
Bioquímica | Ruta de las pentosas fosfato, gluconeogénesis y síntesis de ácidos grasos
Gluconeogénesis
La gluconeogénesis (“formación de azúcar nuevo”) es la síntesis de glucosa a partir de
precursores no carbohidratos.
La glucosa en sangre es la principal fuente de energía en los tejidos, principalmente en el
cerebro que requiere más de 120 g de glucosa al día. Después de haber sido ingerida, la
glucosa es oxidada a piruvato mediante el proceso de glucólisis en el hígado, y el exceso es
almacenado como glucógeno mediante la glucogenogénesis (síntesis de glucógeno). Durante
el ayuno el hígado libera glucosa a la sangre de forma que los tejidos dependientes de glucosa
no sufran por pérdida de energía.
Hay dos mecanismos involucrados en este proceso: la glugenólisis y la gluconeogénesis
Los niveles de glucosa en sangre también deben ser mantenidos durante el ejercicio, y las
células musculares toman glucosa de la sangre para oxidarla y proveer de energía. Las
hormonas, particularmente insulina y glucagón, dictan si la glucosa fluye a través de la
glucólisis o si las reacciones van en la dirección opuesta y se produce glucosa vía glucogénesis.
¿Dónde ocurre la gluconeogénesis? en el hígado
90%; y en el riñón 10%
La mayoría de la glucosa producida en la corteza renal
es usada por la médula renal, pero una parte entra a la
circulación sanguínea.
Resumiendo, la glucogénesis es el proceso por el
cual la glucosa es sintetizada a partir de precursores
no carbohidratos y ocurre principalmente en el
hígado en condiciones de ayuno.
¿Cuáles son los precursores gluconeogénicos?
Los precursores no carbohidratos que pueden ser
convertidos a glucosa incluyen los productos de la
glucólisis lactato y piruvato.
También incluyen al esqueleto carbonado de la mayoría
de los aminoácidos glucogénicos y también a partir
del glicerol proveniente de los triglicéridos.
Precursor: PIRUVATO
Los aminoácidos, el lactato, y el Propionil-CoA proveniente de los
ácidos grasos de cadena impar, convergen en el Oxalacetato, que
es un intermediario del ciclo de Krebs de alta energía, ya que, su
descarboxilación exergónica provee la energía necesaria para la
síntesis del fosfoenolpiruvato.
Los ácidos grasos con número impar de átomos de C producen
Propionil-CoA. Estos carbonos pueden ser precursores de glucosa,
pero es una fuente menor. El propionil-CoA es convertido en Succinil-
CoA (intermediario del ciclo de Krebs) que puede usarse para la
gluconeogénesis. Los otros carbonos de la
cadena impar dan Acetil-CoA, a partir del
cual NO se puede sintetizar glucosa. Por otra
parte, el glicerol pasa directamente a
transformarse en dihidroxiacetona fosfato.

Bioquímica | Ruta de las pentosas fosfato, gluconeogénesis y síntesis de ácidos grasos
El piruvato se produce en el hígado a partir de precursores gluconeogénicos
como el lactato y la alanina.
Precursor: LACTATO
El precursor lactato entra en el hígado, donde es oxidado por la lactato
deshidrogenasa a piruvato, el cual es convertido en glucosa por la
glucogénesis. La glucosa generada al pasar a la sangre es absorbida por el
músculo esquelético, de esta manera, el hígado suministra glucosa para la
contracción muscular.
El músculo obtiene ATP al convertir la glucosa en
lactato. El lactato pasa a la sangre y vuelve al hígado.
Este ciclo de reacciones constituye lo que se llama el
CICLO DE CORI.
Precursor: AMINOÁCIDO
En cuanto a los aminoácidos como precursores, vienen
principalmente del “pool” de aminoácidos en el músculo
donde pueden ser obtenidos a partir de la degradación
de proteínas.
La alanina es el mayor AA gluconeogénico y se produce
en el músculo a partir de otros AAs y luego el destino final
es la glucosa. También hay otros AA como Ser, Thr que
también darán origen al piruvato, terminando al final en la
síntesis de glucosa. Algunos AAs forman intermediarios
del ciclo de Krebs (ej. succinil-CoA; fumarato) los cuales
a su vez pueden también generar glucosa.
La leucina (Leu) y la lisina (Lys) son los únicos AA que no
pueden ser convertidos en Oxalacetato, porque su
ruptura da lugar a Acetil-CoA, y no hay rutas para poder
convertir Acetil-CoA a Oxalacetato y por tanto a glucosa.
Esos dos AAs son aminoácidos cetogénicos, es decir,
son capaces de sintetizar cuerpos cetónicos.
La alanina como el lactato es un precursor importante de la glucosa en el músculo, se forma
del piruvato por transaminación en una reacción catalizada por la Alanina Aminotransferasa.
La reacción inversa tiene lugar en el hígado. A este ciclo de reacciones le llamamos CICLO DE
LA ALANINA
Estas reacciones de transaminación pueden darse para otros AA con α-cetoglutarato para dar
el α-cetoácido correspondiente. La Alanina producida en el músculo es capaz de generar
glucosa en el hígado.
Bioquímica | Ruta de las pentosas fosfato, gluconeogénesis y síntesis de ácidos grasos
Precursor: GLICEROL
Se produce liberándose de las reservas adiposas de triglicéridos dando ácidos grasos y glicerol
catalizado por la lipasa. Esos ácidos grasos sufrirán beta oxidación y formarán Acetil-CoA (no
es precursor de la glucosa).
Este glicerol es luego transformado en dihidroxiacetona fosfato entrando en la vía
gluconeogénica para la síntesis de glucosa. Por otro lado, el glicerol también es capaz de
generar NADH transformándose en dihidroxiacetona fosfato.
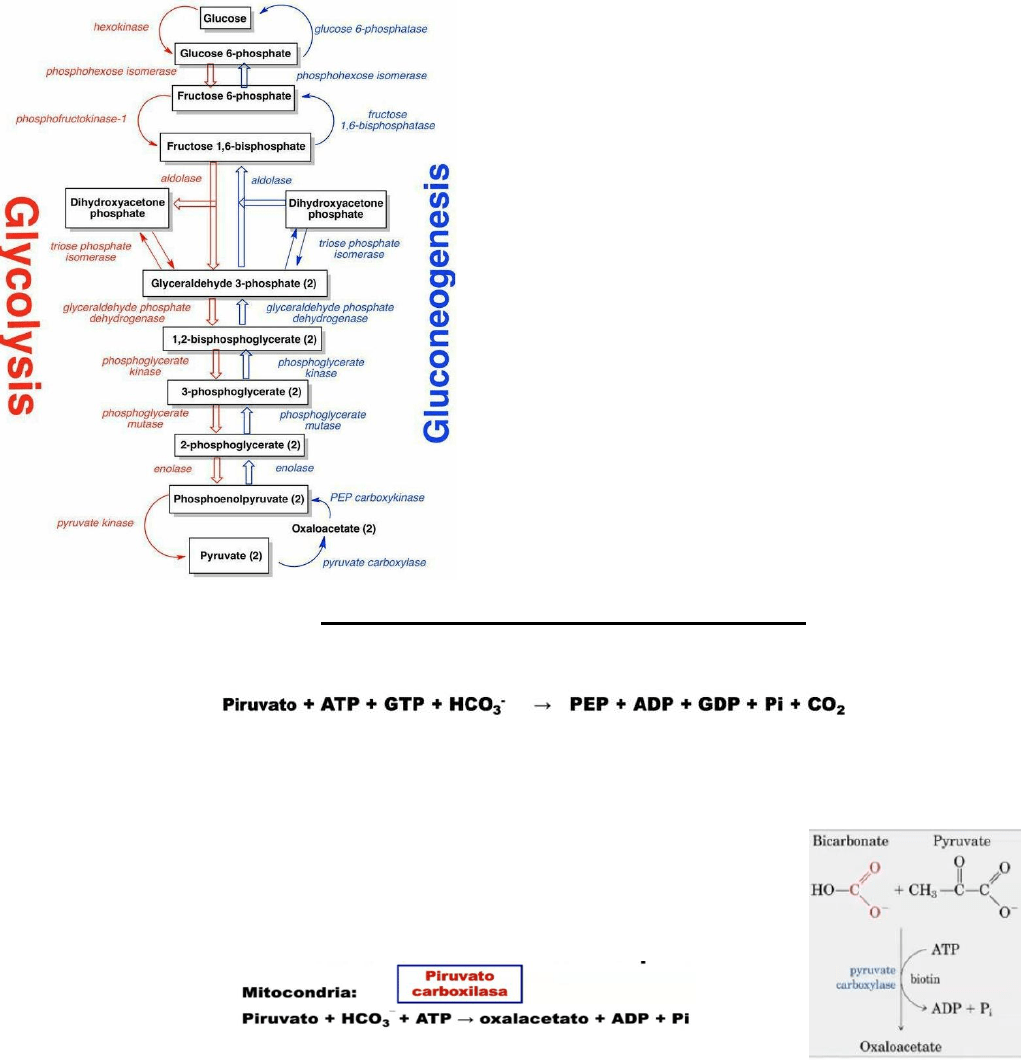
Glucolisis / Gluconeogénesis
Importante recordar que las reacciones 1, 3 y 10 de la glucólisis son muy exergónicas en
condiciones estándar e irreversibles.
La mayoría de las reacciones de la gluconeogénesis son
reacciones inversas de la glucólisis, sin embargo, las
enzimas hexoquinasa, fosfofructoquinasa (PFK-1) y la
piruvato quinasa catalizan reacciones con cambio de
energía muy negativo y deben ser reemplazadas en la
glucogénesis por reacciones que hagan la síntesis de
glucosa termodinámicamente favorable.
1-
Hexoquinasa / glucosa-6-fosfato
3- Fosfofructoquinasa-1 / Fructosa-1,6-bifosfatasa
10- Piruvato quinasa / piruvato carboxilasa + PEP
carboxiquinasa
En la glucólisis todas las reacciones se daban en el
citosol. En el caso del gluconeogénesis, algunas
reacciones son citosólicas, pero otras enzimas, como, por
ejemplo, la glucosa-6-fosfatasa está unida al retículo
endoplásmico, y la piruvato carboxilasa es una enzima
mitocondrial.
También podemos tener otras enzimas que estén en la
mitocondria como la fosfoenolpiruvato carboxiquinasa,
que puede ser mitocondrial o citosólica.
Reacciones de “rodeo” de la gluconeogénesis
1. Conversión de Piruvato en Fosfoenolpiruvato
En la glucólisis, el fosfoenolpiruvato (PEP) se convierte en piruvato mediante la acción de la
piruvato quinasa. Mientras, en la gluconeogénesis, el piruvato se transformará en PEP por la
acción de dos enzimas: la piruvato carboxilasa y la fosfoenolpiruvato carboxiquinasa.
En la mitocondria, el piruvato se carboxilará por medio del
bicarbonato (HCO
3
-
, generado a partir del CO
2
) usando ATP,
convirtiéndose en Oxalacetato y liberando Pi.
Esta reacción es catalizada por la piruvato carboxilasa que usa
biotina.

Bioquímica | Ruta de las pentosas fosfato, gluconeogénesis y síntesis de ácidos grasos
La reacción en que el oxalacetato pasa a convertirse en
fosfoenolpiruvato es catalizada por la fosfoenolpiruvato
carboxiquinasa y puede darse en citosol y en la mitocondria.
Además, para poder darse esta reacción se usa GTP como fuente
de energía, así como el grupo fosfato para el PEP.
La gluconeogénesis necesita NADH en el citosol, según el precursor hay dos caminos:
➔ Malato deshidrogenasa
➔ Lactato deshidrogenasa
En la imagen, podemos observar que el oxalacetato
generado a partir del piruvato no puede atravesar la
membrana mitocondrial. El oxalacetato depende de
equivalentes de reducción en el citosol para transportarse
a través de la membrana mitocondrial como malato o
aspartato.
En el citosol se necesita tener NADH para reducir el 1,3-
bifosfoglicerato a gliceraldehido-3-fosfato durante la
gluconeogénesis.
Para que el malato o el aspartato atraviesen la membrana
mitocondrial se vale de la Lanzadera Malato-Aspartato
(resumen fosforilación oxidativa). En esta lanzadera, el
malato sale de la de la mitocondria y en el citosol se
convierte en oxalacetato, produciendo el NADH requerido
para que la síntesis de glucosa pueda darse.
El oxalacetato que sale al citosol mediante la lanzadera, es
convertido en PEP por la fosfoenolpiruvato
carboxiquinasa citosólica.
Si el precursor de glucosa es lactato el transporte de malato está inhibido, debido a que
ya se genera NADH en el citosol cuando el lactato se transforma en piruvato en la reacción
catalizada por la Lactato deshidrogenasa, siendo luego convertido en oxalacetato por la
piruvato carboxilasa.
Los siguientes pasos a partir del fosfoenolpiruvato generado en este primer “rodeo” ocurren en
el citosol, en pasos reversibles de la glucólisis hasta llegar al gliceraldehído-3-fosfato que se
isomeriza en dihidroxiacetona-fosfato, se condensan y forman la fructosa-1,6-bifosfato.
2. Conversión de Fructosa 1,6 bifosfato en Fructosa 6-fosfato
La enzima fructosa 1,6-bisfosfatasa (FBPasa-1) libera fosfato inorgánico de la fructosa-1,6-
bifosfato convirtiéndola en fructosa 6-fosfato. Por lo tanto, NO es la reacción inversa de la
catalizada por la PFK-1, no se produce ATP al remover el fosfato del C-1 de la fructosa 1,6-
bifosfato porque es un enlace fosfato de baja energía.
En la siguiente reacción de la glucogénesis la
fructosa 6-fosfato se convierte a glucosa-6-fosfato
por la misma isomerasa usada en la glucólisis, la
fosfoglucosa isomerasa.
Lanzadera
Malato-
Aspartato

Bioquímica | Ruta de las pentosas fosfato, gluconeogénesis y síntesis de ácidos grasos
3. Conversión de Glucosa-6-fosfato en Glucosa.
La
glucosa-6-fosfatasa hidroliza el fosfato liberando Pi desde
la glucosa-6-fosfato dando glucosa que puede ser liberada a
la sangre. NO es la reacción inversa, en este caso, a la
Hexoquinasa, NO se genera ATP. El enlace fosfato en la
glucosa-6-fosfato es un enlace de baja energía.
La glucosa-6-fosfatasa está asociada al retículo endoplasmático, además de usarse en la
gluconeogénesis también puede producir glucosa sanguínea por ruptura del glucógeno.
Balance de la Gluconeogénesis
Reacciones secuenciales de la gluconeogénesis empezando por piruvato
Durante las reacciones gluconeogénicas se rompen 6 enlaces de fosfato de alta energía, 4
del ATP y 2 del GTP. Se necesitan 2 piruvato para la síntesis de 1 mol de Glucosa. Se requiere
energía en forma de equivalentes de reducción, como el NADH, los cuales se usan en la
reacción de 1,3-bifosfoglicerato para dar gliceraldehido 3-fosfato.
Se aprecia, que en la gluconeogénesis se usan 2 ATP y 2 GTP más de lo que se usaría en la
reacción inversa de la glucólisis.
Regulación de la Gluconeogénesis
Además de ocurrir en ayuno, la Gluconeogénesis también es estimulada en el ejercicio
prolongado por una dieta rica en proteínas y bajo condiciones de estrés. En condiciones de
ayuno la energía requerida para la gluconeogénesis es obtenida de la beta oxidación.
Los factores que promueven el flujo de carbonos desde el piruvato a la glucosa incluyen la
disponibilidad de sustrato y cambios en la actividad o cantidad de ciertas enzimas claves
de la glucólisis y la gluconeogénesis.
La gluconeogénesis es estimulada por el flujo de los sustratos desde los tejidos periféricos al
hígado. El glicerol es liberado desde el tejido adiposo cuando los niveles de insulina son bajos,
o los niveles de glucagón u otras hormonas gluconeogénicas aumentan en sangre. El lactato
es producido por el músculo durante el ejercicio y por los eritrocitos. Por otro lado, los
aminoácidos son liberados desde el músculo si el nivel de insulina es bajo o cuando el cortisol
es elevado. También se usan aminoácidos para glucogénesis cuando la ingesta de proteínas
es alta y la de carbohidratos es baja.

Bioquímica | Ruta de las pentosas fosfato, gluconeogénesis y síntesis de ácidos grasos
Las cantidades y actividades de las
enzimas características de cada ruta están
controladas de tal manera ambas rutas
que no pueden estar activas
simultáneamente.
Las tres vías de rodeo de la Gluconeogénesis
están reguladas por la piruvato carboxilasa, la
fructosa-1,6-bifosfatasa y la glucosa-6-
fosfatasa.
Regulación de la reacción fosfoenolpiruvato/piruvato
El piruvato, elemento clave en la gluconeogénesis, deriva del lactato o también de AAs
glucogénicos, particularmente la alanina. En condiciones a favor de la gluconeogénesis, el
piruvato no será convertido a Acetil-CoA, ya que, la piruvato deshidrogenasa estará inhibida.
Por lo tanto, el piruvato mediante la acción de la piruvato carboxilasa se convertirá en
oxalacetato, el cual después, se convertirá en PEP, luego a fructosa-1,6-bifosfato para
finalmente terminar en Glucosa.
La piruvato carboxilasa se encuentra activada cuando aumenta
el Acetil-CoA, a diferencia de la piruvato deshidrogenasa que es
inhibida.
La carga energética alta o niveles de precursores de glucosa
altos disminuyen el glucólisis y aumentan la gluconeogénesis.
Regulación de la reacción Fructosa-1,6-bifosfatasa
A diferencia de lo que ocurre con la PFK-1
en que la fructosa-2,6-bifosfato es un
activador de la enzima, en el caso de la
fructosa-1,6-bifosfatasa es un inhibidor al
igual que el AMP.
La fructosa-2,6-bifosfato es el principal
regulador de la PFK-1 y de la fructosa-1,6-
bifosfatasa.
La PFK-2 y la FBpasa-2 forman parte de una enzima bifuncional y están controladas por
fosforilación/desfosforilación. Cuando disminuye la fructosa 2,6-bifosfato se estimula la
gluconeogénesis y cuando aumenta se estimula la glucólisis.

Bioquímica | Ruta de las pentosas fosfato, gluconeogénesis y síntesis de ácidos grasos
Ciclos Fútiles o de Sustrato
Ciclo fútil: ocurre cuando dos vías metabólicas transcurren simultáneamente en
direcciones opuestas, el efecto final es disipar energía en forma de calor.
La ingestión de alcohol y la gluconeogénesis
Mitocondria: piruvato carboxilasa, fosfoenol-piruvato carboxiquinasa, aspartato
aminotransferasa y malato deshidrogenasa
Citosol: fosfoenol-piruvato carboxiquinasa, enolasa, fosfoglicerato mutasa,
fosfoglicerato quinasa, gliceraldehído 3-fosfato-deshidrogenasa, triosa fosfato
isomerasa, aldolasa, fructosa 1,6-bifosfatasa y glucosa isomerasa.
Retículo Endoplasmático: glucosa 6-fosfatasa

05- Gluconeogénesis.pdf
 Estamos procesando este archivo...
Estamos procesando este archivo...
 Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.
Descargar
Estamos procesando este archivo...
Lamentablemente la previsualización de este archivo no está disponible. De todas maneras puedes descargarlo y ver si te es útil.