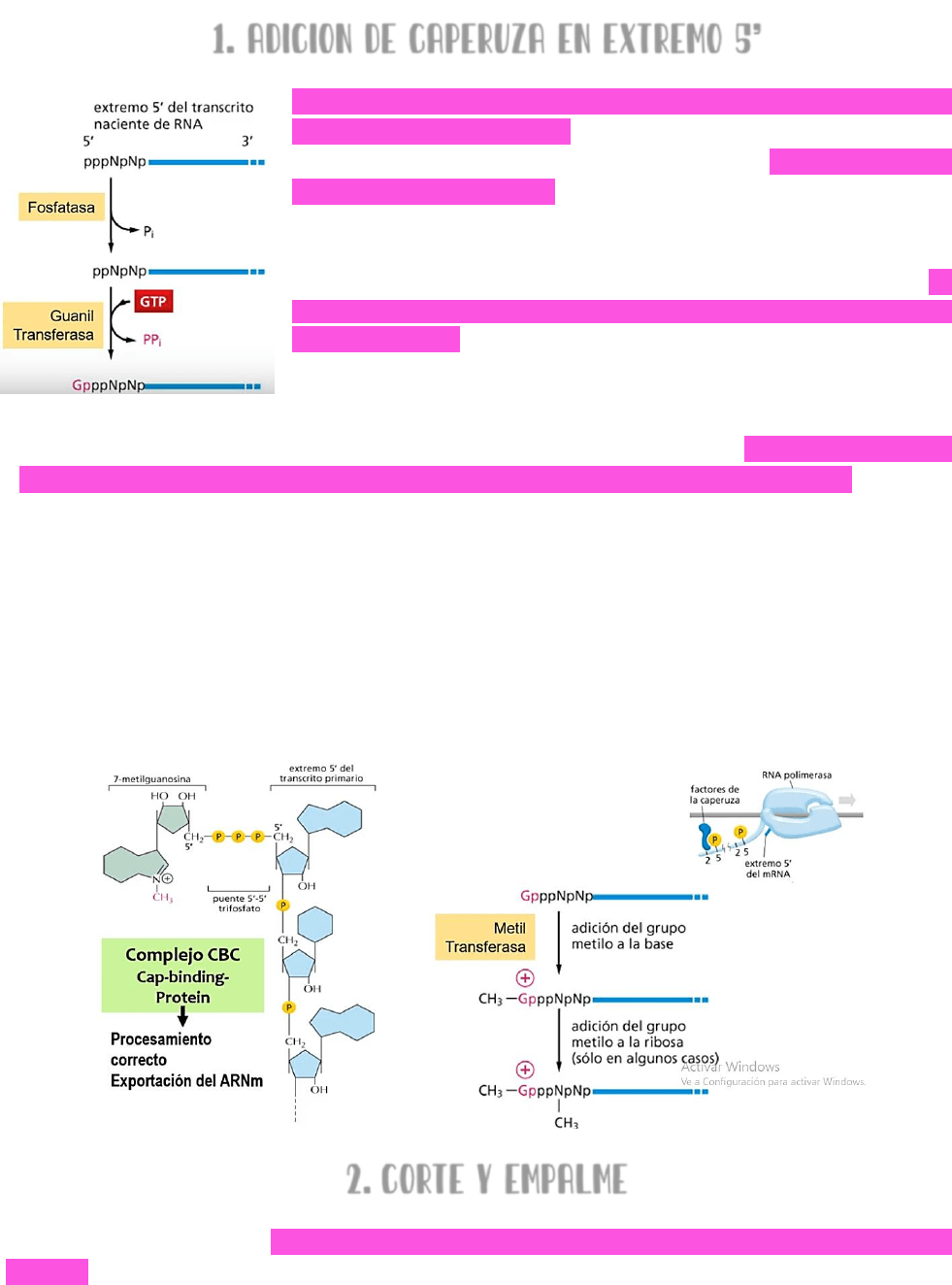
1. ADICION DE CAPERUZA EN EXTREMO 5’
La cola de la ARN polimerasa se fosforila en las serinas
ubicadas en posición 5 → lo cual recluta los factores
necesarios para la adición de la caperuza. El extremo 5’ del
ARN naciente esta libre → por medio de una fosfatasa se
escindirá un grupo fosfato de ese extremo; mediante una
GUANILILTRANSFERASA se le añadirá una guanosina; y por una
METILTRANSFERASA se unirá un grupo metilo en la posición 7. DE
ESTA MANERA, EL EXTREMO 5’ DEL ARN NACIENTE INCORPORA UNA 7-
METILGUANOSINA → el tipo de unión entre la guanosina y el
transcriptor primario será 5’ → 5’.
También, a la 7-metilguanosina → se le añadirá un
COMPLEJO CBC (complejo proteico →
proteína de unión a la caperuza) → esta unión con el complejo CBC PERMITE EL CORRECTO
PROCESAMIENTO DEL ARN MENSAJERO Y SU CORRECTA EXPORTACIÓN HACIA EL CITOSOL.
LA ADICIÓN DE LA CAPERUZA:
protege el ARNm de la posible degradación por medio de enzimas como
exonucleasas.
va a indicar cuál es el extremo 5’; cuando el ARNm llegue al citosol debe ser
reconocido por los ribosomas → y el extremo 5’ debe ser identificado por los
ribosomas para iniciar así la traducción del mensajero.
La caperuza permite que se pueda distinguir al ARNm de otros tipos de ARN.
2. CORTE Y EMPALME
Otro procesamiento que debe sufrir el pre-ARNm para transformarse en un ARN mensajero
maduro → es el corte de intrones y el empalme de exones. Para ello → LA COLA DE LA ARN
POLIMERASA SE DEBE FOSFORILAR EN LAS SERINAS EN POSICIÓN 2 Y 5
→ esto recluta a las
proteínas necesarias para este proceso.
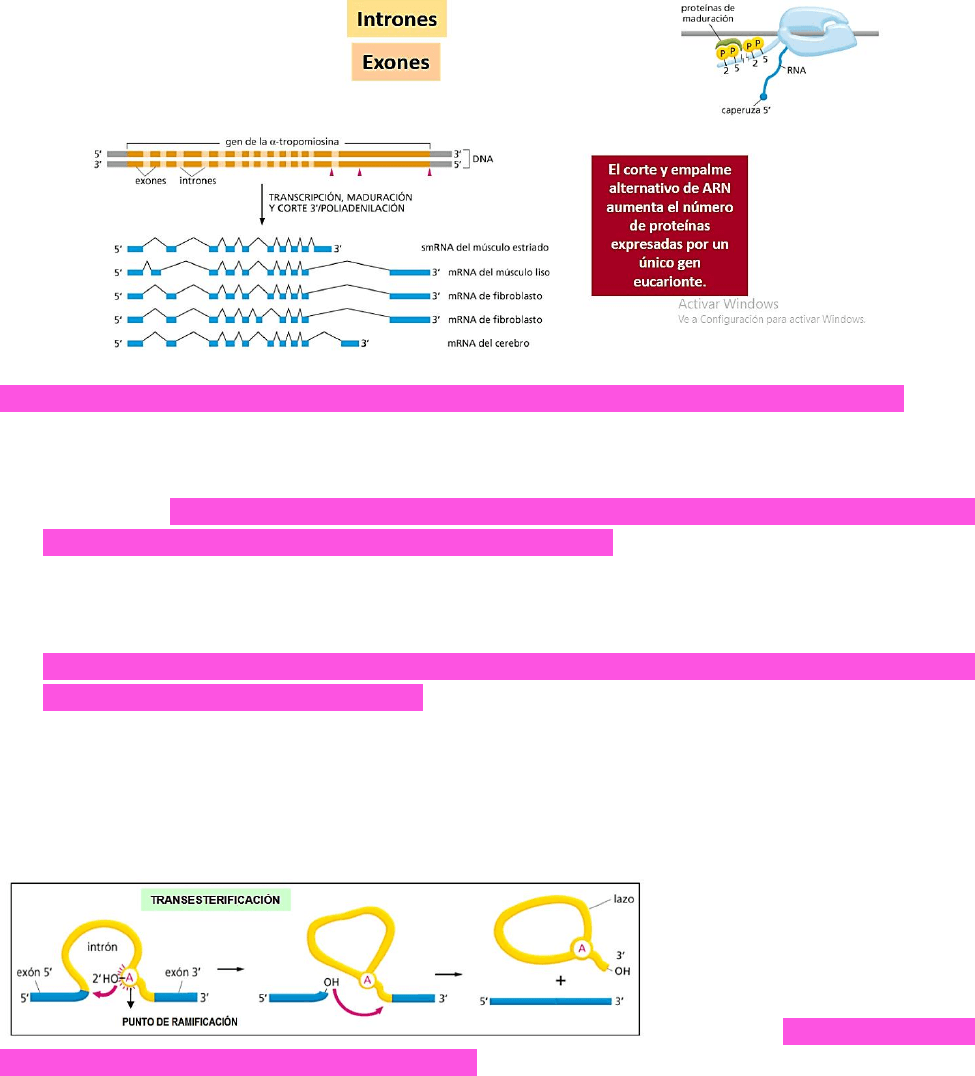
Los genes eucariontes tienen intrones a pesar de ser secuencias no codificantes → los
intrones son secuencias muy largas y cumplen varías funciones:
como el ADN es el material genético, cuando la polimerasa produce la replicación
del ADN → muchos de sus errores pueden caer en estas secuencias de intrones y por
lo tanto transformarse en MUTACIONES SILENCIOSAS → o sea, mutaciones que no
afectarán y podrá obtenerse una proteína normal.
Los transcriptos de muchos genes se pueden cortar y empalmar de maneras diferentes
dando lugar a proteínas distintas de acuerdo al tejido del que se encuentren y
siempre partiendo del mismo gen → este es el caso de la alfa tropomiosina que,
depende del tejido en el que se encuentre, se obtendrán distintas isoformas de la
misma proteína.
Corte y empalme alternativo del ARN → AUMENTA EL NÚMERO DE PROTEÍNAS EXPRESADAS POR
UN ÚNICO GEN EUCARIONTE
.
REACCIÓN DE MADURACIÓN
DEL PRE-ARNm
Esquema → en azul están
los exones y en amarillo el
intrón → EN ESTE INTRÓN VA
A OCURRIR UN REORDENAMIENTO DE LAS BASES → lo cual dejará una adenina desapareada
(conocido como
PUNTO DE RAMIFICACIÓN) → se expondrá un oxhidrilo en posición 2, el cual
es muy reactivo y atacará la unión entre el exón 5’ y el intron. Se produce el corte y queda
libre el exón 5’ → el cual ahora expone un OH libre, el cual ataca a la unión entre el intron
y el exón 3’ → producirá el corte y se desprenderá el intron →
EL CUAL LUEGO SERÁ
DEGRADADO
.
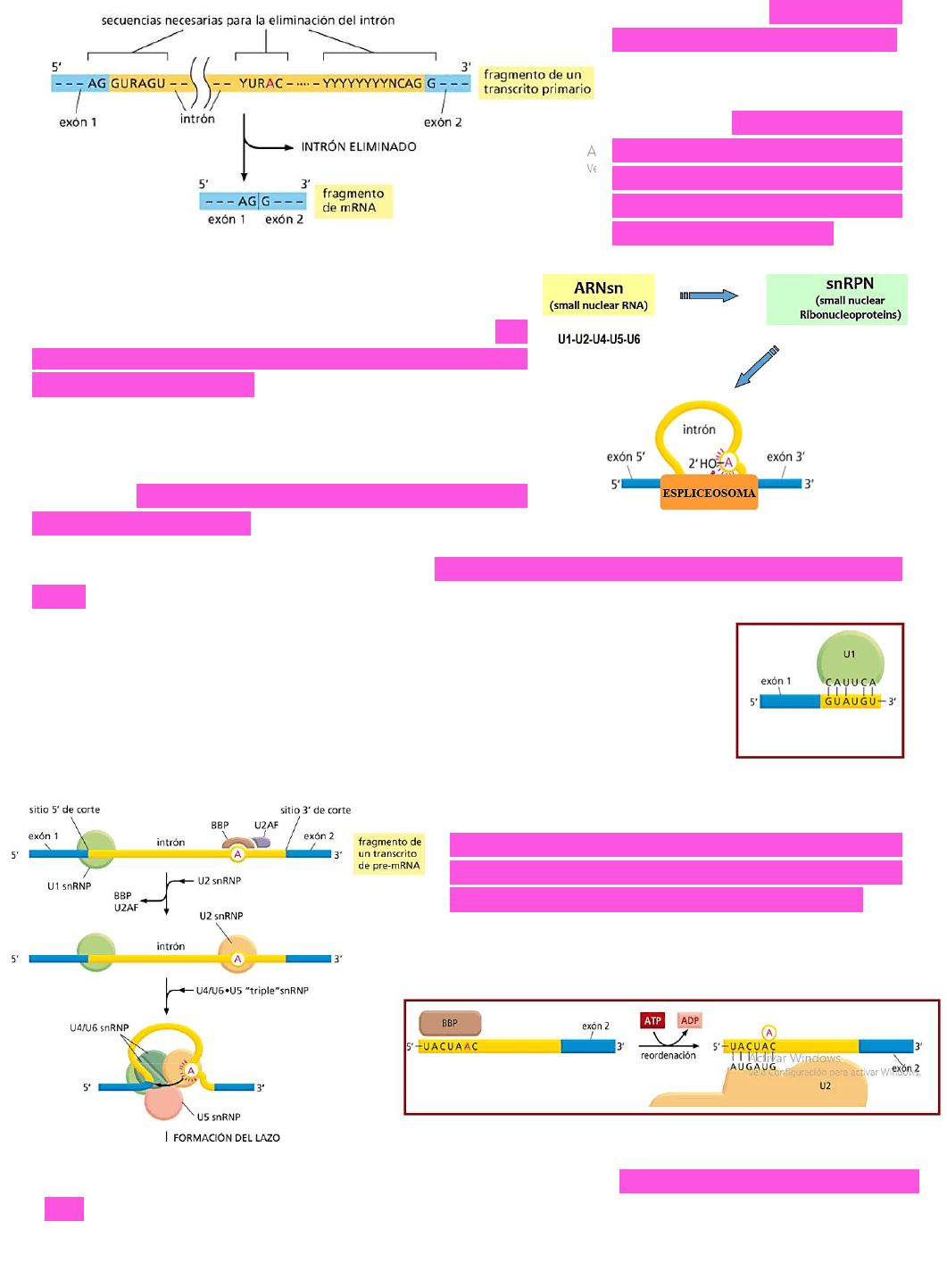
Por otro lado → el exón 5’ y el
exón 3’ se van a empalmar.
¿Cómo se sabe cuáles son las
secuencias que se deben
cortar? → estas secuencias
son generalmente secuencias
consenso que indican cuales
deben ser los sitios de corte
entre el exón y el intrón.
EL CORTE DE INTRONES Y EL EMPALME DE LOS EXONES ES
UN PROCESO CATALIZADO POR ARN
→ muchos ARN
tienen funciones catalizadoras. En este caso → LOS
ARN PEQUEÑOS NUCLEARES SON LOS ENCARGADOS DE
CATALIZAR EL SPLICING → estos ARN pequeños
nucleares se unen con proteínas formando las
RIBONUCLEOPROTEÍNAS PEQUEÑAS NUCLEARES → esto
forma un complejo llamado
SPLICEOSOMA → este
complejo CATALIZARÁ EL CORTE DE INTRONES Y EL
EMPALME DE LOS EXONES.
Existe una RIBONUCLEOPROTEÍNA (U1) que reconoce el sitio de corte entre el exón 1 y el
intron → y se une a este sitio en una forma ENERGÉTICAMENTE FAVORABLE ya que es capaz
de unirse por complementariedad de bases.
Por otro lado → también hay una
PROTEÍNA DE UNIÓN AL SITIO DE
RAMIFICACIÓN
→ que reconoce la adenina que luego queda
desapareada y se une a ese sitio.
Luego → otra ribonucleoproteína (
U2)
desplaza a la proteína de unión al sitio de
ramificación y se unirá a ese sitio en un
proceso energéticamente desfavorable →
necesita ATP → ya que en este proceso va a
quedar desapareada una adenina.
Luego se unirán otras ribonucleoproteínas → U4, U5 y U6 → contribuirán a la formación del
lazo.
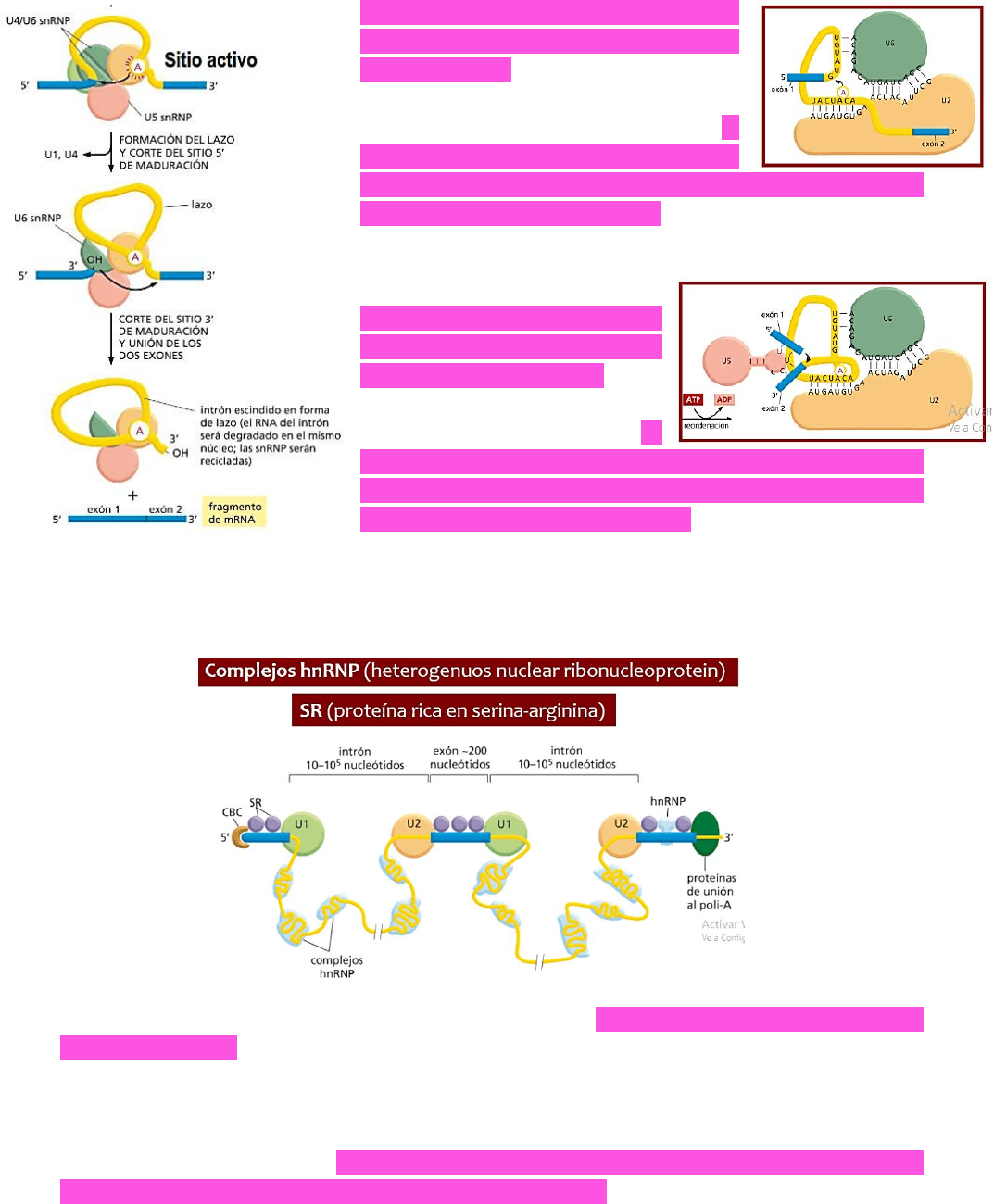
Queda entonces determinado un sitio
activo, que es la adenina
deseapareada → esta andenina
desapareada atacará al sitio de
corte entre el exón 5’ y el intrón → la
adenina produce el corte entre el
exón 5’ y el intrón y de esta manera queda un OH libre
en el exón 5’ en la posición 3’ → este OH libre atacará al
sitio de unión entre el exón 3’ y el intrón.
Se produce el corte y luego,
otra ribonucleoproteína (U5)
permitirá que se acerquen los
dos exones (el 3’ y el 5’) y por
medio de gasto energético
(HIDRÓLISIS DEL ATP) → se
unirán ambos exones y de esta manera queda el
fragmento de ARN mensajero formado por los dos
exones y el lazo queda libre → pero luego será
degradado en el núcleo; por su parte, las ribonucleoproteínas serán recicladas.
¿CÓMO SE ESCOGEN CORRECTAMENTE LOS SITIOS DE MADURACIÓN? Existen dos tipos de
proteínas:
proteínas ricas en serina-arginina → PROTEÍNAS SR → reconocen exones y evitan que
sean degradados → PROTEGEN A LOS EXONES.
COMPLEJOS hnRNP (ribonucleoproteínas nucleares heterogéneas) → se unirán a los
intrones (intrones son secuencias muy largas que tienden a enrollarse) →
EVITAN QUE
LOS INTRONES SE ENROLLEN. Estas proteínas le indican a las ribonucleoproteínas que
forman parte del espliceosoma donde deben ubicarse.
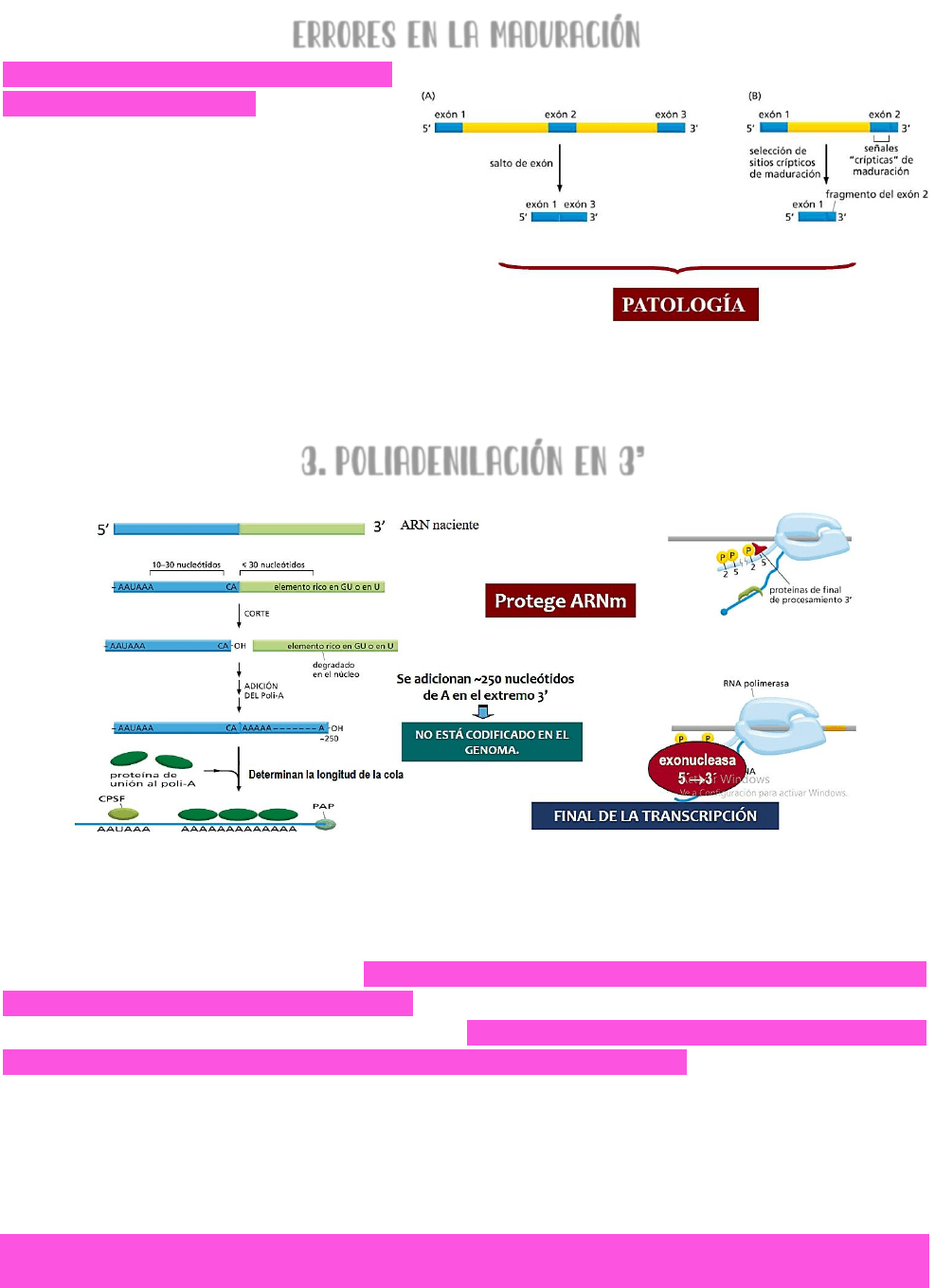
ERRORES EN LA MADURACIÓN
Durante la maduración del ARNm se
pueden cometer errores → se puede
saltar un exón (esquema A) o se
pueden introducir en un exón señales
de corte que no deberían estar y por
lo tanto se tiene un fragmento de un
exón (esquema B).
TODOS LOS ERRORES
QUE SE SUCEDAN DURANTE LA
MADURACIÓN DEL ARNM DAN COMO
RESULTADO PROTEÍNAS ANÓMALAS QUE
PUEDEN LLEVAR A UNA PATOLOGÍA
.
3. POLIADENILACIÓN EN 3’
EL PRE-ARNM SUFRE UN PROCESO DE POLIADENILACIÓN EN EL EXTREMO 3’ → para ello, la cola de
la ARN polimerasa se debe desfosforilar en las serinas que están en posición 5’ → esto va a
reclutar proteínas necesarias para la poliadenilación.
El ARN que se va transcribiendo → hacia el final de la transcripción tendrá una secuencia
consenso rica en adeninas y uracilos → de 10 a 30 nucleótidos posteriores a esta
secuencia presenta una secuencia de corte. Allí se produce el corte, se libera el ARN de la
polimerasa y el ARN quedará con la caperuza en el extremo 5’ → lo que impedirá su
degradación por parte de las exonucleasas.
EN EL EXTREMO 3’ → una POLI-A POLIMERASA va a agregar una cola de adeninas; y al mismo
tiempo se añadirán unas proteínas llamadas
PROTEÍNAS DE UNION A LA POLI-A → estas
proteínas van a determinar la longitud de la cola.
APROXIMADAMENTE → SE ADICIONARAN UNAS 250 ADENINAS EN EL EXTREMO 3’ → ESTO NO ESTÁ
CODIFICADO EN EL GENOMA Y SIRVE PARA PROTEGER AL ARNM DE LA ACCIÓN DE EXONUCLEASAS.
Por otro lado → la polimerasa sigue sintetizando ARNm ya que todavía está unida al ADN
→ esa cadena de mensajero que sigue sintetizando la polimerasa → es una cadena rica
en Guaninas y Uracilos → sin embargo, como carece del capuchón en el extremo 5’ →
será rápidamente degradado por exonucleasas y esto determinará el final de la
transcripción → SE DESENSAMBLARÁ EL COMPLEJO Y LA POLIMERASA IRÁ A TRANSCRIBIR OTRO
ADN
.
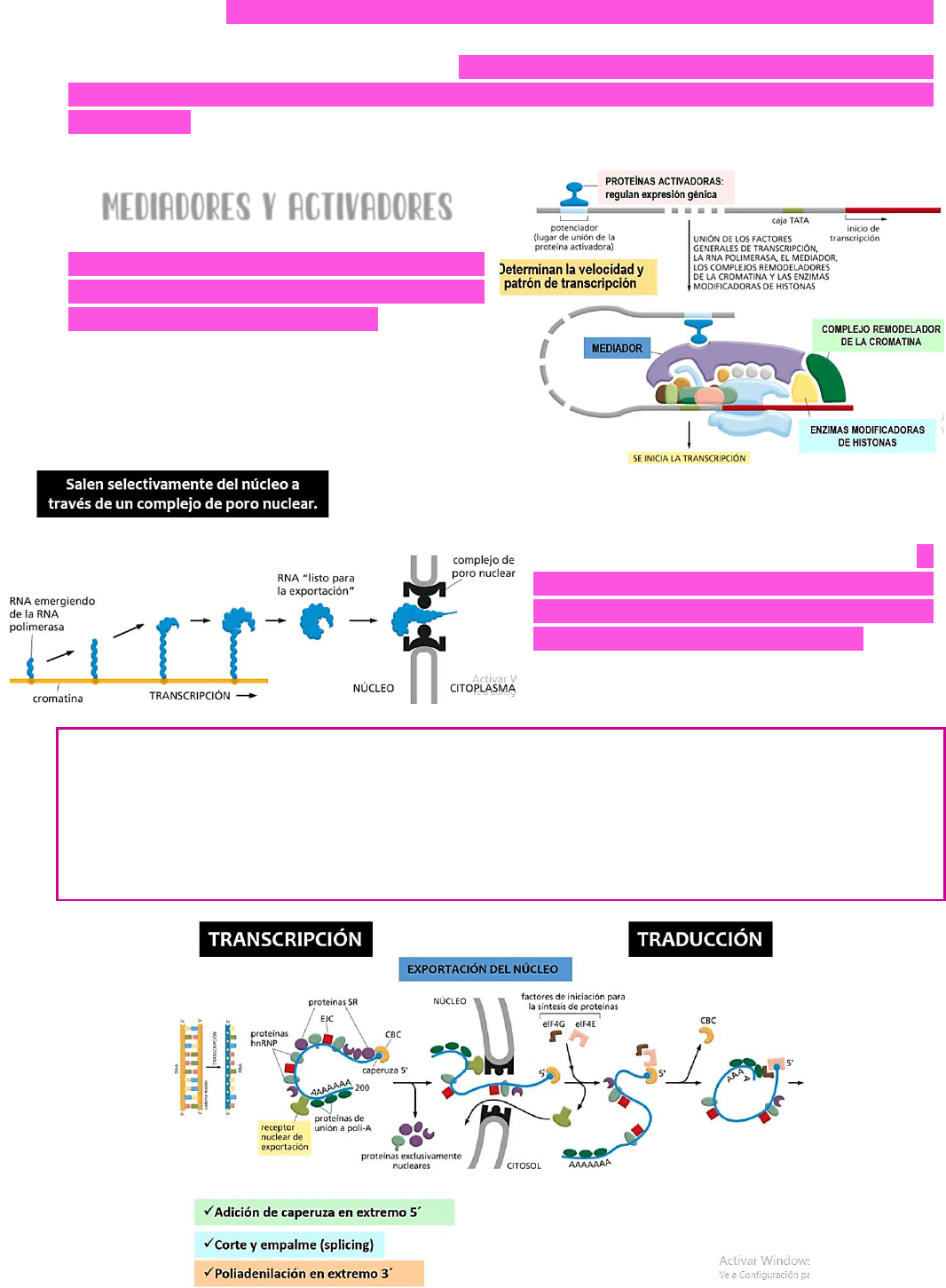
MEDIADORES Y ACTIVADORES
EL proceso de transcripción está muy
regulado por PROTEÍNAS que son
MEDIADORAS O ACTIVADORAS → por
ejemplo, hay proteínas que son
activadoras y que regulan la expresión
génica y determinaran la velocidad y el
patrón de transcripción.
¿CÓMO SALEN DEL NÚCLEO LOS ARNM
MADUROS? Una vez que el pre-ARNm fue
procesado correctamente y está listo
para ser exportado al citoplasma → el
ARNm sale selectivamente del núcleo a
través de un complejo que se conoce
como COMPLEJO DEL PORO NUCLEAR → este
proceso de exportación del ARNm del
núcleo al citoplasma
REQUIERE DE ENERGÍA.
Entonces →
TRANSCRIPCIÓN → un fragmento de ADN se transcribe a un pre-ARNm → este
sufre un procesamiento en el cual se le adhiere una caperuza en el extremo 5’; se cortan
los intrones y se empalman los exones y se produce el agregado de la cola poli-A en el
extremo 3’. Este ARNm maduro será exportado al citoplasma a través del complejo del
poro nuclear y en el citoplasma será reconocido por ribosomas y se iniciará el proceso de
traducción.

30avo teo CICLO CELULAR 1.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.